
Abstract
High-elevation cushion peatlands are typical ecosystems of the Andes above 4000 m of altitude, with an important role in hydrology and global carbon sequestration. In Tropical Andean context, grazing livestock is one of the main threats to cushion peatlands, altering the vegetation and the storage carbon function. The aim of this research is to understand how grazing influences cushion peatland functioning by identifying ecological thresholds for carbon balance process. The study was carried out in four Andean peatlands in the northern part of Colombian Andes during 2019–2020. We established 30 plots of 1 m2 where water table level, vegetation cover, and grazing disturbance were monitored. We also measured CO2 fluxes using an infrared gas analyzer connected to a closed static chamber, which registered net ecosystem exchange and respiration data. Considerable variation in the conservation status of Distichia muscoides cushions was found within the sampled peatlands, reflecting an heterogeneous signal of grazing disturbance that is evident at the plot-specific scale. Decreasing water table level was related with changes in dominant vegetation, from compact cushion species to grasses proliferation, exacerbating disturbance effects and carbon emissions. Mixed-effects logistic regression models showed a carbon balance shift, from CO2 sink to net emitter, in plots with high disturbance intensity and low D. muscoides cover. This study provides information for a better understanding of mountain peatlands functioning in the Tropical Andes and underlines the key role of D. muscoides cushions and the water table in carbon balance shift.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Peatlands are common ecosystem in tropical mountains, and they have one of the largest carbon density of all the ecosystems, with nearly 1000 ton C ha−1 (Hribljan et al. 2016). Peatland carbon storage requires the accumulation of partially decomposed organic matter over thousands of years, which results from the imbalance between primary production and limited soil decomposition. Human disturbances such as grazing livestock affect the stability of the water table and plant composition, which are key elements that have a direct effect on the ability of the system to accumulate peat (Belyea and Malmer 2004; Moore and Dalva 2006). Our understanding on how grazing disturbance alters carbon accumulation process in peatland ecosystems is still scarce in the tropical Andes. Better understanding of carbon balance shifts once it is pushed past a critical threshold is needed. Management strategies for Andean mountain peatlands rely on the development of indicators that help address the impacts of grazing disturbance (Chimner and Welker 2011).
Carbon storage in soils requires an organic matter input provided by litter or old roots from peat-forming plants, such as Distichia muscoides which is a peat-forming cushion plant with high net primary production rates but it’s also dependent of flooding conditions (Cooper et al. 2015). Thus, changes in vegetation composition and water table depth result in the alteration of net carbon assimilation and decomposition rates, (Clymo 1984; Turetsky et al. 2002; Belyea and Malmer 2004). The alteration of the carbon balance happens when stress conditions cause greater respiration loss than the CO2 uptake, due to peat oxidation and acceleration of decomposition rate. The change in the functioning of the system from a carbon sink to a net emitter of CO2 corresponds to a cross of an ecological threshold (Turetsky et al. 2002; Moore and Dalva 2006; Larsen and Alp 2015; Toms and Villard 2015). Although studies aimed at understanding ecological thresholds from empirical data are limited and the scope in decision-making is questioned (Dodds et al. 2010; Hillebrand et al. 2020), this approach could be useful to represent critical conditions of carbon balance shift as key indicator oriented toward management strategies to minimize adverse changes in peat accumulation process (Belyea and Malmer 2004; Johnson 2013; Toms and Villard 2015).
Even though mountain peatlands cover small areas, they have large carbon stocks in the soil due to a combination of high mineral content from the geomorphological process of the mountains and the type of vegetation that forms the peat (Hribljan et al. 2017). The steep topography of the Andes, together with a diverse and complex geomorphological setting of high-altitude lakes and glacial valleys, provides adequate conditions for the formation of wetlands and peatlands with unique vegetation (Cooper et al. 2015). At high elevations of Andes mountains (> 4000 masl), peatland vegetation grows in compact cushion of vascular plants such as D. muscoides (Juncaceae) and Plantago rigida (Sklenář and Jørgensen 1999). Since cushion species at high elevations are the main peat-forming plant and play a key role in carbon sequestration, changes in cushions dominance could be catastrophic for the functioning of mountain peatlands.
