
Abstract
Fire-killed serotinous trees are often dominant species in fire-prone regions with contrasting wet and dry seasons. We studied a serotinous fire-killed conifer (Callitris preissii, Cupressaceae) to identify the influence of fire regime and climate on geographic variation in level of serotiny, recruitment and population structure. We measured population size distributions, seedling recruitment and level of serotiny for sample stands across a climatic gradient from mesic coastal (including two islands) to semi-arid inland sites in SW Australia. Trees on islands were mostly non-serotinous and were only weakly serotinous on the adjacent mainland in the higher rainfall West coast region. In the semi-arid Goldfields and South inland, and the higher rainfall South coast region, trees were more strongly serotinous with up to six cohorts of closed cones retained on trees. Level of serotiny was stronger at the drier end of the climatic gradient where severe to extreme fire danger weather occurs most frequently, and where vegetation type (e.g., shrublands) supports frequent crown-type, stand-replacing fires. Recruitment was most abundant post-fire in all regions, but seedlings also established inter-fire where there was regular seed fall due to low level of serotiny or spontaneous release of seeds from old cones on trees with a high cone load. Population structures for mainland stands shifted from approximately reverse-J (indicating continuous recruitment) in mesic regions to mainly unimodal (establishment mostly after fire) in drier, inland regions, although some long-unburned inland stands showed occasional inter-fire recruitment, likely linked to high rainfall years. Unimodal population structures characterized islands, where previous studies have identified herbivory on seedlings by refugial populations of native macropods as a limiting factor on inter-fire establishment. Understanding landscape context, response to disturbance, biotic interactions and reproductive traits of dominant species is essential in conservation planning in a rapidly changing global environment.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The population dynamic behaviour of species, especially in fire-prone systems, is closely linked to how and when seeds are stored and dispersed relative to the occurrence of conditions favourable for recruitment. In environments where disturbances are episodic seed germination may be cued to disturbance. In fire-prone vegetation, heat and smoke act as cues for germination in many species with soil-stored seeds, or flames kill the above ground part of plants and facilitate release of seeds from canopy seed stores of serotinous species. Seeds of serotinous species must germinate after release as soon as there are suitable temperature and moisture conditions, or they will perish (Lamont et al. 1991). For this reason, serotiny tends to be associated with environments characterized by predictable, recurrent fire intervals, while soil seed stores may predominate in less predictable fire interval environments (Pausas and Keeley 2014).
For widely distributed species serotiny may vary along a continuum from weak to strong and is often associated with environmental gradients, particularly relating to rainfall and fire regime (Cowling and Lamont 1985; Nathan et al.1999; Hernández-Serrano et al. 2013; Ladd et al. 2013). For example, in some Banksia species virtually all seeds are retained indefinitely in serotinous cones until released by fire (e.g., B. cuneata—Lamont et al. 1991), while in others, such as B. attenuata, populations range from strongly serotinous in more fire-prone and less mesic areas to non-serotinous in less fire-prone and more mesic areas (Cowling and Lamont 1985). In the tropical north and arid centre of Australia, Callitris (native cypress pine) species are not serotinous, (Prior et al. 2011; Trauernicht et al. 2012), but in the temperate south most species retain seeds in woody cones for more than one year and range from weakly to strongly serotinous (Ladd et al. 2013 but see Adams 1999). For example, C. verrucosa populations in at least some parts of that species’ range are strongly serotinous and seedling establishment is tightly keyed to fire (Bradstock and Cohn 2002), while in the weakly serotinous C. glaucophylla recruitment is influenced by the interplay of fire regime and episodic periods of above average rainfall (Prior et al. 2011). In a study of serotiny in Pinus pinaster and P. halepensis in Spain, Hernandez-Serrano et al. (2013) found that populations in areas with a high likelihood of crown fires were more strongly serotinous than populations in locations that experienced mainly ground fires. In many Australian genera (e.g., in Banksia, Eucalyptus and Allocasuarina) serotiny is present but weak in most species and there is a regular turnover of closed fruits/cones with only the most recent 1–3 years of fruit/cone crops retaining seeds. Weak serotiny can operate as a ‘bet-hedging’ strategy with inter-fire establishment from a low but regular (annual) seed rain occasionally possible, and able to buffer the population against decline in association with occasionally long inter-fire intervals which exceed average plant longevity (Enright et al. 1998; Maher et al. 2010). Even in most strongly serotinous species (i.e., where seeds are held in closed cones/fruits within the plant canopy for many years and released en masse typically as a result of fire) seeds are eventually shed from older infructescences and can lead to recruitment if moisture conditions are favourable (Schmidberger and Ladd 2020).
Species characterized by strong serotiny should show post-fire cohort recruitment with little recruitment during the inter-fire period (Enright et al. 1998). Stands establishing under such circumstances are even-aged and have unimodal stem size distributions (Keeley et al. 2006). For weakly serotinous species, while post-fire recruitment is still the dominant regeneration mode, the opportunity for inter-fire seedling recruitment increases as the amount of inter-fire seed release increases, and may lead to stands with multi-modal to continuous age and size distributions. Multi-modal to continuous recruitment should become increasingly apparent for old (long unburned) stands as the number of potential recruitment years, and number of seeds released per year, increases. For example, in SE Australian mixed Callitris—Eucalyptus woodlands, C. rhomboidea recruits immediately after fire from a serotinous seed bank, establishing an even-aged cohort. However, once individuals are about 15 years of age, seed fall from ageing cones increases and stands begin to show inter-fire recruitment (Enright et al. 1994).
Callitris is a conifer genus (family Cupressaceae) of predominantly fire-killed tree species that is widely distributed across Australia from tropical to temperate latitudes. While trees of most species are killed by fire, some can survive mild burns (Bradstock and Cohn 2002; Trauernicht et al. 2012). If fires are more frequent than the time taken to reach reproductive age, a seeder species population may face local extirpation due to immaturity risk (Keeley et al. 1999). Alternatively, if fire interval is unusually long, dense post-fire stands may become “locked” (Banks and Pulsford 2001) so that tree growth slows and individuals produce few cones. When a fire eventually occurs in such stands the serotinous seedbank may be depauperate and recruitment limited, as described by Bond et al. (1984) for a South African Protea species.
