
Abstract
Identifying environmental variables that affect the distribution of pest and/or nuisance species promotes effective management strategies by improving urban planning and the targeted application of direct interventions like population control. Australian white ibis Threskiornis moluccus are considered a nuisance species throughout Australia, having increased significantly in abundance in coastal urban landscapes following declines in their native inland habitats over the past three decades. Few management interventions thus far have successfully reduced conflicts between ibis and humans, necessitating a better understanding of conditions driving ibis distribution and abundance. We surveyed 26 ibis colonies throughout the Moreton Bay region in central eastern Australia and quantified the effects of a range of spatial (e.g. connectivity with waste management facilities, urban land and wetlands) and habitat (e.g. nesting tree species, density and size) variables on ibis and nest abundance. Ibis and their nests were most abundant at sites within 10 km of waste management facilities, and < 60 m from residential or industrial estates. Ibis were also more abundant at sites with tree canopy height < 10 m and lower tree species richness. Nests were most abundant on water bound islands with tree density > 0.5 trees/m2. Urbanisation had contrasting effects, with nest numbers increasing with higher connectivity (i.e. closer proximity) to urban land, while ibis numbers decreased (relative to waste management facilities). Multifaceted approaches are required for controlling ibis populations, including restricting access to anthropogenic food resources, managing the condition of remnant habitats, and considering population control at sites with high connectivity to foraging grounds.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Managing native and exotic pest animal species and their effects on ecosystems, agriculture and the economy is a global challenge (Worner and Gevrey 2006). Pest species can be defined as species which cause significant damage to urban areas, crops, and natural ecosystems, outcompete more vulnerable species for space and resources in remnant habitats, alter the natural functioning of an ecosystem, or cause human-animal conflict (Worner and Gevrey 2006; Green and Grosholz 2021). As such, pest species are often perceived as a nuisance, negatively impacting the community, the economy, or environment (Martin et al. 2007). The designation of a species as a ‘pest’ is inherently context-dependent, and often applies to plants and animals that have expanded or been introduced beyond their natural historical ranges. The interactive effects of climate change and expanding urban development can contribute towards an exponential increase in the abundance and impacts of pest species in many settings (Worner and Gevrey 2006; Mackenstedt et al. 2015; Coogan et al. 2017). Increased pest presence in urban areas can heighten human-animal conflict (Coogan et al. 2017), zoonotic disease transmission (Mackenstedt et al. 2015), and damage to property (Soulsbury and White 2015). The financial costs of these impacts can be immense. Australia alone has spent at least US$298.58 billion on controlling pest species and their impacts since 1960 (Bradshaw et al. 2021). Effectively mitigating increases in urban pest species and their impacts requires pest management and urban planning that incorporates a thorough understanding of species ecology and habitat preferences (Martin et al. 2010). Crucially, however, many existing prevention and control mechanisms fail due to a lack of this fundamental knowledge (Corben and Munro 2008; Smith 2009).
Species select habitats based on trade-offs between energy intake, reproductive success, and predation risk (Railsback and Harvey 2002; DeCesare et al. 2014; Lovera et al. 2019). Species colonisation and expansion in urban areas is therefore often mediated by the availability of food, key habitats (i.e. habitats with attributes which support reproductive success and minimise predation risk), and connectivity between populations, habitat, and resources (Worner and Gevrey 2006). Crucially, landscape connectivity can influence these effects across broad spatial scales (Van Langevelde 2015). Identifying characteristics of landscapes and habitats that affect pest species distribution can therefore uncover broad-scale patterns that can be useful in guiding urban planning, prioritising the allocation of current interventions to specific sites and/or identifying likely future locations where pest species might colonise (Moilanen and Nieminen 2002; Hulme 2009; Skidmore et al. 2011). For example, the availability of key habitats within a species home range and their connectedness across landscapes can mediate the abundance of pest species in urban areas (Vergara and Armesto 2009; Henderson et al. 2017). Many species move across heterogeneous landscapes to access sites that maximise food and nesting opportunities, thereby increasing the importance of understanding this landscape connectivity (Lookingbill et al. 2022). Consequently, it is increasingly recognised that a thorough understanding of both the spatial ecology and habitat selection preferences of pest species is crucial to minimising the impacts and colonisation of pest species (Ferguson et al. 2003; Witmer 2007; O’Rourke 2010; Lovera et al. 2019).
The Australian white ibis Threskiornis moluccus, hereafter ibis, is a large (65–75 cm long) Threskiornithid native to eastern Australia that has recently started to be considered by some members of the public and environmental managers as a pest species of national concern in urban settings (Shaw 1999; Martin et al. 2007). Historically rare in urban areas, ibis have expanded from their traditional inland wetland habitats to colonise all Australian States and Territories in recent decades, establishing large, permanent colonies and seasonal nesting colonies within and around coastal cities (Corben and Munro 2006; McKiernan and Instone 2016; IUCN 2022). Ibis are now considered common in urban areas across the eastern seaboard, and typically nest in fragmented remnant wetlands and parklands (Corben and Munro 2006). Ibis nesting colonies, sometimes numbering hundreds of nests and thousands of individuals, initiate complaints from some city residents due to their noise and odour, and may cause conflicts when ibis acclimate to humans, increasing direct interactions like stealing food from humans (Shaw 1999; Corben and Munro 2006; Martin et al. 2007). Nesting colonies can degrade habitats as nests can be large, heavy, and densely populated, smothering vegetation, and causing localised erosion and eutrophication of waterways (Corben and Munro 2008). There are concerns of large colonies competing with native bird species for space, which has been observed in closely related ibis species like the African sacred ibis Threskiornis aethiopicus overseas (Martin 2012). Nesting colonies near roads and airports increase risk of collision with vehicles and aircraft, and therefore risk of injury to people and equipment damage (Shaw 1999). Ibis often feed on litter and garbage produced by people, potentially increasing the risk of spreading diseases including avian influenza virus and Salmonella sp. (Epstein et al. 2006). However, this assumption of ibis being significant disease vectors has been increasingly questioned by more recent evidence (Maute et al. 2019). Regardless, these behaviours have contributed towards the species’ negative public perception as ‘dirty’ (McKiernan and Instone 2016), and contribute towards increasing reports of people harming and killing the species across Australia (McKiernan and Instone 2016).