Andean mountains and in particular their alpine ecosystems are important sources of feed for native and non-native grazers, including Andean camelids at least for the last 9000 years (Browman 1984), especially in dry Puna region of the central Andes, where peatlands provide the evergreen forage for the livestock in the dry season (Maldonado Fonkén 2014). Grazing plays an important role for biomass production because of the historical relationship between native camelids and cushion peatlands (Garcia et al. 2014). However, in the northern Andes, cushion peatlands are instead affected by introduced grazers such as sheep, horses, and cattle, which also generate a disturbance on plant cover due to their heavier bodies and hard hooves (Maldonado Fonkén 2014; Machaca et al. 2018). Trampling by grazers pug peat soil, removing part of the biomass and the carbon stored in peat due to aerobic decomposition (Salvador et al. 2015).
Here, we hypothesized that human disturbances by high intensity grazing, combined with a lowering water table, could negatively affect the carbon balance of cushion peatlands dominated by D. muscoides, shifting the system from a carbon sink to a carbon source. Therefore, the purpose of this study was to assess how grazing and vegetation influence CO2 emissions in Tropical Mountain peatlands and the limit at which the system shifts from a carbon sink to a net carbon source in order to provide management tools that protect carbon storage capacity in peatland ecosystems. Our research focused on (1) comparing environmental drivers and vegetation patterns along a grazing disturbance gradient and (2) identifying response variables that affect carbon balance, using the ecological thresholds approach.
Methods
Study site
The study was conducted at the high elevations of El Cocuy National Park, in the northern Andes of Colombia (Fig. 1). A large portion of the protected area overlaps with an Indigenous territory, as well as private lands managed by farming families who frequently graze their livestock within the protected area. The area is characterized by an annual mean temperature of 6.4 °C and a bimodal precipitation regime, with two wet periods in May and October with an average rainfall of 868.4 mm (López‐Moreno et al. 2022). We randomly selected four peatlands dominated by cushions of D. muscoides, combined with graminoids such as Agrostis breviculmis and Calamagrostis effusa and the moss Breutelia chrysea (Fig. 2). All sites had more than five meters of peat, were located at a similar elevation, and had a long history of livestock use, with evidence of sheep grazing disturbance (Table 1). The peatlands were visited on five occasions during the 2019–2020 period to include the annual climatic variation.

Location of study sites in El Cocuy National Park, in the northern Andes of Colombia

Cushion peatlands of D. muscoides in the sampled sites, located in El Cocuy National Park, northern Andes of Colombia. (Top left) Chapetona; (top right): Corral Grande; (bottom left): Curial and (bottom right): Curial alto
Vegetation sampling and environmental factors
We located a total of 30 1 m2 plots randomly positioned and stratified across the main plant and ground cover gradients observed on the selected peatlands. Plant composition was sampled in all the plots by visual estimation using a 1 m2 quadrat with 10 × 10 cm sub-quadrants (Mueller-Dombois and Ellenberg 1974), where plant and ground covers were estimated to the nearest 1% for each species. From plant composition information, ground cover was classified in cushions of D. muscoides, hollows, dead cushions, or exposed peat- , grass-, and moss-dominated sites. Plant species were identified following standard herbarium identification protocols, and species names were assigned following the international plant name index (IPNI 2023. https://www.ipni.org/ accessed on February 28, 2023).