Callitris preissii Miq. is a small (to 8 m height) fire-killed tree that has a broad but discontinuous distribution in southern Western Australia (WA), occurring on the west and south coasts under a warm Mediterranean winter rainfall climate, and inland under a semi-arid non-seasonal rainfall regime (Table 1). On the west coast the species may form low, closed forests but in arid areas is generally more sparsely distributed in open mallee (Eucalyptus spp.) woodlands and shrublands. The species distribution is disjunct, with the west coast populations separated from inland and south coast populations by many hundreds of kilometers and more than 500 mm difference (240–775 mm) in annual rainfall. As in all the Cupressaceae, seeds are formed in cones, with 6–16 viable seeds cone−1 reported by Ladd et al. (2013) for stands from semi-arid inland WA. While fire is clearly a very important driver of Callitris ecology, fire regimes (especially fire type and frequency), along with climatic conditions (Table 1) and biotic interactions (e.g., herbivory) vary across the geographic range of the species (e.g., Cohn et al. 2013) and may have different effects on recruitment as has been shown for mallee shrublands in semi-arid SE Australia (Kenny et al. 2018). Since the 1990s a positive phase of the Southern Annular Mode has correlated with increased summer precipitation in inland southern WA but decreased precipitation in coastal areas (O’Donnell et al. 2015). This can lead to increased likelihood of fire in both areas due to fuel build up in inland WA from increased summer rainfall in some years and drier conditions near the coast resulting in more severe fires once they are ignited. Understanding the influence of degree of serotiny on resulting tree populations is useful to predict population viability in the inevitable event of fires under a changing climatic regime.
In this study we examined level of serotiny and population structures across the C. preissii rainfall—geographic range in South-Western (SW) Australia. We quantified recruitment of seedlings after recent fire at sites spanning the rainfall and forest fire danger index (FFDI) days gradient occupied by the species. We examine if level of serotiny, population structure and recruitment behaviour changes predictably along fire regime and rainfall gradients and explains observed stand size/age structures.
We hypothesized that;
-
1.
Level of serotiny increases as the probability of stand-replacing fire increases, and so should be weakest for island populations (mesic climate, low surface fuel loads, low probability of ignitions), and strongest for mainland populations in vegetation types conducive to crown fire propagation and characterized by severe fire danger weather (semi-arid climate, severe FFDI days more frequent).
-
2.
Seedling recruitment is more continuous and abundant (and stand density greater) in more mesic areas,
-
3.
Following from the above, population structures are related to level of serotiny and climate—fire regime region, and are -
-
a.
unimodal in the most fire-prone areas where fires are frequent and seedlings recruit only after fire,
-
b.
continuous in areas where fires are less frequent and inter-fire recruitment may be common due to high levels of annual seed fall (individuals only weakly serotinous).
-
a.
Methods
Study sites
Thirty-two Callitris stands were assessed across five regions in SW Australia (Fig. 1), to encompass the range of population structures and geographic distribution of the species: Islands (8 stands), West coast mainland (10 stands), Goldfields (7 stands), South inland (5 stands) and South coast (2 stands). Each stand sampled a different population in terms of time since last fire and/or was separated from other stands by differing vegetation types. This spanned the range 31°11ʹ to 34° 54ʹ S (~ 400 km) and 115°40ʹ to 121°07ʹ E (~ 550 km). Measurements were made in one plot per stand and due to variation in tree density plot size was variable (minimum 200 m2, maximum 0.54 ha) such that from 30 to 200 individuals were sampled per plot (details in Supplementary Table 1). Mean annual rainfall was highest on the West and South coasts (675–775 mm year−1), decreasing with distance eastwards and inland to a minimum of ~ 240 mm year−1 in the Goldfields region (Fig. 1). Sites on the West and South coasts included islands (Garden Island and Bald Island, respectively) where native macropod herbivores, Macropus eugenii, (Tammar Wallaby, Garden Island) and Setonix brachyurus (Quokka, Bald Island), now rare on the mainland, still occur in abundance. Here, we combine data for island stands as a fifth region type separate from adjacent mainland stands due to their very different site characteristics. Full site details are provided in Supplementary Table 1.

Location of regions in SW Australia from which sites in this study were sampled, showing also rainfall isohyets (mm year−1; green–blue lines) and number of forest fire danger Index (FFDI) days year−1 > 50 (red lines, number of days in black). Rainfall data are from the Australian Bureau of Meteorology for the period 1987–2016 inclusive. FFDI days data are from Bannister and Hanstrum (1995)
Fire histories for most sites are unknown. However, three of the Island sample stands regenerated from seed after a large wildfire on Garden Island in 1956 (McArthur 1998), one Goldfields stand (site 6) in 1973 (fire history was verified using publicly available fire history data curated by WA state government, DCBA 2021) and one West Coast stand (at Woodman Point) after a fire of limited extent in 1992 (PGL, personal observation). Ring counts of fire-killed stems at South Coast site 2 ranged from 30 to 42 (stem basal diameters 2.3–8.2 cm) and at Goldfields site 2 from 60 to 68 (basal diameters 4.6–7.5 cm) indicating that at these sites trees established within a 10-year period. These counts indicate that establishment of the majority of the trees was likely to have occurred immediately following a single fire event at each site.
The average number of severe (or higher) Forest Fire Danger Index (FFDI > 50) weather days increases from < 1 day year−1 on the West coast and Islands to from 3 to 7 days year−1 in the South inland, Goldfields and South coast regions (Fig. 1) (Bannister and Hanstrum 1995; Enright and Fontaine 2014), and fire season increases in duration along the same gradient (Walker 1981).