Due to ongoing conflicts between ibis and local residents, natural resource managers have integrated ibis management into broader environmental management programs in many urban areas (Smith 2009). Expensive egg destruction events (including over 5500 eggs in single events) have been employed across eastern Australia (Martin 2012). Egg oiling, which applies a thin layer of household oil to eggs and prevents up to 98% of eggs from hatching, and nest destruction have also been implemented, although these practices are increasingly criticised in the literature as unsuccessful in meaningfully decreasing colony size (Martin et al. 2007, 2010; Smith 2009; Yésou et al. 2017). Instead, studies have indicated that a limited knowledge of the species’ basic ecology, habitat preferences, and movement patterns has resulted in a typically low success of these management actions in abating colony impacts and expansion, especially as current techniques do not account for population-level movements or preferences across landscapes (Martin et al. 2007). Several authors have suggested that existing population management methods simply displace, fragment, and disturb existing colonies, causing them to disperse and expand populations and their impacts spatially (Corben and Munro 2006; Smith 2009). Active interventions also result in ibis also becoming more wary and less approachable, making management practices more difficult (Yésou et al. 2017).
Connectivity between habitat patches and the size of a habitat patch combine to influence species composition and abundance by facilitating foraging success and affecting population viability (Vergara and Armesto 2009; Henderson et al. 2017; Teitelbaum et al. 2020). Connectivity is especially important in fragmented urban areas because it can facilitate movement pathways across these heterogenous landscapes (Martensen et al. 2012). Larger patch area can support a greater abundance of resources, enhanced habitat diversity, reduced edge effects, and therefore larger population size (Uezu et al. 2005; Evans et al. 2009; Martensen et al. 2012). As such, highly mobile species like ibis can reduce movement distances in more connected urban areas due to resource availability and trade-offs between energy gained from a foraging patch versus energy expenditure travelling to a foraging patch (Martin et al. 2011; Teitelbaum et al. 2020). Abundant resources in coastal urban landscapes attract ibis as food is readily available and easily accessible (Smith and Munro 2011; Davis et al. 2021). Waste management facilities and landfills provide an abundant and stable foraging ground, and so often support the highest abundance of foraging ibis in urban areas (Martin et al. 2010). Wetlands provide both foraging and suitable nesting/roosting habitat in urban landscapes where greenspace is often limited (Martin et al. 2011; Davis et al. 2021). While it is likely that these landscape-scale patterns relating to effects of landscape connectivity and patch size contribute towards structuring ibis populations in urban settings (Hamilton et al. 2006; Shanahan et al. 2011), these patterns must also be understood in the context of individual patch-scale attributes to be truly meaningful to management (Vergara and Armesto 2009; Haslem et al. 2021).
Small-scale variations in habitats influence species distribution both within and between habitat patches (Vergara and Armesto 2009). For ibis, habitat characteristics including the size, density and composition of canopy-forming and potential nesting trees (Kentish 1999; Corben and Munro 2006; Martin et al. 2010), and access by predators (Brandis et al. 2011) likely affect their choices of locations for nesting and roosting. For example, species’ nest locations within patches vary spatio-temporally according to characteristics such as tree height or density and wind exposure (Vergara and Armesto 2009), however nesting preferences vary significantly between ibis species and locations. Ibis build nests on the ground in some locations, but ground nesting is often unsafe in urban settings due to increased predation and human interference, resulting in tree nesting being more common (Shaw 1999). Similarly, the use of islands surrounded by water ensures nests have greater protection from flightless predators including rats, cats, and snakes (Shaw 1999; Martin et al. 2010; Brandis et al. 2011). Understanding habitat selection in ibis can identify features of parks and wetlands which drive ibis nesting, thereby informing future planning and restoration projects to minimise the risk of enhancing ibis populations (Singleton et al. 1999). Modifying habitat to reduce reproduction can be successfully implemented for some pest species (Smith and Carlile 1993), and may therefore be an option for ibis as well. For example, mowing of grass in a nesting habitat prior to the commencement of breeding reduced the abundance of a silver gull Larus novaehollandiae colony, a prominent Australian native-invasive (Smith and Carlile 1993). Despite broad recognition of the importance of within-habitat patch factors for the distribution of birds, including ibis, to our knowledge these effects have never been quantified in concert with broader spatial variables, like distances between key foraging and roosting sites, with a view to improving management (Reif et al. 2010).
Globally, conflict between urban pest species and people is increasing (Bradshaw et al. 2021), necessitating a more thorough understanding of the environmental drivers of pest species distribution and abundance (Singleton et al. 1999; Martin et al. 2010). We aim to identify the combined effects of landscape (especially the size and connectedness of nearby waste management facilities, urban areas, and wetlands) and habitat (especially the size, density, and composition of habitat-forming trees) attributes on the abundance and distribution of Australian white ibis and their nests in central eastern Australia. We hypothesised that the number of ibis and ibis nests at a given site is a result of the combined effects of both spatial and vegetation characteristics, and habitat-specific conditions like tree species and the presence of water-bound islands. Specifically, we expect ibis abundance to be greater at sites nearer to waste management facilities and at sites where nests are positioned on small, water-bound islands with dense vegetation. Additionally, we expect nest abundance to be greater at sites nearer to wetlands due to greater nesting habitat suitability, and that ibis will prefer specific tree species as nesting locations.
Methods
Study region and sites
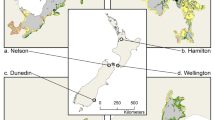
We surveyed 26 ibis colonies (i.e. both nesting and non-nesting colonies) throughout the Moreton Bay region of Southeast Queensland, Australia (Fig. 1). The Moreton Bay region is subject to significant human population increases, with the population increasing by 51,038 people between 2016 and 2021 (Australian Bureau of Statistics 2021), resulting in expansion of residential and urban areas across the region. Simultaneously, there has been steadily increasing reports of the number of ibis nesting locations in residential and recreational areas across the region, with an increased number of reported complaints, typically due to noise and smell (Moreton Bay Regional Council 2021). Since 2014, the Moreton Bay Regional Council has been actively surveying all reported sites where ibis regularly and predictably nest or roost in groups larger than ~ 10 individuals (hereafter ‘colonies’). While detectability of sites may not be perfect, sites for this study were selected from data obtained by Moreton Bay Regional Council (MBRC) of reported nesting and non-nesting ibis colonies between 2014 and 2023. Sites were typically reported to MBRC by members of the public. As additional sites were added to the program as reports of colonies increased, surveys commenced at seven sites only in 2014, with an average of three sites being added to the survey list per subsequent year. Between January 2021 and May 2023, researchers from the University of the Sunshine Coast (UniSC) continued surveys at 26 of the 35 historical ibis sites, omitting sites that were no longer active (i.e. those with no nests or individuals present). These sites covered the breadth of current ibis nesting locations across the region, varying from those positioned within highly urbanised residential areas, to more isolated, wetland-nesting colonies (Fig. 1). In addition to these nesting site data, bird abundance was recorded at three major waste management facilities across the region. Although colony detectability may have missed some colonies in the greater region (e.g. smaller, unreported colonies, or those in rural areas) we are confident we have a robust representative sample of the overall population in the urban areas of Moreton Bay. Additionally, previous studies in urban Sydney found 0.027 colonies per square kilometre (Martin et al. 2010; Regional Development Australia 2021), whereas we found 0.14 colonies per square kilometre in urban Moreton Bay (Wang et al. 2022).