Due to the differential sheep grazing intensity within individual peatlands, disturbance was not homogenous throughout each site. Thus, disturbance was characterized at plot level and nearby area using the percentage of exposed peat cover and the evidence of sheep alteration such as excrements, hoof marks, trails, and/or herbivory. The type of specific disturbance was classified according to a semi-quantitative intensity scale from 1 to 6, where the presence of excrement represented the lowest intensity while pugged and peat implied the most severe (Online Resource: Fig. S1). The disturbance category was developed from combining exposed peat cover and the sheep alteration scale. For water table level, we installed 30 monitoring wells that were located near the 1m2 plots. Water level was measured manually during each visit to the study site. When the water table was above the peatland surface, a positive value was recorded and, if it was below the surface, a negative value was recorded. The water table level was monitored for each plot at each visit to the study site. Also, in each visit, a water sample was collected to measure pH and electrical conductivity (EC). Average monthly air temperature and precipitation were provided by the Instituto de Hidrología, Meteorología y Estudios Ambientales (IDEAM) from a meteorological station closest to the study sites.
CO2 measurements
CO2 flux was measured using an infrared gas analyzer (EGM4; PP Systems, Amesbury, USA) connected to a closed transparent static chamber of 60 cm of diameter and a height of 50 cm (Hutchinson and Mosier 1981). For each measurement, soil and air temperature, relative humidity, atmospheric pressure, and photosynthetically active radiation using a PAR sensor attached to the inside of the chamber were measured (Apogee MQ-200 Quantum meter). Using a total of 105 flux measurements with a duration of 124 s, each was performed in two consecutive phases: (1) a phase where the chamber receives natural solar radiation, registering net ecosystem exchange (NEE), and a second phase where the chamber does not receive radiation, registering autotrophic and heterotrophic respiration (RE). Gross primary production (GPP) was calculated as the difference between net ecosystem exchange and ecosystem respiration for each pair of light and dark, flux measurements.
Statistical analysis
Parametric and non-parametric analyses of variance (one way-ANOVA and Kruskal–Wallis) were performed to identify the differences between the environmental and flux variables among the sampled peatlands. Plant composition was analyzed using a cluster analysis with UPGMA method as well as analysis of similarity between sites and vegetation plots based on the Bray–Curtis dissimilarity matrix among samples. Additional MDS analysis was used to analyze the homogeneity differences in plant composition and the main environmental gradients associated with the changes in species composition. The main environmental patterns were analyzed using principal components analysis (PCA). A repeated-measures analysis with mixed effects was applied using lme4 package to quantify the relationships between explanatory variables (vegetation cover and water table) and CO2 fluxes, where plot and period were incorporated in the models as random effects, and maximum likelihood estimation was performed. CO2 fluxes was the dependent variables with GPP, NEE, and ER analyzed separately. Five NEE outliers were omitted due to equipment problems.
The relationships between vegetation and water table and NEE were analyzed using mixed-effects binomial logistic regression models to identify the ecological thresholds (McCullagh and Nelder 1989; Toms and Villard 2015). NEE was coded as a binary variable, converting negative values to values of 0 (net ecosystem uptake) and positive values to values of 1 (net ecosystem emission). The threshold was determined when the probability of the system going from sink to emitter crossed 0.5 probability (Toms and Villard 2015). The selection of the logistic models that better explained the changes in NEE was made based on the significance of the model and AIC value. Residuals were checked for normality and that homogeneity of variance assumptions was met, and significance was determined at an alpha value of 0.05. Also, average and standard error of NEE values were calculated to evaluate the carbon emissions behavior above and below the determined threshold for all the sampled plots. All data analyses were performed with the open-source software package R, version 4.1.3 2022.
Results
Vegetation patterns and environmental factors
A total of 43 species were registered, including 13 bryophytes and 30 vascular plants. Plot vegetation was dominated by D. muscoides with a mean 54% cover in all study sites (F4,130 = 35.2, p = 0.001; Table 2). Exposed peat was more common in Curial alto and Corral Grande (15 and 16% cover, respectively) and commonly associated with sedges (Carex sp.), grasses (Agrostis breviculmis), rosettes such as Werneria pygmaea, herbs that were common on disturbed non-peatland areas such as Lachemilla nivalis and Oreomyrrhis andicola, and mosses such as Bryum argenteum and Breutelia spp. Five cover types were classified in the plots: 1) D. muscoides, 2) mosses, 3) grasses, 4) exposed peat, and 5) other vascular plants (Table 2).
