
Abstract
The global population of great cormorants (Phalacrocorax carbo L.) is on the rise. These birds, characterized by rapid metabolism, can deposit large quantities of feces, and because they breed on the land but forage on water, both terrestrial and aquatic environments can be simultaneously affected by their activities. The contribution of great cormorants in the dispersal of bacterial and viral pathogens has been immensely studied; whereas, the occurrence of eukaryotic parasites such as protozoans and microsporidians in these birds is little known. The present study investigated the presence of dispersive stages of potentially zoonotic protozoans belonging to the genera Blastocystis, Giardia and Cryptosporidium, and Microsporidia spores in feces collected from birds inhabiting the breeding colony established at one lake island in Poland, Europe. The feces were examined by coprological techniques (staining with iron hematoxylin, Ziehl-Neelsen, and modified Weber’s chromotrope 2R–based trichrome), and with immunofluorescence antibody MERIFLUOR Cryptosporidium/Giardia assay. As found, the Cryptosporidium oocysts were identified rarely in 8% of samples (2/25; 3–5 × 103/g) and no cysts of Giardia and Blastocystis were detected. Microsporidian spores were detected in 4% of samples (1/25) but at very high frequency (4.3 × 104/g). No dispersive stages of parasites were identified in water samples collected from the littoral area near the colony. Despite the profuse defecation of cormorants, their role in the dispersion of the investigated parasites may not be as high as hypothesized.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The global population of great cormorant (Phalacrocorax carbo L.) is on the systematic rise, particularly in some parts of Europe where the number of breeding pairs is estimated to exceed 400,000 (Bregnballe et al. 2014; Klimaszyk and Rzymski 2016). This bird species, exterminated for decades by humans, have become numerous not only due to international and national law enforcements but also as a result of its high ecological adaptation, ability to forage on marine and freshwater environments, lack of regular predators, and increase in fish biomass due to the eutrophication and climate changes (Cramp and Simmons 1997; White et al. 2011; Skov 2011; Klimaszyk and Rzymski 2016). As these birds are generally gregarious, appear collectively, gather in flocks, nest in colonies on land, can feed on a relatively large area (up to 30 km from the colony), and simultaneously on various water systems, they may represent a significant biological factor that could trigger environmental modifications (Klimaszyk et al. 2015a; Klimaszyk and Rzymski 2016).
The effect of P. carbo on nutrient cycling, soil chemistry, terrestrial and aquatic vegetation, excessive algae growth (Ligęza and Smal 2003; Klimaszyk et al. 2015a, b), invertebrate communities (Kolb et al. 2010), and fish populations (Ostman et al. 2013) has been studied extensively (for review see Klimaszyk and Rzymski 2016). Some studies also addressed the role of these birds in dispersion of bacterial pathogens such as Escherichia coli (Klimaszyk 2012; Klimaszyk and Rzymski 2013a, 2016), avian influenza virus (Albini et al. 2014), avian paramyxovirus (Schelling et al. 1999), and West Nile virus (Iashkulov et al. 2008; Table 1). Recent studies also investigated the presence of gastric nematodes in these birds (Dziekońska-Rynko and Rokicki 2008; El-Dakhly et al. 2012).
The occurrence of dispersive stages of intestinal protozoan parasites such as Giardia cysts and Cryptosporidium oocysts in great cormorants is, however, largely unknown and, so far, reported only in two studies examining the bird feces (Medema 1999; Plutzer and Tomor 2009). The parasites were detected in cormorant droppings, but due to low number of samples in both studies, the definite conclusions on the role of cormorants in dispersion of these potential pathogens cannot be drawn. The presence of microsporidian spores in great cormorant, on the other hand, was so far a subject to only one study conducted recently in Slovakia. The spores, identified molecularly as Encephalitozoon cuniculi were detected using PCR in several fecal samples (Malčeková et al. 2013).
