
Abstract
The parallel batches of the same species and geographical origin mushrooms both raw and stir-fried were investigated to get an insight into the content and intake of 137Cs, 40K, and K from mushroom meals. The Boletaceae family species (Baorangia bicolor, Boletus bainiugan, Butyriboletus roseoflavus, Retiboletus griseus, Rugiboletus extremiorientalis, and Sutorius magnificus) were collected from the Midu County (Dali Bai Autonomous Prefecture) in 2018. The activity concentrations of 137Cs in the caps of dried raw mushrooms were in the range 14 ± 1 Bq kg−1 dry biomass (db) (R. griseus) to 34 ± 2 Bq kg−1 db (R. extremiorientalis), and in stems from 16 ± 1 Bq kg−1 db (B. bicolor and B. bainiugan) to 23 ± 1 Bq kg-1 db (R. extremiorientalis). The mean activity concentration in the whole fruiting bodies in all six species was 18 ± 4 Bq kg-1 db. The activity concentrations of 137Cs were roughly the same in both dehydrated materials, stir-fried, and raw mushrooms, while the contents of 40K and stable K were around 2- to 3-fold smaller in stir-fried than raw product. The raw and stir-fried mushrooms on a whole (wet) weight basis showed activity concentrations of 137Cs in the range from 1.2 to 3.2 Bq kg−1 ww (mean 1.9 ± 0.6 Bq kg−1 ww) and 6.0 to 9.4 Bq kg−1 ww (mean 7.0 ± 1.2 Bq kg−1 ww), respectively. Evidently, when expressed on a whole (wet) weight basis, the cooked mushrooms showed on average around 3.5-fold greater activity concentration of 137Cs when compared with raw mushrooms. The 137Cs, 40K, and total K enrichment in stir-frying (in a whole (wet) weight basis for the meal), confronted with the results for dehydrated raw and fried mushrooms, show the direct correlation with loss of mass (largely moisture) during the cooking procedure but not much of 137Cs and 40K. Edible wild mushrooms from Yunnan were little contaminated with radiocaesium. As assessed, the mean radioactivity dose from natural 40K in around 9.3-fold exceeded the dose obtained for artificial 137Cs from stir-fried mushroom meals, which both were very low doses.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Mushrooms that grow in the wild are known to be sensitive to contamination with radiocaesium (134Cs/137Cs) that originates from atmospheric fallout. Historically, the accident at the Chernobyl nuclear power plant in 1986 caused high and long-lasting pollution over significant parts of continental Europe. This included contamination of forest soils with 137Cs in many regions, and consequently also of the wild mushrooms that grow in these forests (Betti et al. 2017; Chiaravalle et al. 2018; Falandysz et al. 2015; Grodzynska 2018; Orita et al. 2017; Travnikova et al. 2002; Tucaković et al. 2018). More recently, a failure of the Fukushima Dai-ichi nuclear power plant in also caused pollution with 137Cs of mushrooms in a region of Japan (Prand-Stritzko and Steinhauser 2018; Steinhauser et al. 2014). There was negligible impact of this incident on continental Asia, e.g., in the region of southwestern China (Falandysz et al. 2016, 2017, 2018). Soil in Yunnan is considered little polluted with 137Cs, and forest topsoil (0–5 cm layer) sampled from the Changning localization (ca. 200 km west of Midu County) in Yunnan in 2016 showed the 137Cs activity concentration at level 4.9 ± 0.6 Bq kg−1 dry weight (Falandysz et al. 2018).
The mycelial network of fungi rapidly absorbs radiocesium that is available in litter and soil and is able to translocate it to the emerging fruiting bodies soon after deposition of radioactive fallout (Stijve and Poretti 1990). The bulk of 137Cs deposited in forest soil is a result of the low radio decay rate (half-life time is 30.17 years) and slow migration down to soil depths that are relevant for incorporation in fungal biological cycles (Tanaka et al. 2018), but the resulting contamination persists for decades. A fraction of the freshly deposited radiocaesium that is not absorbed by the uppermost layers on the forest floor can infiltrate at a faster rate down to lower soil layers (horizon) via pores in the soil structure (Fuji et al. 2014). Thus, it can also be available relatively rapidly for species whose mycelia extend to greater depths in the soil, e.g., the mycelium in some species can extend to depths of 0.5 m (Ingrao et al. 1992).
