
Abstract
Purpose
The impact of land use on biodiversity is a topic that has received considerable attention in life cycle assessment (LCA). The methodology to assess biodiversity in LCA has been improved in the past decades. This paper contributes to this progress by building on the concept of conditions for maintained biodiversity. It describes the theory for the development of mathematical functions representing the impact of land uses and management practices on biodiversity.
Methods
The method proposed here describes the impact of land use on biodiversity as a decrease in biodiversity potential, capturing the impact of management practices. The method can be applied with weighting between regions, such as ecoregions. The biodiversity potential is calculated through functions that describe not only parameters which are relevant to biodiversity, for example, deadwood in a forest, but also the relationships between those parameters. For example, maximum biodiversity would hypothetically occur when the nutrient balance is ideal and no pesticide is applied. As these relationships may not be readily quantified, we propose the use of fuzzy thinking for biodiversity assessment, using AND/OR operators. The method allows the inclusion of context parameters that represent neither the management nor the land use practice being investigated, but are nevertheless relevant to biodiversity. The parameters and relationships can be defined by either literature or expert interviews. We give recommendations on how to create the biodiversity potential functions by providing the reader with a set of questions that can help build the functions and find the relationship between parameters.
Results and discussion
We present a simplified case study of paper production in the Scandinavian and Russian Taiga to demonstrate the applicability of the method. We apply the method to two scenarios, one representing an intensive forestry practice, and another representing lower intensity forestry management. The results communicate the differences between the two scenarios quantitatively, but more importantly, are able to provide guidance on improved management. We discuss the advantages of this condition-based approach compared to pre-defined intensity classes. The potential drawbacks of defining potential functions from industry-derived studies are pointed out. This method also provides a less strict approach to a reference situation, consequently allowing the adequate assessment of cases in which the most beneficial biodiversity state is achieved through management practices.
Conclusions
The originality of using fuzzy thinking is that it enables land use management practices to be accounted for in LCA without requiring sub-categories for different intensities to be explicitly established, thus moving beyond the classification of land use practices. The proposed method is another LCIA step toward closing the gap between land use management practices and biodiversity conservation goals.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
1 Introduction
In recent years, the increasing loss of species and habitats has not only become one of the central issues of environmental policy but also a focus topic for consumers and, thus, for the sustainability strategies of companies. The loss of biodiversity and of ecosystems still continues on a large scale despite the increased awareness of ecosystems and biodiversity (de Groot et al. 2010), and despite it being well established that the transformation and reduction of the suitability of habitats are the lead causes of biodiversity loss (IPBES 2018). The impairment of biodiversity is not least caused by land use processes in global supply chains.
Life cycle assessment (LCA) is a standardized method to assess potential environmental impacts of the production, use, and end of life of products and services. As one approach, LCA is also used to map the potential impacts of land-using processes. Land use and its impacts have been a topic in the LCA research community for about two decades (i Canals et al. 2007; Lindeijer et al. 2002; Koellner et al. 2013a). Impacts of land use on soil quality and soil function have at least partially been covered (i Canals et al. 2007; Beck et al. 2010) and the methods are being improved (Bos et al. 2016). With the aim of addressing these issues, the joint Life Cycle Initiative by the United Nations Environment Programme and the Society of Environmental Toxicology and Chemistry (UNEP-SETAC) framework for land use in LCA (i Canals et al. 2007; Koellner et al. 2013a) presents a consensual basis upon which most of the life cycle impact assessment (LCIA) methods have been and continue to be developed.
Recent reviews of the different models to assess land use and biodiversity in LCA show a growing number of papers and approaches (Michelsen and Lindner 2015; Souza et al. 2015; Curran et al. 2016; Gabel et al. 2016; Pavan and Ometto 2016; Winter et al. 2017). In reviewing the indicators for biodiversity with respect to genomes, species, and ecosystems, Winter et al. (2017) noted that 40% of the methods assess biodiversity at the species level. The use of the number of species, also referred to as species richness, as a metric for biodiversity derives from well-developed methods from biological science. Broadly speaking, the biodiversity of a plot of land is understood as the species richness occurring on that plot. Methods that use this metric based on the species-area relationship from ecology are, for example, de Baan et al. (2013), Chaudhary et al. (2015), and subsequent methods. Many methods for the assessment of biodiversity in LCA are derived directly or indirectly from Koellner and Scholz (2008); see Gabel et al. (2016) for a historical overview and connection between methods. Earlier publications addressing biodiversity in LCA referred only to vascular plants (Koellner 2000), and biomes were used for geographical differentiation (Weidema and Lindeijer 2001). With advances in data availability, the methodology has evolved to provide a broader coverage of taxa and regional differentiation, and among more recent methodologies, Chaudhary and Brooks (2018) used data on mammals, birds, amphibians, reptiles, and plants providing characterization factors for 804 different ecoregions. An ecoregion is a large unit of land or water containing a geographically distinct assemblage of species, natural communities, and environmental conditions (Olson et al. 2001).
It is also possible to assess biodiversity in terms of the conditions under which a certain biodiversity level can be reasonably expected. The biodiversity of a plot of land is understood as an abstract index number calculated from the input variables referring to conditions at the site. Several authors have used this approach to assess biodiversity in LCA (Michelsen 2008; Lindner 2016; Kyläkorpi et al. 2005; Lindqvist et al. 2016). Other condition-based methods for the assessment of biodiversity in LCA use hemeroby as a proxy (Brentrup et al. 2002; Fehrenbach et al. 2015; Coelho and Michelsen 2014; Rossi et al. 2018). The hemeroby concept was first introduced in the 1950s in vegetation science and landscape planning (Kowarik 1990) and was brought to LCA by Brentrup et al. (2002). The term hemeroby is used in landscape ecology as a representation of the distance to nature or naturalness (Fehrenbach et al. 2015), and in these methods, naturalness is implicitly assumed as the ideal condition for biodiversity.
A particularly difficult issue within the land use topic in LCA is the definition of the reference state against which the quality difference is calculated in the UNEP-SETAC framework (Koellner et al. 2013b). The reference state for biodiversity is even more difficult to define because a plethora of underlying values are addressed in biodiversity assessment, which is often not made explicit (Michelsen and Lindner 2015). The natural state of ecosystems seems to be the intuitive reference state of biodiversity. However, there are ecosystems with high conservation values that are not equal to their pristine state. Prime examples for anthropogenic ecosystems are the landscapes formed by low-intensity traditional land use in Europe, like heaths and meadows. Without continued human intervention, most open spaces other than bogs would be closed forests (Landesamt für Umweltschutz Sachsen-Anhalt – Halle 2004). The reference situation for biodiversity in the context of LCA and conservation biology was extensively discussed by Vrasdonk et al. (2019).
Biodiversity is a complex, multi-faceted subject. It is common to use a single relevant aspect as a biodiversity metric (e.g., species richness, percentage of protected habitats, richness of red list species), or to combine several aspects into an index, e.g., species richness and rarity (Geyer et al. 2010). While it is common to measure biodiversity with single indicators such as species richness or the extent of the habitat of different taxa, neither of these gives a complete picture of biodiversity. Additionally, empirical methods are vulnerable to data gaps. Based on the approach presented by Michelsen (2008), Lindner et al. (2014) and Lindner (2016) proposed the use of elements from fuzzy thinking to derive values for favourable biodiversity conditions, instead of the use of a pre-defined land use type and intensity. Differently from Michelsen, Rossi et al. (2018) proposed the use of a continuous classification using the concept of hemeroby, partial biodiversity scores and aggregation into a biodiversity potential, with the aggregation being based on the proposal of Lindner et al. (2014).