Map of included Australian white ibis nesting and non-nesting colonies surveyed throughout Moreton Bay, Queensland, with palustrine wetlands and urban land highlighted in green and yellow respectively, and all other land-use types in grey (e.g., agricultural land, natural (non-wetland), parklands)
Ibis and nest abundance surveys
Data collection at each site was consistent across all years and across both survey groups (Moreton Bay Regional Council and University of the Sunshine Coast). Known sites with active colonies were typically visited once per month during daylight hours (across all seasons) from the time they were added to the program, however this varied occasionally between sites due to weather and other logistical factors. On average each site was surveyed 48 times across the entire study, and a total of 1265 surveys were conducted. Most sites were directly accessible by car or on foot, otherwise (such as those on water bodies) had a standard survey location from which the colony could be seen and counted by at least two surveyors with binoculars. Site perimeter was identified prior to surveys at all sites using Google Earth Pro to ensure totals only accounted for individuals and nests within the site. All colonies were counted using a point-count method from one to three standard viewing areas that allowed observation of the site as a whole between the group. During each survey, researchers visually counted all ibis within the site perimeter, including individuals in flight, in nests and trees, and on the ground, producing a total ibis abundance count for each site per survey. Similarly, all active nests, defined as nests occupied by either adults, juveniles, or eggs were counted within the site perimeter, producing a total nest abundance for each site per survey. Effort was taken to ensure all individuals and nests were accounted for during surveying. However, we acknowledge that imperfect detectability may be present at sites with a high abundance of individuals, and as a result, count data is limited and may produce an under abundance of individuals. Records of all surveys and methods were kept by the Moreton Bay Regional Council until 2021, after which these data were shared with the University of the Sunshine Coast, who continued the surveys in the same manner. Counts generally took approximately 20 min to complete and confirm between the two observers at each site.
Environmental variables
We tested the effects of six spatial landscape variables (Table 1A) on the abundance of ibis and their nests at each site, quantified using QGIS (QGIS Development Team 2022). Variables were chosen based on current knowledge of ibis distribution, foraging, and nesting sites in urban areas, as detailed in Table 1A. Spatial variables could be broadly grouped in three categories: distances, areas, and patch size. Distances quantify the distance (in km) from the middle of sites to the edge of the focal connected habitat or site, and included wetland, urban land and waste management facility metrics. Area metrics quantify the area of each focal habitat (in ha) within strategically selected buffers around each site, and quantified wetlands and urban land cover within the buffer. Buffer distances were selected using literature on Australian water birds and daily urban ibis travel distances (Martin et al. 2010; Martin et al. 2011), resulting in three buffer distances of 5 km, 30 km, and 70 km being chosen. Patch size (in m2), defined as the total area of each habitat patch, was measured using QGIS and NearMap (2022) with ground truthing using a handheld Garmin 70 series GPS unit. Finally, the total area of each nest patch (in m2), defined as the area in which ibis nests are positioned within the broader habitat patch, was measured using a handheld Garmin 70 series GPS in the field.
Habitat (Table 1B) characteristics surveys were undertaken at each site in May 2023 following abundance surveys to avoid disturbing counts. Habitat characteristics could be broadly grouped into three categories: island type, tree characteristics, and tree density. Habitat patches were defined as either water islands, being a patch surrounded by water, or an urban island, being a patch of remnant vegetation or parkland land surrounded by urban land including housing, shopping centres, and roads. Vegetation characteristics of each site included the maximum canopy height of the patch measured using a clinometer, and the number and species richness of trees (defined as woody vegetation > 1.5 m). Tree density was calculated as the number of trees within a patch divided by the size of the patch, giving the density of trees per m2.
Nest characteristics surveys were also undertaken once at each site in 2023 during the end of the breeding season, following abundance surveys to avoid disturbing counts. All nests were surveyed at each site. Nest characteristics included the species of tree in which the nest was positioned, identified to the lowest taxonomic level possible, and nest height was measured using a clinometer.
Statistical analysis
Two analyses were conducted for each of the two dependent variables of nest abundance and total ibis abundance. The first included all data collected from 2014 to 2023, with patterns established across this period using spatial variables only (i.e. those listed in Table 1A). The second included only data from the most recent three years of abundance data only (i.e. from 2021 to 2023), and established patterns across all of the spatial, habitat and nest characteristics (i.e. those listed in Table 1A and 1B). The reason for this split was that we could not be confident that habitat and nest characteristics quantified in 2023 were consistent across longer time frames, and so ran analyses with habitat and nest characteristics on data from 2021 onwards. We are confident that the spatial variables remained consistent throughout the study, and this was further confirmed with satellite imagery from 2014 to 2022 provided by NearMap (2022).
All explanatory variables were checked for collinearity using the Pearson’s r coefficient in R (R Core Team 2023) prior to analysis. Where any two variables correlated with coefficients either <-0.7 or > 0.7 (Ratner 2009), one variable was removed based on the strength of the underlying hypotheses detailed in Table 1. For the analysis with spatial variables only (i.e. all data from 2014 to 2023), the area of wetlands within 5 km (r = 0.733), area of wetlands within 70 km (r = 0.608), area of urban within 5 km (r = -0.800), and area of urban within 70 km (r = -0.659) all correlated with distance to waste management facility, and so were subsequently removed from the model. Given these correlations, area of wetlands within 30 km and area of urban within 30 km remained in the model. For the combined spatial and habitat characteristics analysis (i.e. 2021 to 2023 data), average nest height (m) and maximum canopy height (m) correlated (r = 0.781), with average nest height removed from model due to (1) maximum canopy height likely having a greater influence on nest abundance (Lusk et al. 2006), and (2) maximum canopy height being a simpler and typically more readily available metric for local managers.
We quantified correlations between nest abundance and total ibis abundance with each set of environmental variables (as detailed above for each analysis) using generalised additive models (GAMs) in the mgcv package (Wood 2022) of R. Model selection was undertaken by adding an additional penalty term in the smoothness selection procedure through the select = TRUE command in GAM (Marra and Wood 2011). We checked the significance of the terms in the model of best fit using GAMs. All smooth terms were significant (p = > 0.01) to the model; therefore, no further simplification of the model was performed. The model was calculated using the Tweedie error distribution with a canonical link function. GAM overfitting was minimised by limiting knots to four or fewer (i.e. k = 4) for each variable.
