
Abstract
Xyloglucan is the major hemicellulosic polymer found in the primary cell walls of dicots. Xyloglucan tethers cellulose microfibrils conferring rigidity and strength for maintenance of cell integrity, and it is thought that its metabolism contributes to cell elongation and thus plant growth. Here, we have cloned and characterized a Eucalyptus grandis gene ortholog of the Arabidopsis thaliana MUR3 gene (xyloglucan galactosyltransferase), thus termed EgMUR3. EgMUR3 represents an intronless sequence of 1,854 bp predicted to encode a protein of 617 amino acid residues. It exhibits 73% identity and 82% similarity to the A. thaliana MUR3 gene. To demonstrate that this gene encodes a functional enzyme, the putative ORF was cloned into a binary vector under the control of a constitutive promoter and transformed into the A. thaliana mur3 mutant. The effect of the genetic complementation was investigated by xyloglucan oligosaccharide fingerprinting of wall material. The results confirmed that EgMUR3 represents indeed a xyloglucan galactosyltransferase of E. grandis able to use endogenous substrate(s) in A. thaliana, suggesting that both species share common steps in xyloglucan biosynthesis.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Eucalypts are among the fastest growing trees and are suitable candidates as biomass resource for bioenergy and pulping purposes. Breeding targets have been mainly biomass yield gains, fiber quality, biotic and abiotic stress tolerance, flowering control, and herbicide resistance. The domestication of the Eucalyptus genus is just in its infancy, mainly because classic genetic breeding of forest trees is hampered by the long juvenile stage and many traits of interest can be only assessed at rotation age (Boerjan 2005). Molecular and classic breeding are two approaches taken in parallel toward the designing of elite trees. Biotechnology exploit the advantage of speeding up the investigation of gene function by transgene transference to model plants, while classic breeding offers the opportunity of identifying new genetic elements controlling plant traits through the exploration of the natural diversity.
Comprehension of cell wall metabolism is the direction toward altering biomass for biofuel and pulping purposes in this economically important tree. In spite of literature describing protocols for Eucalyptus spp. transformation (Serrano et al. 1996; Moralejo et al. 1998; Gonzalez et al. 2002), transcriptional analysis of gene expression (Paux et al. 2005; Foucart et al. 2006; Navarro et al. 2009), gene structure, and diversity (McKinnon et al. 2005; Sassaki et al. 2005), there is a lack of report on functional analysis of cell wall-related genes in eucalypt species in comparison to the other important forestry species (Chen et al. 2001; Sibout et al. 2002; Baucher et al. 2003). Much of the research being conducted in this area is under confidentiality policies adopted by private companies. Recently, a paper reported that the overexpression of a gene encoding the Eucalyptus camaldulensis HD-Zip class II transcription factor increased fiber length and growth in transgenic tobacco plants (Sonoda et al. 2009).
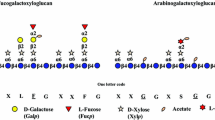
All plant cells are encased in a wall, which consists of complex heteroglycans and polyphenolics. The major load-bearing structure in the dicot primary wall, i.e., the wall of elongating cells, is a cellulose–xyloglucan network, whose metabolism plays a major role in cell elongation (Somerville et al. 2004). Xyloglucan (XyG) coats and permeates cellulose microfibrils (Pauly et al. 1999a), limiting their aggregation and connecting them via tethers that directly or indirectly regulate the mechanical properties of the wall. XyG consists of β-1,4 linked glucosyl residues, which interact tightly and non-covalently with the cellulose microfibrils. Unlike cellulose, XyG contains quite diverse side-chain structures that render the polymer soluble in aqueous solutions. XyG structure and the adopted nomenclature is described by Fry et al. (1993) and Hoffman et al. (2005). The precise distribution and function of the individual side-chain moieties are not known. However, it is thought that the galactosyl residue is needed for solubility of the polymer (de Lima and Buckeridge 2001). Furthermore, fucose residues are thought to flatten the XyG backbone, allowing for an enhanced interaction with cellulose microfibrils (Levy et al. 1997).
With the help of forward and reverse genetic approaches, Arabidopsis mutants have been obtained that shed light on the biosynthesis of XyG. The first and best-known large-screening for cell wall mutants was conducted by Reiter and colleagues by the characterization of mutants with an altered wall-sugar composition (Reiter et al. 1997). Among these so-called mur-mutants, mutants exhibiting a decrease in fucose were represented by mur1, mur2, and mur3. The XyG structure in all of those mutants was altered (Lerouxel et al. 2002). MUR1 represents a GDP-mannose dehydratase, an enzyme needed for the biosynthesis of the precursor GDP-fucose (Zablackis et al. 1996). MUR2 represents an XyG fucosyltransferase (Perrin et al. 1999). MUR3 encodes for a galactosyltransferase, which galactosylates only the third xylose residue in the XXXG core leading to XXLG (Madson et al. 2003). In addition to a substantial reduction in galactosyl moieties, and hence a lack of the fucoyltransferase acceptor substrate site, mur3 XyG also lacks fucosyl residues. Different phenotypes related to different MUR3 alleles have been described. The occurrence of a stop codon after the transmembrane domain (TMD) in another mur3 allele leads to a truncated protein, which caused disorganization of the endomembrane system (Tamura et al. 2005). Despite a significantly altered XyG structure, mur3 plants do not exhibit any apparent change in morphology and/or growth behavior under regular growth conditions (Reiter et al. 1997). However, a change in tensile strength of etiolated hypocotyls was observed (Pena et al. 2004).
In this study, we identified a Eucalyptus grandis MUR3 ortholog here termed EgMUR3. EgMUR3 was overexpressed in the Arabidopsis thaliana mur3 mutant to assess its functionality. EgMUR3 was confirmed to be a functional xyloglucan galactosyltransferase expressed mainly in elongating stem and young leaves of young Eucalyptus urograndis plants.
Materials and methods
Plant lines and growth conditions
Seeds of the mur3 mutant were obtained from the Arabidopsis Biological Resource Center at the Ohio State University (Columbus). The mutation was in an Ecotype Columbia (Col-0) background. After a cold treatment by incubation of the seeds at 4°C in darkness for 48 h, in order to break seed dormancy, A. thaliana Col-0 and mur3 were grown in the greenhouse of the Max-Planck Institute for Molecular Plant Physiology, Golm, Germany, with the following temperature/humidity parameters, 20°C/80% from 6 to 22 h, 18°C/50% from 22 to 6 h. Light intensity was 120 mmol m−2 s−1 until seed harvesting. Long-day condition for flowering induction consisted of 16 h light/8 h dark alternating cycles at 22°C, 120 mmol m−2 s−1 of light intensity and 60% humidity. Short-day conditions for producing leaf material for RNA extraction and cell wall preparation consisted of 8 h light/16 h dark at 22°C, under 120 mmol m−2 s−1 light intensity and 60% humidity.