In this paper, we propose a method based on Michelsen (2008), who used an indicator for the intrinsic value of the biodiversity of an area, which includes aspects of ecosystem scarcity and ecosystem vulnerability, and a specific component that describes the conditions for maintained biodiversity (CMB). The focus of the method described in this paper is equivalent to the CMB component of the method developed by Michelsen (2008). A first proposal for the use of CMB was applied to the case of forests in Norway in which three factors were identified: the amount of decaying wood, areas set aside, and the introduction of alien species (Michelsen 2008). For each of the factors, the author proposed a classification from 0 to 3, representing zero impact to major impact (with a total of four classes). In the CMB case study application, the factors were not weighted, and other relevant factors that were judged to be relevant but not included were mentioned, such as tree species composition, regeneration, and number of large trees (Michelsen 2008).
Some authors have used the conditions approach to assess the impacts of forestry, mining, and agriculture (Eberle and Lindner 2015; Lindqvist et al. 2016; Eberle 2018; Föst 2019; Lindner et al. 2019; Geß 2020). Despite applications of the method already being published, the detailed description of the methodology to address the conditions for maintaining biodiversity using fuzzy thinking and its relationships is limited to Lindner (2016) in German. This paper fills this gap, providing details of the framework for the use of fuzzy thinking and operation for the assessment of biodiversity in LCA, and an example of the method’s use for forestry will be described as a case study for paper production.
2 Method
The aim of this paper is the description of the estimated biodiversity value of the area occupied by a process which uses or occupies land, under the conditions set by the process. In the following subsections, we describe the proposed method in order of decreasing abstraction. Section 2.1 describes the mathematical building blocks of the method; Sect. 2.2 provides guidelines on how to develop the biodiversity potential function, and Sect. 2.3 describes the integration in the LCA framework for land use assessment.
2.1 Method architecture
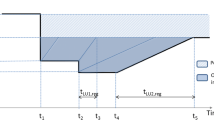
The method follows the principle that any land use activity influences the environmental quality, or quality (Q) (Lindeijer et al. 2002). This principle is mostly agreed upon and forms the basis of the framework for the assessment of land use in LCA proposed by the Life Cycle Initiative (Koellner et al. 2013b). The framework describes any land use process in three dimensions: quality Q, time t, and area A. The impact of a land use unit process on the quality of the land can in many cases be simplified as the product of the quality difference ΔQ (between the quality during use and the reference quality Qref), the duration of the land use Δt, and the area affected A (Fig. 1). The framework goes beyond this simple definition, but for the article at hand, the most relevant aspect is that any method that addresses biodiversity as land quality needs to define the Q axis in a way that allows the framework to be meaningfully applied.

Schematic representation of the impact of a land use on the land-quality (Q), as a function of the duration of occupation (t) and area. Figure based on Koellner et al. (2013a)
In our method, we define biodiversity as land quality Q in the Q,A,t diagram. In order to assess the impacts of land use, we start from the principle that certain conditions are beneficial or damaging to biodiversity. These conditions are our parameters, here \({x}_{i}\). A biodiversity parameter has a contribution to biodiversity yi.
If the contributions are assessed in a categorical manner such as high intensity and low intensity, they can be understood as characteristics of crisp sets. This classification is, of course, a coarse simplification, as these contributions are not necessarily binary, meaning that either the contributions interact or not. Most of our tools for modelling, reasoning and computing are dichotomous, “yes” or “no” scale, meaning a statement can be true or false, and nothing in between, i.e., crisp (Zimmermann 2010). In crisp sets, or classical set theory, an element membership function is binary, 0 or 1 (Guo and Wong 2013). To deal with the problems that arise in the absence of sharply defined criteria of membership, Zadeh (1965) proposed the use of fuzzy sets to address a continuum of grades of membership. A graphical representation of crisp and fuzzy sets is presented in Fig. 2.

Illustration of crisp and fuzzy sets and membership functions
Fuzzy modelling is suitable for the numeric representation of both quantitative and qualitative knowledge (Zadeh 1973). A simple example of crisp and fuzzy set theory is the case of a tall person: using crisp set theory, the person is defined as being “tall” or “not tall,” i.e., there is a cutoff at a certain height (e.g., 190 cm). With fuzzy sets, the person’s height is classified according to the degree of membership in the “tall” category, with increasing degrees of membership defined as a function of height (Ong and Tilahun 2011).
A wide range of scientific fields used fuzzy modelling, with 26 research journals on theory or application of fuzzy logic (Singh et al. 2013). The use of fuzzy logic has been proposed in the field of biodiversity conservation: for example, to assess the sustainability of a protected area ecosystem (Prato 2007), to evaluate the ecological vulnerability of habitats (Caniani et al. 2016), and to avoid false absences of spatial distribution of species (Barbosa 2015).
The method detailed here is designed to make use of knowledge beyond hard numbers, such as the number of species as input, and to avoid the strict definition of land use intensity classes. The method was developed to allow a continuum of values for each parameter x, to represent its influence or contribution y to biodiversity. The method converts knowledge on the influence of certain parameters into fuzzy sets to communicate “high biodiversity.”
2.1.1 Defining biodiversity potential functions and biodiversity contributions (y)
Each singular parameter (x) provides a certain contribution to biodiversity (y). The generic expression of \({y}_{i}({x}_{i})\) is:
While the expression may seem convoluted, it actually describes the Gaussian function, or a simple bell curve with constants α = 2, σ = 0.15, β = 0.5, γ = 0, δ = 1, and ε = 1 in Fig. 3.

Gaussian distribution, or basic function
The six constants α, σ, β, γ, δ, and ε serve to mold the curve into any desired shape depending on the type of contribution (y) that the parameter (x) has on biodiversity. Specifically, the exponent α sets the width of the plateau without affecting the width of the entire bell, σ sets the width of the bell, but not the plateau, β and γ shift the entire curve either in the x- or in the y-directions, and δ and ε shift the peak of the bell either in the x- or y-direction. An example of the effects of varying the constants is presented in Fig. 4.

Examples of Gaussian curve with altered constants α, σ, β, γ, δ, and ε
The impact of land use on biodiversity does not only depend on several factors, here the parameters and their contributions, but also on the interactions between them. The interaction of parameters for crisp sets is defined by a hard border, while for fuzzy sets, the interaction is continuous (Fig. 5).

Illustration of crisp and fuzzy sets intersection
If the individual parameters (x) and biodiversity contributions (y) are related, they can be combined by applying the logical operators AND/OR. The operator AND is used to represent parameters such that both need to be in a good range in order to achieve a high potential biodiversity value. If one of the two parameters is outside of the favorable range, the potential biodiversity is low, regardless of how good the value of that second parameter is, as the second other parameter cannot compensate for the other parameter. The operator OR represents parameters that compensate for each other. In this case, if one parameter is in a beneficial range, the value of the other parameter is less important. A graphical representation is shown in Fig. 6

Strict AND (left) and strict OR (right) relations between parameters
The following paragraphs describe the functions y(x) for different types of interactions of parameters A and B using the nomenclature \({y}_{AB}\left({x}_{A},{x}_{B}\right).\) The simplest definition of a fuzzy intersect between two sets A and B is the multiplication of the membership functions, representing the AND operator:
An example of an AND relation can be found in agricultural processes: to achieve maximum biodiversity, the nutrient balance should be in equilibrium AND pesticide input should be zero. In this case, if only one parameter is in the favorable range, this would not be enough for high biodiversity.
The definition of a fuzzy sum of two sets A and B, which represents the OR operator is described as:
To illustrate the concept of OR relationships, we can think of the example of forestry. Knowing and protecting the most relevant habitats in a managed forest is roughly as good as broadly protecting a large fraction of a managed forest. In this case, one can assume that protecting large tracts of land would also protect the most relevant habitats. If one parameter OR the other is in the beneficial range, biodiversity is potentially high.