As some microsporidian and protozoan parasites are potentially infectious in mammals including human (Ehsan et al. 2015), it is of high priority to conduct further studies elucidating the role of great cormorants in their dissemination. As these birds represent a very important intermediate link in some food webs (Gwiazda et al. 2010, Skov et al. 2014) and a factor facilitating the dislocation of matter between terrestrial and aquatic ecosystems (Marion et al. 1994), it can be rather anticipated that they could also be responsible for high dispersion of parasites because they consume relatively large fish biomass, estimated at 350 g per day (Carss 1997). Various fish species were, in turn, identified as potential reservoirs of protozoan intestinal parasites such as Giardia sp. (Yang et al. 2010; Ghoneim et al. 2012) or Cryptosporidium sp. (Barugahare et al. 2011; Gabor et al. 2011) as well as microsporidian parasites (Lom and Nilsen 2003). Moreover, cormorants are characterized by rapid metabolism and the birds defecate on average 30 g dry weight of droppings per day (Marion et al. 1994). Deposited on relatively small area of colony, their chemical and microbial content can be subsequently transported with surface runoff and/or groundwater to the nearby lake (Klimaszyk and Rzymski 2011, 2013b; Klimaszyk et al. 2015a, b). Therefore, it is of great interest to evaluate the importance of great cormorants as vectors of dispersive stages of intestinal protozoan parasites in terrestrial and aquatic environments.
The present study aimed to investigate the presence of Cryptosporidium oocysts, cysts of Giardia and Blastocystis, and microsporidian spores in fecal samples collected from the colony of P. carbo during the breeding season. The colony, constituted of 170 breeding pairs, was located on the island of recreationally used, eutrophic Lake Chrzypsko (Northern Poland, Europe). To the best of our knowledge, this is the first study not only to survey such number of these birds in this regard but also to highlight that the role of great cormorants in dispersion of human intestinal protozoan and microsporidian parasites may not be as significant as expected.
Material and methods
The cormorant colony
The studied colony inhabits the Lake Chrzypsko (Poland, Europe) which is in the state of moderate eutrophy (Klimaszyk 2012). Owing to its location, the lake is intensely used for recreation. Numerous holiday resorts and bathing places are located on its shores. In the west bay, there is also a rowing training center, the racetrack of which stretches near the cormorant colony. The colony has been existing since the beginning of the twenty-first century (Klimaszyk 2012). It occupies the most northward island of the lake (Fig. 1) at the latitude and longitude of 52o 36′ 57″ N and 16o13′ 23″ E, respectively. An island has an area of 0.9 ha and slight elevation, up to 40 cm above the lake level. The counting of birds was performed by two independent observers during dawn-dusk prior to collection of fecal samples. During the investigated period (June 2013), 170 breeding pairs (approx. 600 individuals including adults and rearing chicks) were recorded. The foraging area of cormorants during this period is in the radius of 50 km from the colony, but outside the breeding season, adult birds and younglings may spread across the Europe (Bregnballe et al. 2014).

The studied island on Lake Chrzypsko (Poland) inhabited by cormorants and sampling points
Samples collection
Samples of cormorant feces were collected from the colony area in June 2013 using 10 trays (60 × 60 cm) located directly beneath bird nests at various points within the island (Fig. 1). The birds’ behavior was observed from the boat using binoculars and feces were systematically collected from each tray. A special care was taken to avoid collecting the fecal samples from the same nest. Plant detritus (leaves, branches) was removed from trays to minimize contamination of samples. Deposited feces were collected to sterile polypropylene tubes by pooling droppings from five different birds as one sample. All samples were immediately preserved with 20 mL potassium dichromate and transported to the laboratory in a lightproof insulated box containing a cooling factor. A total number of 25 pooled samples, and estimated droppings from 125 cormorants, were collected for subsequent parasitological examination. Additionally, samples of lake water (10 L each) were collected from four sampling sites at littoral area near the colony (shore zone) into sterile vessels (Fig. 1). These sites were selected because the shore lake zone was previously shown to be characterized by a high density of fecal bacteria originating from cormorant species (Klimaszyk and Rzymski 2013a; Klimaszyk et al. 2015b).
