
Abstract
Macrobenthic invertebrate communities serve as markers of anthropogenic stress in freshwater ecosystems. In this study, 17 sampling sites were selected from two Nile river subbranches (El-Rayah El-Behery and El-Rayah El-Nassery) and subjected to different anthropogenic influences to explore the ecological environment and characteristics of macrobenthos communities. Macrobenthos were studied using taxonomic diversity and biological trait analysis to investigate how human activity and variation in water quality affect their structure and function. A total of 37 taxa represented by 43,389 individuals were recognized. The communities are composed chiefly of Oligochaeta and aquatic insects. Multivariate statistical analyses found that the most influential environmental variables in the structural and functional community were sodium, dissolved oxygen, silicate, pH, calcium, and cadmium. At high levels of pollution, notably sewage and industrial pollution in the northern part of El-Rayah El-Behery, characteristics such as larger body size, detritus feeders, burrowers, and high tolerance to pollution predominated, whereas at low levels of pollution, features such as small body sizes, scraper and predator feeders, intolerant and fairly tolerant of pollution, and climber and swimmer mobility are predominant. The results confirm our prediction that the distribution of macroinvertebrate traits varies spatially in response to environmental changes. The diversity-based method distinguished impacted sewage and industrial sites from thermal effluent sites, while the trait-based approach illustrated an apparent variance between the ecological status of contaminated regions. Therefore, the biological features should be employed in addition to structural aspects for assessing the biodiversity of macroinvertebrate communities under environmental stressors.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Macrobenthic invertebrates are considered efficient markers to assess and screen the health of freshwater ecosystems because they are prevalent and sensitive to environmental stressors (Piló et al. 2016; Borja et al. 2010). These motivate significant river ecosystem cycle processes, such as organic material retention and breakdown, nutrient and mineral recycling, and contribution to energy processing at various trophic levels (Butkas et al. 2011; Mangadze et al. 2019). Recent studies illustrated a considerable correlation between these invertebrates and ecological issues. Therefore, this correlation analysis has significantly improved river ecosystems (Butkas et al. 2011). Assessing the functional features and structure of the macrobenthic assemblages can highlight how ecological changes affect benthic assemblages (Piló et al. 2016; Kenny et al. 2018; Hu et al. 2019). Several authors concluded that the macrobenthic functional traits could illustrate the environmental disturbance better than these macrobenthic invertebrates themselves (Piló et al. 2016; Nasi et al. 2018; D’Alessandro et al. 2020). There is a strong relationship between macrobenthic invertebrates’ functional features, environmental processes, and ecosystem functions (Cadotte et al. 2011). This macrobenthos can respond swiftly to many stressors (Mouillot et al. 2013; Voß and Schäfer 2017).
Most ecological research focuses on structural diversity, abundance, and biomass of species communities, whereas functional adaptation to environmental variables is rarely evaluated. The functional structure of the macrobenthos is expressed via a collection of traits that define the morphological and behavioral features of the taxa. The stability and function of communities and ecosystems are influenced by their characteristics and interactions (Loreau et al. 2001), which also demonstrate a correlation between alterations in the macrobenthic community and environmental factors (Lavorel and Garnier 2002). Biological trait analysis (BTA) is a representative trait-based technique that utilizes collections of the biological traits of organisms to illustrate the variation pattern of the functional features along spatial or temporal gradients (Bremner et al. 2006). BTA and taxonomic analysis should be integrated (Díaz and Cabido 2001; Villéger et al. 2010). Functional data is quantitatively characterized as a set of biological features, with values supplied for each taxon based on lifestyle and activity mode. Integrating the functional characteristics of species composition with all species’ biomass or abundance values allows for their quantification. This expression, like structural data, can be used to study gradients and connections to environmental variables. Consequently, the methodology establishes links among species, environmental conditions, and ecological activities. However, BTA was initially developed and used for freshwater systems (e.g., Zhang et al. 2021; Laini et al. 2019; Lamouroux et al. 2004; Charvet et al. 2000; Usseglio-Polatera et al. 2000), but BTA in the macrobenthic community does not perform on River Nile before.
The Nile River is the principal supply of fresh water for all purposes and uses in Egypt, supplying the country with more than 95% of its freshwater needs. Many branches, irrigation canals, and streams originate from the Nile to form the agricultural system, extending for 31,000 km (Goher et al. 2019). The irrigation canals and rayahs, like the Nile River, are primarily used for drinking, irrigation, industry, navigation, and fishing. Consequently, due to the increased population and industrial development, massive organic and inorganic waste and heavy metals are thrown into the Nile and travel into the Mediterranean Sea via Sudan and Egypt (Goher et al. 2019, 2021). Metals are more concentrated in sediments than in the water body in rivers and watercourses (Shyleshchandran et al. 2018). They are discharged into the water body under appropriate hydrological and chemical circumstances, resulting in water pollution and harming aquatic organisms (Kouidri et al. 2016). Nutrient, metals, and organic pollution can cause variations in macrobenthos community structures by decreasing the abundance of sensitive species, species richness, and diversity while increasing the abundance of tolerant species (Birk et al. 2020). The authors hypothesized that the diversity-based and trait-based methods would describe the response of the macrobenthic community to different environmental stresses in various approaches.
Therefore, in this study, the diversity-based and trait-based methods were performed (1) to assess the variations in taxonomic and functional macroinvertebrate community structures toward different stressors within El-Behery and El-Nassery rayahs, (2) to determine the significant environmental factors prompting macrobenthos species and their biological traits, and (3) identify the most sensitive traits to environmental stressors.
Materials and methods
Study area and sampling locations
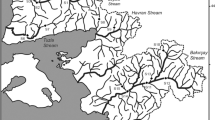
In the Delta region, the Nile is divided into two main branches and four subbranches (irrigation canals, or rayahs in the colloquial dialect): the El-Behery, El-Nassery, El-Tawfiky, and El-Menoufy rayahs (Talab et al. 2016; Bendary and Ibrahim 2021). The El-Behery and El-Nassery rayahs provide fresh water for millions of people in the West Delta area and the Alexandria Governorate (Goher 2015). Both rayahs arise from the Rosetta branch downstream from the Delta Barrage, heading northwest to the Mediterranean Sea. El-Rayah El-Behery is 215 km long that is refilled with water from the Rosetta branch at Damanhur Governorate via the El-Mahmoudia Canal. The El-Mahmoudia Canal is the northern portion of El-Rayah El-Behery (from sites B6–B9). It originated from the Rosetta branch and is considered a multi-stressor area in our study due to the discharged sewage and industrial wastes via Rosetta branch. Unfortunately, Rosetta branch collects massive amounts of pollutants and wastewater from several sources, including the El-Rahawy Drain (El Sayed et al. 2020). El-Rayah El-Nassery is 200 km long and connects to the El-Nubaria Canal, which splits off from El-Rayah El-Behery at Bolin Bridge and runs to the Mediterranean Sea via Mariout Lake. The section from sites N4 to N8 is called El-Nubaria Canal. The EI-Nubaria canal is vital for fishing and agriculture; however, it receives different wastewater from the surrounding farmlands, electric power plants, and small water treatment stations (see Fig. 1 and Table 1 for more details).