Water table monitoring showed that site and time of the year had a significant effect on water table depth, being Curial site the wettest, followed by Corral Grande, Chapetona, and Curial Alto the driest. Water table levels ranged from 10 to 33 cm below the soil surface and were very variable throughout the year, with a mean depth of − 19.6 ± 12.2 cm during the wet season, and -30.6 ± 10.3 cm during the dry season (January and February), when some piezometers were completely dry (Table 2). Plots with less exposed peat showed more stable water table levels throughout the year (F4,130 = 35.2, p = 0.001).
Water table showed a significant positive relationship with D. muscoides cover from the mixed model (Pseudo-R2 = 0.5, p = 0.005) and a negative relationship with grasses (Pseudo-R2 = 0.2, p = 0.06). Plots with water table above 20 cm depth had up to 50% D. muscoides cover, whereas plots with lower water table (< 20 cm from the surface) exhibited an average increase of 15% cover of grasses (Fig. 3). A negative relationship between water table depth and the percentage of exposed peat was also observed (Pseudo-R2 = 0.38, p = 0.03), except in Chapetona site, where almost no exposed peat was recorded.

Linear relationship between water table level (cm) and vegetation cover type (%) for all sampled peatlands of the Cocuy National Park. (Top): D. muscoides cover; (center): grass cover and (bottom): exposed peat cover
The ordination diagram shows that there was a plot-scale heterogeneity regardless of the peatland. Plant composition changed across the water table and disturbance gradients. Plots with higher exposed peat cover had higher dominance of species associated with drier conditions such as Agrostis breviculmis, Lachemilla nivalis, Bryum argenteum, and Oreomyrrhis andicola. In contrast, plots with higher water table level and low disturbance intensity were dominated by D. muscoides and Plantago rigida and had less exposed peat cover (Fig. 4). Exposed peat cover was strongly correlated with disturbance categories (r = 0.89) and was the environmental factor that better explained variance between plots.

NMDS Analysis for the four sites of the Cocuy National Park: Chapetona (CHA), Curial (CU), Curial grande (CUA), and Corral Grande (CO)
Carbon fluxes
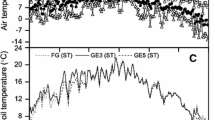
The studied peatlands had an average carbon assimilation rate of −0.04 ± 2.1 g CO2 m−2 h−1. Carbon fluxes were positively related to soil temperature, solar radiation, and water table level. GPP ranged from − 0.7 to − 0.22 g CO2 m−2 h−1 (Table 1). GPP was influenced by PAR, with a significant response function during the hours of highest radiation (around 1900 µmol m-2 s-1; Fig. 5). Carbon assimilation values decrease at low water table levels and increase with increasing water table level (Pseudo-R2 = 0.29, p = 0.04; Fig. 5). In contrast, ER responded oppositely to increasing water tables (Pseudo-R2 = 0.45, p = 0.5). Also, ER had a strong relationship with soil temperature, since ER values increase with soil warming (Pseudo-R2 = 0.35, p = 0.12; Fig. 5).

Linear relationship between net ecosystem exchange (NEE; top) and water table level (cm); gross primary production (GPP; center) versus photosynthetically active radiation (PAR; µmol m−2 s−1); and ecosystem respiration (ER; bottom) and soil temperature (°C) for all sampled peatlands of the Cocuy National Park
D. muscoides exposed peat and grass cover had a significant relationship with NEE. Carbon assimilation decrease in exposed peat-dominated plots, where positive or low NEE values were recorded (Pseudo-R2 = 0.13, p = 0.01). In contrast, plots with high D. muscoides cover had negative NEE values, which means high-carbon assimilation rate (Pseudo-R2 = 0.15, p = 0.0001). Also, high cover of grass was related with positive NEE values. The logistic regression models showed a variation on carbon balance values related to the loss of vegetation cover of the cushions at the plot scale. In both cases when D.muscoides cover decreased and when the exposed peat area increased, a threshold point was found for which there was a greater probability that respiration exceeded assimilation. Plots with less than 33% of D. muscoides cover had a high probability of a net carbon emission, reaching a 0.78 probability of being net carbon emitters when D. muscoides cover was completely lost (Fig. 6). In the same way, plots with more than 25% cover of exposed peat had also a high probability of net carbon emission (Fig. 7). Also, average and standard error of NEE values were related with changes in D. muscoides and exposed peat cover according to the NEE patterns above and below the determined thresholds.