Level of serotiny
Development of Callitris preissii cones has not been specifically studied but in related species cones take two years to mature (Baird 1937) and if non-serotinous will open when mature. Only fully formed and mature cones (based on their dryness and brown/grey colour) were recorded here. Green, immature cones were ignored. The level of serotiny in each plot was assessed by recording the oldest cohort of closed cones on each mature, live tree in the plot. Binoculars were used to observe cones on high branches. Callitris produces annual cone crops along branches in most but not all years, so it is not possible to age cones precisely. Therefore, closed cones were assigned to approximate age cohorts from I (youngest mature cones, located towards the end of the branch; cone age > 1 but < 2 years) to VI (oldest, towards the base of the branch; cone age ≥ 6 years). No closed cones above cohort VI were found on any plants. We categorize level of serotiny as absent to very weak where cones on the majority of trees in a stand are all open, as weak where the majority of trees hold a mix of open and cohort I closed cones, and moderate to strong where the majority of trees hold closed cones in age cohorts II to VI. The serotiny estimate for each region is the average of the proportions of the cohort data from each stand. Significance of difference in level of serotiny between regions was determined based on the cumulative relative frequency distributions using the Kolmogorov–Smirnov test for two independent samples (Norcliffe 1977).
Population size-class structures
Stem diameter was measured at the tree base below where branching occurred for all live and dead standing individuals. Plants under 2 m tall with stem diameter less than 1 cm and no cones were classed as saplings. While there is sometimes a poor relationship between stem diameter and age in C. preissii (O’Donnell et al. 2010), size-class structures are generally useful for inferring population dynamics and have been used for other Callitris species (Prior et al. 2011).
Seedling recruitment after fire
Quantification of C. preissii seedling recruitment after fire was possible for stands in three regions where fires had occurred in the recent past:
-
1.
Woodman Point (West coast)—burned in February 2016.
-
2.
Cape Le Grand National Park (South coast 2)—burned in May 2012.
-
3.
Boorabbin National Park (Goldfields site 2)—burned in December 2007.
At Woodman Point, seedling recruitment (Autumn 2016), and survival (Winter 2017), was assessed in 24 × 0.25 m2 subplots randomly located on a rectangular grid within a 30 × 10 m plot. At Cape Le Grand, juvenile plant density was determined for a single 30 × 30 m plot two years after the fire. At Boorabbin, juvenile plant (age 6 years) density was assessed in nine replicate 10 × 10 m subplots within a 30 × 30 m plot in 2014.
Inter-fire seed rain
Annual seed rain was assessed over two years (September 2014 to August 2016) in mature C. preissii stands on Garden Island (West coast). Seed traps (26 cm diameter mesh-lined buckets held 1 m above ground) were placed in 10 locations under the canopy of mature trees in August 2014. Traps were emptied, and seeds counted and assessed for viability by a cut test, monthly for 2 years.
Results
Level of serotiny
Serotiny was absent to very weak on Islands, was weak for mesic West coast stands, and moderate to strong for semi-arid Goldfields, South inland and South coast stands (Fig. 2). Island (n = 8) stands were characterized by a high proportion of trees with all mature cones open (i.e., they were not serotinous). West coast mainland stands (n = 9) overall displayed weak serotiny, with most cones either open or in age cohort I, but a small proportion of trees (< 10%) had closed cones in older cohorts. Goldfields (n = 6), South inland (n = 5) and South coast (n = 2) stands were all characterized by an increasing proportion of trees with closed cones in cohorts II to VI, and few (< 10%) non-serotinous trees.

Mean proportion of trees showing level of serotiny based on oldest cone cohort (no closed cones—Cohort VI) represented on each tree from sample stands by region (Garden and Bald Islands shown separately; see text) across the range of Callitris preissii in SW Australia from the west coast with < 1 FFDI day year−1 > 50 to the Goldfields with 5–7 FFDI days year−1 > 50. Error bars are 95% confidence intervals. Regions are ordered from least (left) to most (right) serotinous
Stand level serotiny was significantly different between all regions except Goldfields, South Inland and South Coast: The majority of trees on Islands showed no serotiny but the proportion of trees with only open cones was higher on Garden Island (0.98) than on Bald Island (0.67) (D = 0.304, P < 0.001). West Coast stands showed a lower proportion of trees with all cones open than those on Islands (D = 0.230, P < 0.01), but higher than for the Goldfields (D = 0.371, P < 0.001) and other mainland regions. Goldfields, South Inland and South Coast regions all showed a low proportion of trees with no closed cones (0.06 to 0.10).
Population structures
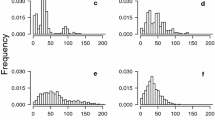
Mean stem diameter size distributions in most regions were unimodal (and ~ normal), with few individuals < 2.5 cm diameter, except for the West Coast (mesic climate) region which showed strong evidence of inter-fire recruitment, (Fig. 3). Four populations from the Goldfields showed strong unimodal size distributions while two contained sapling-sized individuals and more multi-modal to reverse-J distributions: In the long unburnt open woodlands site 5, 34% of plants were saplings all located under and around large live or dead (not fire-killed) trees with many open cones, indicating inter-fire recruitment (Fig. 4). While at site 6, burnt in 1973, 7% of plants were saplings, these were suppressed individuals in a very dense post-fire Callitris stand characterized by a high proportion of small stems.

Mean size-class structures for Callitris preissii sample stands, grouped by region/site type. Saps saplings less than 2 m tall and 1 cm stem diameter. Error bars are 95% confidence intervals

Comparison of population structures at two Goldfields sites showing the structural differences—continuous (although slow) recruitment for Goldfields site 5 (long unburnt) and cohort regeneration after a fire in 1973 (Goldfields site 6)
Modal stem diameter was either 2.5–5 or 5–10 cm for all regions other than Islands, where it was much larger (15–20 cm). Maximum stem size also varied among regions, being larger in the Islands and West coast regions (diameters up to ~ 60 cm) than in the Goldfields, South inland and South coast regions where maximum tree diameter was around 30 cm.