Map of the study area indicating the sampling sites in El-Rayah El-Behery (B1-B9) and El-Rayah El-Nassery, (N1-N8), Nile River
According to the contamination degree (Cd) and potential ecological risk (RI) indices (El Sayed et al. 2022) and water quality (WQ) Egyptian standards (Habash et al. 2018), the study area sites are classified into three main sectors: sector 1 includes sites from B6 to B9 (El-Mahmoudia Canal) with the highest values of contaminations; sector 2 includes sites from N5 to N8 (from El-Nubaria Canal) which recorded the lowest values of contaminations, and sector 3 contains the remains that considered as non-contaminated sites.
Sampling
Nine sites from El-Rayah El-Behery (B1–B9) and eight sites from El-Rayah El-Nassery (N1–N8) were sampled. Sampling was performed seasonally in the spring, summer, and autumn of 2014 and winter of 2015. Water samples were collected using an A 2 L polyvinyl chloride water sampler. An Ekman Grab sampler collected samples of macrobenthic invertebrates (225 cm2). Each specimen was sieved through a 0.5-mm screen and stored in a 10% buffered formalin. In the laboratory, macroinvertebrate individuals were counted and classified to the species or genus level as far as possible.
Environmental data
The following variables were measured as discussed in American Public Health Association–defined methods (APHA 2005): water temperature (°C), pH, total dissolved solids (TDS), dissolved oxygen (DO), chemical oxygen demand (COD), ammonia (NH4–N), nitrogen dioxide (NO2–N), phosphate (PO4–P), silicate (SiO4), sulfate (SO3), calcium (Ca), chlorine (Cl), sodium (Na), potassium(K), lead (Pb), and cadmium (Cd) as shown in the supplementary information (ST1), to inspect the effect of anthropogenic stressors on the macrobenthic species’ distribution and their biological traits.
Biological traits analysis (BTA)
Ten biological traits were chosen depending on their ability to define the critical biological and environmental processes related to anthropogenic and ecological environmental stresses: morphology, body size, longevity, feeding mode, mobility, habitat, substrate affinity, reproductive technique, larval environmental development, and pollution tolerance. The 10 traits were divided into 40 categories that represented the behavior and strategy of the organisms in greater detail. For example, the “feeding mode” trait categories were filter feeders, detritus feeders, collectors/gatherers, scrapers/grazers, engulfers/predators, and scavengers (Table 2). The association of each taxon with the categories of each trait was quantitively scored based on the “fuzzy coding” method (Chevenet et al. 1994; Bremner et al. 2003). Each feature was allocated a functional biological score ranging from 0 to 3, where 0 displays no affinity, 1 or 2 represents moderate affinity, and 3 illustrates a high relationship for the specified trait category. For the BTA, three matrices were created: the density of taxa in each station (matrix “taxa by stations”), the functional biological score of each taxon (matrix “taxa by traits”), and a functional traits score combined with the abundance value for all constituent species in each station (matrix “traits by stations”) (Bremner et al. 2003). To equilibrium the abundance values between abundant and rare species without losing density effects, the initial matrices were transformed by log (1 + x) before the calculations. These three matrices were created for four seasons to determine seasonal impacts. Biological trait data from a range of published resources, journal articles, and online databases (https://www.epa.gov/risk/freshwater-biological-traits-database-traits) were collected to generate the “taxa by traits” matrix.
Statistical analysis
Fuzzy correspondence analysis (FCA) was utilized to arrange the “traits by stations” matrix on a two-dimensional plane. Which traits influenced the species’ distribution inside and between the various sites in rayahs and seasons are clear. The FCA’s correlation ratio (CR) represents each trait’s contribution to the total variance. The features with a CR value larger than 0.1 are regarded as having the most influence on the variation among sites. FCA was conducted using R-4.1.2 (ADE4-ADE4TKGUI) (Thioulouse et al. 1997).
Multivariate statistical analyses were conducted using Canoco 4.5 software (Ter Braak and Smilauer 2002). Principal component analysis (PCA) was used to evaluate seasonal and spatial patterns of environmental factors. Redundancy analysis (RDAs) was done to highlight the correlations between macrobenthic species and environmental factors, likewise the correlation between trait features and environmental factors. RDAs was suitable for data analysis as in the preliminary detrended correspondence analysis (DCA) for species composition and trait; the lengths of the gradient for the first axis was less than 3 SD (Lepš and Šmilauer 2003). Data for the PCA and RDAs were log (x + 1) transformed. The significance of the variables was determined using a Monte Carlo permutation test with 499 permutations, and only the significant environmental factors were plotted in the RDAs.
Non-parametric Kruskal–Wallis tests were done to show the significant differences between the sampling sites and seasons for the environmental variables and macrobenthos structural and trait composition. Statistical calculations were performed using SPSS 24. All statistical analysis was done at a 95% confidence interval.
Results
Environmental variables
Among environmental factors, remarkable variations were found across sampling sites and seasons (Supplementary Tables S1–S5). Figure 2 illustrates the PCA plots of environmental data for the rayahs canals in different seasons. The first two axes of the PCA explained 85.8 to 93.9% of the overall variability. El-Mahmoudia Canal sites B6, B8, and B7 were associated with higher average values of NH4 (2827 µg/l), NO2 (166 µg/l), and PO4 (89 µg/l), respectively, than other sites (p < 0.05). Likewise, B9 showed a lower average content of DO (3.2 mg/l) (p < 0.05) and the highest average value of Ca (40.4 mg/l) (p < 0.05). El-Nubaria Canal sites N8 and N4 had higher average value of TDS (506 mg/l) and SiO4 (3.5 mg/l), respectively (p < 0.05). All environmental variables differed significantly between seasons (p < 0.05) except cadmium. The maximum values of NH4 (6733 mg/l), TDS (637 mg/l), and Ca (47.8 mg/l) were highest in the winter, while the values of NO2 (447 mg/l), Pb (60 µg/l), and pH (8.8) were highest in the autumn, and the highest value of SiO4 (5.4 µg/l) occurred in summer.

Principal component analysis plots of the relevant environmental variables in the El-Rayah El-Behery and El-Rayah El-Nassery for different seasons
Taxonomic composition and seasonal variations of the macrobenthic community
An overall of 37 taxa was recorded (43,389 individuals/m2) belonging to the Insecta, Crustacea, Gastropoda, Bivalvia, and Oligochaeta (Suppl., Table S5). Oligochaeta generally dominated (seven species, 1772 individuals/m2, 69%), especially Limnodrilus udekemianus. The second-largest group was Insecta (15 species, 506 individuals/m2, 20%), with the dominance of Chironomidae larvae. Gastropoda is represented by eight species (201 individuals/m2, 8%), and Bivalvia is represented by four species (67 individuals/m2, 3%). Crustacea (three species, 6 individuals/m2) were the least abundant group. A significant difference among the sampling sites was noticed in the species composition of macrobenthos (p < 0.05). The highest density was recorded at site B9 (5281 individuals/m2), followed by B6 (5262 individuals/m2). Site N8 had the most remarkable diversity (18 species). Site B2 contained the lowest number of taxa and density (six species and 476 individuals/m2). The highly tolerant species, such as L. udekemianus and Chironomidae larvae, were the taxa with the maximum frequency, disseminated at all the sites. The sensitive polluted species such as Cloenon sp., Caenis sp., and Hydropsychidae larvae rarely occurred and disappeared in El-Mahmoudia Canal sites. The species abundance and diversity over the two rayahs increased from the southern sites toward the northern sites, which were mainly characterized by oligochaetes species (Fig. 3). A significant difference between El-Mahmoudia Canal sites and the other remaining sires was observed (p < 0.05). The distribution of benthic macrofauna in the irrigation rayahs varied greatly between seasons. Commonly, the abundance and highest number of taxa were recorded in spring and winter related to summer and autumn (Suppl. Table S6). The highest diversity and density were observed during the winter (3254 individuals/m2 and 29 species). The lowest density was noted during the autumn (1921 individuals/m2), while the lowest number of taxa was observed during the spring (21 species). L. udekemianus and Chironomidae larvae were the dominant species in all seasons. The most abundant individuals in the winter were Micronecta sp., averaging 313.2 individuals/m2. The most abundant individuals in the summer were G. senaariensis, averaging 473.5 individuals/m2.