In the two cases above, the two parameters are strictly AND/OR-related. However, less strict relations are also possible. Inspired by multi-criteria decision theory (see, e.g., Zeleny (2011)), the following operations can be used. They represent a softer relation between the parameters as the fuzzy intersect (soft AND) of their corresponding fuzzy sets:
The corresponding fuzzy sum (soft OR) is then defined as:
Both operations use the exponent p to fine-tune the strictness of the relationship. A value of p = 1 causes the surface defined by xA and xB to take the shape of a flat plane. Values of p above 1 push the corners at (0, 1) and (1, 0) lower (AND) or higher (OR). With p → ∞, the functions become very similar to the strict AND/OR functions described in Fig. 6. Soft relationships of parameters with intermediate values are exemplified in Fig. 7 (soft AND) and Fig. 8 (soft OR).

Soft AND relation between parameters with p = 0.5 (left) and p = 2 (right)

Soft OR relation between parameters with p = 0.5 (left) and p = 2 (right)
It is also possible that one or more parameters \({x}_{B}\) do not entail any biodiversity contribution of their own, but affect the biodiversity contribution function of another parameter \({y}_{A}\left({x}_{A}\right)\). In such a case, we call the main parameter a management parameter \(\left({x}_{A}\right)\), and the modifying parameter, a context parameter \(\left({x}_{B}\right)\). This relation can be expected in situations in which the efficacy of a certain land management measure depends on the landscape context: for example, the distance to a natural area. For the mathematical representation of the biodiversity contribution \({y}_{A}\) as a function of management parameter \({x}_{A}\) and context parameter \({x}_{B}\), we suggest defining \({y}_{A}\left({x}_{A}\right)\) at both extreme ends of the \({x}_{B}\), its range, and then using a function \(h({x}_{B})\) as a sliding weight between the sum of the two \({y}_{A}\left({x}_{A}\right)\) functions. In the absence of further information, the sliding function \(h({x}_{B})\) can be, for example, a simple linear function. The equation for the context parameter \({y}_{A}\left({x}_{A},{x}_{B}\right)\) is:
A visualization of the contribution of the context parameter is shown in Fig. 9. An example would be if \({x}_{A}\) is the size of the natural area (on one’s own land), and \({x}_{B}\) is the distance to the next natural area (outside of one’s own land). The context parameter is able to capture that the perceived quality of the natural land cover in the area of interest is reduced if it is isolated, but its contribution to biodiversity increases if it is part of a network of natural areas.

Context parameter relation
A summary of the equations used in the method architecture and their corresponding operators is presented in Table 1. These contributions also interact. Therefore, the biodiversity contribution functions are aggregated into the biodiversity potential field function. Applying weighting factors zg, which assign different weights to the various biodiversity contributions, yields the biodiversity potential (BP), a potential field function. BP is defined by the biodiversity contribution yij from management parameters xi and context parameters xj, as well as a number k of weighting factors zg for each (yij)g
The potential field function is normalized to the [0, 1] interval, meaning that the sum of all zg needs to be 1.
In summary, the model proposes assessing biodiversity by subdividing it into individual parameters, defining their relationships, and calculating their respective biodiversity contributions. At each step, the intermediate result is normalized to the [0, 1] interval to facilitate the modularity of the system. The biodiversity field function is thus created from standardized blocks bonded by standardized links.
2.2 Development of the field equation
This section addresses LCIA development describing the process of using the regional LCIA models for deriving case-specific biodiversity characterization factors for land using processes. Literature reviews, interviews, and workshops can be carried out to obtain the parameters necessary for the assessment of biodiversity. A key input to determine the parameters, the functions, and the relationships of the contributions is expert knowledge. On the one hand, expert interviews or workshops can be time-consuming; on the other hand, biodiversity expert involvement can unlock a wider range of knowledge when compared to literature.
Fuzzy sets are often used to reflect the inherent subjectivity and imprecision in evaluation processes (Tavana and Sodenkamp 2010), with the linguistic variables of fuzzy logic lending themselves to representing human preferences for example in multi-criteria decision analysis (Yatsalo et al. 2017; Dilli et al. 2018; Ajibade et al. 2019; Das and Pal 2020). As fuzzy logic and fuzzy thinking are suitable for the representation of knowledge, or even intuition, the proposed method is designed to make use of knowledge which can be valuable to improve the assessment of biodiversity in LCA. Also, “biodiversity” as an aggregate indicator contains strong normative components (note that the interview questions in the following subsection deliberately include normative vocabulary like “better”).
2.2.1 Conducting interviews
When carrying out interviews, our suggestion is to use the Convention on Biological Diversity definition as a guardrail for the discussion. In our experience, at any given workshop (or one to one interview) with biodiversity experts, trying to define the subject is likely to lead to extensive discussions. Instead of focusing on the definition of biodiversity, we suggest asking experts to describe “ideal conditions for biodiversity.” This, in our experience, yields converging results. Counter-intuitively, it is actually quite practical to work towards a common understanding of the conditions for biodiversity without explicitly defining it in too much detail. This approach seems to force contributing experts to think of “biodiversity” as a whole, so the normative aspect of aggregating the various aspects of biodiversity into the value of land is included.
In this section, we make suggestions on conducting interviews to develop the biodiversity field functions. We describe the aspects and questions which are relevant while conducting expert interviews. We do not intend this list to be exhaustive, but rather to be seen as general guidance. The answers to the questions refer to the parameters, their respective biodiversity contributions, AND/OR relations between contributions, and the weighting of contributions necessary for the linear aggregation.
As a starting point, researchers seeking to define a field function for the biodiversity potential should conduct an extensive literature review to develop their own preliminary understanding of biodiversity within a given area of interest: for example, an ecoregion. Hereafter, we will refer to ecoregions to communicate differences in the distinct characteristics of a study area for the simplicity of it, and because this has been widely applied in LCA.
The initial literature research may guide the selection of potential interview partners. These could be, for example, the authors of national strategy documents, non-governmental organizations’ reports, or scientific papers on the most relevant threats to biodiversity. A good starting point for scientific publications are the reports of the Intergovernmental Platform on Biodiversity and Ecosystem Services (IPBES), in which scientific knowledge about biodiversity and ecosystem services is aggregated in a curated manner. When talking to the experts, we found that it was better to avoid giving too much background knowledge about LCA and the Life Cycle Initiative framework, and instead focus on describing the state of biodiversity.
In order to determine the potential field functions, the method developer can focus on questions such as the following: How do the rather generic drivers of biodiversity loss described, e.g., in the Millennium Ecosystem Assessment (Millennium Ecosystem Assessment 2005) and the Living Planet Report (WWF 2016) manifest themselves in the specific ecoregion? How relevant are they in this ecoregion? Which national conservation goals are relevant for this ecoregion? The following questions can help to structure the interview:
-
What is the typical biodiversity of this ecoregion?
-
Which anthropogenic activities threaten it?
-
Which conditions on an area plot (or attributes of the plot) indicate places of high or low biodiversity?
-
How can the state of biodiversity be qualitatively linked to anthropogenic activities and/or the conditions or attributes of an area?
-
How can the state of biodiversity be quantitatively linked to anthropogenic activities and/or the conditions or attributes of an area?
-
Are anthropogenic influences and the land conditions or attributes independent from each other? If not, how do they interact?
-
Weighting: what is the appropriate share of the individual influences on the state of biodiversity in the overall state of biodiversity?
Once the questions have been framed, the contribution (y) of the identified parameters (x) has to be established. In the absence of a predefined relationship curve, the function can be drawn from expert knowledge. In the following section, we provide guidance on how to draw functions for the identified parameters regarding their biodiversity contribution, allowing the researcher to draw the functions:
-
1.
What is the relationship of the parameter to biodiversity? Is more generally better? Or the less, the better? This question allows the researcher to define the start and end of the function, i.e., the top or bottom corners.
-
2.
Is having more or having less of the parameter always better? Or does it reach a plateau at some point? Or does it peak and then drop again? This question allows the researcher to define the overall shape of the curve.
-
3.
Does it take a certain level of presence or absence of a parameter to influence the biodiversity? This question allows the researcher to further refine the curve.