Average stand basal area was highest at the wetter, and lowest at the drier end of the rainfall gradient (Fig. 5). It ranged from 35 m2 ha−1 on Garden Island for a closed forest stand initiated after a fire in 1956, to 0.47 m2 ha−1 for a heathy open woodland stand at a site in the Goldfields region. However, low basal area occurred also for a few young post-fire stands in wetter regions (e.g., a ~ 25 year old stand at Woodman Point in the West coast region; see Supplementary Table 1). The density and basal area of standing dead trees was also greater at the wetter end of the rainfall gradient (i.e., dense stands showed evidence of self-thinning), with few or no dead trees in the more open semi-arid area stands.

Basal area (BA) of live and dead Callitris preissii at the study sites, ordered by region/site type as described in the text. Error bars are 95% confidence intervals
Seedling recruitment after fire
Pre-fire tree densities at the three post-burn sites ranged from 544 ha−1 in a semi-arid open woodland stand in the Goldfields, to 933 ha−1 at Woodman Point on the west coast and 2789 ha−1 in a stand at Cape le Grand on the South coast (Table 2) that was only partially burnt. Post-fire seedling densities in the Goldfields and Woodman Point sites were above pre-fire mature stand densities but were lower at Cape le Grand at the time of assessment. At Woodman Point (West coast, highest rainfall), wildfire burned a small area in summer 2016, killing all C. preissii trees, with seedling recruitment commencing in the following winter. Seedling establishment was highly variable spatially (72.2 ± CI 42.5 m−2), with highest seedling densities located in shallow (micro-topographic) depressions between dead parent trees. By the end of the first summer after fire (8 months after seeds germinated) only 22% of the seedlings were still alive (15.6 ± 20.0 m−2, Table 1). At Cape Le Grand (South coast, mesic) a patchy fire burned part of a larger sampled stand, killing all trees in the burn area. Seedling recruitment was two orders of magnitude lower than at Woodman Point (0.19 m−2) and was the lowest overall among the three burn sites. At Boorabbin (Goldfields, lowest rainfall) the fire was patchy but extensive, with high variation in post-fire recruitment and juvenile plant density (0.91 ± 0.51 m−2) seven years after fire. In all three cases, seedlings were only found in areas that had been burnt, and there was no evidence of herbivory on seedlings.
Inter-fire seed release
Very weak to no serotiny in stands on Garden Island meant that the annual seed crop was nearly all released each year and the seasonal pattern of seed release could be clearly identified. Seed release occurred predominantly from January to June, with a peak in January–February (Fig. 6). Putatively fertile (seeds with plump embryo) seed rain ranged from 2 to 30 seeds m−2 month−1 with total mean annual seed fall in excess of 100 seeds m−2.

Mean monthly fertile seed rain for Callitris preissii from September 2014 to August 2016 (averaged over two years) from 10 seed rain traps on Garden Island. Error bars are 95% confidence intervals
Discussion
In this study we found a gradient of serotiny in the fire-killed conifer tree species, C. preissii, from absent (cones on nearly all trees opening each year upon ripening) to weak (cones on most trees closed for only 1–2 years) to moderate—strong (cones on most trees closed for 2–6 years) across the species’ geographic range. However, stand size structures and imputed population dynamics did not always conform with the hypothesized patterns, with other factors, including rainfall, fuel and ignition availability, herbivory and fire patchiness likely implicated in a fuller understanding of this species’ demographic behaviour.
Callitris is loosely considered a serotinous genus (e.g., O’Donnell et al. 2010) but there is considerable variation in degree of serotiny, both between, and within species (Ladd et al. 2013; Crisp et al. 2019). As hypothesized, level of serotiny in C. preissii increased with increasing occurrence of extreme fire danger weather conditions across the regions sampled here. Recent analyses confirm this pattern based on a range of other measures of fire hazard: Kala et al. (2020) report the annual sum of daily FFDI increasing from ~ 2000 for the Islands and West coast regions, to 3000–4000 for South inland and South coast, and up to 5000 for Goldfields, while Dowdy (2018) provide similar results for 1-, 5- and 10-year return periods for extreme FFDI days for climate data spanning 1950–2016.
However, the FFDI is calculated solely from current and antecedent weather variables (Luke and McArthur 1978) and does not include other important drivers of fire regime such as frequency of natural ignitions, and fuel type/load and continuity. The annual pattern of ground flash lightning strikes in SW Australia parallels that for extreme FFDI days, with lightning strikes more than twice as frequent in the Goldfields and South inland and coast areas (1–2 km−2 year−1) as on the West coast and Islands (0.5–1 km−2 year−1) (Dowdy and Kuleshov 2014). Fuel types and continuity are also important in determining fire severity and vary with rainfall from high density woodlands—forests on the West coast and Islands (rainfall 675–775 mm year−1), to closed canopy shrublands on the South coast (rainfall 626 mm year−1), and open Callitris—mallee (Eucalyptus spp.) woodlands with (often discontinuous) grassy or heathy surface fuels in the Goldfields (rainfall 240 mm year−1) and South inland (330 mm year−1, Fig. 1) (Bradstock and Cohn 2012; Keith et al. 2012). The combination of fire weather, ignitions and fuel type (load and continuity) should thus support crown-type fire most frequently in the South coast region, where all conditions for fire occurrence are regularly available, followed by the Goldfields and South inland regions where low fuel continuity may limit fire in some years, but where grassy fuels can raise fuel continuity quickly after good rains (Table 2). Fire should occur least frequently on islands despite high fuel loads since likelihood of ignition is low (low lightning strike rate and no human occupation prior to European settlement) and fire weather conditions are rarely extreme. Observed levels of serotiny in C. preissii conform with these expectations and support our first hypothesis.