Taxa distribution within El-Rayah El-Behery and El-Rayah El-Nassery stations. The size of the squares is proportional to the density (the bigger squares, the higher density)
Fuzzy correspondence analysis (FCA)
Biological trait scores obtained using the “fuzzy coding” technique are shown in the supplementary data (Table S7). In all seasons, the chi-square test (or Fisher’s exact test) was done for all traits and revealed a significant correlation (p < 0.001) between trait categories and sample sites. FCA for the matrix (“traits by stations”) was conducted separately for the four seasons (Fig. 4). In the spring, the first two FCA axes expressed 59% of the overall variance, with 39% accounted for by axis 1 and 20% by axis 2. The influence of each trait on this variance is explained in the CR (Table 3). The main biological features separated along FCA axis 1 were body size, mobility, longevity, and substrate affinity. In the summer, the FCA axes represent 68% of the overall variance; axis 1 counted 53% and axis 2 15% of the variability. The biological traits on FCA axis 1 were morphology, body size, habitat, reproductive technique, and larval development. Mobility was highly correlated with both axes. In the autumn, 55% of the overall variance was expressed by the first two FCA axes: 36% by axis 1 and 19% by axis 2. Body size, longevity, and mobility were correlated with axis 1, and habitat and larval development were associated with axis 2. Larval development was connected with axes 1 and 2, and pollution tolerance and substrate affinity were not correlated with axes 1 and 2. During the winter, the first two FCA axes represented 64% of the overall variance. Axis 1 explained 45% of the overall variability in body size, longevity, mobility, habitat, reproductive technique, and larval development. Body size distribution and longevity categories along axis 2 explained 19% of the overall variability.

Factor map of the sampling stations obtained by FCA for different seasons: A spring, B summer, C autumn, and D winter
The high-contaminated sector had similar distributions of traits, dominated by species tolerant of pollution, detritus feeders, burrowers, mud substrate affinity, and large body size (P4, F2, mob1, SA5, and S4). At the same time, the low-contaminated sector is characterized by the following trait categories, very tolerant and fairly tolerant of pollution, filter-feeder, scraper/grazer, and climber (P4, P3, F2, F4, and mob5). The non-contaminated sites had higher percentages of small body size, engulfer/predator and scavenger feeders, and swimmers (S1, F5, F6, and mob6) than the El-Mahmoudia canal and El-Nubaria Canal sites. A significant difference among seasons was observed for the trait categories of macrobenthos (p < 0.05). Regarding the different contaminated sectors, Kruskal–Wallis test exposed that there were significant variances (p < 0.05) in trait categories (i.e., S3, L2, L4, F1, F4, H2, H4, SA2, mob4, and LD3).
Associations between species and environmental variable
RDA was conducted to study the correlations between macrobenthic invertebrate species and environmental factors (Fig. 5A). The cumulative variance of taxa data along the first two axes and the cumulative variance of the taxa-environment relationship along the first two axes constituted the same percentage, 44.9% of the total variability. The arrows describe the environmental factors and point in the direction of the factor with the maximum variation. Na, DO, SiO4, and pH were the greatest significant variables in taxa occurrence (p < 0.05). Monte Carlo permutation tests demonstrated that Na was the most significant environmental factor associated with community composition (p < 0.05; F-ratio = 4.64). The second most significant factor (p < 0.05; F-ratio = 1.99) was DO, which influenced the overall variance of species composition. The plot shows that strong relationships existed between (NH4, Na, and TDS) and the individuals of (Branchiura sowerbyi, Bulinus truncates, Viviparus contectus, Lanistes carinatus, and Helobdella conifer). The DO was the most effective variable for the distribution of Micronecta sp., Cloenon sp., Ischnura sp., and Caridina nilotica. SiO4 was the most effective variable for distributing Hydroptilidae larvae, Caenis sp., and Melanoides tuberculata.

A RDA triplot illustrating associations between macroinvertebrate species and the most significant environmental variables. See Fig. 3 for complete species names. B RDA triplot explaining associations between macroinvertebrate traits and the most important environmental variables. See Table 2 for the meaning of the trait codes
Relations between traits and environmental variable
Based on RDA analysis, the cumulative percentage variance along the first two axes recorded 47.9% of the total variance of macrobenthic trait categories, and the cumulative percentage variance of trait features-environment relation along the first axes 1 and 2 recorded 59.2% of the total variance. The RDA results showed that Ca and Cd (p < 0.05; F-ratio = 4.9 and 2.02, respectively) significantly affected the trait categories distribution. Burrow-dwelling, mud substratum affinity, burrowers, detritus feeder, > 20-mm-large body size, and very tolerant pollution (H1, SA5, mob1, F2, S4, and P4) were associated with high levels of Ca and Cd. Free living, bentonic larval development, and small body size (5–10 mm) (H2, LD2, S2) were related to the DO concentration (Fig. 5B).
Discussion
Environmental variables of El-Rayah El-Behery and El-Rayah El-Nassery
El-Mahmoudia Canal was the most polluted area in our study. High concentrations of nitrogen and phosphorous nutrient salts besides cations and anions caused due to these numerous anthropogenic stressors (Goher et al. 2021), affecting biodiversity and causing disruption in macroinvertebrates (El-Shabrawy and Goher 2012; El-Otify and Iskaros 2015). El Sayed et al. (2022) also recorded that the various anthropogenic stressors in El-Mahmoudia Canal contributed to higher aggregations of heavy metals. The highest concentrations of most abiotic parameters were recorded in the El-Mahmoudia Canal, indicating its state of constant contamination. This result is in line with El Sayed et al. (2020) in Rosetta branch and (Talab et al. 2016) in the rayahs. The low DO levels in this canal may be correlated with the extreme transmission of organic pollutants and nutrients. In this study, the highest average value of silica was recorded at site N4 (3.5 mg/l), and the lowest average value was noted at site B5 (1.7 mg/l) in contravention with other nutrients; this indicated that sewage and industrial effluents do not significantly affect the distribution of reactive silica in irrigation canals, where the accurate concentrations of river silica levels were shown to be dependent on the motion of underlying soils and the degradation of silicate rocks (Wang et al. 2013; El-Otify and Iskaros 2015).