Results
Data patterns
The highest recorded nest abundance was 400 nests at Esme Avenue, Caboolture in February 2015: a site consisting of multiple water islands totalling 1236 m2 patch size. The mean nest abundance for all surveys was 11.80 (± 30.63 SD). The highest recorded ibis abundance was also at Esme Avenue, at 2800 individuals in February 2015. The mean ibis abundance for all sites was 101.06 (± 242.70 SD). Esme Avenue consistently harboured the largest number of nests since 2015 in Moreton Bay. Although Esme Avenue had the largest abundance of individuals (mean = 447 ± 660, maximum count 2800), the Caboolture Waste Management Facility (mean = 701 ± 241, maximum count = 1221 individuals) and the Dakabin Waste Management Facility (mean = 557 ± 222, maximum count = 1114 individuals) also consistently exhibited high numbers of ibis in Moreton Bay. These waste management facilities did not, however, ever record any nesting activity.
Eucalypts (native) and broad-leaved pepper Schinus terebinthifolius (non-native) contained the highest abundance of nests recorded in 2023, although nesting was not limited to these species (Table 2). However, broad-leaved pepper was present at only one nesting site, and was the only tree species present on the water-bound island. Conversely, nests were present in Eucalypts across four sites, and in Melaleuca (native) across three sites. Given we identified such a dominance of site-specific trends in nesting trees, and many tree species were only present in one site, nesting tree species was unable to be analysed further spatially and so was excluded from subsequent analyses. It is possible that nesting tree species was less important for nest selection than spatial or habitat variables in this study, however, more detailed research is required to address this.
Spatial effects
Ibis nest abundance at sites across all data (i.e. 2014–2023) was best explained by the combined effects of island type (Fig. 2A-D, F = 290.7, P < 0.001), the distance of sites to waste management facilities (Fig. 2A, F = 21.72, P < 0.001), urban land (Fig. 2B, F = 94.89, P < 0.001) and wetlands (Fig. 2C, F = 83.23, P < 0.001), and the area of urban land within 30 km (Fig. 2D, F = 64, P < 0.001). This model explained 35% of variance overall (R2 = 0.35). Ibis nest abundance was highest at sites on water islands and furthest from waste management facilities and urban land, nearer to wetlands, and with a higher extent of urbanisation within 30 km.

Generalised additive model (GAM) output of the best fit model describing the Australian white ibis nest abundance (panels A-D) and Australian white ibis abundance (panels E-H). These analyses quantified the effects of spatial variables only across all data collected within the Moreton Bay Region, Australia between 2014 and 2023. Error bars indicate 95% confidence intervals
Ibis abundance at sites across all data (i.e. 2014–2023) was best explained by the combined effects of island type (Fig. 2E-H, F = 34.29 P < 0.001), the distance of sites to waste management facilities (Fig. 2E, F = 343.23, P < 0.001), urban land (Fig. 2F, F = 14.06, P < 0.001) and wetlands (Fig. 2G, F = 8.90, P < 0.001), and total area of urban land within 30 km of a site (Fig. 2H, F = 40.60, P < 0.001). This model explained 46% of variance overall (R2 = 0.46). Ibis abundance was highest at sites on water islands that were nearer to waste management facilities, approximately 40 m from urban land, further from wetlands, and with a lower extent of urbanisation within 30 km.
Combined habitat and spatial effects
Ibis nest abundance at sites across the 2021–2023 dataset (i.e. the most recent three years) which included habitat variables was best explained by the combined effects of distance of sites to waste management facilities (Fig. 3A, F = 3.79, P = 0.04) and urban land (Fig. 3B, F = 6.4, P = 0.01), total patch size (Fig. 3C, F = 26.80, P < 0.01), and tree density (Fig. 3D, F = 17.30, P < 0.001). This model explained 44% of variance overall (R2 = 0.44). There was no significant difference in ibis nest abundance between urban and water islands once habitat variables were accounted for (Fig. 3A-D). Ibis nest abundance was highest at sites that were closest to waste management facilities, further from urban land, when patch size was < 50 000 m2 or > 150 000 m2, and when tree density was higher.

Generalised additive model (GAM) output of the best fit model describing the Australian white ibis nest abundance (panels A-D) and Australian white ibis abundance (panels E-J). These analyses quantified the effects of both spatial and patch characteristics variables using the subsetted 2021–2023 data collected within the Moreton Bay Region, Australia. Data was subsetted as we could not be confident in using current patch characteristics data for surveys greater than two years old. Error bars indicate 95% confidence intervals
Ibis abundance across the 2021–2023 dataset (i.e. the most recent three years) was best explained by the combined effects of island type (Fig. 3E-J, F = 95.41, P < 0.001), the distance of sites to waste management facilities (Fig. 3E, F = 23.6, P < 0.001) and urban land (Fig. 3F, F = 23.56, P < 0.001), total patch size (Fig. 3G, F = 58.10, P < 0.001), area of urban land within 30 km (Fig. 3H, F = 48.30, P < 0.001), maximum canopy height (Fig. 3I, F = 92.32, P < 0.001), and tree species richness (Fig. 3J, F = 17.43, P < 0.001). This model explained 57% of variance overall (R2 = 0.57). Ibis abundance was highest at sites on urban islands that were closer to waste management facilities, around 30 m from urban land, when patch size was > 50,000 m2, with a lower extent of urbanisation within 30 km, a lower maximum canopy height, and lower tree species richness.
Discussion
Globally, the impacts of pest species are expanding and intensifying as human impacts and urbanisation promote increased interactions between humans and wildlife (Bradshaw et al. 2021). Understanding the environmental drivers of pest species populations and their impacts is crucial in optimising future urban planning decisions to minimise pest species spread (Martin et al. 2010), and in identifying locations and actions to prioritise when they do establish (Singleton et al. 1999; Venette et al. 2010). In this study, landscape connectivity and local habitat characteristics jointly shaped ibis nest and individual abundance across an urbanising landscape in eastern Australia. The effects of connectivity with waste management facilities were consistent with our expectations for ibis abundance, with greater abundance at sites nearer to the facilities. However, results were variable and often counter to expectations for ibis nest abundance. Opposing trends were found for the effects of wetlands on ibis nest abundance versus ibis abundance, likely indicating differing preferences for ibis site selection for nesting versus roosting or feeding. Effects of site connectivity with urban land remained consistent when accounting for habitat condition. While water islands harboured more individuals and nests in the combined analyses, water islands had either lower ibis abundance, or island type did not affect ibis nest abundance once habitat variables were incorporated into analysis, indicating habitat metrics shaped distribution greater than islands. Sites with greater tree density supported a greater abundance of nests, likely due to a greater availability of suitable vegetation on which the animals build nest structures. Conversely, sites with lower tree canopy height and tree species richness had greater ibis abundance, indicating effects of homogenous stands of smaller trees on enhancing ibis abundance. Our findings show that ibis population abundance and distribution, and therefore their likely impacts on people, are shaped by a complex synergy of both spatial and habitat metrics. This means that ibis management must take a strategic approach to managing populations that incorporates information regarding site landscape connectivity, ways to restrict access to key anthropogenic food resources, and managing tree assemblages and condition in remnant habitat patches.