Where fires are infrequent some pines show only weak serotiny or have dispensed with it altogether (Schwilk and Ackerly 2001). This empirical observation is supported by modelling (Tonnabell et al. 2012) which showed that low predictability of fires could lead to weak serotiny as an evolutionary outcome. Nevertheless, weak serotiny can still be highly advantageous, with even 1–2 years of canopy storage sufficient to ensure massive post-fire recruitment (Enright et al. 1998). Continental shelf islands off the south and west coasts of Australia (such as those studied here) were unoccupied at European contact, and likely for at least 6–8000 years (time since post-glacial separation; Backhouse 1993), so that over this time-frame fires would have been infrequent, starting only from lightning ignitions on the islands themselves. C. preissii stands there would normally have the opportunity to grow old, characterized by large trees with high annual cone loads that could provide a massive seed rain after infrequent crown fires. Thus, the weak to absent serotiny on islands is not due to a total absence of crown fires but rather to their very infrequent occurrence. These would occur typically in mid-late summer when cones were mature, so an ephemeral seedbank would be present at that time (as shown by our seed rain results). With long fire free intervals there is also the possibility of inter-fire recruitment if seeds are shed annually, as occurs in other Callitris species from tropical north and arid central Australia where the species are non-serotinous (Ladd et al. 2013). Seed rain was only quantified on Garden Island due to inland populations being too distant to allow regular monitoring of seed shed. It would have been useful to have both inter-fire and after-fire seed rain data from inland populations but this was not logistically possible, especially after fire, as seed shed occurs within a few days. In populations with stronger serotiny annual seed rain will be minimal in young stands but will increase as trees grow and cone cohorts age, lose water connection to the tree and open to release seeds. That inter-fire recruitment had occurred in the most arid areas (Goldfields site 5) indicates inter-fire seed can enable plants to establish.
Population structures have been used in a number of studies to help understand the dynamics of tree populations (O’Connor and Goodall 2017) and the ecological processes driving them (Peck et al. 2014). In many such studies age structure is preferred, but in species that establish after a severe disturbance almost all plants are the same age. The size-class pattern will indicate if the population is healthy with a range of tree sizes or is “locked” where intraspecific competition limits plant growth and often reproductive potential (Bond et al. 1984) indicating a population that could be improved by thinning intervention. In terms of reproductive potential, it is the size of the individual rather than its age that is important. Although it was not always possible to know when plants established in the populations examined here, the relative proportion of different-sized individuals is likely to have remained consistent as the populations aged. Population structures, along with ring counts, have been used elsewhere in the study of Callitris ecology (Clayton-Greene and Ashton 1990; Bradstock and Cohn 2002; Prior et al. 2011, 2018). Most studies have been on C. glaucophylla, C. columellaris and C. intratropica, all species of tropical northern, or arid central Australia, that are non-serotinous. In some cases, the population structures showed a reverse-J pattern and in other cases structures were unimodal, with recruitment attributed to sporadic high rainfall years (Read 1995). C. preissii structures varied from unimodal, with plants mostly established immediately after disturbance by fire, to reverse-J, indicating more continuous recruitment. We hypothesized that continuous recruitment would occur in the higher rainfall areas while recruitment in arid areas would be episodic due to limited suitable recruitment opportunities (e.g., following wet years and/or fires) and could give rise to unimodal or multi-modal population structures depending on the type of establishment opportunity. For example, Cohn et al. (2013) found that stand structure for C. glaucophylla in semi-arid SE Australia depended on both rainfall and herbivory, with recruitment limited in dry years and in the presence of browsing impacts from rabbits but favoured in wetter years and in the absence of rabbits.
In accordance with our second and third hypotheses, some stands in the mesic West coast region showed evidence of continuous recruitment, while most stands in the more fire-prone semi-arid Goldfields and South inland regions showed unimodal distributions characteristic of post-fire episodic recruitment dynamics. However, Island and South coast populations, both in higher rainfall (mesic) regions also showed unimodal size distributions and so did not conform with these hypotheses. The reasons for absence of continuous recruitment in two of the three mesic region stands can be explained by differences in level of serotiny (and of associated fire regime) and herbivory. South coast stands occur in a region with moderate rainfall (~ 625 mm year−1) and support a dense shrubland. These are highly flammable (Prober et al. 2016), are able to carry crown-type fires at short fire intervals (Keith et al. 2012) and occur in a region with high fire danger weather (Bannister and Hanstrum 1995) and moderate frequency of lightning strikes (Table 1), so that all of the drivers for fire are regularly available. Stands here are characterized by strong serotiny and so also by cohort recruitment following stand-replacing fires. Nevertheless, in some landscape positions such as around rocky outcrops, fires may be discontinuous and leave patches of unburnt vegetation which can act as local refugia (e.g., South Coast site 2; see Supplementary Table 1).
Island stands also show a unimodal size structure, however, winter rainfall is reliable and serotiny is weak, so that continuous recruitment might be expected. Here, herbivory best explains the initial development of a unimodal (post-fire cohort) population structure and this is maintained by herbivory and low light levels in dense stands as trees mature. Recruitment of Callitris has been reported elsewhere to be affected by grazing animals, both native, and introduced (kangaroos, wallabies, rabbits, deer; Austin and Williams 1988; Mackenzie and Keith 2009; Cohn et al. 2013; Prior et al. 2018). The islands off the coast of southern Western Australia were isolated from the mainland by rising sea level in the early to mid Holocene (Backhouse 1993) and were unoccupied by humans until the arrival of Europeans ~ 200 years ago since there was no reliable source of fresh water. In the absence of people, fires would result only from infrequent lightning strikes in association with suitable fire weather and fuel moisture/load conditions. After European settlement fires were much more frequent. While occasional fires in the 1800s were recorded from Garden Island (McArthur 1998) most of the current Callitris forest there is the result of recruitment after a severe and extensive fire in the summer of 1956 which affected more than half of the island (Baird 1958). McArthur (1998) examined recruitment after a high severity fire of more limited extent on the island in 1991, reporting that Callitris seedlings established and survived in animal exclosures, but not outside the exclosures where grazing by the Tammar Wallaby removed all seedlings. These results are also supported by reported Callitris recruitment failure on the neighbouring Rottnest Island due to herbivory by another still abundant native macropod, the quokka (Setonix brachyurus) (Rippey and Hobbs 2003). In mesic areas, while there is likely sufficient annual seedfall and winter rainfall in most years to support recruitment of new individuals, macropod browsing precludes any such recruitment except after large fires which produce high seedfall over a large area. Animal impact is then dissipated such that some seedlings have time to establish and grow to a size where they are no longer susceptible to grazer-induced mortality. At three of the west coast mainland sites where there are currently no herbivores, seedlings and saplings are present.