Macrobenthic invertebrate community structures
The quantitative and qualitative investigation of the macrobenthos assemblage in El-Behery and El-Nassery rayahs demonstrated that it was mainly composed of Oligochaeta and aquatic insects. L. udekemianus and Chironomidae larvae were the most abundant and widely distributed species. The community composition of macrobenthos in the irrigation canals of the Nile River has not changed much since previous investigations (Mola and Abdel Gawad 2014; Saad Abd El-Halim et al. 2015; Khalifa and Bendary 2016; El-Damhogy et al. 2017; Bendary and Ibrahim 2021). Yap et al. (2006) described that Limnodrilus sp. are contaminant-resistant species that can survive in habitats with poor water quality. Rashid and Pandit (2014) reported that oligochaetes were the most common benthic macroinvertebrates because they can live in many different environments and tolerate hypoxia due to the breakdown of organic matter in the background. Al-Shami et al. (2010) reported high populations of Chironomus sp. in six Malaysian rivers as the water quality changed from high to moderate contamination. Chironomidae are poor water quality indicators in freshwater environments because of their abundance in sewage-polluted areas (Serra et al. 2017). This dominance of Chironomidae and Oligochaeta was also recorded in other rivers affected by anthropogenic stressors, e.g., the Langat River, Malaysia (Azrina et al. 2006), the Bílina River, Czech Republic (Orendt et al. 2012), the Bzura River, Poland (Grzybkowska et al. 2015), and the Hex River, South Africa (Erasmus et al. 2021).
The reduction in species diversity and abundance at site B2 could be due to the thermal effluence from the electricity plant. Worthington et al. (2015) and Han et al. (2017) stated that the thermal discharge reduced the abundance and richness of macrobenthic invertebrates. Increased Ca levels can induce shell calcification in warm waters (Abd El-Wakeil et al. 2015), which could explain the rise in molluscan quantity at thermal-polluted site B9. Hussein et al. (2011) stated that freshwater snails display a high degree of tolerance and flexibility in a specific range of physicochemical variation. The increase in the quantity of sensitive polluted species Cloenon sp., Caenis sp., and Hydropsychidae larva in the non-contaminated area is explained by Ephemeroptera and Trichoptera taxa increase with an increase in water quality (Erasmus et al. 2021).
Functional diversity pattern of macrobenthic invertebrates
The current investigation of macrobenthic fauna discovered functional diversity patterns associated with contamination aspects and environmental variables. The maximum characteristic patterns were revealed for body size, mobility, feeding mode, substrate affinity, and pollution tolerance. This outcome is in line with other studies in which traits associated with mobility and feeding have been determined to be the most accountable for discriminating between species communities (Paganelli et al. 2012). In all seasons, however, feeding mode traits were weakly associated with both FCA axes; this may be due to the consistent domination of only categories, e.g., in all samples, taxa with detritus feeders predominated over taxa with other feeding modes. The variety of larvae produced by an organism indicates its response to environmental variations. Taxa with planktonic larval development possess a higher spreading potential and a lower hazard of extinction than organisms with other forms of development (McHugh and Fong 2002); this explains the domination of taxa with planktonic larvae in the sites of high environmental stress in the study area.
BTA showed that trait distributions in the El-Mahmoudia Canal were relatively similar. Eutrophication has been a major environmental problem in this region because of excessive nutrient effluents. Waste effluents carrying inorganic and organic contaminants have been shown to impact macrobenthic species assemblages in previous investigations (El Sayed et al. 2020; Bendary and Ibrahim 2021), where the biological traits that reflected the nutrient effluents were mud substratum affinity, large body size, detritus feeders, burrowers, and high tolerance of pollution. The current study’s outcomes align with those of Llanos et al. (2020). Body size is macrobenthos’s most direct and instinctive feature (Edegbene et al. 2020; Odume 2020). Typically, the body size of macrobenthic organisms is correlated with stressor fluctuations, but our results recorded that the body size increased with increasing inorganic and organic contamination. This result is consistent with earlier research indicating that macrobenthic organisms with large body sizes are further common in habitats with somewhat extreme environmental stresses than organisms with small body sizes (Gusmao et al. 2016; Odume 2020; Dong et al. 2021). Additional research has shown that the body size of macrobenthos individuals within a species increases as they approach a pollution source (Ryu et al. 2011). Nevertheless, few studies have indicated that individual size is inversely correlated with the intensity of pollution; i.e., the higher the level of contamination, the smaller the individuals (Piló et al. 2016; Edegbene et al. 2020).
Feeding features convey information about an ecosystem’s ecological status and structural complexity (Tselepides et al. 2000; Bremner et al. 2003; Törnroos and Bonsdorff 2012). In the present study, detritus feeder was the most prevalent feature at highly contaminated sites (El-Mahmoudia Canal). While at less-contaminated sites (El-Nubaria Canal), scrapers were the most pervasive feature. In contrast, the scavenger carnivores and predators were the predominant feeding modes in non-contaminated locations; this could result from higher sediment quality, which allows organisms with diverse feeding habits to increase (Rosenberg 2001; Gusmao et al. 2016). Gaston et al. (1998) and Rakocinski et al. (2000) observed that subsurface deposit feeders predominated in areas with high organic and metal concentrations. In contrast, the number of members of other trophic groups, including carnivores, was low. According to Rawer-Jost et al. (2000) and Erasmus et al. (2021), anthropogenic influences cause a decrease in the number of scrapers.
Mobility of taxa and environmental disruptions were strongly associated (Nasi et al. 2018). In our study, mobility feature altered from burrowers at highly contaminated sites to climber and swimmers at less-contaminated sites and non-contaminated sites, respectively; this is favorable for the macrobenthos because it broadens their range of activity, increases the availability of food, and helps them quickly avoid polluted sediments (Dong et al. 2021).
This gradient and variation in feeding and mobility features in different sectors of the study area demonstrated that the trait-based method could evaluate the variation in the ecological status of the river ecosystem.
Environmental driving factors analysis
The environmental variables influencing macrobenthic organisms are very complicated, and the associations between environmental factors and community structure differ between locations (Zhang et al. 2021). The richness of highly tolerant taxa increased from Sites N1 and B2 towards Sites B6 and B9 with cumulative nutrient (NH4, NO2, PO4) and metal (Na, Ca, Cd, and Pb) concentrations, signifying that each nutrient and metal affected the community structure.