The importance of waste management facilities for urban bird species has been documented previously for multiple species including ibis (Baxter and Robinson 2007). We found colonies within 10 km of a waste management facility had the highest ibis and nest abundance in the combined analysis, thereby aligning closely with the results of previous studies (Martin et al. 2010). Although, no nesting colonies in this study established directly within waste management facilities, with colonies forming greater than 1.8 km away from facilities instead. Analyses with only spatial variables indicated that ibis nest abundance increased with increasing distance from waste management facilities. These findings indicate one of two patterns occurring. Firstly, habitat attributes, especially tree density, has such a mediating effect on the abundance of ibis that patterns are reversed when only spatial variables are accounted for. Alternatively, nest data prior to, and after 2020, had such differing relationships with waste management facilities that this was reflected in analyses. This might, however, be considered unlikely as there was no meaningful change in the size and/or management of focal waste management facilities over this period. Irrespective of the underlying cause of patterns, these results jointly indicate that connectivity with, and access to, waste management facilities are a major driver of ibis distribution and abundance in this region.
Ibis have double the foraging success in waste management facilities compared to natural foraging grounds (Murray and Shaw 2009; Calle and Gawlik 2011), likely leading to greater reproductive success (Martin et al. 2010). Restricting access to refuse in waste management facilities may therefore decrease ibis abundance by reducing reproductive success (Martin et al. 2010). This could include managing or restricting the openness of landfill sites to reduce accessibility by ibis, including by more regularly covering edible refuse with at least 15 cm of soil (Martin et al. 2010). However, further research regarding such interventions is required as the effects of changing waste management procedures on ibis remains understudied (Baxter and Robinson 2007; Martin et al. 2010). Reducing connectivity between colonies and waste management facilities by decreasing preferred nesting habitat (especially sites with tree density greater than 0.5 trees/m2) at sites closer than 10 km to waste management facilities may further reduce population size. The strategic placement of any new waste management facilities across landscapes should therefore be a management priority. For example, councils could avoid constructing waste management facilities near areas where human-animal conflict occurs, such as next to major roads or airports, to minimise the chance of collisions.
While urbanisation typically has negative impacts on biodiversity, many generalist bird species can excel in an urban environment when provided the opportunity and resources (Ortega-Álvarez and MacGregor-Fors 2009). We found that ibis and nest abundance was consistently greater at nesting colonies positioned more than 40 m from urban areas. Crucially, however, all colonies within this study were positioned within 70 m of urbanisation: a very short distance for a species with a potential daily home range at the scale of 10s of kilometres (Martin et al. 2011). These findings align with previous studies which found that the most abundant ibis colonies establish within 100 m of urban areas (Martin et al. 2010). Ibis species are able to nest in highly urbanised areas, such as the extremely urbanised Delhi Zoo in Delhi, India (Urfi 2010), and these areas often correspond to greater fledgling success due to greater nearby resources (Corben and Munro 2006). Therefore, while analyses found effects of intermediate distances to urban in this study, these effects are occurring at a very narrow spatial scale. While ibis can therefore be considered synanthropic and abundant in urban areas, they still prefer to nest in locations with some buffering from people and potentially other factors human-associated which have not yet been explored.
The area of nearby urbanisation has mixed effects on patterns for both ibis nest and individual abundance. While lower extents of urban land within 30 km consistently resulted in lower ibis abundance, greater extents were preferred for nesting in the spatial only analysis, and urban area was dropped from nest models when habitat variables were included. Broadly, these differences indicate ibis exploit separate foraging and nesting/roosting grounds to maximise foraging and reproductive success in urban landscapes (Smith and Munro 2011). Effects of lower extents of urbanisation on ibis abundance support the notion that ibis prefer patchy urban landscapes with interspersed green space, often on the periphery of large cities, as nesting and roosting locations (Martin et al. 2010). This highlights the need to manage both foraging and nesting/roosting sites, and minimise connectivity between the two, rather than focusing solely on nesting colonies, which has historically been the preferred form of ibis management (Martin et al. 2007). Our results also emphasise the adaptability of ibis, with large colonies able to successfully establish close to and within urban areas when habitat preferences are met.
Greenspace is typically limited in urban areas meaning that a higher abundance and diversity of avian species can often be found in small, remnant habitat patches due to a lack of alternative habitats (Irvine et al. 2009; Aida et al. 2016; Von Thaden et al. 2021). Wetlands can provide such locations as they are often among the few remnant vegetation areas left in urbanising landscapes due to the complexities of building homes or industrial estates within them (Davis et al. 2021). Despite this, ibis distribution was more influenced by anthropogenic attributes of landscapes, including waste management facilities and urban areas, than by wetlands in this study. We found opposing effects of wetlands on ibis and nest abundance in urban wetlands in the spatial only analysis, and wetlands were dropped altogether from models once habitat variables were included. Broadly, we found that the most abundant nesting colonies established within 1 km of a wetland in the spatial analysis. Conversely, ibis abundance increased exponentially further from wetlands. Wetlands provide a relatively stable nesting habitat in urban landscapes where land use is constantly changing (Davis et al. 2021), and where predators (like foxes, cats, and dogs) and human interference are abundant (Shaw 1999; Martin et al. 2010). Further, there is evidence some bird species feed nestlings a more natural diet which is of higher nutritional value, rather than food from anthropogenic sources which adults commonly consume (Lenzi et al. 2019). Again, these opposing trends highlight the importance of connectivity between foraging and nesting/roosting sites, but perhaps also connectivity between multiple foraging grounds, and the need to manage this connectivity to manage urban pest species.