Cloning of EgMUR3
A BAC library of E. grandis maintained at the Genomics Laboratory (BIOAGRO/UFV, Brazil) was used to clone EgMUR3. BACs were pooled and screened by PCR for genes involved in cell wall metabolism with primers designed according to EST data obtained from a private EST database (at http://www.lge.ibi.unicamp.br/eucalyptus). A positive BAC clone (87E04) of approximately 180 kb containing a putative ortholog of the MUR3 gene of A. thaliana was identified using the following primer pairs: egmur for 5′-GGCCACATTGTCCTTGTTTT-3′ and egmur rev 5′-GGGAGTTTCCAAATGGCTCT-3′. A shotgun library of this BAC was constructed in order to obtain the complete sequence of EgMUR3. The library was constructed after physical shearing of the insert fragment released from the BAC clone 87E04 using the TOPO® Shotgun Subcloning kit (Invitrogen). Sheared fragments were ligated into a pCR®4Blunt-TOPO® plasmid creating the EUGR_SG_006 subclone library containing inserts of approximately 1.5 kb. Large-scale sequencing of this library was performed (MegaBACE 500 sequencer, Amersham Pharmacia) using the universal primers SP6 (5′-ATTTAGGTGACACTATAG-3′) and T7 (5′-TAATACGACTCACTATAGGG3′), which border the pCR®4Blunt-TOPO® cloning site, resulting in the complete sequence of the 180-kb genomic insert. The raw data was converted to FASTA query sequences and subjected in batches to BLAST X analysis. One of the shotgun sequences showed a high sequence homology to the A. thaliana MUR3 gene. Plasmid DNA of subclones of the EUGR_SG_006 library displaying sequence overlap to the A. thaliana MUR3 ortholog was isolated, and the insert between SP6 and T7 primers was fully sequenced in order to obtain the full EgMUR3 gene sequence. Reads were processed with Phred (>20) for base calling, cross_match for removing vector sequences, and Phrap and Consed for assembly (Ewing and Green 1998; Ewing et al. 1998; Gordon et al. 1998). The final contig was analyzed using GENSCAN (http://genes.mit.edu/GENSCAN.html) to determine the full EgMUR3 sequence.
Sequence analysis
Peptide properties were predicted by ProtParam (http://www.expasy.org/cgi-bin/protparam). The Conserved Domains Database (http://www.ncbi.nlm.nih.gov/Structure/cdd/cdd.shtml) was searched for putative catalytic domains. Analysis of EgMUR3 amino acid sequence for transmembrane domain was performed with TMHMM v. 2.0 (http://www.cbs.dtu.dk/services/TMHMM-2.0) and TopPred (http://bioweb.pasteur.fr/seqanal/interfaces/toppred.html). Hydrophobic cluster analysis (Callebaut et al. 1997; Gaboriaud et al. 1987) was performed at http://mobyle.rpbs.univ-paris-diderot.fr/cgi-bin/portal.py?form=HCA.
Phylogenetic analysis
Phylogenetic and molecular evolutionary analyses were conducted using MEGA v. 4 (Tamura et al. 2007) and box-shading with GeneDoc (Nicholas et al. 1997). The evolutionary history was inferred using the neighbor-joining method (Saitou and Nei 1987). The optimal tree had the sum of branch length = 5.95201462. Next to the branches, the percentage of replicate trees in which the associated taxa clustered together in the bootstrap test (1,000 replicates) is shown (Felsenstein 1985). The tree was drawn to scale, with branch lengths in the same units as those of the evolutionary distances used to infer the phylogenetic tree. The evolutionary distances were computed using the Dayhoff matrix-based method (Schwarz and Dayhoff 1979) and are in the units of the number of amino acid substitutions per site. All positions containing gaps and missing data were eliminated from the dataset. There were a total of 33 positions in the final dataset.
Construction of pBinAr-EgMUR3 and plant transformation
Primers containing adaptamers for KpnI and SalI (egmur3_kpn_for 5′- GGTACCATGAGACGCCGTTCGTCGGCGA-3′ and egmur3_sal_rev 5′-GTCGACTCATGACTGGTCTCTCTGCTCGTT-3′) were used to amplify the EgMUR3 coding sequence from the BAC clone 87E04. A high-fidelity PCR with Pfu Turbo DNA polymerase (Invitrogen) was facilitated according to the manufacturer's recommendations. About 20 ng of BAC DNA were used as template in the PCR. A gradient procedure was used, starting from 65°C to 55°C (−1°C/cycle). Other steps consisted of 25 amplification cycles with primer annealing temperature set to 56°C. After the last cycle, 0.5 µl of Taq DNA polymerase was added to the mixture and an additional amplification cycle was performed during 10 min.
The PCR product was directly cloned into pCR®II-TOPO® vector (Invitrogen) according to manufacturer's instructions. The ligation reaction was transformed into one-shot competent bacteria (Invitrogen). Ampicillin-resistant colonies were picked, and plasmid DNA was extracted by alkaline lysis method (Sambrook and Russell 2001). Plasmidial DNA was analyzed by digestion with restriction enzymes, and a positive clone was selected and fully sequenced with egmur3_kpn_for and egmur3_sal_rev primers to confirm for the absence of mutations in the PCR-amplified EgMUR3 ORF. The 1.8-kb insert corresponding to the EgMUR3 ORF was excised from pCR®II-TOPO®-EgMUR3 with KpnI and SalI restriction enzymes, gel-purified using the Nucleospin Extract II kit (Machery-Nagel), and ligated into the KpnI–SalI restriction sites in the MCS (multiple cloning site) of pBinAr (Hofgen and Willmitzer 1990), a pBIN19 derivative (Bevan 1984) containing the 35S promoter from the cauli-flower mosaic virus.
The pBinAr-EgMUR3 construct was transformed and amplified into Escherichia coli DH5α. The pBinAr-EgMUR3 plasmid was then purified and transformed into Agrobacterium tumefaciens strain GV 3101 (pMP90) by electroporation (Mersereau et al. 1990). After confirmation that the A. tumefasciens strain contained the proper EgMUR3 gene, pBinAr-EgMUR3 and an empty vector control (pBinAr) were transformed into A. thaliana mur3 mutant by A. tumefaciens-mediated transformation (Bechtold and Pelletier 1998; Clough and Bent 1998). Infiltrated plants were grown for production of seeds. Sterilized seeds from infiltrated plants were placed on Petri dishes with 1% (w/v) sucrose AMOZ media (11.0 g L−1 MS medium, 2.5 g L−1 MES, 7 g L−1 bacto-agar, pH 5.7) plus antibiotics (50 µg mL−1 kanamycin, 250 µg ml−1 β-bactyl) for the selection of transformant plants. Plants were grown in long-day phytotron as described before.