In semi-arid inland areas fire frequency is related to vegetation type and fuel connectivity (O’Donnell et al. 2011b; Gosper et al. 2018). In the semi-arid parts of its range C. preissii most commonly establishes after fire, aided by its moderate—strong serotiny. Fires most frequently occur in a dry year that follows one or more years of above average rainfall that increases the surface (grassy) fuel load (Turner et al. 2008; O’Donnell et al. 2011a), but fires may still be patchy due to discontinuity of fuels. This was the case for our Goldfields site 2 fire in 2007 and the Cape Le Grand (South coast 2) fire in 2012. Seedling establishment in semi-arid areas is also strongly related to moisture availability (Elliott et al. 2019). The advantage of serotiny is that there is mass seed release after fire so there are many chances for some seedlings to survive out of many germinants. However, moisture conditions still need to be suitable to encourage germination and sustain new recruits. Prior et al. (2018) determined that for successful recruitment of Callitris glaucophylla in arid inland Australia there needed to be between 600 and 720 mm of rain over the 2 years after seed release. It is likely that similar rainfall is necessary for C. preissii to recruit seedlings, as seeds are of similar size (Ladd unpublished). For the West and South coast sites where we assessed seedling recruitment, such rainfall would occur in most years. In contrast, for the Goldfields this is unusual, but for the two years following the December 2007 Boorabbin fire more than 600 mm of rainfall was received (nearest BOM station—Dedari, 2008–2009, BOM 2018) and recruitment was abundant. Like C. glaucophylla, C. preissii should also be able to recruit between fires if seed is available in years when higher than normal rainfall occurs. At our Goldfields site 5 there were a few large trees that had heavy cone loads (many open) with saplings present surrounding both live and dead large trees. Thus, inter-fire recruitment has occurred for C. preissii from time to time over recent decades, as also found for eucalypt seedlings in long unburnt Eucalyptus salubris (serotinous seeder species) woodlands that occur in the same region (Gosper et al. 2018).
Comparison with obligate seeder Eucalyptus woodlands in the Goldfields region highlights the differences between these two ecosystems based on species traits. The eucalypts shed lower branches and self-thin over time, leading to low live tree density and a sparse, grassy understorey. In contrast C. preissii is usually associated with, an often discontinuous shrubby understorey, that can support grassy fuels after higher than average rainfall. Also, plants retain low branches and if established as a dense stand after fire may become “locked”, with low mortality, slow growth and low reproduction—as found at Goldfields site 6 where < 30% of plants had accumulated only two or more cohorts of cones in 47 years. The propensity for fire differs due to the sparse vegetation under the eucalypts, the tall, steeply branched architecture of the trees and lack of persistent decorticating bark (Gosper et al. 2018) compared with the more continuous canopy and understorey in the Callitris vegetation and retention of low branches. C. preissii stands rarely exceed 140 years old in semi-arid heathland (O’Donnell et al. 2010). In contrast, eucalypt woodland patches may survive to 400 years (Gosper et al. 2018). Thus, due to their suite of traits, and the traits of species with which they associate, Callitris more frequently follows the fire route to regeneration than does the seeder eucalypt.
There is considerable uncertainty about how different species and the vegetation in which they occur may change in the future. Anthropogenic climate change and other human impacts lead to shifts in fire regimes (Rodman et al. 2021) and as C. preissii is spread over a wide geographic and climatic range there will be differences in the intensity of effects on the species in different parts of its range. In areas of higher human population such as on the West coast, fires are actively suppressed and seedling recruitment can occur without disturbance in these mesic woodland stands as native macropod grazers have been largely eliminated. In semi-arid SE Australia, Prober et al. (2012) hypothesized that grassy woodlands could become more Callitris dominated due to climate change, while more frequent fire has been predicted to lead to declines in C. endlicherii woodlands in NSW (Denham et al. 2016) and in C. glaucophylla in South Australia in parts of its range that lack fire refuges if rainfall decreases (Prior et al. 2018). However, these species differ from C. preissii because they are non-serotinous and must rely on infrequent wet years for recruitment and cannot accumulate a seed bank. Climate projections for Western Australia to the end of the twenty-first century (Western Australian DPIRD 2021) predict a widespread increase in temperature, lower rainfall along the west coast, but little change in rainfall inland. This implies that inter-fire recruitment in the more mesic parts of the state is less likely than at present but fires may be more frequent. Inland population recruitment may not be very different from now and would still be most commonly related to fire, but higher temperatures may reduce recruitment through their interactive effect with moisture (i.e., higher evapotranspiration). With increasing anthropogenic influence on environmental conditions globally, controls on the demography of plant populations may be vastly different from those in the past, particularly in terms of recruitment (Boisvert-Marsh et al. 2019). There is clearly some lability in the way C. preissii is adapted to the climate—fire regimes in different parts of its range. However, the rapidity and types of environmental changes in the future—such as in climate, fire regime and populations of grazing and browsing animals—will be influential in how successful it will be in adapting to future conditions.
Data availability
All data presented in this study are available from the first author upon reasonable request.
Code availability
N/A.