Probability of net carbon emission according to D. muscoides cover. Red line represents the ecological threshold where there is a 50% probability of a positive carbon balance for all sampled peatlands of the Cocuy National Park

Probability of net carbon emission according to exposed peat cover. Red line represents the ecological threshold where there is a 50% probability of a positive carbon balance for all sampled peatlands of the Cocuy National Park
Discussion
The variation in species composition along the disturbance gradient suggests that the signal for grazing disturbance is heterogeneous within the peatlands, and it is evident at plot scale rather than on the whole peatland as a unit. The variation in vegetation cover along the disturbance gradient and water table fluctuation could lead different trajectories of plants composition. The relationship between low water table and high grass cover showed how graminoids proliferate at dryer areas, suppressing cushions species and favoring the proliferation of species more adapted to grazing use and drier condition such as Agrostis breviculmis, Lachemilla sps, and Oreorrhymmys andicola (Benavides et al. 2013ab; Loza Herrera et al. 2015; Greenwood et al. 2019). Water table drawdown could be also favored by trampling and grazing, by reducing cushions surface humidity (Machaca et al. 2018; Zhong et al. 2020), which could also accelerate graminoids rise and peat decomposition. Also, oral communications with local people and field observations showed that sheep did not forage Distichia sp. itself, but rather the graminoids growing over the cushions, evidencing that sheep grazing pressure only occurred in drier areas with palatable plant cover. This interaction could lead to positive feedback between graminoids proliferation favored by water table drawdown and increasing disturbance, opening new questions such as whether the presence of grasses is explained, to a greater extent, by disturbance or by the low water table.
Our results for net ecosystem uptake were in a range of − 0.001 and − 0.67 g CO2 m−2 h−1, much lower than those reported for cushion-dominated grazed peatlands in Ecuador (− 1.25 g CO2 m−2 h−1) (Sánchez Garces et al. 2017). Besides both studies being carried out at similar altitudes (> 4200 m) and hydrological conditions, the Ecuadorian peatland barely had exposed peat cover as well as non-cushion plant abundance, which could explain the carbon balance differences (Urbina and Benavides 2015; Sánchez Garces et al. 2017).Water table also showed a spatial relationship with carbon balance values, with high ER values and low positive NEE values (lower C uptake) at low water table level (Zhong et al. 2020). Water table drawdown increases the oxygen diffusion within the unsaturated peat and promotes peat oxidation by an increase in soil microorganism ER activity, producing carbon loss (Moore and Dalva 2006; Jaatinen et al. 2008; Hribljan et al. 2014).
This research allowed us to understand the influence of water table fluctuations on the instantaneous CO2 fluxes, highlighting the importance of short-term variation understanding to infer the effects of water table drawdown if it is more prolonged in time. Besides the explored methods were limited to explain the variability of peat accumulation process at large temporal scales, drier conditions could increase decomposition rate of peat and grasses proliferation, leading to a rapid loss of stored carbon and driving to an ecosystem shift from carbon sink to a net carbon emitter. Although cloud cover was not considered in the analysis, it has a significant effect on carbon balance shift, having a positive impact on CO2 assimilation rate (Planas et al. 2020). However, the one-year monitoring of CO2 fluxes expects to lead us to include annual climatic variation and rule out the effect of cloudiness over the NEE results.