To confirm the transgenic status of antibiotic resistant plants, total RNA was extracted from leaf material of 5-week-old plants using the RNeasy Plant Mini Kit (Qiagen). For cDNA synthesis, contaminant genomic DNA was removed with the DNA-free™ DNAse Treatment & Removal Reagents (Ambion®). The cDNA synthesis was performed with Bioscript reverse transcriptase (Bioline USA Inc.) according to manufacturer's recommendations.
EgMUR3 message was detected by PCR with EgMUR3 specific primers (egmur3RT_for 5′-CCATTGCGGTTAGGACTGAG-3′ and egmur3RT_rev 5′-TGCTCTTAGTTCCCACCACTT-3′) using the cDNA library as template. The constitutive APT gene (adenine phosphoribosyltransferase 1, At1g27450) was amplified as internal control with the primers: APT_for 5′-TCCCAGAATCGCTAAGATTGCC-3′ and APT_rev 5′-CCTTTCCCTTAAGCTCTG-3′. The RT-PCR was performed with the recombinant Taq DNA polymerase from Invitrogen according to manufacturer's recommendations. Extension time was 1.5 min, and annealing temperature was 55°C. To confirm the specificity of primers used to amplify EgMUR3, a control PCR using EgMUR3 primers and cDNA preparation from WT as template was conducted. MUR3 gene primers were mur3RT_for 5′-GCACCAGTAGCCAATTCTAG-3′ and mur3RT_rev 5′-GTCGCATACCAACCTTCATC-3′. In all RT-PCR analysis performed, 27 cycles were used.
Differential expression analysis of EgMUR3 by RT-qPCR
E. urograndis plants were grown in pots with soil for 90 days under regular irrigation regime. Samples of roots, apical portion of stem, and expanding leaves were collected and pooled (two pools of three plants) for RNA extraction. Total RNA was extracted (Wang et al. 2008), and total RNA concentration was measured spectrophotometrically (Hitachi U2000 spectrophotometer) under 260 and 280 nm wavelengths. RNA concentration of all samples was adjusted to 300 ng µL−1. Physical integrity of the RNA was assessed in 1% agarose gel (Wilson and Walker 2000). Afterwards, 2 µg of total RNA from each organ were treated with DNAse I (Promega, USA) according to the manufacture's recommendations. The cDNA synthesis was carried out with Superscript II (Invitrogen, USA). Control experiments consisted of omitting Superscript II in two samples from each tissue. The RT-PCR was conducted from the cDNA library and the controls with primers for the EgMUR3 gene and for two housekeeping genes: histone H2B and ribosomal protein L23A. EgMUR3 primers for qPCR were based on the EgMUR3 sequence. The following EgMUR3 primers were used: qEgMUR3_for 5′ GACAGCGAAGAACCAGCACTC 3′ and qEgMUR3_rev 5′ GAGTTTCCAAATGGCTCTCAGTC 3′. Primers for constitutive genes were designed according to EST database information for Eucalyptus (http://www.lge.ibi.unicamp.br/eucalyptus/). Expression of the histone H2B and the ribosomal protein L23A were monitored in each qPCR experiment. Primers for histone H2B were: H2B_for 5′ GAGCGTGGAGACGTACAAGA 3′ and H2B_rev 5′ GGCGAGTTTCTCGAAGATGT 3′. Primers for the ribosomal protein L23A were: RPtnL23A_for 5′ AAGGACCCTGAAGAAGGACA 3′ and RPtnL23A_rev 5′ CCTCAATCTTCTTCATCGCA 3′. All primers were used at a final concentration of 0.12 pmol in each primer-specific reaction. The 20-µL final volume reaction contained 1× High Fidelity PCR Buffer; 3.0 mM MgCl2, 25 µM of each dNTPs; 0.04 µL of ROX; 0.08 µL BSA; 2.0 µL SYBR Green (1:10,000 dilution), and 0.25 U Taq Platinum (all reagents from Invitrogen, USA). Three technical and two biological replicates for each sample were performed. The real-time PCR was conducted in a Step-One Plus equipment (Applied Biosystems, USA).
Primers validation based on the relative standard curve was performed with three technical replicates of leaf samples from the biological pool 1 as four geometric dilutions (1:5) of the cDNA reaction (1, 5, 25, and 125 times diluted). Afterwards, 0.28 µL of the cDNA sample was used per reaction. The PCR program consisted of 90°C for 5 min, 40 cycles of 95°C for 15 s, 58°C for 30 s, and 72°C for 30 s during the amplification steps. The dissociation curve was determined starting from 58°C increasing 0.3°C each 20 s until 95°C was reached. Primer pairs assessed exhibited determination coefficients above 95%, variance coefficient in two repetitions less than 10%, amplification efficiencies of 95% to 100% and slope values near to −3.0 (Applied Biosystems, USA).
The relative quantitation was performed by the comparative C T (∆∆C T) method using the Step-One Plus C T (∆∆C T) program (Applied Biosystems, USA). The cycling conditions were the same as for the relative standard curve. The experimental design consisted of two biological pools with three technical replicates for each sample: root, stem, or leaves.
Data analysis was carried out with the Step-One Plus software v. 2.0 (Applied Biosystems, USA). Threshold cycle (Ct) values for the technical replicates were calculated out and set by the relative quantitation method of copy-level amplification (Ali-Benali et al. 2005). In order to determine the relative expression of the target genes in the respective tissues, homogeneity was assessed by the Bartlett test (p = 0.05) for the three technical replicates of each biological sample. For comparative test, root sample was used. Relative EgMUR3 expression level in the analyzed tissues was assessed by comparison with the relative expression level of the housekeeping genes (Nicot et al. 2005). The expression level (R) was then determined: \( R = {{2}^{ - \Delta \Delta {\rm{Ct}}}} \), where \( ^{ - \Delta \Delta {\rm{Ct}}} = \left( {\Delta {C_{\rm{T}}}\,{\hbox{of}}\,EgMUR3} \right) - \left( {\Delta {C_{\rm{T}}}\,{\hbox{of}}\,{\hbox{the}}\,{\hbox{housekeeping}}\,{\hbox{genes}}} \right) \). Comparative analysis was performed for both constitutive genes. The variance components comprised of the results obtained from gene expression analysis in both biological replicates of each organ in comparison with both constitutive genes. The relative EgMUR3 expression in each organ was evaluated by ANOVA and the average of each treatment compared by Scott–Knot test (p = 0.05).
Cell wall analysis
For the characterization of XyG in WT, mur3 and the generated transgenic plants oligosaccharide mass profiling was performed using a purified xyloglucan-specific endo-β-1,4-glucanase (Pauly et al. 1999b) in combination with MALDI-TOF mass spectrometry (Lerouxel et al. 2002). The XyG oligosaccharides released by the digestion were assessed in a Voyager DE-Pro MALDI-TOF (Applied Biosystems, Germany) equipment using an acceleration voltage of 20,000 V and a delay time of 350 ns. Mass spectra were obtained in the positive reflection mode.