Parastiological examination of feces
All fecal samples were examined using coprological methods. From each pooled fecal sample, four smears were made. One direct wet smear was immediately microscopically examined under high dry power (total magnification ×400). The remaining three smears were stained with either: (i) modified Weber’s chromotrope 2R–based trichrome stain for Microsporidia spores (Weber et al. 1992), (ii) Ziehl-Neelsen stain for Cryptosporidium oocysts, or (iii) iron hematoxylin stain for cysts of Giardia and Blastocystis (Garcia 2001). Stained smears were microscopically screened using an oil-immersion objective (total magnification ×1000).
Additionally, to confirm identification of Cryptosporidium oocysts and/or Giardia cysts, all positive specimens were tested using a direct immunofluorescence antibody (IFA) test kit, MERIFLUOR Cryptosporidium/Giardia (Meridian Diagnostic, Cincinnati, Ohio, USA), was used according to the manufacturer’s instructions.
Parastiological examination of water samples
All water samples were examined using modified U.S. Environmental Protection Agency Method 1623 (U.S. Environmental Protection Agency 1999). The sediment was obtained by filtration using SM 16274 filter chamber (Sartorius, Germany) on cellulose acetate membranes with a nominal pore size of 0.8 μm (Merck Millipore, Ireland). The filters were then dissolved in acetone according to Graczyk et al. 1997. Each sample concentrate was analyzed using the Ziehl-Neelsen (Cryptosporidium oocysts), modified Weber’s chromotrope 2R-based trichrome stain (Microsporidia spores) and iron hematoxylin (Giardia and Blastocystis cysts) methods, and immunofluorescent assay (IFA).
Results
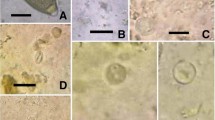
Cryptosporidium oocysts were identified in 2/25 (8%) of pooled fecal samples of great cormorant (Fig. 2a). All samples detected as positive by Ziehl-Neelsen staining were also positive by the immunofluorescence technique. In both Cryptosporidium-positive samples, a small number of oocysts, i.e., five to ten per slide, were detected, at frequency of 3 × 103/g and 5 × 103/g of feces. The mean length (±SD) and width (±SD) of identified oocysts was 5.0 (±0.0) and 5.4 (±0.5) μm, respectively. Spores of Microsporidia (Fig. 2b) were identified only in 1/25 (4%) of pooled fecal samples but at high concentration of 4.3 × 104/g of feces. The mean length (±SD) and width (±SD) of these spores was 1.8 (±0.4) and 1.2 (±0.2) μm, respectively. None of the investigated pooled fecal samples contained detectable cysts of Giardia and Blastocystis.

Spores of Microsporidia stained with modified Weber’s chromotrope 2R-based trichrome (a) and oocysts of Cryptosporidium stained with Ziehl-Neelsen (b), identified in Phalacrocorax carbo feces
None of investigated water sample was identified to contain microsporidian spores and dispersive stages of Cryptosporidium, Blastocystis and Giardia.
Discussion
The birds represent an important factor harboring and dispersing the microorganisms, including pathogens (Graczyk et al. 1998; Okulewicz 2014). The bird microbiota has been demonstrated to be affected by many different factors, such as infections and general health status, diet, and local microbial communities in environment (Palmgren et al. 1997; Lu et al. 2003; Santos et al. 2012). A main route through which the birds can take part in the dispersion of various microorganisms, including protozoa and microsporidia, is fecal excretion. Despite that great cormorants were previously reported to deposit large amounts of feces within the colonized areas (Marion et al. 1994; Klimaszyk and Rzymski 2016), the present study indicates that their role in dispersion of intestinal protozoan parasites may be largely limited and decidedly lower than theoretically expected. Some colonies of this bird may, however, still represent a source of dispersion of other human pathogens (Table 1).