It also helps to interpret the relationship by drawing the general shape of the biodiversity contribution functions by hand. In our experience, we have found that there are a few basic recurring shapes. We call them type 1a, type 1b, type 2a, type 2b, type 3, type 4a, and type 4b, shown in Fig. 10. An example of the values for the constants α, σ, β, γ, δ, and ε to obtain the different types of curves is presented in Table 2, and the relationship of type 4 is linear and simply described as the equation of a line of positive or negative slope.

The four main types of curves representing the relationship of a parameter to its biodiversity contribution
Type 1a represents an immediate sharp drop in biodiversity. This is usually observed in pesticide input. They are actively ecotoxic (by design), and small amounts can have a high negative impact on living organisms. The marginal biodiversity decline decreases with rising parameter values (because there is little biodiversity left to be affected) and the curve levels off at the low end of the [0, 1] interval. Type 1b represents the reverse function and can represent, for example, microhabitats where additional microhabitats add value to biodiversity, but their contribution to biodiversity decreases when they are no longer special and do not add value to biodiversity, no matter how many of these new microhabitats are created.
Type 2a reflects cases where the parameter has little effect at first, but the biodiversity contribution drops after a threshold. This is typical for fertilizer input, as many ecosystems can tolerate some nutrient imbalance, but the resilience ceases after a certain amount. This curve also levels off toward the lower end. Type 2b describes the reverse function; an example is deadwood in a forest, where no or very little deadwood adds no contribution to biodiversity; an increased amount of deadwood contributes to biodiversity, and then levels off, which means that adding more deadwood would not increase its contribution to biodiversity.
Type 3 describes a rise in biodiversity, which then drops off after a local maximum. This is the case for some measurements of farming intensity in regions where the desired biodiversity has been formed by traditional land management, e.g., mowing of grassland in western and southern Europe. In such places, low-intensity management means regular, selective rejuvenation, which secures habitats for desired flora and fauna. If the management is too intensive, biodiversity is much lower than without management at all, so this curve also levels off toward the lower end.
Type 4 is a simple line from one corner to the opposite corner, which is the top left to bottom right (4a) or the bottom left to top right (4b) corner. Such a function can be used for parameters that yield a biodiversity level that is proportional to the parameter value, or this function might serve as a proxy or interim function for parameters for which only “more is worse” or “more is better” is known, e.g., simply knowing that the removal of biomass is worse for biodiversity.
Once a biodiversity potential field is defined as a function of relevant input parameters, its application is relatively simple. We estimate that around five to ten parameters would be defined depending on the land use investigated. The field function yields one data point as output: the biodiversity level of the affected land plot under the conditions defined by the input parameters.
2.3 Integration to the LCA land use framework
The impact of land use activities on biodiversity may differ depending on their location. Such a difference can be captured by a regional or ecoregion factor. For the method at hand, we propose that the value of the region or ecoregion is assessed as part of the Q, but independent of the contribution of land use to biodiversity.
Differences in biodiversity which are specific to the location are accounted for using, for example, an ecoregion-specific factor \({w}_{ER}\); the development of this factor is not part of this paper. Most importantly, the method proposed is designed to accommodate any weighting. Nevertheless, one example is the ecoregion weighting factor calculated from the areas of ecoregions and their species richness compiled in the WildFinder database (World Wildlife Fund 2006), used by Lindner (2016).
The reference state quality level (Qref) is set as the maximum biodiversity at a location within the given ecoregion. BP is normalized to the interval [0, 1] within each ecoregion, so BPref is 1 by definition, and a biodiversity gap can be calculated. This gap is the potential that is not realized when the conditions are not ideal. It describes the difference in quality needed to calculate the impact of each land using process. Accordingly, the quality difference is:
The BP of a given patch of land is described as a function of parameter values \({x}_{ER,i}\) that are relevant for that specific ecoregion \({w}_{ER}\). The methodology laid out here is to be understood as a toolbox for developers of biodiversity functions, i.e., \(BP\left({y}_{ER,i}\right)\) has to be defined for each ecoregion. This methodology ties them all together through a common architecture of the method.
3 Application example for forestry
In order to provide a tangible understanding of the use of the method, the method will be applied to forestry. Forestry products have been widely investigated in LCA with 22 peer-reviewed papers, four original reports, and two databases analyzed by Klein et al. (2015). Forestry or forest products have been used to test the application of the different methods for assessing the impact of land use on biodiversity (Michelsen 2008; Michelsen et al. 2014; Lindner 2016; Rossi et al. 2018; Côté et al. 2019; Myllyviita et al. 2019; Turner et al. 2019). Here, we demonstrate the use of the method proposed with a practical example of the simplified case of paper production in Finland.
In order to develop the potential field functions, the first question that has to be answered is the location where the land use takes place. Here, we will use ecoregional borders from Olson et al. (2001). With the exception of smaller areas in the north and south of Finland, most of the territory coincides with the ecoregion bordering the Scandinavian and Russian Taiga (PA 0608). Finland’s pine and spruce vegetation cover maps are available through a governmental online map (Makisara et al. 2019). The overall location of the forests was considered sufficient to consider that it overlaps the Scandinavian and Russian Taiga ecoregion without the need of geospatial analysis tools.
The second step is the determination of the parameters that contribute to biodiversity in this ecoregion. Here, we will report the results based on expert consultation. Four experts were interviewed in the year 2014: Prof. Rainer Luick who is active in nature conservation and environmental protection at the Rottenburg University of Applied Forest Sciences, Timo Lehesvirta who has developed the biodiversity program of the Finnish forestry company UPM Kymmene, Petri Ahlroth, head of the Natural Environment Centre at the state-owned Finnish Environment Institute (Suomen ympäristökeskus, SYKE) and Lauri Saaristo, head of the Nature, Environment and Waters department at the Finnish forestry consultancy Tapio. It was communicated to the experts that in order to create a unified biodiversity indicator, substantial simplifications were necessary. The experts did not construct any of the functions by themselves. The numerical definitions were ultimately set by the lead author using expert knowledge as input.
Compiling the information from the interviews, eleven main parameters were identified; they can be generally described as the diversity of native vegetation, the age structure of standing timber, the amount and diversity of deadwood, the number and diversity of microbiotopes, the size of riparian strips, the size of protected areas, and the frequency of disturbances. Other parameters, such as the interconnectedness of protected areas, were rejected either due to the difficulty in assessing them or in order to limit the number of parameters.
3.1 Description of identified parameters
Native species is a common indicator of biodiversity, and in this case study, the assessment is limited to trees. Exotic vegetation is an indicator of lower quality of biodiversity, as it limits the area and may compete with native species. The number of native tree species excluding the most common, namely pine, spruce, and birch, was recorded as number/ha, while exotic vegetation was measured in % area coverage. The number of native species ranges from 0 to 25/ha, and exotic vegetation ranges from 0 to 100%.
The age structure of the tree population is considered by the experts to be an important indicator for the quality of biodiversity, not only because the existence of native tree species is important but also because a varying age structure is desirable, as it allows different communities to thrive. The tree age structure was grouped into young trees under 20 years of age and old trees over 150 years of age. With help from the experts, the scale of the age structure classes was defined as 0 to 100 trees per hectare for young trees and 0 to 20 trees per hectare for old growth. No parameter for middle-aged trees was defined, because these can safely be assumed to exist in most places.
Deadwood is a very important parameter in many forest-dominated ecoregions. It is a fundamental part of the food chain of a forest ecosystem; as it decays, it provides food for decomposers and other organisms. Although the presence or absence of deadwood is a suitable indication of the biodiversity quality, the age of the deadwood is also of significant importance. Other factors of importance are the stage of decomposition, its diameter, and whether it is standing or laying. Here we will use three simplified decomposition classes: class I being fresh deadwood; class II is deadwood whose trunk is covered in moss and where the bark and branches are falling off; and class III is the stage where the trunk is falling apart. These classes are easily identified by a forest manager, but can also be distinguished with reasonable accuracy by a non-expert who has been adequately briefed in the characteristics. These parameters are recorded in m3/ha. With help from the experts, the scale of the deadwood classes was defined as 0 to 10; 0 to 30; and 0 to 20 m3/ha for classes I, II, and III, respectively.