Results
Cloning and sequence analysis of EgMUR3 gene
EgMUR3 sequence (GenBank accession no. GU013763) consisted of an intronless sequence of 1,854 bp coding for a predicted protein with 617 amino acid residues with a deduced molecular mass of 70.8 kDa and an isoeletric point of 8.19. Alignment analysis revealed a striking similarity of 82% and identity of 73% over 617 amino acids between EgMUR3 and the MUR3 protein from A. thaliana (Fig. 1). Searches for catalytic domains in EgMUR3 protein were performed by comparing its amino acid sequence with others, available through the Conserved Domains Database (http://www.ncbi.nlm.nih.gov/Structure/cdd/cdd.shtml). This analysis revealed that EgMUR3 contains a pfam03016 motif from the exostosin superfamily, ubiquitous to GT47 glycosyltransferases from many organisms including microorganisms, animal, fungi, and plant. Local alignment of their predicted catalytic domains (GT47 signature motif for exostosins) revealed similarity and identity of 96% and 90%, respectively. Searches of Genolyptus database (http://www.lge.ibi.unicamp.br/eucalyptus), which contains a large collection of Eucalyptus ESTs from different organs and species, did not return any nucleotide or amino acid sequence showing significant homology to EgMUR3.

Sequence alignment of EgMUR3 and MUR3 amino acid sequences and prediction of transmembrane domain and DxD motifs in EgMUR3. Exostosin motif is underlined. The putative transmembrane domain (TMD) is boxed and encompasses amino acids 26 to 48 as determined by TMHMM 2.0. The enzyme stem rich in charged residues was localized by TopPred after TMD and before the exostosin motif. Hydrophobic cluster analysis (HCA) identified four candidate DxD motifs (the four numbered boxes along the EgMUR3 amino acid sequence relates to the encircled residues in the HCA map)
TMHMM v. 2.0, used to predict transmembrane helices in proteins, identified an N-terminal cytoplasmic tail followed by a stretch of hydrophobic residues in EgMUR3, which is consistent with a putative transmembrane domain (TMD) typical of type II glycosyltransferases (Fig. 1). In addition, TopPred analysis showed that the charge distribution along EgMUR3 is in agreement with typical type II GTs, and the most negative values for hydrophobicity observed after the TMD could correspond to the enzyme stem (not shown). Hydrophobic cluster analysis (HCA) of EgMUR3 revealed four putative DxD motifs (Fig. 1). This analysis takes into consideration the hydrophobicity of the amino acids and the folding possibilities in aqueous environment to give rise to catalytic pockets in the enzyme. These sites could potentially accept a UDP sugar, the putative donor substrate to this type of GT.
Phylogenetic analysis
Alignment of EgMUR3 with relevant GTs from the GT47 family was performed, and a phylogenetic tree with the evolutionary relationship between them is shown in Fig. 2. Family GT47 GTs are abundant in plants and exhibit a diversity of enzyme specificities, including β(1,2)-galactosyltransferase, β(1,3)-xylosyltransferase, β(1,4)-xylosyltransferase and possibly also α(1,5)-arabinosyltransferase and β(1,4)-glucuronyltransferase activities. The GT47 family might comprise genes encoding GTs involved in cell wall architecture and storage carbohydrates. Despite the abundance of GT47 family members to date, only the function of a few of those have been determined, and representative members for some subfamilies are still not functionally characterized, hampering the assignment of putative enzyme activities for related entries in the same subfamily branch. The branches comprising FRA8 and IRX10 are better characterized and contain GTs involved in xylan or pectin biosynthesis. NpGUT1 is a pectin glucuronyltransferase involved in the transfer of glucuronic acid to the rhamnogalacturonan II domain in pectin (Iwai et al. 2002), whereas FRA8 and PoGT47C are involved in xylan biosynthesis (Zhong et al. 2005). According to expression profile analysis (Aspeborg et al. 2005), poplar PttGT47A has been suggested to transfer glucuronyl side chains to rhamnogalacturonan II in ray cells or alternatively, to 4-O-methylglucuronoxylan during secondary wall thickening of poplar stem. Nevertheless, poplar PttGT47A was closely related to IRX10 and IRX10-L (Fig. 2), and in two recent studies, IRX10 and IRX10-L were shown to participate in the xylan backbone biosynthesis (Brown et al. 2009 and Wu et al. 2009). The authors showed that the double mutant irx10 irx10-l exhibits strong reduction of xylan content, polymerization, and β(1,4)-xylosyltransferase activity. Furthermore, Wu et al. 2009 demonstrated that the N-terminal portion of IRX10/IRX10-L contains a signal peptide determinant for xylan biosynthesis and that NpGUT1 lacks this N-terminal portion. Interestingly, IRX10 N-terminal portion fused to NpGUT1 conferred the ability of NpGUT1 to successfully rescue the irx10 phenotypes. Recently, F8H, another gene involved in xylan biosynthesis, has been characterized. F8H shares the same biochemical function with FRA8, as shown by complementation studies of fra8 mutant (Lee et al. 2009). A gene encoding for an alpha-1,5-arabinosyltransferase (ARAD1) of A. thaliana (Harholt et al. 2006) was also included in our analysis. EgMUR3 and ARAD1 share 20% aminoacidic identity (8e-07) and were distantly related in our phylogenetic tree. In addition to MUR3, a poplar entry (gi, 224065537) was found to be highly conserved with EgMUR3 and could potentially be a poplar xyloglucan galactosyltransferase.

Evolutionary relationship of EgMUR3 with other GT47 glycosyltransferases. Only GT47 members showing relevant homology with EgMUR3 or functionally characterized GTs were selected for the analysis. Common name was used according to the authors that described the respective gene/enzyme and gi numbers were used for the not yet characterized GT47 members. Po and At after each GT mean Poplar or A. thaliana, respectively. Common names represent: MUR3/KAM1—xyloglucan galactosyltransferase from A. thaliana; EgMUR3—xyloglucan galactosyltransferase from E. grandis; NpGUT1—pectin glucuronyltransferase from Nicotiana plumbaginifolia; PoGT47C—glycosyltransferase involved in the biosynthesis of glucuronoxylan in Populus alba × tremula; FRA8—glucuronoxylan glucuronyltransferase from A. thaliana; ARAD1—alpha-1,5-arabinosyltransferase from A. thaliana; ARAD2—ARAD1 homolog from A. thaliana; PttGT47A—glycosyltransferase possibly involved in the transference of glucuronyl side chains to 4-O-methylglucuronoxylan of secondary wall or to rhamnogalacturonan II in the ray cells; PttGT47B and PttGT47C—up-regulated during secondary wall thickening of poplar cambial region; XGD1—xylogalacturonan xylosyltransferase 1 from A. thaliana; F8H—glycosyltransferase that complements fra8 mutation in A. thaliana; IRX10-L and IRX10—involved in glucuronoxylan biosynthesis in A. thaliana
EgMUR3 expression analysis in E. urograndis
Endogenous expression of EgMUR3 was investigated in roots, expanding leaves and growing stems of 90-day-old E. urograndis plants by qPCR. EgMUR3 transcript abundance differs in the investigated organs. EgMUR3 expression is almost tenfold higher in growing stem in comparison to roots and about fivefold higher in young expanding leaves (Fig. 3).