It is important to fully elucidate the biological vectors of dispersive stages of protozoan and microsporidian parasites. These microorganisms are resistant to various environmental conditions, can lead to serious, acute gastrointestinal infections in human, and are usually characterized by the low infectious dose (Szumowski and Troemel 2015; Messner and Berger 2016). The presence of Cryptosporidium oocysts and Giardia cysts in source waters have already caused numerous documented outbreaks related in both drinking and recreational waters (Karanis et al. 2007). Various birds have already been demonstrated to contribute to contamination of surface waters with dispersive stages of these parasites, including species and genotypes representing a threat to human health (Smith et al. 1993; Graczyk et al. 1998; Majewska et al. 2009). In a Hungarian survey investigating feces of different bird species, one Giardia sp. cyst was identified microscopically and the presence of Cryptosporidium sp. was confirmed with PCR but the study examined only a single fecal sample collected from cormorant (Plutzer and Tomor 2009). Cryptosporidium oocysts (but not Giardia cysts) were also detected in feces collected from ten P. carbo individuals in the Netherlands inhabiting areas nearby man-made human reservoirs. The prevalence amounted to 20% and mean concentration in positive samples was estimated at 64 oocysts per gram—high enough to significantly contribute to the contamination of water reservoir (Medema 1999). Contrary to these findings, the present study, which employed fecal samples collected from significantly greater number of birds (125 individuals) found that the prevalence of Cryptosporidum oocysts was very low and no Blastocystis or Giardia cysts were present. This indicate that this species, at some inhabited sites, may not represent a significant source of dispersive stages of human protozoan parasites—particularly if one considers that none of investigated protozoans were identified in lake water near the colony.
The infection of the gastrointestinal tract by Microsporidia can also lead to severe, persistent diarrhea (Didier 2005). As shown, microsporidian species known to infect humans such as E. hellem are present in aquatic bird species including Anas platyrhynchos, Anser anser, Balearic pavonina, Cygnus atratus, C. melanocoryphus, C. olor and Coscoroba coscoroba (Słodkowicz-Kowalska et al. 2006). The E. cuniculi (but not E. hellem, E. intestinalis or E. bieneusi) was found in the Slovakian pilot study examining 40 samples of great cormorant feces at a relatively high prevalence of 42.5% (Malčeková et al. 2013). The present study showed decidedly lower frequency of spores in investigated population of 125 birds indicating that the role of great cormorants in Microsporidia dispersion may be highly site specific. However a relatively high number of spores were identified in one pooled sample (4.3 × 104/g); no dispersive stages of Microsporidia were identified in lake water near the colony. It should be highlighted that however the most widely used staining method to detect spores (chromotrope 2R modified trichome) was employed in the present study, it does not allow to distinguish particular species or genotypes of microsporidia. This is important if one considers that only some species (at least 15 from over 1200 identified so far) are known to be pathogenic for humans (Ramanan and Pritt 2014). These, in turn, can be identified by means of immunofluorescence assays using polyclonal or monoclonal antibodies and/or PCR (Ramanan and Pritt 2014). Further investigations are required to fully elucidate the environmental conditions contributing to the presence of microsporidian dispersive stages in cormorant feces, and to estimate risks for human health.
The present study was limited only to one great cormorant colony situated at the lake of low human pressure; therefore, the results should be treated cautiously upon any extrapolation. Sewage discharge can lead to increased contamination of water with dispersive stages of parasites and their presence in biota including waterfowls (Słodkowicz-Kowalska et al. 2015); thus, the prevalence of studied parasites may be different at sites varying in human pressure. It should be, however, highlighted that great cormorants usually nest within areas of negligible human impact (Klimaszyk and Rzymski 2016).
Conclusions
The present study investigated the presence of dispersive stages of potentially zoonotic protozoans belonging to the genera Blastocystis, Cryptosporidium and Giardia, and Microsporidia spores in feces of great cormorant. It was hypothesized that due to specific behavior and metabolism, these birds may represent an important vector for these parasites. Contrary to this, the prevalence of Cryptosporidium oocysts and microsporidian spores was very low, and no cysts of Giardia and Blastocystis were identified. The study indicates that this species may not play, at least at certain locations, a profound role in the dissemination of investigated parasites. Further research employing immunological and molecular methods is necessary to elucidate exact species of Microsporidia, and evaluate whether cormorants may disseminate those associated with human infection.