Biotopes are areas associated with a particular set of the ecological community, and microbiotopes are small areas that are distinct from the overall biotope. Microbiotopes support biodiversity and are an indicator of biodiversity in itself, and the area is considered a valuable habitat. In the case of forests in Finland, one example of a biotope is a water hole in the forest. Riparian zones, or the area in the interface between water and land, their size and conservation status are important for maintaining biodiversity. Here, microbiotopes and shore zones are considered as valuable habitat parameters. For this parameter, the number, size, and area are important. Thus, valuable habitats will be measured both in numbers [1/ha] and in area [%].
Protected areas are designated to protect biodiversity and play a fundamental role in the conservation of biodiversity. They can be designated by law or voluntarily set aside. Definitions of protected areas vary among countries, and here no legal definition is taken into account. Instead, the parameter is the simple % of the unused area. The importance of the protected areas is not only limited to their total size; the shape of the protected areas, their configuration, and their purpose are also important, but these aspects have not been quantified. Neither does the likeliness of persistence of the protected status into the future make any difference for the valuation, because any calculation of the quality Q refers only to Q(t) at a given point in time.
Another parameter relevant to biodiversity is the frequency of disturbances, such as low-intensity fires. Fire regimes are important because some species are dependent on the post-fire succession process (a prominent example is the black fire beetle Melanophila acuminata). Here, fire regime is recorded as number of fires on the study area per year in 10-4/a, simplified from 10-4km2/km2a.
3.2 Contribution curve
Guided by the interview questions, the experts described or drew biodiversity contribution functions. The shape of the resulting curves corresponded to one of the curve types. The constants of the basic function were altered to result in curves that were similar in shape to those drawn or described by the experts or to meet specific fixed points described by the experts. The resulting curves were presented to the experts and adjustments were made where necessary. The final curves, representing the contribution of individual parameters to biodiversity, are presented in Fig. 11. A compilation of the parameters, their scales, and the constants used to construct the contribution curves are presented in Table 3.

Biodiversity contribution of parameters relevant for forest ecoregion grouped by a tree age structure, b tree species diversity, c deadwood, d protected areas, and e disturbances
3.3 Parameters relationships
Once the parameters were identified and their contribution curves established, their relationships had to be determined. For parameters that are related, an AND/OR, strict/soft were defined based on the information from the discussion with the experts.
The age structure is captured using algebraic sums, expressing that we need both young and aged trees, using a strict AND. This way, it is assumed that middle-aged trees are always present.
To achieve high biodiversity, both the number of tree species beyond the three most common species spruce, pine, and birch, has to be high, and the share of exotic species has to be low. The area of native vegetation is captured here by the negative contribution of the exotic species parameter. The relationship between these two parameters is represented as strict AND operation.
The deadwood classes are related parameters. The maximum biodiversity values are realized when deadwood of several decay stages is found. If only one deadwood class exists, there is a contribution to biodiversity, as it is considered better than the total absence of deadwood. However, the contribution is limited. This is translated as a soft AND, a compromise intersect, using a p = 2 in Eq. 4.
The amount and number of valuable habitats, as well as protected areas, here recorded as the parameter ‘unused areas’, translate independent concepts of conservation. The protected area and the existence of valuable habitats represent similar but not identical aspects of biodiversity. The existence of both can be considered complementary, and it can be said that they could be sufficient on their own. However, the existence of all parameters is better than only one. Therefore, the relationship is modelled with a soft OR. As the parameters can overlap considerably, a p = 5 was applied, communicating a near redundancy between these parameters.
The fire regime is a single parameter that is not related to the other parameters. Therefore, its contribution will be applied directly without being linked to the contribution of other parameters.
3.4 Weighting of relationships
The five relationships, namely age structure, species diversity, deadwood, protected areas, and fire regime, all amount to the total contribution to biodiversity. An equal weight among these relationships gives a contribution of 20% for each. From the discussion with the experts, deadwood is the most important parameter for biodiversity in forests, and it was indicated that if only one indicator could be investigated, deadwood would be the chosen parameter. The importance of fire regimes is limited to a small area, and the time intervals between fires in the same area can span several decades. Therefore, deadwood weighting was increased to 30% and fire regime weighting was reduced to 10%. These values are applied to equation 7 with the zg scaled from 0 to 1. However, if there is no evidence that some parameters are more important for the biodiversity of the respective study region than others, an equal weighting can be assumed.
3.5 Application to paper production
A graphical summary of all parameters, their relationship operators, and the weighting is shown in Fig. 12. To give an example, the biodiversity potential function of the Scandinavian and Russian Taiga ecoregion is applied to a simplified product system. Here, paper production is defined as 100% fresh pulp derived from Finnish forestry. To produce 1 kg of paper, our simplified system requires 0.75 kg of pulp and 0.25 kg of additives. For the production of 1000 kg of pulp, 5 m3 of wood is processed; therefore, for 1 kg of paper, 0.000375 m3 of wood is required. The forest productivity is around 4.1 m3/ha a. When calculated for 1 m3 of wood, 2.439 m2 a is required, and for 1 kg of paper, the area requirement is 9.15 m2*a.

Summary of parameters, their relationship, and weighting used in the case study
Two scenarios are defined, providing a practical example of how this method can contribute to the assessment of management practices of a forestry product. The scenarios were designed to be realistic, with scenario A being representative of intensive clear-cut forestry with no exotic tree species present, most deadwood removed, a very low share of valuable or protected habitats present, and forest management to avoid fire. Scenario B represents less-intensive forestry practices, with native tree species at different ages, deadwood of different classes, protected and valuable habitats, and occasional fires.
The method was applied to the two scenarios for a simplified paper production. The field equation was developed for one ecoregion. Both scenarios take place in the same ecoregion, thus weighting between the regions (wER) is not necessary. The BP was calculated using the weighting factors developed. The input and results are presented in Table 4 and Fig. 13. Note that in Fig. 13, higher values represent a higher contribution to biodiversity, i.e. a more preferable situation.

Results of two case study scenarios, where A represents intensive management practices and B are less intensive management practices. Higher values represent a higher contribution to biodiversity, i.e., a more preferable situation
The results of the case study show the expected overall lower contribution to biodiversity for the intensive scenario A compared to scenario B. For scenario A, the management regime reaches a BP of about 7 % (meaning 93% unrealized potential) and scenario B reaches a BP of 42 % (68% unrealized), with all individual parameters of scenario A having a lower biodiversity contribution than the parameters in scenario B. Note that for simplicity, identical yields are assumed between the scenarios.
4 Discussion
The main strength of the “conditions” approach is the attempt to describe biodiversity in its entirety. Michelsen (2008) suggests calculating biodiversity as a combination of key factors weighted according to the characteristics of the ecoregion. The method proposed here can be seen as a refined and further-developed version of that approach. Here, the components are reformulated to include more ecoregion information and more local information, respectively. In particular, region-specific key factors are reformulated as the biodiversity potential BP(xi) to provide a more systematic framework for identifying parameters and defining their individual or combined relations with biodiversity.