In this study, RDA revealed that Na, DO, SiO4, and pH were the most influential environmental parameters in the macrobenthic community. Clements and Kotalik (2016) reported that anthropogenic disturbances, including industrial and agricultural discharges, are the sources of high concentrations of Na and Ca in the river ecosystems. Lethal effects of significant ions on aquatic macrobenthic are highly varied but mostly connected with increased ionic pressure and the metabolic cost associated with osmoregulation (Williams et al. 1999; Lob and Silver 2012; Cormier et al. 2013). Dissolved oxygen is one of the most significant vital abiotic influences on the aquatic ecosystem. The solubility and availability of nutrients are affected by dissolved oxygen; releasing nutrients from sediments accelerated under hypoxia and hence influences the production of aquatic ecosystems (Abdelmongy and El-Moselhy 2015). Ekau et al. (2010) recorded that the variations in dissolved oxygen concentrations affected the persistence of certain macrobenthic species, consequently on the species composition in the ecosystem. According to the present results, the study area recorded a widespread oxygen variation representing high hypoxia at site B9 (1.8 mg/l) during the autumn as a result of low water levels, in addition to untreated sewage, agricultural, and thermal discharge to high oxygen levels at site N8 (10.9 mg/l) in summer. This significant range in oxygen concentration in the water column has an immediate impact on the oxygenation procedure in the essential sediment deposit, resulting in macrobenthos devastations (Belal et al. 2016). In our study, oligochaetes were generally negatively correlated with DO concentrations. Lim et al. (2006) reported that oligochaetes were the most adaptable under anoxia. The positive correlation between Micronecta sp. and DO attention revealed the sensitivity of Micronecta sp. to pollution.
Because Micronecta sp. breathe DO and do not come up for air, they cannot survive in oxygen-depleted waters and are severely impacted by pollution (Gogala 2009). de Santiago et al. (2020) reported that silicate is the most significant nutrient-influenced macrobenthic community composition. Silica is abundant and constantly prevalent in surface water as dissolved, suspended, and colloidal forms. Dissolved formulae primarily consist of silica acid (El Sayed et al. 2020). Recent studies recorded a linear statistical association between the density of macrobenthos and silicate (Karlson et al. 2007; Ekeroth et al. 2016). pH is the crucial factor for the water criteria and organisms in the aquatic ecosystem (Goher et al. 2014) as it regulates the metal solubility and influences the natural environment. pH range (6.0–9.0) is the potential value for most aquatic organisms, but pH value (7) is the optimum value of the organism’s activity (Chin 2000). Many studies confirmed the significance of pH on macrobenthic organisms’ distribution and composition (Corfield 2000; Tripole et al. 2006; Almagro-Pastor et al. 2015).
The RDA model demonstrated that the distribution of trait categories was significantly influenced by calcium and cadmium. Oug et al. (2012) said that increased cadmium levels are associated with ecological functioning changes. In our study at high cadmium concentrations, trait features such as burrow-dwelling, mud substratum affinity, burrower, detritus feeders, large body size, and very tolerant pollution were predominant. In contrast, at low levels, free living, bentonic larval development, climber mobility, and small body size were characteristic. Leung and Tam (2013) stated that high Cd concentration is negatively associated with the macrobenthos community. Cadmium is one of the maximum significant heavy metals that may cause alterations in the functional traits of macrobenthos communities (Hu et al. 2019). In general, taxonomic and functional trait–based techniques explained that the significant factors responsible for the fluctuations in the macroinvertebrate community structure were the water chemistry variables, especially the metal concentrations. The taxonomic-based method demonstrated an apparent variance between diverse stressors (sewage, agriculture, and thermal effluents). The functional-based process discriminated a clear difference in the ecological status of contaminated sectors. Therefore, the integration between taxonomic and functional diversity can be a technique for assessing the environmental threat in a holistic sight (Ferreira et al. 2012).
Conclusion
The current study on macrobenthic communities in El-Behery and El-Nassery rayahs recorded that the BTA approach perceived specific functional traits associated with pollution gradients and environmental variables. In the northern portion of El-Rayah El-Behery (El-Mahmoudia Canal), the distribution and trait patterns of the macrobenthic fauna indicated a significant level of eutrophication, where the primary functional features of the macrobenthic fauna shifted from small body size, carnivores, crawlers, swimmers, and pollution-intolerant species to large body size, detritus feeders, burrowers, and pollution-tolerant species. The traits “body size,” “feeding mode,” “mobility,” “habitat,” and “pollution tolerance” were identified as potential markers of the response of the macrobenthic fauna to the contaminations in the irrigation rayahs. The environmental parameters impacting macrobenthos’ structural and functional composition in irrigation canals were Na, DO, SiO4, pH, Ca, and cadmium. Human activities and uncontrolled sewage, industrial, and agricultural waste in the study area affected the taxonomic and trait compositions of the macrobenthos. Thus, enhancing the macrobenthos communities will require improved watershed ecology and pollution control.
Data availability
All data will be available from the corresponding author upon request.
Code availability
Non-applicable.
References
Abd El-Wakeil KF, Obuid-Allah AH, Mohamed AH, Abd El-Aziz FEZA (2015) A comparison of molluscan communities in a section of the Nile River at Assiut, Egypt, with different chemical and thermal pollution. Molluscan Res 35:73–80. https://doi.org/10.1080/13235818.2014.919693
Abdelmongy AS, El-Moselhy KhI (2015) Seasonal variations of the physical and chemical properties of seawater at the northern red sea, Egypt. Open J Ocean Coastal Sci 2:1–17. https://doi.org/10.15764/OCS.2015.01001
Almagro-Pastor V, Conradi M, DelValls TA, Riba I (2015) Alterations in the macrobenthic fauna from Guadarranque River (Southern Spain) associated with sediment-seawater acidification deriving from CO2 leakage. Mar Pollut Bull 96:65–75. https://doi.org/10.1016/j.marpolbul.2015.05.044
Al-Shami SA, Rawi CSM, HassanAhmad A, Nor SAM (2010) Distribution of Chironomidae (Insecta: Diptera) in polluted rivers of the Juru River Basin, Penang, Malaysia. J Environ Sci 22:1718–1727. https://doi.org/10.1016/S1001-0742(09)60311-9