References
Albini K, Sigrist G, Keller H (2014) Shedding of zoonotic pathogens and analysis of stomach contents in great cormorants (Phalacrocorax carbo sinensis) from Switzerland between 2007 and 2012. Schweiz Arch Tierheilkd 156:389–394
Artois M, Manvell R, Fromont E, Schweyer JB (2002) Serosurvey for Newcastle disease and avian influenza a virus antibodies in great cormorants from France. J Wildlife Dis 38:169–171
Barugahare R, Dennis MM, Becker JA, Slapeta J (2011) Detection of Cryptosporidium molnari oocysts from fish by fluorescent-antibody staining assays for cryptosporidium spp. affecting humans. Appl Environ Microbiol 77:1878–1880
Bregnballe T, Lynch J, Parz-Gollner R, Marion L, Volponi S, Paquet J-Y, Carss DN, van Eerden MR (2014) Breeding numbers of great cormorants Phalacrocorax carbo in the Western Palearctic, 2012–2013. IUCN-Wetlands International Cormorant Research Group Report. - Scientific Report from DCE–Danish Centre for Environment and Energy No. 99
Carss DN (1997) Techniques for assessing cormorant diet and food uptake: towards a consensus view. Suppl Ric Biol Selvaggina 26:197–230
Cramp S, Simmons KEL (1997) The birds of the Western Palearctic: Vol. 1. Oxford University Press, Oxford-New York
Didier ES (2005) Microsporidiosis: an emerging and opportunistic infection in humans and animals. Acta Trop 94:61–76
Dziekońska-Rynko J, Rokicki J (2008) Infestation of the black cormorants (Phalacrocorax carbo sinensis L. 1758) from the colony on the Selment Wielki, with the nematode Contracaecum rudolphii Hartwich, 1964 (Nematoda, Anisakidae). Oceanol Hydrobiol Stud 37:61–71
Ehsan A, Geurden T, Casaert S, Paulussen J, De Coster L, Schoemaker T, Chalmers R, Grit G, Vercruysse J, Claerebout E (2015) Occurrence and potential health risk of Cryptosporidium and Giardia in different water catchments in Belgium. Environ Monit Assess 187:6
El-Dakhly KM, El-Nahass E, Uni S, Tuji H, Sakai H, Yanai T (2012) Levels of infection of gastric nematodes in a flock of great cormorants (Phalacrocorax carbo) from Lake Biwa, Japan. J Helminthol 86:54–63
Gabor LJ, Srivastava M, Titmarsh J, Dennis M, Gabor M, Landos M (2011) Cryptosporidiosis in intensively reared barramundi (Lates calcarifer). J Vet Diagn Investig 23:383–386
Garcia LS (2001) Diagnostic Medical Parasitology, 4th edn. ASM Press, Washington
Ghoneim NH, Abdel-Moein KA, Saeed H (2012) Fish as a possible reservoir for zoonotic Giardia duodenalis assemblages. Parasitol Res 110:2193–2196
Graczyk TK, Cranfield MR, Fayer R (1997) Recovery of waterborne oocysts of Cryptosporidium from water samples by the membrane-filter dissolution method. Parasitol Res 83:121–125
Graczyk TK, Fayer R, Trout JM, Lewis EJ, Farley CA, Sulaiman I, Lal AA (1998) Giardia sp. cysts and infectious Cryptosporidium parvum oocysts in the feces of migratory Canada geese (Branta canadensis). Appl Environ Microbiol 64:2736–2738
Gwiazda R, Jarocha K, Szarek-Gwiazda E (2010) Impact of small cormorant (Phalacrocorax carbo sinensis) roost on nutrients and phytoplankton assemblages in the littoral regions of submontane reservoir. Biologia 65:742–748
Iashkulov KB, Shchelkanov MI, L'vov SS, Dzhambinov SD, Galkina IV, Fediakina IT, Bushkieva BT, Morozova TN, Kireev DE, Akanina DS, Litvin KE, Usachev EV, Prilipov AG, Grebennikova TV, Gromashevskiĭ VL, Iamnikova SS, Zaberezhnyĭ AD, L'vov DK (2008) Isolation of influenza virus A (Orthomyxoviridae, influenza A virus), Dhori virus (Orthomyxoviridae, Thogotovirus), and Newcastle's disease virus (Paromyxoviridae, Avulavirus) on the Malyi Zhemchuzhnyi Island in the north-western area of the Caspian Sea. Vopr Virusol 53:34–38