Designing and building urban developments near wetlands is common practice to maximise recreation value for people (through walking tracks, fishing, and water sports etc.). However, such urban planning, especially in areas where wetlands contain isolated islands surrounded by water with a high density of trees, may in fact encourage the settlement of ibis in these locations. Further, enhancing wetlands by revegetation in and around urban areas combined with reducing access to urban resources and nesting/roosting habitat may shift colonies from more urbanised areas, such as waste management facilities and parklands (Martin et al. 2010). Still, spatial metrics were the most significant drivers of ibis abundance in this study across all four statistical analyses, and therefore management of the landscape should remain at the forefront of ibis management.
Avoidance of predators is a key habitat consideration for animals (Fattebert et al. 2019; Lovera et al. 2019). We found that ibis nest and individual abundance was greater on water-bound islands than at sites surrounded exclusively by urban land in the spatial analysis. These effects were opposite for ibis abundance when habitat variables were account for, and were not significant at all for nest abundance; likely due to mediating effects of patch size and tree density in this analysis influencing ibis abundance greater than island type. The use of water-bound islands likely increases protection from predators, thereby increasing safety of adults, nest and chicks (Martin et al. 2010).
Within-patch habitat characteristics affect abundance and colonisation of birds in many settings (Ortega-Álvarez and MacGregor-Fors 2009; Kellner 2018), and modified the effects of some spatial patterns of ibis in this study. Our results show that ibis did not necessarily select one tree species over another, preferring to select habitats with higher tree density, and foraging/roosting sites with lower maximum canopy height, and lower tree species richness. Nest abundance increased exponentially when tree density was greater than 0.5 trees per m2, with lower densities supporting colonies of less than 10 nests. Ibis tend to nest in dense colonies (Shaw 1999), and increasing tree density likely facilitates this colonial nesting. Higher tree density can also increase nest safety through protection and concealment (Götmark et al. 1995), which may be a crucial safety measure for species nesting in urban landscapes (Martin et al. 2010). We found the most abundant ibis colonies established in habitats where the maximum canopy height was less than 10 m, while a similar study in Centennial Park, Sydney, found nests to be predominately constructed on vegetation under 1 m in height as available vegetation was scarce (Corben and Munro 2006). Varying canopy height provides distinct opportunities for colonisation, hence canopy height influences distribution of avian species (Kellner 2018). We found ibis abundance to be greater at sites with lower tree species richness. This pattern could be explained by ibis preferring sites with homogenous stands of trees and therefore typically lower habitat heterogeneity. Indeed, this homogeneity likely contributes to the reliability of our counting methods. Counting individuals in large colonies, such as those present in the waste management facilities of our study, comes with the inherent risk of underestimating abundance. Though, the significant differences in colony size between nesting colonies and colonies in the three waste management facilities means (1) most colonies were easily counted due to small size (typically less than 20 individuals, barring a few sites with ~ 100 individuals) and unrestricted visibility (open low-laying canopy, low density of vegetation); (2) the waste management facilities were exponentially larger in spatial extent than that of other nesting or roosting colonies, and as such under-counting abundance would not significantly impact the data.
Another reason for this pattern might be the result of long-term impacts of high-density ibis nesting at some sites. Here, ibis nesting behaviour compresses tree canopies and can cause localised nutrient enrichment and erosion, thereby potentially selecting for only the hardiest of tree species at these sites, reducing species richness. However, these hypotheses require further testing. These fine-scale spatial analyses provide guidelines for parkland planning. Strategic revegetation can reduce pest abundance, and has successfully decreased the abundance of a native pest, noisy miner Manorina melanocephala, while enhancing other native species abundance through re-establishing the understory of various New South Wales woodlands (Lindenmayer et al. 2018). Careful consideration when vegetating parklands and other urban landscapes, especially when positioned near foraging grounds, could minimise attractiveness and available space in the park deterring large ibis colonies. Planting a diverse range of taller trees in low densities within parklands and urban areas may therefore discourage colony establishment.
The suite environmental variables that affect the distribution of pest species across landscapes is often complex and driven at multiple spatial scales (Hamilton et al. 2006; DeCesare et al. 2014). Our results support the notion that a multifaceted approach is necessary for managing ibis populations across landscapes. Such programs must focus on targeting access to resources, connectivity between colonies and foraging grounds, and minimising preferred habitat attributes like low tree species richness and dense tree stands. Waste management facilities had the most consistent and significant effect on ibis abundance in this study, with connectivity to waste management facilities being a major driver in ibis distribution. Stakeholders should incorporate improving waste management facility policies in ibis reduction plans (Martin et al. 2010), especially by reducing the amount and accessibility of available refuse onsite. Connectivity to urban areas and the intensity of urban development was secondary to this, with large ibis colonies always establishing close (≤ 70 m) to urban areas. This study shows that planting a diverse range of taller trees in low densities in greenspaces, nearby urban areas may discourage colony establishment. Creating areas in wetlands which provide suitable nesting area, could entice nesting colonies away from urban land and into wetlands. We suggest that the multi-scale spatial analysis conducted here could be useful in a variety of contexts, and for a variety of pest species. For example, managing the most costly pests in Australia like the feral cat Felis catus, or black rat Rattus rattus (Bradshaw et al. 2021). Based on our findings, we suggest pest managers seeking to control nuisance bird populations in urban landscapes by: (1) targeting both foraging and breeding/roosting sites by reducing access to both resources and suitable nesting/roosting habitat, (2) considering landscape connectivity between sites when managing specific locations, and (3) incorporating both spatial and habitat metrics into study systems to determine the most significant drivers of abundance.
Data availability
The datasets used and/or analysed and associated statistical code used during the current study are available in the supplementary materials and from the corresponding author(s) on reasonable request.