With one-year observations, the logistic regression models showed a carbon unbalance related to the disturbance signal at a plot scale, from which respiration exceeded assimilation. Carbon balance shift from negative to positive NEE was observed both for the decrease in D. muscoides cover, and the increase of exposed peat cover, pointing out an ecological threshold from C sink to C source (Pelletier et al. 2015). Areas with less than 33% of D. muscoides cover and more than 25% of exposed peat cover are likely to have crossed the ecological threshold and they are losing the stored carbon, compromising the persistence of the ecosystem (Sánchez Garces et al. 2017).
For ecological thresholds models, single drivers were analyzed following the best statistical models, confounding variation for the identification of threshold responses (Simmonds et al. 2017). However, other abiotic variables could interfere, highlighting the importance of considering complementary drivers that also affect or interact with NEE such as water table level, microtopography, water chemistry, and climate variability (Belyea and Malmer 2004). Furthermore, grazing pressure combined with environmental changes could bring forward the carbon balance shift at lower disturbance intensity (Machaca et al. 2018), which could lead us to devise management strategies that delay threshold points such as develop cushion restoration and restrict livestock access at drier season.
Little is known about how ecological processes response to threshold cross or the capacity of vegetation and humidity to recover after certain disturbance intensity (Benavides et al. 2013a,b). Evidence of disturbance effect will be dependent on the spatial scale measure, where the response variable and the environmental drivers selected will determine the identification of a threshold for carbon balance shift (Belyea and Malmer 2004). Despite better understanding of the resilience of the systems is required, small spatial and temporal scale processes such as NEE could be an early-warning signal for ecosystem decline (Eppinga et al. 2009).
Conclusion
This study provides information for a better understanding of the functioning of mountain peatlands of the Tropical Andes, crucial in carbon storage and water regulation (Salvador et al. 2015), and underlines the key role of D. muscoides cushions and the water table in the peat accumulation process. Considerable variation in the conservation status of D. muscoides cushions was found within the sampled peatlands, showing an heterogenous signal of grazing disturbance and an effect on the carbon balance at microscale level. Since carbon balance determines a key function of peatlands, high-resolution variables like NEE could be a useful response variable to anticipate responses in larger-scale processes (Eppinga et al. 2009). This hypothesis needs further exploration through additional data for a better characterization of spatial patterns on carbon balance behavior. Rather than providing specific number of vegetation cover at which the system crosses non-return state, these results could guide decision-makers about consistent monitoring of specific response variables, and appropriate spatial and temporal scale at which thresholds may arise (Huggett 2005).
Data availability
The data presented in this study are available on request from the corresponding author. The species data are attached in supplementary material.
References
Belyea LR, Malmer N (2004) Carbon sequestration in peatland: patterns and mechanisms of response to climate change. Glob Change Biol 10:1043
Benavides J (2013a) Perturbaciones en las turberas de páramo: la acción del hombre y el clima. In: Cortés-Duque J, Sarmiento C (eds) Visión socio ecosistémica de los páramos y la alta montaña colombiana: memorias del proceso de definición de criterios para la delimitación de páramos. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt, Bogotá, pp 81–87