Karanis P, Kourenti C, Smith H (2007) Waterborne transmission of protozoan parasites: a worldwide review of outbreaks and lessons learnt. J Water Health 5:1–38
Klimaszyk P (2012) May a cormorant colony be a source of coliform and chemical pollution in a lake? Oceanol Hydrobiol Stud 41:67–73
Klimaszyk P, Rzymski P (2011) Surface runoff as a factor determining trophic state of midforest lake (Piaseczno Małe, North Poland). Pol J Environ Stud 5:1203–1210
Klimaszyk P, Rzymski P (2013a) Impact of cormorant (Phalacrocorax carbo sinensis L.) colonies on microbial pollution in lakes. Limnol Rev 13:139–145
Klimaszyk P, Rzymski P (2013b) Catchment vegetation can trigger lake dystrophy through changes in runoff water quality. Ann Limnol Int J Limnol 49:191–197
Klimaszyk P, Rzymski P (2016) The complexity of ecological impacts induced by great cormorants. Hydrobiologia 771:13–31
Klimaszyk P, Piotrowicz R, Rzymski P (2015a) Changes in the ecosystem of shallow softwater lake induced by the great cormorant roosting colony. J Limnol 74:114–122
Klimaszyk P, Brzeg A, Rzymski P, Piotrowicz R (2015b) Black spots for aquatic and terrestrial ecosystems: impact of a perennial cormorant colony on the environment. Sci Total Environ 517:222–231
Kolb SG, Jerling L, Hamback PA (2010) The impact of cormorants on plant-arthropod food webs on their nesting islands. Ecosystems 13:353–366
Ligęza S, Smal H (2003) Accumulation of nutrients in soils affected by perennial colonies of piscivorous birds with reference to biogeochemical cycles of elements. Chemosphere 52:595–602
Lom J, Nilsen F (2003) Fish microsporidia: fine structural diversity and phylogeny. Int J Parasitol 33:107–127
Lu J, Idris U, Harmon B, Hofacre C, Maurer JJ, Lee MD (2003) Diversity and succession of the intestinal bacterial community of the maturing broiler chicken. Appl Environ Microbiol 69:6816–6824
Majewska AC, Graczyk TK, Słodkowicz-Kowalska A, Tamang L, Jedrzejewski S, Zduniak P, Solarczyk P, Nowosad A, Nowosad P (2009) The role of free-ranging, captive, and domestic birds of Western Poland in environmental contamination with Cryptosporidium parvum oocysts and Giardia lamblia cysts. Parasitol Res 104:1093–1099
Malčeková B, Valenčáková A, Molnár L, Kočišová A (2013) First detection and genotyping of human-associated microsporidia in wild waterfowl of Slovakia. Acta Parasitol 58:13–17
Marion L, Clergeau P, Brient L, Bertu G (1994) The importance of avian-contributed nitrogen (N) and phosphorus (P) to Lake Grand-Lieu, France. Hydrobiologia 279(280):133–147
Medema GJ (1999) Cryptosporidium and Giardia: new challenges to the water industry. Ph.D. Thesis. University of Utrecht
Messner MJ, Berger P (2016) Cryptosporidium infection risk: results of new dose-response modeling. Risk Anal 36:1969–1982
Okulewicz A (2014) Helminths in migrating and wintering birds recorded in Poland. Ann Parasitol 60:19–24
Ostman O, Boström MK, Bergström U, Andersson J, Lunneryd SG (2013) Estimating competition between wildlife and humans-a case of cormorants and coastal fisheries in the Baltic Sea. PLoS One 8:e83763
Palmgren H, Sellin M, Bergstrom S, Olsen B (1997) Enteropathogenic bacteria in migrating birds arriving in Sweden. Scand J Infect Dis 29:565–568