The soil layer where a fungus has its highest density of mycelium and the extent of depth and space to which the hyphae penetrate within layers depends largely on the species. For ectomycorrhizal fungi, the soil distribution of taxa is likely species-dependent, and if the soil profile is favorable, the hyphae follow the roots of their symbiotic partners to lower depths. For example, the ectomycorrhizal Amanita strobiliformis had the highest density in the 6–12 cm layer, and for the saprobic Agaricus bernardii, hyphae live at least down to a depth of 30 cm (Borovička et al. 2014). The long-term retention of 137Cs in forest topsoil results in prolonged contamination of mushrooms with this nuclide (Falandysz and Borovička 2013).
Susceptibility to nuclide uptake/bio-concentration typically occurs over a wider range of fungal species (Cocchi et al. 2017; Falandysz et al. 2019d; Mietelski et al. 2010; Zalewska et al. 2016) and fruiting bodies both of the same or several species collected in the same season from the same forested area, usually shows a range of different 137Cs activity concentrations. The variability in uptake/bio-concentration efficiency of 134Cs/137Cs between species and its activity concentration in fungal fruiting bodies is also a function of the stable caesium (133Cs) status of the mushroom (Yoshida et al. 2000), but this important aspect has been little studied so far. Additionally, within species, the developmental stage (size) of the fruiting body (e.g., for Amanita muscaria) can also influence the activity concentration (Falandysz et al. 2019b).
Many regions of the world are habitats to mushrooms that can be foraged in the wild including the Yunnan province of China, which has high biodiversity with nearly 900 edible species (Wu et al. 2010). Twenty years before the nuclear accident at Chernobyl, mushrooms were already recognized as a possible source of 137Cs contamination (coming from the nuclear weapon explosions) for humans (Kiefer and Maushart 1965). However it has also been observed that intake through this pathway could be reduced by cooking processes, e.g., blanching or boiling with excess of water, can decrease the mushroom content of artificial 137Cs and the alkali metals, e.g., Li and Cs, which leak into the discarded water phase (Consiglio et al. 1990; Daillant et al. 2013; Pankavec et al. 2019; Skibniewska and Smoczyński 1999; Stijve 1994).
In a number of Asian cuisines, stir-frying of foods including mushrooms, in a wok type pan, is a popular method of cooking. Information on the possible effect of stir-frying mushrooms in a wok, on the 137Cs, 40Potassium (40K), and total Potassium (K) content and their intake from such stir-fried mushrooms meals, are not available in the literature. We investigated the potential effect of stir-frying in a moderate volume of oil using traditional Chinese cooking practice, on the content of 137Cs, 40Potassium (40K), and total Potassium (K) in dishes made of six species of mushrooms widely foraged in Yunnan.
Materials and methods
Fungal materials
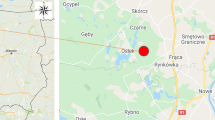
Mushrooms were collected from the geochemically anomalous region from the Midu County (Dali Bai Autonomous Prefecture) located in the west-central region of Yunnan province in China in 2018 (Fig. 1). Species such as Baorangia bicolor (Kuntze) G. Wu, Halling & Zhu L. Yang (earlier called Boletus bicolor Peck) and the sample whole (fresh) weight was 433 g (n = 21) for uncooked and 402 g (n = 22) for stir-fried fruiting bodies; Boletus bainiugan Dentinger (earlier called in SW China Boletus edulis Bull.) weighing 280 g (n = 5) and 292 g (n = 6); Butyriboletus roseoflavus (earlier called Boletus speciosus Forst.) weighing 299 g (n = 5) and 342 g (n = 5); Retiboletus griseus (Frost) Manfr. Binder & Bresinsky (earlier called Boletus griseus Frost) weighing 366 g (n = 8) and 341 g (n = 8); Rugiboletus extremiorientalis (Lj.N. Vassiljeva) G. Wu & Zhu L. Yang (earlier called Leccinum extremiorientale (Lj.N. Vassiljeva)) weighing 440 g (n = 12) and 312 g (n = 12), and Sutorius magnificus (W.F. Chiu) G. Wu & Zhu L. Yang (earlier called Boletus magnificus W.F. Chiu) weighing 154 g (n = 6) and 312 g (n = 7) (Cui et al. 2016; Feng et al. 2012) were sampled.

Map of the area from the Dali Autonomous Prefecture in Yunnan Province where mushrooms were collected; 25° 20′ 34″ N 100° 29′ 35″ E (Google maps)
Cooking
Fruiting bodies that were randomly selected for stir-frying pools were collected at the same time and location as the raw counterpart pools. Each collected fruiting body within a pool was individually cleaned from foreign debris, and the morphological parts (cap and stem) were separated. All specimens not subjected for stir-frying were sliced and dried for 24 h to a constant mass at 65 °C in a food dehydrator (Ultra FD1000, Ezidri, Australia), and ground to a fine powder using a clean porcelain mortar and pestle and stored dry in sealed polyethylene bags until analysis.