The originality of the proposed method is that it allows differentiation within the same land use class beyond categorical intensity classes of low and high intensity. This is an improvement to the methods assessing biodiversity in LCA, be it conditions-based methods or species richness-based methods. The method proposed by Coelho and Michelsen (2014) was also built on Michelsen (2008), but was unable to delineate different management regimes because it used hemeroby as a proxy for CMB. The methods based on species richness and the species-area relationship only distinguish between predefined land use classes, such as agriculture, grassland, and forest and coarse intensity subclasses. To further refine the subclasses, it would be necessary to construct a regression curve for the species-area relationship requiring a statistically viable number of species counts. For species richness methods, the more differentiation is desired, the more classes need to be defined, and the species data required would increase exponentially. The use of fuzzy thinking provides a continuous scale that allows for management practices to be reflected in LCA, eliminating the need for the existence of an adequate pre-defined land use intensity class. Methods based on conditions for biodiversity can include expert estimates that are both valuable and plausible but would require extensive and expensive studies to validate in any specific case empirically.
A biodiversity potential field function can be constructed using input data that is typically documented within a company. Typical parameters are, for example, plot size, fertilizer and pesticide amounts, and harvested biomass. If input data are not documented in company records, they can be obtained from other sources where they are readily documented, e.g., GIS layers from governmental bodies or nature conservation NGOs. If more specific primary data is unavoidable, data can be obtained, e.g., from satellite images, which is still easier than a field survey for a species inventory.
Expert knowledge goes somewhat beyond codified knowledge, which is generally advantageous for capturing the breadth of biodiversity. A case can be made for including local and possibly indigenous knowledge about biodiversity (e.g., in complex systems such as agroforestry), which the method can accommodate. However, relying on expert knowledge can also make it harder to pinpoint data gaps. It is possible that in some cases, the expert-based fuzzy method unconsciously hides knowledge gaps, leading to the exclusion of relevant parameters. It may also happen that, in an effort to keep the calculations relatively simple, more complex interactions between parameters are omitted (e.g., interconnectedness of protected areas). The method architecture is flexible enough to appreciate unknown biodiversity. For example, the weighting factor z for a group of parameters could be increased if there is reason to believe that the elements of biodiversity depending on these parameters are underrepresented in both codified research and expert knowledge. We see this as a possibility, but have not developed a methodological procedure for such a case.
When comparing different locations, the biodiversity potential is weighted across ecoregions. The term ecoregion is used in the broad sense to describe bio-geographical differences between regions. The method can be applied for different classifications, such as finer-grained resolutions. The ecoregion-specific development of biodiversity potential field functions captures relevant aspects for biodiversity that are distinct from other areas and allows the comparison of impacts across ecoregions. The modular architecture of the methodology means that additional field functions for other ecoregions can be added subsequently without the need to adjust existing field functions. Ecoregion factors are independent of the field function within the respective ecoregions, and thus can be easily altered.
One advantage of the proposed methodology is the definition of the reference state. The method prescribes how to define the reference state rather than setting a specific definition, meaning that they are not predefined, but are definable. The reference state is implicitly defined with the development of an ecoregion’s biodiversity potential field function; in other words, a distance-to-target approach. Each ecoregion-specific reference state would be set through the development of the biodiversity potential function, so there is not just one reference state but a number of ecoregion-specific reference states.
In any given ecoregion, the reference state for biodiversity is the set of conditions under which the biodiversity potential reaches its maximum. Any biodiversity level lower than that is considered an impact, even if the reference state is only achievable through continuous anthropogenic intervention. In an ecoregion where intermittent disturbance yields the highest biodiversity value, this would be reflected in the fuzzy system regardless of whether the disturbance is natural or anthropogenic. In another ecoregion, stable conditions may yield the highest biodiversity value, so there the reference state would be more static. The expression “maintain” (in “conditions for maintained biodiversity”) refers to the long-term valuation of biodiversity, potentially including cases in which the highest valued biodiversity may not be naturally self-maintaining.
The possibility to elegantly include all land use types that may occur in a given ecoregion in one unified function is a strength of the proposed methodology. It may also be a potential weakness if the function uses 20 or more input parameters, which would then make the function difficult to construct. Also, there is a risk that, if biodiversity potential field functions are generated in specific case studies, the parameter choice could be biased toward those parameters that are particularly relevant to the industrial branch in question as, for example, agriculture, forestry, and open-pit operations. If such a bias is unavoidable, any publication about the study must be transparent and explicit about it.
We are aware that the accuracy of the biodiversity potential field function may be overstated. While the result can be calculated to the nth decimal, it is paramount to keep in mind that each contribution function is a simplification, an abstraction from reality, and so are the AND/OR relationships. Results should, as always in LCA, be treated with caution.
We have provided an illustrative example of the case of forestry in Finland, but stress that the method proposed is applicable to any land use, and has already been tested for other cases (Eberle and Lindner 2015; Eberle 2018; Föst 2019; Lindner et al. 2019), and the method is explicitly designed to be applied to any land use anywhere in the world. In the sections describing the method, illustrative examples were given for different types of land uses and parameters.
The specific example is about land use, but the method is applicable for land use change as well. It provides a Q(t) value depending on a number of inputs. Where land use change occurs, two Q(t) values have to be calculated and compared. Typically, the reference state can then be eliminated from the calculation, so that only the quality before and after the change remain. The same principle is to be applied in consequential studies; the quality that would be achieved without the trigger for the consequences is to be compared to the quality that is achieved as a consequence of the trigger. However, what the method does not cover are transitional practices causing land use change. It is designed for assessing the biodiversity value in reasonably stable situations (including “before” and “after” situations of land use change), but not the specific progression of the Q(t) function in the transition phase. For example, slash-and-burn agriculture transforms forests into arable land or pasture. It is possible to quantify the biodiversity value of the forest and the pasture, but not the slashing-and-burning itself.
5 Conclusion and outlook
The method provides a mathematical framework to systematically describe regionally desired biodiversity. The biodiversity potential field function is created from standardized blocks of parameters bonded by standardized links modelled using AND/OR relationships, yet offers great flexibility to honor the nuances of biodiversity across the world. The novelty is that is moves beyond the categorization of land use intensity classes focusing on the practices which are more beneficial or damaging to biodiversity in the area of study. This presents an opportunity for conservation authorities and conservation NGOs to fill the framework with specific requirements. This method brings the assessment of biodiversity in LCA closer to corporate environmental management. Defining the parameters, contributions, and their interactions to reflect conservation goals of different regions, and companies can measure through LCA how far from these goals their product’s land use is.
We foresee that biodiversity potential functions are most likely to be developed for ecoregions, and despite them being rather large units of land, this can be a tedious process for developers, but rewarding when it allows practitioners to use the methodology with relatively low effort, bringing the benefit of being able to distinguish and characterize management practices. Once the method is applied for different ecoregions and land uses, there might be an opportunity to simplify the construction of some of the field equations and parameters by transfer of knowledge, such as within the same biome or certain parameters for some land use practices. For example, Maier et al. (2019) use a similar, but expanded, method architecture; Lindner et al. (2019) build on both Maier et al. (2019) and approach presented here, but do define land use classes. Also, developing or using the field functions for one ecoregion may not be applicable to all ecosystems: for example, a wetland that is within a forest ecoregion’s boundaries.
Overall, in attempting to model reality, and as with any model, recommendations derived from it have to be carefully interpreted, especially in such early stages of development. Yet, we believe that our methodology is a practical way forward as it allows LCA developers to provide LCA users with tools that yield actionable results regarding impacts on biodiversity. The method has been applied for a variety of cases in different fields, but these potential field functions may need refinement as the overall methodology matures.