Plutzer J, Tomor B (2009) The role of aquatic birds in the environmental dissemination of human pathogenic Giardia duodenalis cysts and Cryptosporidium oocysts in Hungary. Parasitol Int 58:227–231
Ramanan P, Pritt BS (2014) Extraintestinal microsporidiosis. J Clin Microbiol 52:3839–3844
Santos SS, Pardal S, Proença DN, Lopes RJ, Ramos JA, Mendes L, Morais PV (2012) Diversity of cloacal microbial community in migratory shorebirds that use the Tagus estuary as stopover habitat and their potential to harbor and disperse pathogenic microorganisms. FEMS Microbiol Ecol 82:63–74
Schelling E, Thur B, Audige L (1999) Epidemiological study of Newcastle disease in backyard poultry and wild bird populations in Switzerland. Avian Pathol 28:263–272
Skov H (2011) Waterbird populations and pressures in the Baltic Sea. Norden Publ
Skov H, Jepsen N, Baktoft H, Jansen T, Pedersen S, Koed A (2014) Cormorant predation on PIT-tagged lake fish. J Limnol 73:177–186
Słodkowicz-Kowalska A, Graczyk TK, Tamang L, Jedrzejewski S, Nowosad A, Zduniak P, Solarczyk P, Girouard AS, Majewska AC (2006) Microsporidian species known to infect humans are present in aquatic birds: implications for transmission via water? Appl Environ Microbiol 72:4540–4544
Słodkowicz-Kowalska A, Majewska AC, Rzymski P, Skrzypczak Ł, Werner A (2015) Human waterborne protozoan parasites in freshwater bivalves (Anodonta anatina and Unio tumidus) as potential indicators of fecal pollution in urban reservoir. Limnologica 51:32–36
Smith HV, Brown J, Coulson JC, Morris GP, Girdwood RWA (1993) Occurrence of oocysts of Cryptosporidium sp. in Larus spp. gulls. Epidemiol Infect 110:135–143
Szumowski SC, Troemel ER (2015) Microsporidia-host interactions. Cur Opinion Microbiol 26:10–16
Tausova D, Dolejska M, Cizek A, Hanusova L, Hrusakova J, Svoboda O, Camlik G, Literak I (2012) Escherichia coli With extended-spectrum beta-lactamase and plasmid-mediated quinolone resistance genes in great cormorants and mallards in Central Europe. J Antimicrob Chemother 6:1103–1107
U.S. Environmental Protection Agency (1999) Method 1623: Cryptosporidium and Giardia in water by filtration/IMS/FA. Publication EPA-821-R-99-006. U.S. Environmental Protection Agency Office of Water, Washington, D.C
Weber R, Bryan RT, Owen RL, Wilcox CM, Gorelkin L, Visvesvara GS (1992) Improved light-microscopical detection of microsporidia spores in stool and duodenal aspirates. The enteric opportunistic infections working group. N Engl J Med 326:161–166
White CR, Boertmann D, Gremillet D, Butler PJ, Green JA, Martin GR (2011) The relationship between sea surface temperature and population growth of great cormorants near Disco Bay, Greenland. Ibis 153:170–174
Yang R, Reid A, Lymbery A, Ryan U (2010) Identification of zoonotic Giardia genotypes in fish. Int J Parasitol 40:779–785
Acknowledgments
Piotr Rzymski is supported by the Foundation for Polish Science (FNP) (START 091.2016).
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible editor: Robert Duran
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Rzymski, P., Słodkowicz-Kowalska, A., Klimaszyk, P. et al. Screening of protozoan and microsporidian parasites in feces of great cormorant (Phalacrocorax carbo). Environ Sci Pollut Res 24, 9813–9819 (2017). https://doi.org/10.1007/s11356-017-8652-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-017-8652-y