All specimens subjected for stir-frying were sliced, separately caps and stems, pooled accordingly and stir-fried using one of the traditional Yunnan methods in moderate amount of hot vegetable oil (15 to 55 g and depending on the sample size) in a wok pan for 10 min. After cooking, the excess oil was drained away and the fried mushrooms were cooled and transferred into fresh polyethylene jars (screw capped, 0.5 L), weighted, deep frozen (− 20 °C), freeze-dried for 72 h, then reweighed (to calculate moisture content), homogenized using a blender with steel-less blades and plastic bowl, and kept tightly closed in screw capped polycarbonate jars which were packed individually into sealed polyethylene bags and stored in a refrigerator. Immediately prior to the instrumental analysis, all the fungal materials were deep frozen (− 20 °C) and freeze-dried for 72 h (Labconco Freeze Dry System, Kansas City, MO, USA), so that the activity levels of nuclides were determined in fully dehydrated materials.
Analysis
Activity concentrations of 137Cs and 40K were determined using a gamma spectrometer with a coaxial HPGe detector and with a relative efficiency of 18% and a resolution of 1.9 keV at 1.332 MeV (with associated electronics). Quantitation was carried out using the equation:
where Ni is the number of counts after background correction, ε(E) is the detector efficiency for photons with energy E, y is the emission probability, and t is the measurement time in seconds.
All measurements were preceded by a background measurement (time 80,000 s or 250,000 s), and background counts were subtracted (using the GENIE 2000 program). The lower limit of detection was at 0.10 Bq kg−1 dry biomass (db). The equipment was calibrated using a multi-isotope standard, and the method was fully validated (Falandysz et al. 2019d; Wang et al. 2015). The reference material “Standard solution of gamma emitting isotopes, code BW/Z-62/27/07” produced at the IBJ-Świerk near Otwock in Poland was used for preparing reference samples for equipment calibration. The radionuclides used in the multicomponent reference solution during equipment calibration were 137Cs at 1.5% and several other nuclides with an approximation uncertainty level at 0.80-2.1%.
The same geometry of cylindrical dishes with 40 mm diameter (as used for the measurement of collected samples) was used for reference samples during equipment calibration. Calibration was carried out using standards with a density of approximately 1 g cm3(liquid) with different heights: 3, 6, 9, 15, and 25 mm, which allows the selection of the appropriate calibration for samples of different thickness layer. All numerical data obtained were recalculated for dehydrated fungal material (freeze-dried), and all data were decay-corrected back to the time of sampling. Concentrations of total (stable) K (Table 2) were calculated from the 40K activity concentration (mean value) in natural K, which is in the range 27.33 to 31.31 Bq g−1 of K (Samat et al. 1997).
Results and discussion
137Cs, 40K, and K in dried raw mushrooms
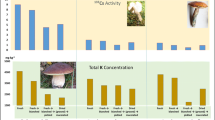
The activity concentrations of 137Cs in pooled samples of caps of the species were in the range 14 ± 1 Bq kg−1 db in Retiboletus griseus to 34 ± 2 Bq kg−1 db in Rugiboletus extremiorientalis. Values of 137Cs in the pooled samples of stems were from 16 ± 1 Bq kg−1 db in Baorangia bicolor and Boletus bainiugan to 23 ± 1 Bq kg−1 db in Rugiboletus extremiorientalis. The mean activity concentration in the whole fruiting bodies in all six species was 18 ± 4 Bq kg−1 db (Table 1).
The data obtained for 137Cs confirm that wild mushrooms foraged from the Yunnan province in China show low contamination levels, and the activity concentrations were within the range of values reported so far both in wild and cultivated mushrooms from southwestern Asia (Falandysz et al. 2015, 2016, 2017. 2018; Tuo et al. 2014; Wang et al. 2015). However, 137Cs at a somewhat greater activity concentration, i.e., 210 Bq kg−1 db (range 150 ± 9 to 340 ± 22 Bq kg−1 db) has been found in the shaggy (scaly or woolly) chanterelle Gomphus floccosus (Schv.) Sing in one study (Tuo et al. 2014), current name Turbinellus floccosus (Schwein.) Earle ex Giachini and Castellano (Index Fungorum 2020). Also, mushrooms from the subalpine region of the Minya Konka (Mount Gongga or Gongga Shan) in the Eastern Tibetan Plateau, showed, on average, higher levels of 137Cs than the present study, i.e., in the range 62 ± 6 to 280 ± 150 Bq kg−1 db in caps and 62 ± 6 to 400 ± 72 Bq kg−1 db in stems (Falandysz et al. 2018).