APHA (2005) Standard methods for the examination of water and wastewater, 21st edn. American Public Health Association/American Water Works Association/Water Environment Federation, Washington
Azrina MZ, Yap CK, Rahim Ismail A et al (2006) Anthropogenic impacts on the distribution and biodiversity of benthic macroinvertebrates and water quality of the Langat River, Peninsular Malaysia. Ecotoxicol Environ Saf 64:337–347. https://doi.org/10.1016/j.ecoenv.2005.04.003
Belal AAM, El-Sawy MA, Dar MA (2016) The effect of water quality on the distribution of macro-benthic fauna in Western Lagoon and Timsah Lake, Egypt I. Egypt J Aquat Res 42:437–448. https://doi.org/10.1016/j.ejar.2016.12.003
Bendary RE, Ibrahim SM (2021) Diversity and density of macrobenthic invertebrates associated with macrophytes in the El-Rayah El-Nasery and El-Rayah El-Behery, Nile river, Egypt. Egypt J Aquat Biol Fish 25:511–526. https://doi.org/10.21608/EJABF.2021.203279
Birk S, Chapman D, Carvalho L et al (2020) Impacts of multiple stressors on freshwater biota across spatial scales and ecosystems. Nat Ecol Evol 4:1060–1068. https://doi.org/10.1038/s41559-020-1216-4
Borja Á, Elliott M, Carstensen J et al (2010) Marine management - tsowards an integrated implementation of the European marine strategy framework and the water framework directives. Mar Pollut Bull 60:2175–2186. https://doi.org/10.1016/j.marpolbul.2010.09.026
Bremner J, Rogers SI, Frid CLJ (2003) Assessing functional diversity in marine benthic ecosystems: a comparison of approaches. Mar Ecol Prog Ser 254:11–25. https://doi.org/10.3354/MEPS254011
Bremner J, Rogers SI, Frid CLJ (2006) Methods for describing ecological functioning of marine benthic assemblages using biological traits analysis (BTA). Ecol Indic 6:609–622. https://doi.org/10.1016/j.ecolind.2005.08.026
Butkas KJ, Vadeboncoeur Y, Vander Zanden MJ (2011) Estimating benthic invertebrate production in lakes: a comparison of methods and scaling from individual taxa to the whole-lake level. Aquat Sci 73:153–169. https://doi.org/10.1007/s00027-010-0168-1
Cadotte MW, Carscadden K, Mirotchnick N (2011) Beyond species: functional diversity and the maintenance of ecological processes and services. J Appl Ecol 48:1079–1087. https://doi.org/10.1111/j.1365-2664.2011.02048.x
Charvet S, Statzner B, Usseglio-Polatera P, Dumont B (2000) Traits of benthic macroinvertebrates in semi-natural french streams: an initial application to biomonitoring in Europe. Freshw Biol 43:277–296. https://doi.org/10.1046/J.1365-2427.2000.00545.X
Chevenet F, Doléadec S, Chessel D (1994) A fuzzy coding approach for the analysis of long-term ecological data. Freshw Biol 31(3):295–309. https://doi.org/10.1111/j.1365-2427.1994.tb01742.x
Chin DA (2000) Water- resources engineering (pp. 585–635). New Jersey: Prentice-Hall Inc. Upper Saddle River
Clements WH, Kotalik C (2016) Effects of major ions on natural benthic communities: an experimental assessment of the US Environmental Protection Agency aquatic life benchmark for conductivity. Freshw Sci 35:126–138. https://doi.org/10.1086/685085
Corfield J (2000) The effects of acid sulphate run-off on a subtidal estuarine macrobenthic community in the Richmond River, NSW, Australia. ICES J Mar Sci 57:1517–1523. https://doi.org/10.1006/jmsc.2000.0901
Cormier SM, Suter GW, Zheng L, Pond GJ (2013) Assessing causation of the extirpation of stream macroinvertebrates by a mixture of ions. Environ Toxicol Chem 32:277–87. https://doi.org/10.1002/etc.2059
D’Alessandro M, Porporato EMD, Esposito V et al (2020) Common patterns of functional and biotic indices in response to multiple stressors in marine harbours ecosystems. Environ Pollut 259:113959. https://doi.org/10.1016/j.envpol.2020.113959
de Santiago K, Palmer TA, Wetz MS, Beseres Pollack J (2020) Response of macrobenthic communities to changes in water quality in a subtropical, microtidal estuary (Oso Bay, Texas). Exp Results 1:1–9. https://doi.org/10.1017/exp.2020.44
Díaz S, Cabido M (2001) Vive la différence: plant functional diversity matters to ecosystem processes. Trends Ecol Evol 16:646–655. https://doi.org/10.1016/S0169-5347(01)02283-2
Dong JY, Zhao L, Yang X et al (2021) Functional trait responses of macrobenthos to anthropogenic pressure in three temperate intertidal communities. Front Mar Sci 8:1–15. https://doi.org/10.3389/fmars.2021.756814
Edegbene AO, Arimoro FO, Odume ON (2020) Exploring the distribution patterns of macroinvertebrate signature traits and ecological preferences and their responses to urban and agricultural pollution in selected rivers in the Niger Delta ecoregion, Nigeria. Aquat Ecol 54:553–573. https://doi.org/10.1007/s10452-020-09759-9
Ekau W, Auel H, Pörtner HO, Gilbert D (2010) Impacts of hypoxia on the structure and processes in pelagic communities (zooplankton, macro-invertebrates and fish). Biogeosciences 7:1669–99. https://doi.org/10.5194/bg-7-1669-2010
Ekeroth N, Blomqvist S, Hall POJ (2016) Nutrient fluxes from reduced Baltic Sea sediment: effects of oxygenation and macrobenthos. Mar Ecol Prog Ser 544:77–92. https://doi.org/10.3354/meps11592
El Sayed SM, Hegab MH, Mola HRA et al (2020) An integrated water quality assessment of Damietta and Rosetta branches (Nile River, Egypt) using chemical and biological indices. Environ Monit Assess 192:1–6. https://doi.org/10.1007/s10661-020-8195-4
El Sayed SM, Salem G. Salem, Abdo MH, et al (2022) Spatial distribution of heavy metals and ecological risk assessment for the main sub-branches (Rayahs) sediments of Nile River. Peer Review. https://doi.org/10.21203/rs.3.rs-1247777/v1
El-Damhogy KA, Fishar MR, Mola HR, El-Naggar MM, SA, (2017) The relationship between macrobenthic invertebrates and those associated with plants (Myirophyllum spicatum) in River Nile at Qanater region, Egypt. J Egypt Acad Soc Environ Dev D, Environ Stud 18:21–32. https://doi.org/10.21608/JADES.2017.62640
El-Otify AM, Iskaros IA (2015) Water quality and potamoplankton evaluation of the Nile River in upper Egypt. Acta Limnol Bras 27:171–190. https://doi.org/10.1590/S2179-975X4014
El-Shabrawy GM, Goher ME (2012) Limnology-Limnology of the River Nile. UNESCO, Eolss Publ
Erasmus JH, Lorenz AW, Zimmermann S et al (2021) A diversity and functional approach to evaluate the macroinvertebrate responses to multiple stressors in a small subtropical austral river. Ecol Indic 131:108206. https://doi.org/10.1016/j.ecolind.2021.108206
Ferreira M, Wepener V, van Vuren JHJ (2012) Aquatic invertebrate communities of perennial pans in Mpumalanga, South Africa: a diversity and functional approach. African Invertebr 53:751–768. https://doi.org/10.5733/afin.053.0212
Gaston GR, Rakocinski CF, Brown SS, Cleveland CM (1998) Trophic function in estuaries: response of macrobenthos to natural and contaminant gradients. Mar Freshw Res 49:833–846. https://doi.org/10.1071/MF97089