References
Adams R (1999) Germination of Callitris seeds in relation to temperature, water stress, priming, and hydration-dehydration cycles. J Arid Environ 43:437–448
Austin MP, Williams OB (1988) Influence of climate and community composition on the population demography of pasture species in semi-arid Australia. Vegetatio 77:43–49
Backhouse J (1993) Holocene vegetation and climate record from Barker Swamp, Rottnest Island, Western Australia. J R Soc WA 76:53–61
Baird AM (1937) The suspensor and embryo of Actinostrobus. J R Soc WA 23:89–95
Baird AM (1958) Notes on the regeneration of vegetation of Garden Island after the 1956 fire. J R Soc WA 14:102–107
Banks J, Pulsford I (2001) Dendrochronology of Australian cypress pines. In: Dargavel J, Hart D, Libbis B (eds) Perfumed pineries. Environmental history of Australia’s Callitris forests. Centre for Resource and Environmental Studies, The Australian National University, Canberra, pp 30–38
Bannister AJ, Hanstrum BN (1995) A climatology of extreme fire weather days in southern Western Australia. Bureau of Meteorology internal report, Perth
Boisvert-Marsh L, Périé C, de Blois S (2019) Divergent responses to climate change and disturbance drive recruitment patterns underlying latitudinal shifts of tree species. J Ecol 107:1956–1969
BOM (2018) Bureau of meteorology. http://www.bom.gov.au/climate/data/. Accessed Sept 2018
Bond WJ, Vlok J, Viviers M (1984) Variation in seedling recruitment of Cape Proteaceae after fire. J Ecol 72:209–221
Bradstock RA, Cohn JS (2002) Demographic characteristics of mallee pine (Callitris verrucosa) in fire-prone mallee communities of central New South Wales. Aust J Bot 50:653–665
Bradstock RA, Cohn JS (2012) Fire regimes and biodiversity in semi-arid mallee ecosystems. In: Bradstock RA, Williams JE, Gill AM (eds) Flammable Australia. Cambridge Uni. Press, Cambridge, pp 238–258
Burrows ND, Ward B, Robinson AD (1995) Jarrah forest fire history from stem analysis and anthropological evidence. Aust Forestry 58:7–16
Clayton-Greene KA, Ashton DH (1990) The dynamics of Callitris columellaris/Eucalyptus albens communities along the Snowy River and its tributaries in south-eastern Australia. Aust J Bot 38:403–412
Cohn JS, Lunt ID, Bradstock RA, Hua Q, McDonald S (2013) Demographic patterns of a widespread long-lived tree are associated with rainfall and disturbance along a rainfall gradient in SE Australia. Ecol Evol 3:2169–2182
Cowling RM, Lamont BB (1985) Variation in serotiny of three Banksia species along a climatic gradient. Aust J Ecol 10:345–350
Crisp MD, Cook LG, Bowman DMJS, Cosgrove M, Isagi Y, Sakaguchi S (2019) Turnover of southern cypresses in the post-Gondwanan world: extinction, transoceanic dispersal, adaptation and rediversification. New Phytol 221:2308–2319
DCBA (2021) https://catalogue.data.wa.gov.au/dataset/dbca-fire-history. Accessed 12 July 2021
Denham AJ, Vincent BE, Clarke PJ, Auld TD (2016) Responses of tree species to a severe fire indicate major structural change to Eucalyptus-Callitris forests. Plant Ecol 217:617–629
Dowdy AJ (2018) Climatological variability of fire Weather in Australia. J Appl Meteor Clim 57:221–234. https://doi.org/10.1175/JAMC-D-17-0167.1
Dowdy AJ, Kuleshov Y (2014) Climatology of lightning activity in Australia: spatial and seasonal variability. Aust Meteor Ocean J 64:103–108
Elliott CP, Lewandrowski W, Miller BP, Barrett M, Turner SR (2019) Identifying germination opportunities for threatened plant species in episodic ecosystems by linking germination profiles with historic rainfall events. Aust J Bot 67:256–267
Enright NJ, Fontaine JB (2014) Climate change and the management of fire-prone vegetation in Southwest and Southeast Australia. Geograph Res 52:34–44
Enright NJ, Miller BP, Crawford A (1994) Environmental correlates of vegetation patterns and species richness in the northern Grampians, Victoria. Aust J Ecol 19:159–168
Enright NJ, Marsula R, Lamont BB, Wissel C (1998) The ecological significance of canopy seed storage in fire-prone environments: a model for non-sprouting shrubs. J Ecol 86:946–959
Gosper CR, Yates CJ, Prober SM, Parsons BC (2012) Contrasting changes in vegetation structure and diversity with time since fire in two Australian Mediterranean-climate plant communities. Austral Ecol 37:164–174
Gosper CR, Yates CJ, Cook GD, Harvey JM, Liedloff AC, McCaw WL, Theile KR, Prober SM (2018) A conceptual model of vegetation dynamics for the unique obligate-seeder eucalypt woodlands of south-western Australia. Austral Ecol 43:681–695
Hernández-Serrano A, Verdu M, González-Martínez SC, Pausas J (2013) Fire structures pine serotiny at different scales. Am J Bot 100:2349–2356
Kala J, Tenna AS, Rudloff D, Andrys J, Rieke O, Lyons TJ (2020) Evaluation of the weather research and forecasting model in simulating fire weather for the south-west of Western Australia. Int J Wildl Fire 29:779–792
Keeley JE, Neeman G, Fotheringham CJ (1999) Immaturity risk in a fire dependent pine. J Mediterranean Ecol 1:41–48
Keeley JE, Fotheringham CJ, Baer-Keeley M (2006) Demographic patterns of post-fire regeneration in Mediterranean-climate shrublands of California. Ecol Monogr 76:235–255
Keith DA, McCaw L, Whelan RJ (2012) Fire regimes in Australian heathlands and their effects on plants. In: Bradstock RA, Williams JE, Gill AM (eds) Flammable Australia. Cambridge Uni. Press, Cambridge, pp 199–237
Kenny SA, Bennett AF, Clarke MF, Morgan JW (2018) Time-since-fire and climate interact to affect the structural recovery of an Australian semi-arid plant community. Austral Ecol 43:456–469
Ladd PG, Midgely JJ, Nield AP (2013) Serotiny in southern hemisphere conifers. Aust J Bot 61:486–496
Lamont BB, Connell SW, Bergl SM (1991) Seed bank and population dynamics of Banksia cuneata: the role of time, fire, and moisture. Bot Gazette 152:114–122
Luke RH, McArthur AG (1978) Bushfires in Australia. Forestry and Timber Bureau, CSIRO, Canberra