40K in caps was in the range 890 ± 96 Bq kg−1 db (Baorangia bicolor) to 1100 ± 93 Bq kg−1 db (Retiboletus griseus) and in stems 500 ± 110 Bq kg−1 db (Sutorius magnificus) to 1000 ± 110 Bq kg−1 db (Retiboletus griseus). The whole fruiting bodies showed activity concentrations of 40K in the range from 740 Bq kg−1 db (Baorangia bicolor) to 1100 Bq kg−1 db (Retiboletus griseus), and the mean value for all six species was 840 ± 120 Bq kg−1 db. The distribution of 137Cs and 40K between morphological parts of the fruiting bodies (QuotientCap/Stem) was similar (QC/S ~ 1) in Boletus bainiugan, Rugiboletus extremiorientalis, and Retiboletus griseus, but favored the caps (mean QC/S 1.6) in Baorangia bicolor, Butyriboletus roseoflavus, and Sutorius magnificus. The overall mean distribution ratios for all the fruiting bodies sampled were QC/S 1.1 ± 0.2 for 137Cs and QC/S 1.5 ± 0.3 for 40K.
The total K content determined was in the range of 32,000 ± 3400 mg kg−1 db (Baorangia bicolor) to 40,000 ± 3300 mg kg−1 db (Retiboletus griseus) in the caps, and 19,000 ± 3000 mg kg−1 db (Baorangia bicolor and Sutorius magnificus) to 37,000 ± 3800 mg kg−1 db (Retiboletus griseus) in the stems. Soil mushrooms, including species of the family Boletaceae, are rich in K. In the present study, the total K contents in the whole fruiting bodies were in the range 26,000 ± 3200 mg kg−1 db (Baorangia bicolor) to 39,000 ± 3500 (Retiboletus griseus) mg kg−1 db, and the overall mean was 30,000 ± 4400 mg kg−1 db.
Data obtained on K in species in the present study agree with the results obtained for B. edulis that grows in European forests, which contained the element in the range 25,000 ± 4000 to 29,000 ± 3000 mg kg−1 db (caps) and 16,000 ± 4000 to 20,000 ± 3000 mg kg−1 db (stems) (Falandysz et al. 2011; Frankowska et al. 2010; Zhang et al. 2010). The small fluctuation in the concentration range of K in species such as Boletus edulis from a range of the geographical regions is relatively narrow. It is thought to be a result of the essentiality and homeostatic regulation of this major metallic element absorbed from soil by mushrooms (Stijve 1996).
137Cs, 40K, and K in stir-fried mushrooms
The activity concentrations of 137Cs were roughly the same in both dehydrated samples, stir-fried and raw mushrooms, while the contents of 40K and stable K were around 2- to 3-fold smaller in stir-fried than raw produce (Table 1). In detail, the overall mean of the activity concentration of 137Cs in the stir-fried mushrooms (whole fruiting bodies) for six species was 16 ± 4 Bq kg−1 db (range 11 ± 2 to 22 ± 2 Bq kg−1 db), and of 40K, was 320 ± 92 Bq kg−1 db (range 190 ± 80 to 480 ± 110 Bq kg−1 db). The overall mean content of K in stir-fried mushrooms was 11,000 ± 4600 mg kg−1 db, and the range for all species was 7000 ± 2800 to 18,000 ± 3600 mg kg−1 db (Table 1). Clearly, the content of 40K and K in the stir-fried mushrooms when normalized to dry biomass basis was far less than in dehydrated raw mushrooms (humidity in the range from 88.2% in Rugiboletus extremiorientalis to 91.7% in Retiboletus griseus) (Falandysz et al. 2019c). The vegetable oil used in stir-frying of mushrooms showed activity concentration of 137Cs in 0.24 ± 0.05 Bq per g, and was free of K.
Data obtained (Table 1) shows higher retention of 137Cs than of 40K (and stable K) in the course of stir-frying. This may be due to the difference in the distribution of Cs and K in cell structures and their binding sites. Potassium is a major monovalent element that is found in high concentrations in mushrooms and is a key component of the cellular protoplasm. Breakout from cell walls due to the effects of high-temperature cell shrinkage during the course of stir-drying can favor the release of K, but can have a lower effect on Cs. Also a preferable leaching of K from cells into the residual oily phase is possible. Stir-frying causes partial dehydration of cooked foodstuffs and resulting in an increase in the proportion of dry matter in a mushroom meal but also of some less volatile or non-volatile compounds of the metallic elements, e.g., mercury (Falandysz et al. 2019a, c). The volume or proportion of oil per unit of stir-fried mushrooms depends on the cooking practice or on local tradition in the different regions of Yunnan. In the present study, fried mushrooms lost from 36 to 55% of the original humidity and absorbed almost all the entire volume of the oil added (from 24 to 32 g of oil per 100 g) to raw (fresh) mushrooms, leaving a small residual volume (0.82 to 5.7 g per dish) (Falandysz et al. 2019c).