Relative expression of EgMUR3 in roots, leaves, and stem of E. urograndis. Relative expression level of EgMUR3 normalized to root (fold-change = 1) is shown in the columns. Letters indicate statistic differences of transcript level. Statistics consisted of ANOVA and Scott–Knot test at 5% probability
Generation and identification of transformant plants
The 4.6-kb contig obtained from the sequenced subclones of the EUGR_SG_006 library showing sequence overlap to the A. thaliana MUR3 ortholog contained the complete coding sequence of the EgMUR3 gene. PCR of BAC 87E04 DNA as template and EgMUR3 specific primers with KpnI and SalI adaptamers for cloning into pBinAr MCS resulted a PCR product of approximately 1.8 kb that facilitate the construction of pBinAr-EgMUR3 plant transformation vector. This vector was successfully transformed into A. thaliana mur3 plants using floral dip method, and transformants were identified by growth on kanamycin media.
Phenotypic alterations were not observed in transgenic plants constitutively expressing EgMUR3. After 5 weeks, leaf material was collected for transgenic and cell wall analysis. Transgenic status was confirmed by RT-PCR for most of the plants selected on kanamycin media (Fig. 4a). Since MUR3 transcript is still present in mur3 plants (The mur3 mutant allele has a miss-sense mutation) and EgMUR3 shares high homology of sequence with MUR3, primers for characterization of the transformants were designed to a region of minimal alignment between EgMUR3 and MUR3. To confirm the specificity of the primers, a PCR test was performed with Col-0 cDNA preparation from young leaves as template and primers for the endogenous MUR3 or EgMUR3. MUR3 message was detected in the cDNA preparation from Col-0 leaf material only with the MUR3 specific primers, but not with EgMUR3 primers, confirming that EgMUR3 primers were specific for the detection of transgenic plants (Fig. 4b). Plants with strong and weak expression of EgMUR3 were probably identified due to positional effects of the inserted transgene (Fig. 4a).

Transcript analyses by RT-PCR of Arabidopsis lines. a EgM1 to EgM12—EgMUR3 transformant plants, BAC—DNA of BAC 85E07 was used as template (positive control), Col-0 and mur3—DNA from Col-0 and mur3 plants were used as templates (negative controls). b RT-PCR with Col-0 cDNA preparation to test the specificity of the primers used in the analysis of transgenic plants. APT+—amplification with primers for the adenine phosphoribosyltransferase 1 constitutive gene, EgMUR3—amplification using EgMUR3 primers, MUR3—amplification using A. thaliana MUR3 primers, APT−—Test for genomic DNA contamination using APT primers in a sample which reverse transcriptase was not added during cDNA synthesis
Cell wall analysis
Cell wall material was prepared from young leaves of Col-0, mur3, empty vector control plants, EgM4 (weak expressor), EgM9 and EgM10 (strong expressors). Cell wall material was digested with XEG, and the released XyG oligosaccharides (XyGOs) were analyzed by MALDI-TOF MS. MUR3 has previously been shown to add a galactosyl unit solely to the third xylosyl residue from the non-reducing end in the XXXG xyloglucan oligosaccharide core (Madson et al. 2003). Since fucosylation at this position depends on previous galactosylation, all galactosylation (XXLG) and fucosylation (XXFG and XLFG) ions are lacking in mur3 mutant (Fig. 5). In the EgM9 and EgM10 transformant plants, the peaks corresponding to m/z 1,393.2 (XXFG) and 1,555.2 (XLFG) were restored, confirming that EgMUR3 possesses a xyloglucan galactosyltransferase activity. Also, EgMUR3-expressing plants exhibited an increase in fucosylation, galactosylation, and O-acetylation (XXLG/XLXG, XXFG and XLFG) in comparison to Col-0 (Fig. 5, lower panel), suggesting that the abundance of the galactosyltransferase gene product might be limiting or that the E. grandis EgMUR3 might have different enzymatic properties. Despite the EgM4 line exhibiting kanamycin resistance and EgMUR3 expression, the XyG structure was not restored. Thus, this line cannot be considered to contain a functional MUR3.

XyG MALDI-TOF MS spectra derived from Arabidopsis leaf cell wall material of the various lines. The lower panel shows the relative abundance of XyGO ions as displayed above. Ions representing known XyGOs are labeled using the one-letter code according to Fry et al. 1993. O-acetylated residues in XyGOs in the MALDI-TOF MS spectra are underlined. Error bars represent the standard deviation from two technical and two biological replicates
Discussion
Cell wall genes from woody species have successfully complemented A. thaliana, proving to be a suitable and fast approach for investigation of gene function in tree species (Dornelas et al. 2004; Park et al. 2003; Zhou et al. 2006). In this study, transformation of Arabidopsis with a newly identified and cloned EgMUR3 gene from E. grandis permitted the successful identification of this gene as a xyloglucan galactosyltransferase.
In the A. thaliana genome, MUR3 is represented as a single-copy gene, and its non-functional redundancy is confirmed by the impact of mur3 mutation on xyloglucan structure (Reiter et al. 1997; Tamura et al. 2005). Although the subcellular localization of EgMUR3 needs to be confirmed, in silico analysis determined that it is a type II GT that carries a TMD. EgMUR3 is expected to be anchored in the Golgi, with its C-terminal portion probably facing the Golgi lumen, as is the case with other type II GTs (Fleischer 1981). EgMUR3 has a stretch of charged amino acids after the TMD that could be the “stem” segment, which is responsible for the proper positioning of the catalytic domain of anchored GTs away from the lipid bilayer in the lumen face (Breton et al. 2001). EgMUR3 belongs to the GT47 CAZy family represented by several exostosin-like glycosyltransferases. Analysis within the exostosin domain of EgMUR3 (pfam03016) revealed important putative DxD motifs that might be directly involved in the galactosyltransferase activity. Several GTs that utilize nucleotide sugars as donor substrate and sugar moieties as acceptor contain conserved DxD motifs [aspartic acid (D), any (X), aspartic acid (D)] (Breton and Imberty 1999). Variants of these motifs are EDD, EED, or [DE] × [DE], where (E) represents glutamic acid. These residues are negatively charged in the cellular environment and therefore might contact divalent cations, which are believed to coordinate the binding between substrates and enzyme, functioning as nucleotide binding domain.