Benavides J, Vitt D, Wieder R (2013b) The influence of climate change on recent peat accumulation patterns of Distichia muscoides cushion bogs in the high-elevation tropical Andes of Colombia. J Geophys Res Biogeosci 118:1627
Browman D (1984) Pastoralism and development in high Andean arid lands. J Arid Environ 7:313–328
Chimner RA, Welker JM (2011) Influence of grazing and precipitation on ecosystem carbon cycling in a mixed-grass prairie. Pastor Res Policy Pract 1:1–15
Clymo R (1984) The limits to peat bog growth. Philos Trans R Soc Lond B Biol Sci 303:605–654
Cooper D, Kaczynski K, Slayback D, Yager K (2015) Growth and organic carbon production in peatlands dominated by Distichia muscoides, Bolivia, South America. Arct Antarct Alp Res 47:505
Dodds WK, Clements WH, Gido K, Hilderbrand RH, King RS (2010) Thresholds, breakpoints, and nonlinearity in freshwaters as related to management. J North Am Benthol Soc 29:988–997
Eppinga MB, Rietkerk M, Wassen MJ, De Ruiter PC (2009) Linking habitat modification to catastrophic shifts and vegetation patterns in bogs. Plant Ecol 200:53–68
Garcia MC, Meneses RI, Naoki K, Anthelme F (2014) Métodos para evaluar el efecto del pastoreo sobre las comunidades vegetales de bofedales. Ecol En Boliv 49:91–103
Greenwood JL, Stark LR, Chiquoine LP (2019) Effects of rate of drying, life history phase, and ecotype on the ability of the moss Bryum argenteum to survive desiccation events and the influence on conservation and selection of material for restoration. Front Ecol Evol 7:388
Hillebrand H, Donohue I, Harpole WS, Hodapp D, Kucera M, Lewandowska AM, Merder J, Montoya JM, Freund JA (2020) Thresholds for ecological responses to global change do not emerge from empirical data. Nat Ecol Evol. 4(11):1502–1509. https://doi.org/10.1038/s41559-020-1256-9
Hribljan JA, Kane ES, Pypker TG, Chimner RA (2014) The effect of long-term water table manipulations on dissolved organic carbon dynamics in a poor fen peatland. J Geophys Res Biogeosci 119:577–595
Hribljan JA, Suárez E, Heckman KA, Lilleskov EA, Chimner RA (2016) Peatland carbon stocks and accumulation rates in the Ecuadorian páramo. Wetl Ecol Manag 24:113–127
Hribljan JA, Suárez E, Bourgeau-Chavez LL, Endres SL, Lilleskov EA, Chimbolema S, Wayson C, Serocki E, Chimner RA (2017) Multidate, multisensor remote sensing reveals high density of carbon-rich mountain peatlands in the páramo of Ecuador. Glob Change Biol 23:5412–5425
Huggett A (2005) The concept and utility of ‘ecological thresholds’ in biodiversity conservation. Biol Conserv 124:301–310. https://doi.org/10.1016/j.biocon.2005.01.037
Hutchinson GL, Mosier AR (1981) Improved soil cover method for field measurement of nitrous oxide fluxes. Soil Sci Soc Am J 45:311–316. https://doi.org/10.2136/sssaj1981.03615995004500020017x
IPNI (2023) International Plant Names Index. Published on the Internet http://www.ipni.org, The Royal Botanic Gardens, Kew, Harvard University Herbaria & Libraries and Australian National Herbarium. Accessed 05 May 2023
Jaatinen K, Laiho R, Vuorenmaa A, del Castillo U, Minkkinen K, Pennanen T, Penttilä T, Fritze H (2008) Responses of aerobic microbial communities and soil respiration to water-level drawdown in a northern boreal fen. Environ Microbiol 10(2):339–353. https://doi.org/10.1111/j.1462-2920.2007.01455.x
Johnson CJ (2013) Identifying ecological thresholds for regulating human activity: Effective conservation or wishful thinking? Biol Conserv 168:57–65
Larsen S, Alp M (2015) Ecological thresholds and riparian wetlands: an overview for environmental managers. Limnology 16:1–9
López-Moreno JI, Rojas-Heredia F, Ceballos JL, Moran E, Alonso E, Vidaller I, Deschamps-Berger C, Revuelto J (2022) Recent evolution of glaciers in the Cocuy-Güican Mountains (Colombian Andes) and the hydrological implications. Land Degrad Dev 33:2606–2618
Loza Herrera S, Meneses R, Anthelme F (2015) Comunidades vegetales de los bofedales de la Cordillera Real (Bolivia) bajo el calentamiento global Plant communities of high-Andean wetlands of the Cordillera Real (Bolivia) in the face of global warming. Ecol En Boliv 50:39–56