Data from the literature shows that when frying (flat pan) mushrooms Cantharellus cibarius and Boletus edulis, a substantial portion of 137Cs leaked out into the oily residue, which could be discarded, but the possible intake of 137Cs with fried mushroom meals has not been estimated (Steinhauser and Steinhauser 2016). Wetting or wet-cleaning of Cantharellus cibarius and Boletus edulis from foreign debris before frying could increase the loss of 137Cs into the oily residue. The rate of leaching of 137Cs in washed chanterelles was 19 to 29 %, and from 2 to 16 % from mushrooms that were cleaned dry. The corresponding rates for Boletus edulis were 6 % and 0 % respectively (Steinhauser and Steinhauser 2016).
In an earlier study, Kenigsberg et al. (1996) reported that frying of mushrooms decreased the activity concentration of 137Cs by 70 % (probably when calculated on a dry biomass basis in relation to the activity concentration in dehydrated uncooked mushrooms) but without information on the mushroom species studied or on the intake of the nuclide based on the whole weight of the fried mushroom meal.
137Cs, 40K, and K in mushroom meals, possible intake, and exposure to radioactivity
All of the species surveyed in this study were edible, both stems and caps of the fruiting bodies. Thus, data on 137Cs, 40K, and K in raw mushrooms and mushroom meals were recalculated for appropriate meal portions for stems and caps, and are expressed for the whole fruiting bodies on a whole weight basis (Table 2).
The raw mushrooms and stir-fried mushroom meals on a whole weight showed activity concentrations of 137Cs respectively in the range from 1.2 to 3.2 Bq kg−1 ww (mean 1.9 ± 0.6 Bq kg−1 ww) and from 6.0 to 9.4 Bq kg−1 ww (mean 7.0 ± 1.2 Bq kg−1 ww). Evidently, when expressed on a whole (produce) weight basis, the stir-fried meals showed on average around 3.5-fold greater activity concentration of 137Cs when compared with raw mushrooms.
In the case of 40K, the activity concentration in raw mushrooms was in the range of 82 to 91 Bq kg−1 ww (mean 87 ± 4 Bq kg−1 ww), and in the stir-fried mushroom meals, in the range of 100 to 200 Bq kg−1 ww (mean 140 ± 32 Bq kg−1 ww). Analogically, the content of stable K in fresh mushrooms was in the range from 2700 to 3200 mg kg−1 ww (mean 3100 ± 200 mg kg−1 ww) and in cooked produce, from 3800 to 7700 mg kg−1 ww (mean 5000 ± 1300 mg kg−1 ww).
Potassium (both 40K and total K) were slightly enriched in the whole weight stir-fried mushroom meals when compared with the whole weight raw mushrooms (Table 2), and this outcome was very different when compared with data expressed on dry biomass basis (Table 1). Thus, it can be expected that in the course of stir-frying, a large portion of potassium escaped out of the mushroom meals, while for 137Cs, the rate of loss was lower.
It has been estimated from a previous report (Falandysz et al. 2019c) that during the mushroom foraging season in southwestern China, certain elements of the population can consume a 100-g portion of stir-fried mushrooms daily, per capita, over a period of several weeks. On this basis, the estimated average intake of 137Cs, 40K, and total K with a single meal per capita would be 0.70 ± 0.12 Bq, 14 ± 3 Bq, and 500 ± 130 mg, and 4.9 ± 0.9 Bq, 96 ± 23 Bq, and 3500 ± 910 mg respectively for weekly intake. The estimated daily intake of 137Cs, 40K, and total K normalized to per kilogram of body mass (bm; typical weight of 60 kg per individual) was 12 ± 2 mBq, 230 ± 52 mBq, and 8.3 ± 2.2 mg respectively, and the corresponding weekly intake was 82 ± 14 mBq, 1600 ± 360 mBq, and 58 ± 15 mg respectively (Table 3).