References
Ajibade FO, Olajire OO, Ajibade TF, Nwogwu NA, Lasisi KH, Alo AB, Owolabi TA, Adewumi JR (2019) Combining multicriteria decision analysis with GIS for suitably siting landfills in a Nigerian state. Environmental and Sustainability Indicators 3–4:100010. https://doi.org/10.1016/j.indic.2019.100010
Barbosa AM (2015) fuzzySim: applying fuzzy logic to binary similarity indices in ecology. Methods Ecol Evol 6(7):853–858. https://doi.org/10.1111/2041-210X.12372
Beck T, Bos U, Wittstock B, Baitz M, Fischer M, Sedlbauer K (2010) LANCA. Land use indicator value calculation in life cycle assessment. Fraunhofer Verlag, Stuttgart
Bos U, Horn R, Beck T, Lindner JP, Fischer M (2016) LANCA — characterization factors for life cycle impact assessment. Version 2.0. Fraunhofer Verlag, Stuttgart, Stuttgart
Brentrup F, Küsters J, Lammel J, Kuhlmann H (2002) Life Cycle Impact assessment of land use based on the hemeroby concept. Int J Life Cycle Assess 7(6):339–348. https://doi.org/10.1007/BF02978681
Caniani D, Labella A, Lioi DS, Mancini IM, Masi S (2016) Habitat ecological integrity and environmental impact assessment of anthropic activities: A GIS-based fuzzy logic model for sites of high biodiversity conservation interest. Ecol Indic 67:238–249. https://doi.org/10.1016/j.ecolind.2016.02.038
Chaudhary A, Brooks TM (2018) Land use intensity-specific global characterization factors to assess product biodiversity footprints. Environ Sci Technol 52(9):5094–5104. https://doi.org/10.1021/acs.est.7b05570
Chaudhary A, Verones F, de Baan L, Hellweg S (2015) Quantifying land use impacts on biodiversity: combining species-area models and vulnerability indicators. Environ Sci Technol 49(16):9987–9995. https://doi.org/10.1021/acs.est.5b02507
Coelho CRV, Michelsen O (2014) Land use impacts on biodiversity from kiwifruit production in New Zealand assessed with global and national datasets. Int J Life Cycle Assess 19(2):285–296. https://doi.org/10.1007/s11367-013-0628-7
Côté S, Bélanger L, Beauregard R, Thiffault É, Margni M (2019) A conceptual model for forest naturalness assessment and application in Quebec’s Boreal Forest. Forests 10(4):325. https://doi.org/10.3390/f10040325
Curran M, de Souza DM, Antón A, Teixeira RFM, Michelsen O, Vidal-Legaz B, Sala S, i Canals LM (2016) How well does LCA model land use impacts on biodiversity?—a comparison with approaches from ecology and conservation. Environ Sci Technol 50(6):2782–2795. https://doi.org/10.1021/acs.est.5b04681
Das B, Pal SC (2020) Assessment of groundwater vulnerability to over-exploitation using MCDA, AHP, fuzzy logic and novel ensemble models: a case study of Goghat-I and II blocks of West Bengal. India Environmental Earth Sciences 79(5):104. https://doi.org/10.1007/s12665-020-8843-6
de Baan L, Mutel CL, Curran M, Hellweg S, Koellner T (2013) Land use in life cycle assessment: global characterization factors based on regional and global potential species extinction. Environ Sci Technol 47(16):9281–9290. https://doi.org/10.1021/es400592q
de Groot R, Fisher B, Christie M (2010) The economics of ecosystems and biodiversity: ecological and economic foundations. In: Kumar P (ed) The economics of ecosystems and biodiversity. Ecological and Economic Foundations. Earthscan, London
Dilli R, Argou A, Pilla M, Pernas AM, Reiser R, Yamin A (2018) Fuzzy logic and MCDA in IoT resources classification. In: Computing, Haddad (Ed.) 2018 – The 33rd Annual ACM Symposium, pp 761–766
Eberle U (2018) Land use impacts: comparing Irish and German milk production. In: Kasetsart University, King Mongkut's University of Technology Thonburi, National Science and Technology Development Agency (eds), p 90
Eberle U, Lindner JP (2015) Biodiversity impact: case study beef production. In: Scalbi S, Loprieno AD, Sposato P (eds) International conference on Life Cycle Assessment as reference methodology for assessing supply chains and supporting global sustainability challenges. LCA for feeding the planet and energy for life, pp 302–306
Fehrenbach H, Grahl B, Giegrich J, Busch M (2015) Hemeroby as an impact category indicator for the integration of land use into life cycle (impact) assessment. Int J Life Cycle Assess 20(11):1511–1527. https://doi.org/10.1007/s11367-015-0955-y
Föst P (2019) Biodiversitätswirkung der Bereitstellung von Batterierohstoffen. Master, Hochschule Bochum
Gabel VM, Meier MS, Kopke U, Stolze M (2016) The challenges of including impacts on biodiversity in agricultural life cycle assessments. J Environ Manage 181:249–260. https://doi.org/10.1016/j.jenvman.2016.06.030
Geß A (2020) Biodiversity Impact Assessment of Grazing Sheep. In: Albrecht S (ed) Ökobilanz-Werkstatt
Geyer R, Stoms DM, Lindner JP, Davis FW, Wittstock B (2010) Coupling GIS and LCA for biodiversity assessments of land use. Part 1: Inventory modeling. Int J Life Cycle Assess 15(5):454–467. https://doi.org/10.1007/s11367-010-0170-9
Guo ZX, Wong WK (2013) 2 - Fundamentals of artificial intelligence techniques for apparel management applications. In: Guo ZX, Leung SYS (eds) Wong WK. Woodhead Publishing Series in Textiles. Woodhead Publishing, Optimizing decision making in the apparel supply chain using artificial intelligence (AI), pp 13–40
i Canals LM, Bauer C, Depestele J, Dubreuil A, Knuchel RF, Gaillard G, Michelsen O, Müller-Wenk R, Rydgren B (2007) Key elements in a framework for land use impact assessment within LCA (11 pp). Int J Life Cycle Assess 12(1):5–15. https://doi.org/10.1065/lca2006.05.250
IPBES (2018) The IPBES assessment report on land degradation and restoration. Bonn, Germany
Klein D, Wolf C, Schulz C, Weber-Blaschke G (2015) 20 years of life cycle assessment (LCA) in the forestry sector: state of the art and a methodical proposal for the LCA of forest production. Int J Life Cycle Assess 20(4):556–575. https://doi.org/10.1007/s11367-015-0847-1
Koellner T (2000) Species-pool effect potentials (SPEP) as a yardstick to evaluate land-use impacts on biodiversity. J Cleaner Prod 8(4):293–311. https://doi.org/10.1016/S0959-6526(00)00026-3
Koellner T, de Baan L, Beck T, Brandão M, Civit B, Goedkoop M, Margni M, i Canals LM, Müller-Wenk R, Weidema B, Wittstock B (2013a) Principles for life cycle inventories of land use on a global scale. Int J Life Cycle Assess 18(6):1203–1215. https://doi.org/10.1007/s11367-012-0392-0
Koellner T, de Baan L, Beck T, Brandão M, Civit B, Margni M, i Canals LM, Saad R, de Souza DM, Müller-Wenk R (2013b) UNEP-SETAC guideline on global land use impact assessment on biodiversity and ecosystem services in LCA. Int J Life Cycle Assess 18(6):1188–1202. https://doi.org/10.1007/s11367-013-0579-z
Koellner T, Scholz RW (2008) Assessment of land use impacts on the natural environment. Int J Life Cycle Assess 13(1):32–48. https://doi.org/10.1065/lca2006.12.292.2
Kowarik I (1990) Some responses of flora and vegetation to urbanization in Central Europe. In: Sukopp H, Hejny S, Kowarik I (eds) Urban ecology. Plants and plant communities in urban environments, SBP Academic, pp 45–75
Kyläkorpi L, Rydgren B, Ellegard A, Miliander S, Grussel E (2005) The biotope method 2005. A method to assess the impact of land use on biodiversity, Vattenfall, Stockholm
Landesamt für Umweltschutz Sachsen-Anhalt – Halle (ed) (2004) Karte der Potentiellen Natürlichen Vegetation Erstellung und Anwendung, Sonderheft 2, Bonn
Lindeijer E, Müller-Wenk R, Steen B (2002) Impact assessment of resources and land use. In: Udo de Haes HA (ed) Life-cycle impact assessment. Striving towards best practice. SETAC Press, Pensacola
Lindner J, Niblick B, Luick R, Eberle U, Schmincke E, Bos U, Schwarz S, Blumberg M, Urbanek A (2014) Proposal of a unified biodiversity impact assessment method. In: Schenck R, Huizenga D (eds) LCA Food 2014. Proceedings of the 9th International Conference on Life Cycle Assessment in the Agri-food Sector : 8–10 October 2014, San Francisco. ACLCA, Vashon
Lindner JP (2016) Quantitative Darstellung der Wirkungen landnutzender Prozesse auf die Biodiversität in Ökobilanzen. Doctoral dissertation, University of Stuttgart
Lindner JP, Fehrenbach H, Winter L, Bischoff M, Bloemer J, Knuepffer E (2019) Valuing biodiversity in life cycle impact assessment. Sustainability 11(20):5628. https://doi.org/10.3390/su11205628
Lindqvist M, Palme U, Lindner JP (2016) A comparison of two different biodiversity assessment methods in LCA—a case study of Swedish spruce forest. Int J Life Cycle Assess 21(2):190–201. https://doi.org/10.1007/s11367-015-1012-6
Maier S, Lindner J, Francisco J (2019) Conceptual Framework for Biodiversity Assessments in Global Value Chains. Sustainability 11:1841
Makisara K, Katila M, Perasaari J (2019) National Forest Inventory. https://kartta.paikkatietoikkuna.fi/?lang=en. Accessed 29 Jan 2020
Michelsen O (2008) Assessment of land use impact on biodiversity. Int J Life Cycle Assess 13(1):22–31. https://doi.org/10.1065/lca2007.04.316
Michelsen O, Lindner J (2015) Why include impacts on biodiversity from land use in LCIA and how to select useful indicators? Sustainability 7(5):6278–6302. https://doi.org/10.3390/su7056278
Michelsen O, McDevitt JE, Coelho CRV (2014) A comparison of three methods to assess land use impacts on biodiversity in a case study of forestry plantations in New Zealand. Int J Life Cycle Assess 19(6):1214–1225. https://doi.org/10.1007/s11367-014-0742-1
Millennium Ecosystem Assessment (2005) Ecosystems and human well-being. Synthesis. Island Press, Washington D.C.