Functional analysis of EgMUR3
Analysis of EgMUR3 expression in E. urograndis by qPCR revealed that EgMUR3 is mainly expressed in elongating stems and expanding leaves of young plants. In roots, transcripts were detected, but in much lower levels.
In order to investigate the ability of EgMUR3 to rescue the mur3 phenotype, an oligosaccharide mass profiling (Lerouxel et al. 2002; Obel et al. 2006) of EgMUR3-transformed lines was performed. The results obtained from MALDI-TOF MS analysis clearly confirmed that EgMUR3 gene encodes for a galactosyltransferase that recognizes acceptor substrates for the xyloglucan biosynthesis in A. thaliana and decorates the third xylosyl residue within the XXXG core structure just like MUR3. This was confirmed by the restoration of the fucosylated side chains leading to the detection of XXFG and XLFG peaks in the transgenic plants in contrast to mur3 plants. A compensation for the lack of galactosylation/fucosylation at the third xylosyl seemed to take place, as detected by the remarkable increase of XLXG abundance when one compares mur3 and Col-0 spectra. On the other hand, EgM9 and EgM10 transformants exhibited an increase of fucosylated XyGOs (XXFG and XLFG) and a decrease of XXXG in comparison to Col-0 levels. These results support the idea that XXXG is a core structure for further decorations of XyG, since the lack of galactosylation/fucosylation in mur3 leads to increased levels of XXXG and XXLG/XLXG, whereas the constitutive expression of EgMUR3 in Arabidopsis mur3 plants decreases XXXG core below to the WT levels. Additionally, EgMUR3 overexpression remarkably increases fucosylation level concomitantly with an increase of O-acetyl substitutions. This suggests that the expression level of endogenous MUR3 in Col-0 plants might be limiting with regards to XyG galactosylation and further substitutions associated with it. Alternatively, MUR3 from E. grandis might have different enzymatic properties than that of A. thaliana resulting in overgalactosylation of XyG.
Overexpression of the fucosyltransferase AtFUT1 in A. thaliana also led to a high xyloglucan O-acetylation (Perrin et al. 2003). In contrast, mutants with reduced or lack of fucose in their XyG structure (mur1 and mur2) exhibited a significant reduction of O-acetylation, suggesting that fucosyl residues on the XyG structure are the primary determinant for galactosyl O-acetylation. However, the biological role of O-acetyl-substituents on XyG or any polysaccharide remains unknown. Structurally, O-acetylation has been suggested to block enzymatic degradation of XyG (Pauly and Scheller 2000).
In A. thaliana, different MUR3 mutant alleles exert different effects on plant development (Madson et al. 2003; Reiter et al. 1997; Tamura et al. 2005; Tedman-Jones et al. 2008). Madson et al. (2003) reported that the galactosylation and consequently, fucosylation at the third xylosyl residue (XXXG) is completely abolished while the galactosylation at the second xylosyl (XLXG) is enhanced in mur3 mutants (mur3-1 and mur3-2) carrying amino acid substitutions in different positions within the exostosin domain, without major consequences for normal plant development. In contrast, Tamura et al. (2005) reported drastic effects due to the expression of a truncated form of KATAMARI1/MUR3 protein in the plant. These authors reported the loss of physical integrity of cytoskeleton elements that could be somehow connecting to the KATAMARI1/MUR3 protein. More recently, Tedman-Jones et al. (2008) isolated mur3/cie1, an Arabidopsis mutant showing constitutive defense responses in the petiole. cie1 carries two amino acids substitutions in the MUR3 gene: one at the putative exostosin domain and another at the C-terminal region.
One issue that remains to be solved in XyG biosynthesis is the precise mechanism by which sugar moieties are added to XyG. In E. grandis, for instance, the basic XyG structure is constituted of XXXG and XXXXG, but also many galactosylated XyGOs forms can be found, such as XXLG/XLXG, XLLG, XLJG, and XLXXG (Dr. Marcos S. Buckeridge, personal communication). It has yet to be determined how many glycosyltransferases are required to produce all those XyGOs or even if a single or a few xyloglucan galactosyltransferase could have their transferase activity modulated by other factors at the cell level in the context of the XyG biosynthesis.
The functional characterization of novel eucalypts genes showing temporal or tissue-specific expression offers new opportunities for studies on modification of plant metabolism. For instance, since EgMUR3 relates to primary cell wall metabolism in young expanding stem and leaves, functional studies using EgMUR3 promoter to target gene expression of candidate genes controlling plant development and other aspects of cell wall metabolism could be investigated. Biological processes such as the early patterns of lignin biosynthesis and deposition, carbon relations through alteration of the sink strength, resistance responses to plant pathogens which preferentially start invading young tissues, and metabolism of many plant hormones which control cell wall constitution and the overall plant architecture are just few examples of potential studies using the EgMUR3 promoter.
As functional genomic reports for this important forestry species become available, the understanding of how particular genes may affect plant development and biomass properties will pave the way for designing trees more suitable for pulping and bioenergy.