The average effective daily and weekly doses of gamma radioactivity received per capita from 137Cs and 40K contained in stir-fried mushroom meals in this study have been estimated as 0.0091 ± 0.015 μSv and 0.064 ± 0.011 μSv for 137Cs and 0.085 ± 0.019 μSv and 0.60 ± 0.14 μSv for 40K (Table 3). The average daily and weekly effective doses received by consumers from 137Cs and 40K expressed per kilogram of body mass have been assessed as 0.00015 ± 0.00003 μSv and 0.0011 ± 0.0007 μSv for 137Cs and 0.0014 ± 0.0004 μSv and 0.010 ± 0.002 μSv for 40K (Table 3). Therefore, on a single meal or annual basis, a mean radioactivity dose from natural 40K (0.51 μSv per kilogram of body mass) in around 9.3-fold exceeded the dose obtained for artificial 137Cs (0.055 μSv), which both are very low.
Adequate potassium daily intake for adults has been set for 4700 mg (NIH 2019), and a serving composed of 100 g stir-fried (whole weight) mushroom meals containing between 380 and 770 mg of K (mean 500 ± 130 mg; Table 2) and assuming that absorption rate by body is 85 to 90% classifies the mushroom meals in this study among a good potential dietary source of potassium. Mushrooms that are conserved are much poorer source of potassium (Pankavec et al. 2019) than the stir-fried products.
Conclusions
Examination of the parallel batches of the same species and geographical origin both uncooked and stir-fried mushrooms showed that the activity concentrations of 137Cs were roughly the same in both dehydrated samples, stir-fried, and raw mushrooms, while the contents of 40K and stable K were around 2- to 3-fold smaller in stir-fried than raw produce. The stir-fried mushroom meals on a whole (wet) weight exhibited on average around 3.5-fold greater activity concentration of 137Cs when compared with raw mushrooms. The 137Cs enrichment in cooking, and less of 40K (in a whole (wet) weight basis for the meal), confronted with the results for dried raw and fried mushrooms show the direct correlation with loss of mass (largely moisture during the cooking procedure) but not much of 137Cs and 40K; therefore, activity concentrations of both nuclides in stir-fried mushrooms increase. Hence, exposure to radiocaesium from meals made of the stir-fried mushrooms substantially contaminated with this nuclide can be a more significant source than speculated earlier. However, edible wild mushrooms from Yunnan are little contaminated with radiocaesium. An assessed, the mean radioactivity dose from natural 40K in around 9.3-fold exceeded the dose obtained for artificial 137Cs from stir-fried mushroom meals in Yunnan, which both were very low doses. The effect of the stir-frying process on 137Cs and other mineral constituent content in mushroom meals has to be clarified from the ongoing studies.
References
Betti L, Palego L, Lucacchini A, Giannaccini G (2017) 137Caesium in samples of wild-grown Boletus edulis Bull. from Lucca province (Tuscany, Italy) and other Italian and European geographical areas. Food Addit Contam A 34:49–55
Borovička J, Mihaljevič M, Gryndler M, Kubrová J, Žigová A, Hršelová H, Řanda Z (2014) Lead isotopic signatures of saprotrophic macrofungi of various origins: tracing for lead sources and possible applications in geomycology. Appl Geochem 43:114–120
Chiaravalle AE, Mangiacotti M, Marchesani G, Bortone N, Tomaiuolo M, Trotta G (2018) A ten-year survey of radiocontamination of edible Balkan mushrooms: Cs-137 activity levels and assessed dose to the population. Food Control 94:263–267
Cocchi L, Kluza K, Zalewska T, Apanel A, Falandysz J (2017) Radioactive caesium (134Cs and 137Cs) in mushrooms of the genus Boletus from the Reggio Emilia in Italy and Pomerania in Poland. Isot Environ Health Stud 53:620–627
Consiglio G, Gattavecchia E, Tonelli D, Cocchi L (1990) La radioattività nei funghi: attualità del problema ed opportunità di un approfondimento. Riv Micol 33:227–231
Cui Y, Feng B, Wu G, Xu J, Yang Z (2016) Porcini mushrooms (Boletus sect. Boletus) from China. Fungal Divers. https://doi.org/10.1007/s13225-015-0336-7
Daillant O, Boilley D, Josset M, Hettwig B, Fischer HW (2013) Evaluation of radiocaesium contamination in mushrooms and influence of treatment after collection. J Radioanal Nucl Chem 297:437–441