Mackenzie BDE, Keith DA (2009) Adaptive management in practice: conservation of a threatened plant population. Ecol Manag Rest 10:S129–S135
Maher KA, Hobbs RJ, Yates CJ (2010) Woody shrubs and herbivory influence tree encroachment in sandplain heathlands of southwestern Australia. J Appl Ecol 47:441–450
McArthur WM (1998) Changes in species and structure in the major plant communities of Garden Island, WA 1990–1996. HMAS Stirling, Environmental working paper No. 11
Nathan R, Safriel UN, Noy-Meir I, Schiller G (1999) Seed release without fire in Pinus halepensis, a Mediterranean serotinous wind-dispersed tree. J Ecol 87:659–669
Norcliffe GB (1977) Inferential statistics for geographers. Hutchinson and Co., London, p 272p
O’Connor TG, Goodall VL (2017) Population size structure of trees in a semi-arid African savanna: species differ in vulnerability to a changing environment and reintroduction of elephants. Austral Ecol 42:664–676
O’Donnell AJ, Cullen LE, McCaw WL, Boer MM, Grierson PF (2010) Dendroecological potential of Callitris preissii for dating historical fires in semi-arid shrublands of southern Western Australia. Dendrochronologia 28:37–48
O’Donnell AJ, Boer MM, McCaw WL, Grierson PF (2011a) Climate anomalies drive wild-fire occurrence and extent in semi-arid shrublands and woodlands of Southwest Australia. Ecosphere 2:127
O’Donnell AJ, Boer MM, McCaw WL, Grierson PF (2011b) Vegetation and landscape connectivity control wildfire intervals in unmanaged semi-arid shrublands and woodlands in Australia. J Biogeogr 38:112–124
O’Donnell AJ, Cook ER, Palmer JG, Tuerney CSM, Page FM, Grierson PF (2015) Autumn precipitation in Northwest Australia is unprecedented within the last two centuries. PLoS ONE 10(6):e0128533. https://doi.org/10.1371/journal.pone.0128533
Pausas JG, Keeley JE (2014) Evolutionary ecology of seeding and sprouting in fire-prone ecosystems. New Phytol 204:55–65
Peck JE, Zenner EK, Brang P, Zingg A (2014) Tree size distribution and abundance explain structural complexity differentially within stands of even-aged and uneven-aged structure types. Eur J for Res 133:335–346
Prior LD, McCaw WL, Grierson PF, Murphy BP, Bowman DMJS (2011) Population structures of the widespread Australian conifer Callitris columellaris are a bio-indicator of continental environmental change. For Ecol Manag 262:252–262
Prior LD, Hua Q, Bowman DMJS (2018) Demographic vulnerability of an extreme xerophyte in arid Australia. Aust J Bot 66:26–38
Prober SM, Hilbert DW, Ferrier S, Dunlop M, Gobbett D (2012) Combining community-level spatial modelling and expert knowledge to inform climate adaptation in temperate grassy eucalypt woodlands and related grasslands. Biodivers Conserv 21:1627–1650
Prober SM, Yuen E, O’Connor MH, Schultz L (2016) Ngadju kala: Australian Aboriginal fire knowledge in the Great Western Woodlands. Austral Ecol 41:716–732
Read J (1995) Recruitment characteristics of the white cypress pine (Callitris glaucophylla) in arid South Australia. Range J 17:228–240
Rippey ME, Hobbs RJ (2003) The effects of fire and quokkas (Setonix brachurus) on the vegetation of Rottnest Island, Western Australia. J R Soc WA 86:49–60
Rodman KC, Veblen TT, Andrus RA, Enright NJ, Fontaine JB, Gonzalez AD, Redmond MD, Wion AP (2021) A trait-based approach to assessing resistance and resilience to wildfire in two iconic North American conifers. J Ecol 109:313–326
Schmidberger JW, Ladd PG (2020) Geographic distribution and the reproductive and demographic ecology of two congeneric seeder and resprouter tree species. For Ecol Manag 475:118428. https://doi.org/10.1016/j.foreco.2020.118428
Schwilk DW, Ackerly DD (2001) Flammability and serotiny as strategies: correlated evolution in pines. Oikos 94:326–336
Tonnabel J, van Dooren JM, Midgley J, Haccou P, Mignot A, Ronce O, Olivieri I (2012) Optimal resource allocation in a serotinous non-sprouting plant species under different fire regimes. J Ecol 100:1464–1474
Trauernicht C, Murphy BP, Portner BDMJS (2012) Tree cover-fire interactions promote the persistence of a fire-sensitive conifer in a highly flammable savanna. J Ecol 100:958–968
Turner D, Ostendorf B, Lewis M (2008) An introduction to patterns of fire in arid and semi-arid Australia 1998–2004. Range J 30:95–107
Walker J (1981) Fuel dynamics in Australian vegetation. In: Gill M, Groves RH, Noble IR (eds) Fire and the Australian Biota. Australian Academy of Science, Canberra, pp 121–128
Western Australian DPIRD (2021) Climate projections for Western Australia. Western Australian Department of Primary Industries and Regional Development. https://www.agric.wa.gov.au/climate-change/climate-projections-western-australia. Accessed 13 July 20121
Acknowledgements
Access to Garden Island was facilitated by the Australian Department of Defence. S. Barrett and G. Regan provided access to Bald Island. R. Miller provided three population structures from the West coast region. J. Fontaine assisted with identifying some fire histories.
Funding
Open Access funding enabled and organized by CAUL and its Member Institutions. This work was partly funded by a grant from the Australian Department of Defence (Garden Island components).
Author information
Authors and Affiliations
Contributions
All authors contributed to the study conception, design and data collection. PGL and NJE compiled and analysed the data and wrote the first draft. All authors commented on draft versions of the manuscript and contributed to write-up of the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
All the authors declared that they have no conflict of interest.
Additional information
Communicated by Brian J. Harvey.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Ladd, P.G., Zhao, X. & Enright, N.J. Fire regime and climate determine spatial variation in level of serotiny and population structure in a fire-killed conifer. Plant Ecol 223, 849–862 (2022). https://doi.org/10.1007/s11258-022-01243-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11258-022-01243-8