Gogala A (2009) Micronecta Poweri (Douglas & Scott) In Slovenia (Heteroptera: Corixidae). Acta Entomol Slov 17:79–82
Goher ME (2015) Monitoring of present environmental status of El-Rayahs ,Nile River, Final Report prepared to National Institute of Oceanography and Fisheries, Fresh Water and Lakes Division, Cairo, Egypt. 439 p
Goher ME, Hassan AM, Abdel-Moniem IA et al (2014) Evaluation of surface water quality and heavy metal indices of Ismailia Canal, Nile River, Egypt. J Aquat Res 40:225–233. https://doi.org/10.1016/j.ejar.2014.09.001
Goher ME, Ali MHH, El-Sayed SM (2019) Heavy metals contents in Nasser Lake and the Nile River, Egypt: an overview. Egypt J Aquat Res 45:301–312. https://doi.org/10.1016/j.ejar.2019.12.002
Goher ME, Mangood AH, Mousa IE, Galem SG, Hussein MM (2021) Ecological risk assessment of heavy metal pollution in sediments of Nile River, Egypt. Environ Monit Assess 193(11):703. https://doi.org/10.1007/s10661-021-09459-3
Grzybkowska M, Dukowska M, Michałowicz J, Leszczyńska J (2015) Trace metal concentrations in free-ranger, tube-dweller chironomid larvae and a weakly polluted fluvial sediment. Oceanol Hydrobiol Stud 44:445–455. https://doi.org/10.1515/ohs-2015-0042
Gusmao JB, Brauko KM, Eriksson BK, Lana PC (2016) Functional diversity of macrobenthic assemblages decreases in response to sewage discharges. Ecol Indic 66:65–75. https://doi.org/10.1016/j.ecolind.2016.01.003
Habash ASHH, El-Molla AM, Shaban MSMA, Abdelall MA (2018) Tailor-made protocol for assessing water quality of irrigation canals: case study of El-Nubaria canal, Egypt. Water Sci 32:380–399. https://doi.org/10.1016/j.wsj.2018.08.001
Han JS, Wang JH, Kim JE et al (2017) The effects of thermal discharge on benthic macroinvertebrate communities structure in Buso StreamTitle. Korean J Environ Biol 35:83–94. https://doi.org/10.11626/KJEB.2017.35.1.083
Hu C, Dong J, Gao L et al (2019) Macrobenthos functional trait responses to heavy metal pollution gradients in a temperate lagoon. Environ Pollut 253:1107–1116. https://doi.org/10.1016/j.envpol.2019.06.117
Hussein M, Obuid-Allah A, Mahmoud A, Fangary H (2011) Population dynamics of freshwater snails (Mollusca: Gastropoda) at Qena Governorate, Upper Egypt. Egypt Acad J Biol Sci B Zool 3:11–22. https://doi.org/10.21608/eajbsz.2011.14309
Karlson K, Hulth S, Rosenberg R (2007) Density of Monoporeia affinis and biogeochemistry in Baltic Sea sediments. J Exp Mar Bio Ecol 344:123–135. https://doi.org/10.1016/j.jembe.2006.11.016
Kenny AJ, Jenkins C, Wood D et al (2018) Assessing cumulative human activities, pressures, and impacts on North Sea benthic habitats using a biological traits approach. ICES J Mar Sci 75:1080–1092. https://doi.org/10.1093/icesjms/fsx205
Khalifa N, Bendary RE (2016) Composition and biodiversity of zooplankton and macrobenthic populations in El-Rayah El-Menoufy. Int J Appl Environ Sci ISSN 11:683–700
KouidriDali youcef MN, Benabdellah I et al (2016) Enrichment and geoaccumulation of heavy metals and risk assessment of sediments from coast of Ain Temouchent (Algeria). Arab J Geosci 9:1–9. https://doi.org/10.1007/s12517-016-2377-y
Laini A, Viaroli P, Bolpagni R, Cancellario T, Racchetti E, Guareschi S (2019) Taxonomic and functional responses of benthic macroinvertebrate communities to hydrological and water quality variations in a heavily regulated river. Water (Switzerland) 11:1–18. https://doi.org/10.3390/w11071478
Lamouroux N, Dolédec S, Gayraud S (2004) Biological traits of stream macroinvertebrate communities: effects of microhabitat, reach, and basin filters. J N Am Benthol Soc 23(3):449–466. https://doi.org/10.1899/0887-3593(2004)023%3c0449:BTOSMC%3e2.0.CO;2
Lavorel S, Garnier E (2002) Predicting changes in community composition and ecosystem functioning from plant traits: revisiting the Holy Grail. Funct Ecol 16:545–556. https://doi.org/10.1046/j.1365-2435.2002.00664.x
Lepš J, Petr Šmilauer (2003) Multivariate analysis of ecological data using CANOCO. Cambridge Univ. Press
Leung JYS, Tam NFY (2013) Influence of plantation of an exotic mangrove species, Sonneratia caseolaris (L.) Engl., on macrobenthic infaunal community in Futian Mangrove National Nature Reserve, China. J Exp Mar Bio Ecol 448:1–9. https://doi.org/10.1016/j.jembe.2013.06.006
Lim HS, Diaz RJ, Hong JS, Schaffner LC (2006) Hypoxia and benthic community recovery in Korean coastal waters. Mar Pollut Bull 52:1517–1526. https://doi.org/10.1016/j.marpolbul.2006.05.013
Llanos EN, SarachoBottero MA, Jaubet ML et al (2020) Functional diversity in the intertidal macrobenthic community at sewage-affected shores from Southwestern Atlantic. Mar Pollut Bull 157:111365. https://doi.org/10.1016/j.marpolbul.2020.111365
Lob DW, Silver P (2012) Effects of elevated salinity from road deicers on Chironomus riparius at environmentally realistic springtime temperatures. Freshw Sci 31:1078–1087. https://doi.org/10.1899/12-095.1
Loreau M, Naeem S, Inchausti P et al (2001) Ecology: biodiversity and ecosystem functioning: Current knowledge and future challenges. Science 80-(294):804–808. https://doi.org/10.1126/science.1064088
Mangadze T, Dalu T, William Froneman P (2019) Biological monitoring in southern Africa: a review of the current status, challenges and future prospects. Sci Total Environ 648:1492–1499. https://doi.org/10.1016/j.scitotenv.2018.08.252
McHugh D, Fong PP (2002) Do life history traits account for diversity of polychaete annelids? Invertebr Biol 121:325–338. https://doi.org/10.1111/j.1744-7410.2002.tb00133.x
Mola HRA, Abdel Gawad SS (2014) Spatio-temporal variations of macrobenthic fauna in Lake Nasser khors, Egypt. Egypt J Aquat Res 40:415–423. https://doi.org/10.1016/J.EJAR.2014.12.001
Mouillot D, Graham NAJ, Villéger S et al (2013) A functional approach reveals community responses to disturbances. Trends Ecol Evol 28:167–177. https://doi.org/10.1016/j.tree.2012.10.004
Nasi F, Nordström MC, Bonsdorff E et al (2018) Functional biodiversity of marine soft-sediment polychaetes from two Mediterranean coastal areas in relation to environmental stress. Mar Environ Res 137:121–132. https://doi.org/10.1016/j.marenvres.2018.03.002
Odume ON (2020) Searching for urban pollution signature and sensitive macroinvertebrate traits and ecological preferences in a river in the Eastern Cape of South Africa. Ecol Indic 108:105759. https://doi.org/10.1016/j.ecolind.2019.105759
Orendt C, Wolfram G, Adámek Z et al (2012) The response of macroinvertebrate community taxa and functional groups to pollution along a heavily impacted river in Central Europe (Bílina River, Czech Republic). Biologia (Bratisl) 67:180–199. https://doi.org/10.2478/s11756-011-0158-3
Oug E, Fleddum A, Rygg B, Olsgard F (2012) Biological traits analyses in the study of pollution gradients and ecological functioning of marine soft bottom species assemblages in a fjord ecosystem. J Exp Mar Bio Ecol 432–433:94–105. https://doi.org/10.1016/j.jembe.2012.07.019