Falandysz J, Borovička J (2013) Macro and trace mineral constituents and radionuclides in mushrooms: health benefits and risks. Appl Microbiol Biotechnol 97:477–501
Falandysz J, Frankowska A, Jarzyńska G, Dryżałowska A, Kojta AK, Zhang D (2011) Survey on composition and bioconcentration potential of 12 metallic elements in King Bolete (Boletus edulis) mushroom that emerged at 11 spatially distant sites. J Environ Sci Health B 46:231–246
Falandysz J, Zalewska T, Krasińska G, Apanel A, Wang Y, Pankavec S (2015) Evaluation of the radioactive contamination in Fungi genus Boletus in the region of Europe and Yunnan Province in China. Appl Microbiol Biotechnol 99:8217–8224
Falandysz J, Zalewska T, Apanel A, Drewnowska N, Kluza K (2016) Evaluation of the activity concentrations of 137Cs and 40K in some Chanterelle mushrooms from Poland and China. Environ Sci Pollut Res 23:20039–20048
Falandysz J, Zhang J, Zalewska T (2017) Radioactive artificial 137Cs and natural 40K activity in 21 edible mushrooms of the genus Boletus species from SW China. Environ Sci Pollut Res 24:8189–8199
Falandysz J, Saniewski M, Zhang J, Zalewska T, Liu H, Kluza K (2018) Artificial 137Cs and natural 40K in mushrooms from the subalpine region of the Minya Konka summit and Yunnan Province in China. Environ Sci Pollut Res 25:615–627
Falandysz J, Dryżałowska A, Zhang J, Wang Y (2019a) Mercury in raw mushrooms and mushrooms stir-fried in deep oil. J Food Compos Anal 82:103239
Falandysz J, Saniewski M, Zalewska T, Zhang J (2019b) Pollution by radiocaesium of fly agaric Amanita muscaria in fruiting bodies decrease with a developmental stage. Isot Environ Health Stud 55:317–324
Falandysz J, Zhang J, Mędyk M, Zhang J (2019c) Mercury in stir-fried and raw mushrooms from the Boeltaceae family from the geochemically anomalous region in the Midu county, China. Food Control 102:17–21
Falandysz J, Zalewska T, Fernandes A (2019d) 137Cs and 40K in Cortinarius caperatus mushrooms (1996 – 2016) in Poland - bioconcentration and estimated intake: 137Cs in Cortinarius spp. from the Northern Hemisphere from 1974 – 2016. Environ Pollut 255:113208
Feng B, Xu J, Wu G, Zeng NK, Li YC, Tolgor B, Kost GW, Yang ZL (2012) DNA sequence analyses reveal abundant diversity, endemism and evidence for Asian origin of the porcini mushrooms. PLoS ONE 7(3):e37567
Frankowska A, Ziółkowska J, Bielawski L, Falandysz J (2010) Profile and bioconcentration of minerals by King Bolete (Boletus edulis) from the Płocka Dale in Poland. Food Addit Contam B 3:3–6
Fuji K, Ikeda S, Akama A, Komatsu M, Takahashi M, Kaneko S (2014) Vertical migration of radiocesium and clay mineral composition in five forest soils contaminated by the Fukushima nuclear accident. Soil Sci Plant Nutr 60:751–764
Grodzynska GA (2018, 2017) Radionuclide contamination macromycetes (in Russian). Visn Nac Acad Nauk Ukr (6):61–76. https://doi.org/10.15407/visn2017.06.061
Index Fungorum (2020) http://www.indexfungorum.org/Names/Names.asp
Ingrao G, Belloni P, Santaroni GP (1992) Mushrooms as biological monitors of trace elements in the environment. Journal of Radioanalytical and Nuclear Chemistry Articles 161(1):113–120
Kenigsberg J, Belli M, Tikhomirov F, Buglovaa E, Shevchuk V, Renaudd P, Maubertd H, Bruke G, Shutov V (1996) Exposures from consumption of forest produce. In: Karaoglou A, Desmet G, Kelly GN, Menzel HG (eds) The radiological consequences of the Chernobyl accident. European Commission and the Belarus, Russian and Ukrainian ministers on Chernobyl Affairs, Emergency Situations and Health. EUR 16544 EN, Brussel, pp 271–181
Kiefer H, Maushart R (1965) Erhöhter Cs-137-Gehalt im menschlichen Körper nach Polzegenuß. Direct Informantion, Atompraxis, p 15
Mietelski JW, Dubchak S, Błażej S, Anielska T, Turnau K (2010) 137Cs and 40K in fruiting bodies of different fungal species collected in a single forest in southern Poland. J Environ Radioact 101:706–711
NIH (2019) National Institute of Health; https://ods.od.nih.gov/factsheets/Potassium-HealthProfessional/, retrieved on January 30, 2019