References
Ali-Benali MA, Alary R, Joudrier P, Gautier MF (2005) Comparative expression of five Lea Genes during wheat seed development and in response to abiotic stresses by real-time quantitative RT-PCR. Biochim Biophys Acta 1730:56–65
Aspeborg H, Schrader J, Coutinho PM, Stam M, Kallas A, Djerbi S, Nilsson P, Denman S, Amini B, Sterky F, Master E, Sandberg G, Mellerowicz E, Sundberg B, Henrissat B, Teeri TT (2005) Carbohydrate-active enzymes involved in the secondary cell wall biogenesis in hybrid aspen. Plant Physiol 137:983–997
Baucher M, Halpin C, Petit-Conil M, Boerjan W (2003) Lignin: genetic engineering and impact on pulping. Crit Rev Biochem Mol 38:305–350
Bechtold N, Pelletier G (1998) In planta Agrobacterium-mediated transformation of adult Arabidopsis thaliana plants by vacuum infiltration. Meth Mol Biol 82:259–266
Bevan M (1984) Binary Agrobacterium vectors for plant transformation. Nucl Acids Res 12:8711–8721
Boerjan W (2005) Biotechnology and the domestication of forest trees. Curr Opin Biotech 16:159–166
Breton C, Imberty A (1999) Structure/function studies of glycosyltransferases. Curr Opin Struct Biol 9:563–571
Breton C, Mucha J, Jeanneau C (2001) Structural and functional features of glycosyltransferases. Biochimie 83:713–718
Brown DM, Zhang ZN, Stephens E, Dupree P, Turner SR (2009) Characterization of IRX10 and IRX10-like reveals an essential role in glucuronoxylan biosynthesis in Arabidopsis. Plant J 57:732–746
Callebaut I, Labesse G, Durand P, Poupon A, Canard L, Chomilier J, Henrissat B, Mornon JP (1997) Deciphering protein sequence information through hydrophobic cluster analysis (HCA): current status and perspectives. Cell Mol Life Sci 53:621–645
Chen CY, Baucher M, Holst Christensen J, Boerjan W (2001) Biotechnology in trees: towards improved paper pulping by lignin engineering. Euphytica 118:185–195
Clough SJ, Bent AF (1998) Floral dip: a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J 16:735–743
de Lima DU, Buckeridge MS (2001) Interaction between cellulose and storage xyloglucans: the influence of the degree of galactosylation. Carbohydr Polym 46:157–163
Dornelas MC, do Amaral WAN, Rodriguez APM (2004) EgLFY, the Eucalyptus grandis homolog of the Arabidopsis gene LEAFY is expressed in reproductive and vegetative tissues. Braz J Plant Physiol 16:105–114
Ewing B, Green P (1998) Base-calling of automated sequencer traces using phred II. Error probabilities. Genome Res 8:186–194
Ewing B, Hillier L, Wendl MC, Green P (1998) Base-calling of automated sequencer traces using phred I. Accuracy assessment. Genome Res 8:175–185
Felsenstein J (1985) Confidence-limits on phylogenies—an approach using the bootstrap. Evolution 39:783–791
Fleischer B (1981) Orientation of glycoprotein galactosyltransferase and sialyltransferase enzymes in vesicles derived from rat-liver golgi-apparatus. J Cell Biol 89:246–255
Foucart C, Paux E, Ladouce N, San-Clemente H, Grima-Pettenati J, Sivadon P (2006) Transcript profiling of a xylem vs phloem cDNA subtractive library identifies new genes expressed during xylogenesis in Eucalyptus. New Phytol 170:739–752
Fry SC, York WS, Albersheim P, Darvill A, Hayashi T, Joseleau JP, Kato Y, Lorences EP, Maclachlan GA, Mcneil M, Mort AJ, Reid JSG, Seitz HU, Selvendran RR, Voragen AGJ, White AR (1993) An unambiguous nomenclature for xyloglucan-derived oligosaccharides. Physiol Plant 89:1–3
Gaboriaud C, Bissery V, Benchetrit T, Mornon JP (1987) Hydrophobic cluster-analysis—an efficient new way to compare and analyze amino-acid-sequences. FEBS Lett 224:149–155
Gonzalez ER, de Andrade A, Bertolo AL, Lacerda GC, Carneiro RT, Defavari VAP, Labate MTV, Labate CA (2002) Production of transgenic Eucalyptus grandis × E-urophylla using the sonication-assisted Agrobacterium transformation (SAAT) system. Funct Plant Biol 29:97–102
Gordon D, Abajian C, Green P (1998) Consed: a graphical tool for sequence finishing. Genome Res 8:195–202
Harholt J, Jensen JK, Sorensen SO, Orfila C, Pauly M, Scheller HV (2006) ARABINAN DEFICIENT 1 is a putative arabinosyltransferase involved in biosynthesis of Pectic Arabinan in Arabidopsis. Plant Physiol 140:49–58
Hoffman M, Jia ZH, Pena MJ, Cash M, Harper A, Blackburn AR, Darvill A, York WS (2005) Structural analysis of xyloglucans in the primary cell walls of plants in the subclass Asteridae. Carbohydr Res 340:1826–1840
Hofgen R, Willmitzer L (1990) Biochemical and genetic-analysis of different patatin isoforms expressed in various organs of potato (Solanum-Tuberosum). Plant Sci 66:221–230
Iwai H, Masaoka N, Ishii T, Satoh S (2002) A pectin glucuronyltransferase gene is essential for intercellular attachment in the plant meristem. Proc Natl Acad Sci U S A 99:16319–16324
Lee CH, Teng Q, Huang WL, Zhong RQ, Ye ZH (2009) The F8H glycosyltransferase is a functional paralog of fra8 involved in glucuronoxylan biosynthesis in Arabidopsis. Plant Cell Physiol 50:812–827
Lerouxel O, Choo TS, Seveno M, Usadel B, Faye L, Lerouge P, Pauly M (2002) Rapid structural phenotyping of plant cell wall mutants by enzymatic oligosaccharide fingerprinting. Plant Physiol 130:1754–1763
Levy S, Maclachlan G, Staehelin LA (1997) Xyloglucan sidechains modulate binding to cellulose during in vitro binding assays as predicted by conformational dynamics simulations. Plant J 11:373–386
McKinnon GE, Potts BM, Steane DA, Vaillancourt RE (2005) Population and phylogenetic analysis of the cinnamoyl coA reductase gene in Eucalyptus globulus (Myrtaceae). Aust J Bot 53:827–838
Madson M, Dunand C, Li XM, Verma R, Vanzin GF, Calplan J, Shoue DA, Carpita NC, Reiter WD (2003) The MUR3 gene of Arabidopsis encodes a xyloglucan galactosyltransferase that is evolutionarily related to animal exostosins. Plant Cell 15:1662–1670
Mersereau M, Pazour GJ, Das A (1990) Efficient transformation of Agrobacterium-tumefaciens by electroporation. Gene 90:149–151
Moralejo M, Rochange F, Boudet AM, Teulieres C (1998) Generation of transgenic Eucalyptus globulus plantlets through Agrobacterium tumefaciens mediated transformation. Aust J Plant Physiol 25:207–212
Navarro M, Marque G, Ayax C, Keller G, Borges JP, Marque C, Teulieres C (2009) Complementary regulation of four Eucalyptus CBF genes under various cold conditions. J Exp Bot 60:2713–2724
Nicholas KB, Nicholas HBJ, Deerfield DW (1997) GeneDoc: analysis and visualization of genetic variation. EMBNEWNEWS 4:14
Nicot N, Hausman JF, Hoffmann L, Evers D (2005) Housekeeping gene selection for real-time RT-PCR normalization in potato during biotic and abiotic stress. J Exp Bot 56:2907–2914
Obel N, Erben V, Pauly M (2006) Functional wall glycomics through oligosaccharide mass profiling. In: The science and lore of the plant cell wall: Biosynthesis, structure and function Ed. T. Hayashi. BrownWalker Press, Boca Raton, Florida, USA. pp 258–266