References
Aida N, Sasidhran S, Kamarudin N, Aziz N, Puan CL, Azhar B (2016) Woody trees, green space and park size improve avian biodiversity in urban landscapes of Peninsular Malaysia. Ecol Ind 69:176–183
Australian Bureau of Statistics (2021) Moreton Bay. ABS Website, accessed 01 August 2023
Baxter AT, Robinson AP (2007) A comparison of scavenging bird deterrence techniques at UK landfill sites. Int J Pest Manage 53:347–356
Bradshaw CJ, Hoskins AJ, Haubrock PJ, Cuthbert RN, Diagne C, Leroy B, Andrews L, Page B, Cassey P, Sheppard AW (2021) Detailed assessment of the reported economic costs of invasive species in Australia. NeoBiota 67:511–550
Brandis K, Kingsford R, Ren S, Ramp D (2011) Crisis water management and ibis breeding at Narran Lakes in arid Australia. Environ Manage 48:489–498
Calle L, Gawlik DE (2011) Anthropogenic food in the diet of the Sacred Ibis, Threskiornis aethiopicus, a non-native wading bird in southeastern Florida, USA. Fla Field Naturalist 39:1–38
Coogan SC, Machovsky-Capuska GE, Senior AM, Martin JM, Major RE, Raubenheimer D (2017) Macronutrient selection of free-ranging urban Australian white ibis (Threskiornis moluccus). Behav Ecol 28:1021–1029
Corben D, Munro U (2006) Clutch size and fledgling success of the Australian white ibis Threskionis Molucca in an urban environment. Corella 30:55–58
Corben DT, Munro U (2008) The abundance and movements of the Australian white ibis Threskiornis Molucca in an urban environment. Corella 32:58–65
Davis S, Brandis KJ, Stow AJ (2021) Population genetics informs the management of a controversial Australian waterbird. Conserv Genet 22:1023–1034
DeCesare NJ, Hebblewhite M, Bradley M, Hervieux D, Neufeld L, Musiani M (2014) Linking habitat selection and predation risk to spatial variation in survival. J Anim Ecol 83:343–352
Epstein JH, McKee J, Shaw P, Hicks V, Micalizzi G, Daszak P, Kilpatrick AM, Kaufman G (2006) The Australian white ibis (Threskiornis molucca) as a reservoir of zoonotic and livestock pathogens. EcoHealth 3:290–298
Evans KL, Newson SE, Gaston KJ (2009) Habitat influences on urban avian assemblages. Ibis 151:19–39
Fattebert J, Morelle K, Jurkiewicz J, Ukalska J, Borkowski J (2019) Safety first: seasonal and diel habitat selection patterns by red deer in a contrasted landscape. J Zool 308:111–120
Ferguson AW, Klukowski Z, Walczak B, Clark SJ, Mugglestone MA, Perry JN, Williams IH (2003) Spatial distribution of pest insects in oilseed rape: implications for integrated pest management. Agric Ecosyst Environ 95:509–521
Götmark F, Blomqvist D, Johansson OC, Bergkvist J (1995) Nest site selection: a trade-off between concealment and view of the surroundings? J Avian Biol: 305–312
Green SJ, Grosholz ED (2021) Functional eradication as a framework for invasive species control. Front Ecol Environ 19:98–107
Hamilton GS, Mather PB, Wilson JC (2006) Habitat heterogeneity influences connectivity in a spatially structured pest population. J Appl Ecol 43:219–226
Haslem A, Clarke RH, Holland GJ, Radford JQ, Stewart A, Bennett AF (2021) Local management or wider context: what determines the value of farm revegetation plantings for birds? J Appl Ecol 58:2552–2565
Henderson CJ, Gilby BL, Lee SY, Stevens T (2017) Contrasting effects of habitat complexity and connectivity on biodiversity in seagrass meadows. Mar Biol 164:1–9
Hulme PE (2009) Trade, transport and trouble: managing invasive species pathways in an era of globalization. J Appl Ecol 46:10–18
Irvine KN, Devine-Wright P, Payne SR, Fuller RA, Painter B, Gaston KJ (2009) Green space, soundscape and urban sustainability: an interdisciplinary, empirical study. Local Environ 14:155–172
IUCN (2022) The IUCN Red List of Threatened Species. Version 2022-2. https://www.iucnredlist.org/species/22697519/93618773. Accessed 10 August 2023
Kellner KF (2018) Local-scale habitat components driving bird abundance in eastern deciduous forests. Am Midl Nat: 52–65
Kentish B (1999) Breeding of Australian white ibis, straw-necked ibis and silver gulls on Winter Swamp, Ballarat from 1990–1997. Corella 23:37–42
Lenzi J, González-Bergonzoni I, Machín E, Pijanowski B, Flaherty E (2019) The impact of anthropogenic food subsidies on a generalist seabird during nestling growth. Sci Total Environ 687:546–553
Lindenmayer DB, Blanchard W, Crane M, Michael D, Florance D (2018) Size or quality. What matters in vegetation restoration for bird biodiversity in endangered temperate woodlands? Austral Ecol 43:798–806
Lookingbill TR, Minor ES, Mullis CS, Nunez-Mir GC, Johnson P (2022) Connectivity in the urban landscape (2015–2020): who? Where? What? When? Why? And how? Curr Landsc Ecol Rep 7:1–14
Lovera R, Fernández MS, Cavia R (2019) Small rodent species on pig and dairy farms: habitat selection and distribution. Pest Manag Sci 75:1234–1241
Lusk JJ, Smith SG, Fuhlendorf SD, Guthery FS (2006) Factors influencing northern bobwhite nest-site selection and fate. J Wildl Manag 70:564–571
Mackenstedt U, Jenkins D, Romig T (2015) The role of wildlife in the transmission of parasitic zoonoses in peri-urban and urban areas. Int J Parasitology: Parasites Wildl 4:71–79
Marra G, Wood SN (2011) Practical variable selection for generalized additive models. Comput Stat Data Anal 55:2372–2387
Martensen AC, Ribeiro MC, Banks-Leite C, Prado PI, Metzger JP (2012) Associations of forest cover, fragment area, and connectivity with neotropical understory bird species richness and abundance. Conserv Biol 26:1100–1111
Martin J (2012) Investigations into the ecology and management of a nuisance native bird in urban environments: the Australian white ibis. University of Wollongong, School of of Biological Sciences
Martin JM, French K, Major RE (2007) The pest status of Australian white ibis (Threskiornis molucca) in urban situations and the effectiveness of egg-oil in reproductive control. Wildl Res 34:319–324
Martin J, French K, Major R (2010) Population and breeding trends of an urban coloniser: the Australian white ibis. Wildl Res 37:230–239
Martin JM, French K, Ross GA, Major RE (2011) Foraging distances and habitat preferences of a recent urban coloniser: the Australian white ibis. Landsc Urban Plann 102:65–72
Maute K, Webb C, Phalen D, Martin J, Hobson-Peters J, Major RE, French K (2019) Clean bill of health? Towards an understanding of health risks posed by urban ibis. J Urban Ecol 5:juz006
McIntosh AR, McHugh PA, Plank MJ, Jellyman PG, Warburton HJ, Greig HS (2018) Capacity to support predators scales with habitat size. Sci Adv 4:eaap7523