Orita M, Kimura Y, Tiara Y, Fukuda T, Takahashi J, Gutevych O, Chornyi S, Kudo T, Yamashita S, Takamura N (2017) Activities concentration of radiocesium in wild mushroom collected in Ukraine 30 years after the Chernobyl power plant accident. PeerJ 6:e4222. https://doi.org/10.7717/peerj.4222
Pankavec S, Hanć A, Barałkiewicz D, Dryżałowska A, Zhang J, Falandysz J (2019) Mineral constituents of conserved white button mushrooms: similarities and differences. Roczn Państw Zakł Hig (Ann Nat Inst Hyg) 71:15–25
Prand-Stritzko B, Steinhauser G (2018) Characteristics of radiocesium contaminations in mushrooms after the Fukushima nuclear accident: evaluation of the food monitoring data from March 2011 to March 2016. Environ Sci Pollut Res 25:2409–2416
Samat SB, Green S, Beddoe AH (1997) The 40 K activity of one gram of potassium. Phys Med Biol 42:407–413
Skibniewska KA, Smoczyński S (1999) Wpływ obróbki kulinarnej na poziom radiocezu w grzybach. Rocz Panstw Zakl Hig 50:157–167
Steinhauser G, Steinhauser V (2016) A simple and rapid method for reducing radiocesium concentrations in wild mushrooms (Cantharellus and Boletus) in the course of cooking. J Food Prot 79:1995–1999
Steinhauser G, Brandl A, Johnson TE (2014) Comparison of the Chernobyl and Fukushima nuclear accidents: a review of the environmental impacts. Sci Total Environ 470-471:800–817
Stijve T (1994) Extraction of radiocesium from contaminated mushrooms. Observ Mycol (Bull de l’Observatoire Mycologique) 6:2–9
Stijve T (1996) Potassium content and growth rate of higher fungi. Aust Mycol Newsl 15:70–71
Stijve T, Poretti M (1990) Radioactivity in mushrooms. Mushroom 8:5–9
Tanaka S, Adati T, Takahashi T, Fujiwara K, Takahashi S (2018) Concentrations and biological half-life of radioactive cesium in epigeic earthworms after the Fukushima Dai-ichi Nuclear Power Plant accident. J Environ Radioact 192:227–232
Travnikova IG, Shutov VN, Bruk GY, Balonov MI, Skuterud L, Strand P, Pogorely JA, Burkova TF (2002) Assessment of current exposure levels in different populations groups of the Kola Peninsula. J Environ Radioact 60:235–248
Tucaković I, Barišić D, Grahek Ž, Kasap A, Širić I (2018) 137Cs in mushrooms from Croatia sampled 15-30 years after Chernobyl. J Environ Radioact 181:147–151
Tuo F, Xu C, Zhang J, Li W, Zhou Q, Zhang Q, Su X (2014) Measurement of activity concentrations for 137Cs and 40K in edible wild mushrooms collected from Mangshi, Yunnan province and evaluation of dose to adult (in Chinese). Chin J Radiol Med Prot 34:621–625
Wang Y, Zalewska T, Apanel A, Zhang J, Wiejak A, Falandysz J (2015) 137Cs, 134Cs and natural 40K in sclerotia of Wolfiporia extensa fungus collected across of the Yunnan land in China. J Environ Sci Health B 50:654–658
Wu SR, Luo XL, Liu B, Gui MY (2010) Analyse and advise to research and development of wild edible fungi. Food Sci Technol 35:100–103 (in Chinese with English abstract)
Yoshida S, Muramatsu Y, Steiner M, Belli M, Pasquale A, Rafferty B, Rühm W, Rantavaara A, Linkov I, Dvornik A, Zhuchenko T (2000) Relationship between radiocesium and stable cesium in plants and mushrooms collected from forest ecosystems with different contamination levels. P-11-244, 1-6
Zalewska T, Cocchi L, Falandysz J (2016) Radiocaesium in Cortinarius spp. mushrooms in the regions of the Reggio Emilia in Italy and Pomerania in Poland. Environ Sci Pollut Res 23:23169–23174
Zhang D, Frankowska A, Jarzyńska G, Kojta AK, Drewnowska M, Wydmańska D, Bielawski L, Wang J, Falandysz J (2010) Metals of King Bolete (Boletus edulis) collected at the same site over two years. Afr J Agric Res 5:3050–3055
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Georg Steinhauser
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Jerzy Falandysz is a visiting professor at affiliations 2 and 3.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Falandysz, J., Zhang, J. & Saniewski, M. 137Cs, 40K, and K in raw and stir-fried mushrooms from the Boletaceae family from the Midu region in Yunnan, Southwest China. Environ Sci Pollut Res 27, 32509–32517 (2020). https://doi.org/10.1007/s11356-020-09393-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-020-09393-w