Park YW, Tominaga R, Sugiyama J, Furuta Y, Tanimoto E, Samejima M, Sakai F, Hayashi T (2003) Enhancement of growth by expression of poplar cellulase in Arabidopsis thaliana. Plant J 33:1099–1106
Pauly M, Scheller HV (2000) O-acetylation of plant cell wall polysaccharides: identification and partial characterization of a rhamnogalacturonan O-acetyl-transferase from potato suspension-cultured cells. Planta 210:659–667
Pauly M, Albersheim P, Darvill A, York WS (1999a) Molecular domains of the cellulose/xyloglucan network in the cell walls of higher plants. Plant J 20:629–639
Pauly M, Andersen LN, Kauppinen S, Kofod LV, York WS, Albersheim P, Darvill A (1999b) A xyloglucan-specific endo-beta-1, 4-glucanase from Aspergillus aculeatus: expression cloning in yeast, purification and characterization of the recombinant enzyme. Glycobiology 9:93–100
Paux E, Carocha V, Marques C, de Sousa AM, Borralho N, Sivadon P, Grima-Pettenati J (2005) Transcript profiling of Eucalyptus xylem genes during tension wood formation. New Phytol 167:89–100
Pena MJ, Ryden P, Madson M, Smith AC, Carpita NC (2004) The galactose residues of xyloglucan are essential to maintain mechanical strength of the primary cell walls in Arabidopsis during growth. Plant Physiol 134:443–451
Perrin RM, DeRocher AE, Bar-Peled M, Zeng W, Norambuena L, Orellana A, Raikhel NV, Keegstra K (1999) Xyloglucan fucosyltransferase, an enzyme involved in plant cell wall biosynthesis. Science 284:1976–1979
Perrin RM, Jia ZH, Wagner TA, O'Neill MA, Sarria R, York WS, Raikhel NV, Keegstra K (2003) Analysis of xyloglucan fucosylation in Arabidopsis. Plant Physiol 132:768–778
Reiter WD, Chapple C, Somerville CR (1997) Mutants of Arabidopsis thaliana with altered cell wall polysaccharide composition. Plant J 12:335–345
Saitou N, Nei M (1987) The neighbor-joining method—a new method for reconstructing phylogenetic trees. Mol Biol Evol 4:406–425
Sambrook J, Russell D (2001) Molecular cloning: a laboratory manual. Cold Spring Harbor Laboratory Press, USA
Sassaki FT, Campos-Pereira T, Maia ID (2005) The post-transcriptional gene silencing pathway in Eucalyptus. Genet Mol Biol 28:496–500
Schwarz R, Dayhoff M (1979) Matrices for detecting distant relationships. In: Atlas of protein sequences Ed. M Dayhoff, National Biomedical Research Foundation, pp 353–358
Serrano L, Rochange F, Semblat JP, Marque C, Teulieres C, Boudet AM (1996) Genetic transformation of Eucalyptus globulus through biolistics: complementary development of procedures for organogenesis from zygotic embryos and stable transformation of corresponding proliferating tissue. J Exp Bot 47:285–290
Sibout R, Baucher M, Gatineau M, Van Doorsselaere J, Mila I, Pollet B, Maba B, Pilate G, Lapieffe C, Boerjan W, Jouanin L (2002) Expression of a poplar cDNA encoding a ferulate-5-hydroxylase/coniferaldehyde 5-hydroxylase increases S lignin deposition in Arabidopsis thaliana. Plant Physiol Bioch 40:1087–1096
Somerville C, Bauer S, Brininstool G, Facette M, Hamann T, Milne J, Osborne E, Paredez A, Persson S, Raab T, Vorwerk S, Youngs H (2004) Toward a systems approach to understanding plant cell walls. Science 306:2206–2211
Sonoda T, Koita H, Nakamoto-Ohta S, Kondo K, Suezaki T, Kato T, Ishizaki Y, Nagai K, Lida N, Sato S, Umezawa T, Hibino T (2009) Increasing fiber length and growth in transgenic tobacco plants overexpressing a gene encoding the Eucalyptus camaldulensis HD-Zip class II transcription factor. Plant Biotechnol 26:115–120
Tamura K, Shimada T, Kondo M, Nishimura M, Hara-Nishimura I (2005) KATAMARI1/MURUS3 is a novel Golgi membrane protein that is required for endomembrane organization in Arabidopsis. Plant Cell 17:1764–1776
Tamura K, Dudley J, Nei M, Kumar S (2007) MEGA4: molecular evolutionary genetics analysis (MEGA) software version 4.0. Mol Biol Evol 24:1596–1599
Tedman-Jones JD, Lei R, Jay F, Fabro G, Li XM, Reiter WD, Brearley C, Jones JDG (2008) Characterization of Arabidopsis mur3 mutations that result in constitutive activation of defence in petioles, but not leaves. Plant J 56:691–703
Wang X, Tian W, Li Y (2008) Development of an efficient protocol of RNA isolation from recalcitrant tree tissues. Mol Biotechnol 38:57–64
Wilson K, Walker K (2000) Principles and techniques of practical biochemistry. Cambridge University Press, Cambridge
Wu AM, Rihouey C, Seveno M, Hornblad E, Singh SK, Matsunaga T, Ishii T, Lerouge P, Marchant A (2009) The Arabidopsis IRX10 and IRX10-LIKE glycosyltransferases are critical for glucuronoxylan biosynthesis during secondary cell wall formation. Plant J 57:718–731
Zablackis E, York WS, Pauly M, Hantus S, Reiter WD, Chapple CC, Albersheim P, Darvill A (1996) Substitution of L-fucose by L-galactose in cell walls of Arabidopsis mur1. Science 272:1808–1810
Zhong RQ, Pena MJ, Zhou GK, Nairn CJ, Wood-Jones A, Richardson EA, Morrison WH, Darvill AG, York WS, Ye ZH (2005) Arabidopsis fragile fiber8, which encodes a putative glucuronyltransferase, is essential for normal secondary wall synthesis. Plant Cell 17:3390–3408
Zhou GK, Zhong RQ, Richardson EA, Morrison WH, Nairn CJ, Wood-Jones A, Ye ZH (2006) The poplar glycosyltransferase GT47C is functionally conserved with Arabidopsis Fragile fiber8. Plant Cell Physiol 47:1229–1240
Acknowledgements
The authors would like to thank Kirk Schnorr, Novozymes, Bagsvaerd, Denmark, for the gift of XEG, Lutz Neumetzler and Katrin Geisler, Max-Planck Institute for molecular plant physiology, Golm, Germany, for technical assistance with the MALDI-TOF MS and plant transformation and Prof Giancarlo Pasquali (UFRGS, Brazil) for information concerning E. grandis constitutive genes. This work was supported by CNPq (Nacional Council for Scientific and Technological Development, Brazil; Grant 200875/2005-1) and DAAD (Deutscher Akademisher Austausch Dienst; Grant Ref. 415; PKZ A/05/50777).
Open Access
This article is distributed under the terms of the Creative Commons Attribution Noncommercial License which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by W. Boerjan
Rights and permissions
Open Access This is an open access article distributed under the terms of the Creative Commons Attribution Noncommercial License (https://creativecommons.org/licenses/by-nc/2.0), which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
About this article
Cite this article
Lopes, F.J.F., Pauly, M., Brommonshenkel, S.H. et al. The EgMUR3 xyloglucan galactosyltransferase from Eucalyptus grandis complements the mur3 cell wall phenotype in Arabidopsis thaliana . Tree Genetics & Genomes 6, 745–756 (2010). https://doi.org/10.1007/s11295-010-0288-8
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11295-010-0288-8