Machaca NC, Condori B, Pardo AR et al (2018) Effects of grazing pressure on plant species composition and water presence on bofedales in the Andes mountain range of Bolivia. Mires Peat 21:1–15
Maldonado Fonkén MS (2014) An introduction to the bofedales of the Peruvian High Andes. Mires Peat 15:1–13
McCullagh P, Nelder J (1989) Binary data. In: McCullagh P, Nelder JA (eds) Generalized linear models. Springer, Boston, pp 98–148
Moore T, Dalva (Mike) M (2006) The influence of temperature and water table position on carbon dioxide and methane emissions from laboratory columns of peatland soils. J Soil Sci 44:651–664
Mueller-Dombois D, Ellenberg H (1974) Aims and methods of vegetation ecology. Wiley, Hoboken
Pelletier L, Strachan IB, Roulet NT, Garneau M, Wischnewski K (2015) Effect of open water pools on ecosystem scale surface-atmosphere carbon dioxide exchange in a boreal peatland. Biogeochemistry 124:291–304
Planas A, Chimner R, Hribljan J, Lilleskov E, Fuentealba B (2020) The effect of water table levels and short-term ditch restoration on mountain peatland carbon cycling in the Cordillera Blanca, Peru. Wetl Ecol Manag 28:51
Salvador F, Monerris J, Rochefort L (2015) Peatlands of the Peruvian Puna ecoregion: types, characteristics and disturbance. Mires Peat 15:1
Sánchez Garces M, Chimner R, Hribljan J, Lilleskov E, Suarez E (2017) Carbon dioxide and methane fluxes in grazed and undisturbed mountain peatlands in the Ecuadorian Andes. Mires Peat 19:1–18
Simmonds JS, van Rensburg BJ, Maron M (2017) Non-random patterns of vegetation clearing and potential biases in studies of habitat area effects. Landsc Ecol 32:729–743
Sklenář P, Jørgensen PM (1999) Distribution patterns of páramo plants in Ecuador. J Biogeogr 26:681–691
Toms J, Villard M-A (2015) Threshold detection: matching statistical methodology to ecological questions and conservation planning objectives. Avian Conserv Ecol. https://doi.org/10.5751/ACE-00715-100102
Turetsky M, Wieder K, Halsey L, Vitt D (2002) Current disturbance and the diminishing peatland carbon sink. Geophys Res Lett 29:21–31
Urbina J, Benavides J (2015) Simulated small scale disturbances increase decomposition rates and facilitates invasive species encroachment in a high elevation tropical Andean Peatland. Biotropica 47:143
Zhong Y, Jiang M, Middleton B (2020) Effects of water level alteration on carbon cycling in peatlands. Ecosyst Health Sustain 6:1–29
Acknowledgements
The authors thank the Laboratory of Ecosystems and Climate Change and El Cocuy National Park for logistical support. Also thank Marcos Correa, Anamaria Rozo, and Luisa Merchán for field assistance.
Funding
Open Access funding provided by Colombia Consortium. This project was partially funded by the Ditter Mosquera Grant from Pontificia Javeriana University of Bogotá. We thank El Cocuy National Park institution for allowing access, transport, and accommodation for our work at the National Park.
Author information
Authors and Affiliations
Contributions
VMA and JC contributed to study conception and methodology. VMA contributed to research design, data collection, and samples processing . VMA and JCB contributed to formal analysis and data curation. VMA contributed to writing-original draft preparation. JCB contributed to writing—review and supervision. VMA and JCB contributed to project administration and funding acquisition. All authors reviewed the manuscript and approved the submitted version.
Corresponding author
Ethics declarations
Competing interests
The authors declare that the research was conducted in the absence of any commercial or financial interest to disclose.
Additional information
Communicated by Paul Ramsay.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Martínez-Amigo, V., Benavides, J.C. Carbon balance shift in mountain peatlands along a gradient of grazing disturbance in the tropical Andes (Colombia). Plant Ecol 224, 1049–1058 (2023). https://doi.org/10.1007/s11258-023-01356-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11258-023-01356-8