Paganelli D, Marchini A, Occhipinti-Ambrogi A (2012) Functional structure of marine benthic assemblages using Biological Traits Analysis (BTA): a study along the Emilia-Romagna coastline (Italy, North-West Adriatic Sea). Estuar Coast Shelf Sci 96:245–256. https://doi.org/10.1016/j.ecss.2011.11.014
Piló D, Ben-Hamadou R, Pereira F et al (2016) How functional traits of estuarine macrobenthic assemblages respond to metal contamination? Ecol Indic 71:645–659. https://doi.org/10.1016/j.ecolind.2016.07.019
Rakocinski CF, Brown SS, Gaston GR et al (2000) Species-abundance-biomass responses by estuarine macrobenthos to sediment chemical contamination. J Aquat Ecosyst Stress Recover 7:201–214. https://doi.org/10.1023/A:1009931721009
Rashid R, Pandit AK (2014) Macroinvertebrates (oligochaetes) as indicators of pollution: a review. J Ecol Nat Environ 6:140–144. https://doi.org/10.5897/jene2014.0443
Rawer-Jost C, Böhmer J, Blank J, Rahmann H (2000) Macroinvertebrate functional feeding group methods in ecological assessment. Hydrobiologia 422–423:225–232. https://doi.org/10.1007/978-94-011-4164-2_18
Rosenberg R (2001) Marine benthic faunal sucessional stages and related sedimentary activity. Sci Mar 65:107–119. https://doi.org/10.3989/scimar.2001.65s2107
Ryu J, Khim JS, Kang SG et al (2011) The impact of heavy metal pollution gradients in sediments on benthic macrofauna at population and community levels. Environ Pollut 159:2622–2629. https://doi.org/10.1016/j.envpol.2011.05.034
SaadAbd El-Halim A, Emam WM, Mola HRA, Omar HM (2015) Effect of pollution on macrobenthic invertebrates in some localities along the River Nile at Great Cairo, Egypt. Aquat Biol Fish 19:1–11
Serra SRQ, Graça MAS, Dolédec S, Feio MJ (2017) Chironomidae traits and life history strategies as indicators of anthropogenic disturbance. Environ Monit Assess 189:326. https://doi.org/10.1007/s10661-017-6027-y
Shyleshchandran MN, Mohan M, Ramasamy EV (2018) Risk assessment of heavy metals in Vembanad Lake sediments (south-westcoast of India), based on acid-volatile sulfide (AVS)-simultaneously extracted metal (SEM) approach. Environ Sci Pollut Res 25(8):7333–7345. https://doi.org/10.1007/s11356-017-0997-8
Talab AS, Goher ME, Ghannam HE, Abdo MH (2016) Chemical compositions and heavy metal contents of Oreochromis niloticus from the main irrigated canals (rayahs) of Nile Delta. Egypt J Aquat Res 42:23–31. https://doi.org/10.1016/j.ejar.2016.01.003
Ter Braak CJF, Smilauer P (2002) Canoco 4.5 software for canonical community ordi_nation. Microcomputer Power, Ithaca NY, US
Thioulouse J, Chessel D, Dolédec S, Olivier JM (1997) ADE-4: a multivariate analysis and graphical display software. Stat Comput 7:75–83. https://doi.org/10.1023/A:1018513530268
Törnroos A, Bonsdorff E (2012) Developing the multitrait concept for functional diversity: lessons from a system rich in functions but poor in species. Ecol Appl 22:2221–2236. https://doi.org/10.1890/11-2042.1
Tripole S, Gonzalez P, Vallania A et al (2006) Evaluation of the impact of acid mine drainage on the chemistry and the macrobenthos in the Carolina stream (San Luis-Argentina). Environ Monit Assess 114:377–389. https://doi.org/10.1007/s10661-006-4941-5
Tselepides A, Papadopoulou KN, Podaras D et al (2000) Macrobenthic community structure over the continental margin of Crete (South Aegean Sea, NE Mediterranean). Prog Oceanogr 46:401–428. https://doi.org/10.1016/S0079-6611(00)00027-6
Usseglio-Polatera P, Bournaud M, Richoux P, Tachet H (2000) Biomonitoring through biological traits of benthic macroinvertebrates: how to use species trait databases? Hydrobiologia 422–423:153–162. https://doi.org/10.1007/978-94-011-4164-2_12
Villéger S, Miranda JR, Hernández DF, Mouillot D (2010) Contrasting changes in taxonomie vs. functional diversity of tropical fish communities after habitat degradation. Ecol Appl 20:1512–1522. https://doi.org/10.1890/09-1310.1
Voß K, Schäfer RB (2017) Taxonomic and functional diversity of stream invertebrates along an environmental stress gradient. Ecol Indic 81:235–242. https://doi.org/10.1016/j.ecolind.2017.05.072
Wang B, Liu CQ, Wang F et al (2013) Diatoms modify the relationship between dissolved silicon and bicarbonate in impounded rivers. J Limnol 72:494–504. https://doi.org/10.4081/jlimnol.2013.e40
Williams DD, Williams NE, Cao Y (1999) Road salt contamination of groundwater in a major metropolitan area and development of a biological index to mon. Related papers Ecology and product ion of invert ebrat es in a Canadian coldwat er spring-springbrook syst em ROAD SALT CONTAMINATION OF GR
Worthington TA, Shaw PJ, Daffern JR, Langford TEL (2015) The effects of a thermal discharge on the macroinvertebrate community of a large British river: implications for climate change. Hydrobiologia 753:81–95. https://doi.org/10.1007/s10750-015-2197-1
Yap CK, Rahim AI, Azrina MZ, Ismail A, Tan SG (2006) The influential of physico-chemical parameters on the distributions of oligochateas (Limnodrilus sp.) at the polluted downstream of the tropical Langat River, Peninsular Malaysia. J Appl Sci Environ Manag 10:135–140. https://doi.org/10.4314/jasem.v10i3.17332
Zhang Q, Yang T, Wan X et al (2021) Community characteristics of benthic macroinvertebrates and identification of environmental driving factors in rivers in semi-arid areas – a case study of Wei River Basin, China. Ecol Indic 121:107153. https://doi.org/10.1016/j.ecolind.2020.107153
Funding
Open access funding provided by The Science, Technology & Innovation Funding Authority (STDF) in cooperation with The Egyptian Knowledge Bank (EKB).
Author information
Authors and Affiliations
Contributions
- Designed the field study and achieved the fieldwork: M.Goher, G.El Shabrawy
- Identification of macrobenthic species and laboratory work: R. Bendary, Sh.Ibrahim, M. Goher, M. Khalil
- Performed the statistical analysis of the data: R. Bendary
- Wrote the manuscript: R. Bendary, M.Goher
- Review and editing: all authors
Corresponding author
Ethics declarations
Ethics approval
Not applicable.
Consent to participate
All authors voluntarily agree to participate in this research study.
Consent to publish
All authors voluntarily approved the publication of this research study.
Competing interest
The authors declare no competing interests.
Additional information
Responsible Editor: Thomas Hein
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Bendary, R.E., Ibrahim, S.M., Goher, M.E. et al. Taxonomic and functional structure of macrobenthic invertebrate communities and their response to environmental variables along the subbranches of the Nile River (rayahs), Egypt. Environ Sci Pollut Res 30, 28803–28817 (2023). https://doi.org/10.1007/s11356-022-24140-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-022-24140-z