Myllyviita T, Sironen S, Saikku L, Holma A, Leskinen P, Palme U (2019) Assessing biodiversity impacts in life cycle assessment framework - comparing approaches based on species richness and ecosystem indicators in the case of Finnish boreal forests. J Clean Prod 236:117641. https://doi.org/10.1016/j.jclepro.2019.117641
Olson DM, Dinerstein E, Wikramanayake ED, Burgess ND, Powell GVN, Underwood EC, D’amico JA, Itoua I, Strand HE, Morrison JC, Loucks CJ, Allnutt TF, Ricketts TH, Kura Y, Lamoreux JF, Wettengel WW, Hedao P, Kassem KR (2001) Terrestrial ecoregions of the world. A new map of life on Earth. Bioscience 51(11):933. https://doi.org/10.1641/0006-3568(2001)051[0933:TEOTWA]2.0.CO;2
Ong HC, Tilahun S (2011) Integrating fuzzy preference in genetic algorithm to solve multiobjective optimization problems. Far East J Math Sci 55:165–179
Pavan ALR, Ometto AR (2016) Regionalization of land use impact models for life cycle assessment. Recommendations for their use on the global scale and their applicability to Brazil. Environ Impact Assess Rev 60:148–155. https://doi.org/10.1016/j.eiar.2016.05.001
Prato T (2007) Assessing ecosystem sustainability and management using fuzzy logic. Ecol Econ 61(1):171–177. https://doi.org/10.1016/j.ecolecon.2006.08.004
Rossi V, Lehesvirta T, Schenker U, Lundquist L, Koski O, Gueye S, Taylor R, Humbert S (2018) Capturing the potential biodiversity effects of forestry practices in life cycle assessment. Int J Life Cycle Assess 23(6):1192–1200. https://doi.org/10.1007/s11367-017-1352-5
Singh H, Gupta MM, Meitzler T, Hou ZG, Garg KK, Solo AMG, Zadeh LA (2013) Real-life applications of fuzzy logic. Adv Fuzzy Syst 2013:1–3. https://doi.org/10.1155/2013/581879
Souza DM, Teixeira RFM, Ostermann OP (2015) Assessing biodiversity loss due to land use with Life Cycle Assessment: are we there yet? Glob Change Biol 21(1):32–47. https://doi.org/10.1111/gcb.12709
Tavana M, Sodenkamp MA (2010) A fuzzy multi-criteria decision analysis model for advanced technology assessment at Kennedy Space Center. J Oper Res Soc 61(10):1459–1470. https://doi.org/10.1057/jors.2009.107
Turner PAM, Ximenes FA, Penman TD, Law BS, Waters CM, Grant T, Mo M, Brock PM (2019) Accounting for biodiversity in life cycle impact assessments of forestry and agricultural systems—the BioImpact metric. Int J Life Cycle Assess 24(11):1985–2007. https://doi.org/10.1007/s11367-019-01627-5
Vrasdonk E, Palme U, Lennartsson T (2019) Reference situations for biodiversity in life cycle assessments: conceptual bridging between LCA and conservation biology. Int J Life Cycle Assess 24(9):1631–1642. https://doi.org/10.1007/s11367-019-01594-x
Weidema BP, Lindeijer E (2001) Physical impacts of land use in product life cycle assessment. Final report of the EURENVIRON-LCAGAPS sub-project on land use, Lyngby
Winter L, Lehmann A, Finogenova N, Finkbeiner M (2017) Including biodiversity in life cycle assessment – state of the art, gaps and research needs. Environ Impact Assess Rev 67:88–100. https://doi.org/10.1016/j.eiar.2017.08.006
World Wildlife Fund (2006) WildFinder: Online database of species distributions. www.worldwildlife.org/WildFinder. Accessed Mar 2017
WWF (2016) Living Planet Report 2016. Risk and resilience in a new era WWF International, Gland, Switzerland
Yatsalo B, Korobov A, Martínez L (2017) Fuzzy multi-criteria acceptability analysis: a new approach to multi-criteria decision analysis under fuzzy environment. Expert Syst Appl 84:262–271. https://doi.org/10.1016/j.eswa.2017.05.005
Zadeh LA (1965) Fuzzy sets. Inform Control 8(3):338–353. https://doi.org/10.1016/S0019-9958(65)90241-X
Zadeh LA (1973) Outline of a new approach to the analysis of complex sand decision processes. IEEE Trans Syst Man Cybern B Cybern SMC 3(1):28–44. https://doi.org/10.1109/TSMC.1973.5408575
Zeleny M (2011) Multiple criteria decision making (MCDM). From paradigm lost to paradigm regained? J Multi-Crit Decis Anal 18(1–2):77–89. https://doi.org/10.1002/mcda.473
Zimmermann HJ (2010) Fuzzy set theory. WIREs Comp Stat 2(3):317–332. https://doi.org/10.1002/wics.82
Acknowledgements
We thank the experts who participated in the interviews, namely Rainer Luick, Timo Lehesvirta, Petri Ahlroth, and Lauri Saaristo. The method described in this paper is also part of the doctoral work of the lead author, Jan Paul Linder (reference: Lindner 2016).
Funding
Open Access funding enabled and organized by Projekt DEAL. This work has been partially funded by the German Federal Agency for Nature Conservation with funds from the German Federal Ministry for the Environment, Nature Conservation and Nuclear Safety, and by the Erich Ritter Foundation.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no competing interests.
Additional information
Communicated by Thomas Koellner.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Lindner, J.P., Eberle, U., Knuepffer, E. et al. Moving beyond land use intensity types: assessing biodiversity impacts using fuzzy thinking. Int J Life Cycle Assess 26, 1338–1356 (2021). https://doi.org/10.1007/s11367-021-01899-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11367-021-01899-w