McKiernan S, Instone L (2016) From pest to partner: rethinking the Australian White Ibis in the more-than-human city. Cult Geographies 23:475–494
Meyer-Gleaves S, Jones DN (2007) Relative abundance of Australian White Ibis Threskiornis molluca across the greater Brisbane region. Pest or Guest: Zool Overabundance: 142–149
Moilanen A, Nieminen M (2002) Simple connectivity measures in spatial ecology. Ecology 83:1131–1145
Moreton Bay Regional Council (2021) Regional Growth Management Strategy 2041
Murray NJ, Shaw PP (2009) Foraging behaviour and success of Australian white ibis Threskiornis Molucca in an urban environment. Notornis 56:201–205
NearMap (2022) NearMap photomaps. http://www.nearmap.com.au
O’Rourke M (2010) Linking habitat diversity with spatial ecology for agricultural pest management. Cornell University
Ortega-Álvarez R, MacGregor-Fors I (2009) Living in the big city: effects of urban land-use on bird community structure, diversity, and composition. Landsc Urban Plann 90:189–195
QGIS Development Team 2022 QGIS Geographic Information System. Open Source Geospatial Foundation
R Core Team (2023) A language and environment for statistical computing. R Foundation for Statistical Computing. Vienna, Austria
Railsback SF, Harvey BC (2002) Analysis of habitat-selection rules using anindividual‐based model. Ecology 83:1817–1830
Ratner B (2009) The correlation coefficient: its values range between + 1/– 1, or do they? J Target Meas Anal Mark 17:139–142
Regional Development Australia (2021) RDA Sydney
Reif J, Jiguet F, Šťastný K (2010) Habitat specialization of birds in the Czech Republic: comparison of objective measures with expert opinion. Bird Study 57:197–212
Santana Marques P, Resende Manna L, Clara Frauendorf T, Zandonà E, Mazzoni R, El-Sabaawi R (2020) Urbanization can increase the invasive potential of alien species. J Anim Ecol 89:2345–2355
Shanahan DF, Miller C, Possingham HP, Fuller RA (2011) The influence of patch area and connectivity on avian communities in urban revegetation. Biol Conserv 144:722–729
Shaw PP (1999) Integrated management systems for Australian white ibis (Threskiornis molucca) on the Gold Coast, Queensland, Australia. 1999 Bird Strike Committee-USA/Canada, First Joint Annual Meeting, Vancouver, BC, p30
Singleton GR, Leirs H, Hinds LA, Zhang Z (1999) Ecologically-based management of rodent pests–re-evaluating our approach to an old problem. Ecologically-based Management of Rodent Pests. Australian Centre for International Agricultural Research (ACIAR), Canberra 31: 17–29
Skidmore AK, Franklin J, Dawson TP, Pilesjö P (2011) Geospatial tools address emerging issues in spatial ecology: a review and commentary on the Special Issue. Int J Geogr Inf Sci 25:337–365
Smith ACM (2009) Population ecology of the Australian white ibis, Threskiornis molucca. The urban environment. Faculty of Science. University of Technology, Open Publications of UTS Scholars
Smith GC, Carlile N (1993) Methods for population control within a silver gull colony. Wildl Res 20:219–225
Smith ACM, Munro U (2011) Local and regional movements of the Australian white ibis Threskiornis Molucca in eastern Australia. Corella 35:89–94
Soulsbury CD, White PC (2015) Human–wildlife interactions in urban areas: a review of conflicts, benefits and opportunities. Wildl Res 42:541–553
Teitelbaum CS, Hepinstall-Cymerman J, Kidd-Weaver A, Hernandez SM, Altizer S, Hall RJ (2020) Urban specialization reduces habitat connectivity by a highly mobile wading bird. Mov Ecol 8:1–13
Uezu A, Metzger JP, Vielliard JM (2005) Effects of structural and functional connectivity and patch size on the abundance of seven Atlantic Forest bird species. Biol Conserv 123:507–519
Urfi AJ (2010) Using heronry birds to monitor urbanization impacts: a case study of Painted Stork Mycteria leucocephala nesting in the Delhi Zoo, India. Ambio 39:190–193
Van Langevelde F (2015) Modelling the negative effects of landscape fragmentation on habitat selection. Ecol Inf 30:271–276
Venette RC, Kriticos DJ, Magarey RD, Koch FH, Baker RH, Worner SP, Gómez Raboteaux NN, McKenney DW, Dobesberger EJ, Yemshanov D (2010) Pest risk maps for invasive alien species: a roadmap for improvement. Bioscience 60:349–362
Vergara PM, Armesto JJ (2009) Responses of Chilean forest birds to anthropogenic habitat fragmentation across spatial scales. Landscape Ecol 24:25–38
Von Thaden J, Badillo-Montaño R, Lira-Noriega A, García-Ramírez A, Benítez G, Equihua M, Looker N, Pérez-Maqueo O (2021) Contributions of green spaces and isolated trees to landscape connectivity in an urban landscape. Urban Forestry Urban Green 64:127277
Wang S, Liu Y, Feng Y, Lei Z (2022) Spatially-explicit prediction of low-density peri-urban development: comparison between urban and rural scenarios in the Moreton Bay Region in South East Queensland, Australia. Environ Plann B: Urban Analytics City Sci 49:1820–1837
Witmer GW (2007) The ecology of vertebrate pests and integrated pest management (IPM). USDA National Wildlife Research Center-Staff, p 730
Wood S (2022) Mgcv: mixed GAM computation vehicle with GCV/AIC/REML smoothness estimation. R Package Version 1:8–36
Worner S, Gevrey M (2006) Modelling global insect pest species assemblages to determine risk of invasion. J Appl Ecol 43:858–867
Yésou P, Clergeau P, Bastian S, Reeber S, Maillard J-F (2017) The Sacred Ibis in Europe: ecology and management. Br Birds 110:197–212
Acknowledgements
The authors thank Moreton Bay Regional Council for both funding the project and providing access to the historical dataset used in this study. The authors also thank Angela Webb and Brittany Elliott for their contributions to the conceptualisation of this project and their contributions the field.
Funding
This work was supported by Moreton Bay Regional Council.
Open Access funding enabled and organized by CAUL and its Member Institutions
Author information
Authors and Affiliations
Contributions
All authors (Ben Gilby, Dominique Potvin, Caitlin Willis, Darian Cadwaladr and Erin Wills) contributed to the study conception and design, and data collection. Analysis was performed by Caitlin Willis, Dominique Potvin and Ben Gilby. The first draft of the manuscript was written by Caitlin Willis and Ben Gilby and all authors commented on previous versions of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Willis, C.S., Potvin, D.A., Cadwaladr, D. et al. Landscape and habitat characteristics jointly drive the urban distribution of a nuisance species. Urban Ecosyst (2024). https://doi.org/10.1007/s11252-024-01594-2
Accepted:
Published:
DOI: https://doi.org/10.1007/s11252-024-01594-2