
Abstract
Exponential increases in anthropogenic noise and light pollution have accompanied growth of the built environment. Noise and light cause negative consequences for birds, such as disrupted navigation during migration, mortality from collisions with windows and other infrastructure, and reduced reproductive success, as well as some positive consequences, such as expanded night niches for behaviors associated with feeding, territoriality, and mating. Relatively less is known about noise and light effects on annual survival of non-migratory birds, so we conducted an exploratory study to examine variation in adult survival rates of seven avian species in relation to noise and light pollution. We used 20 years of band-resight data collected as a part of the Neighborhood Nestwatch Program (NN), a citizen science project run by the Smithsonian Migratory Bird Center, at 242 sites in greater Washington, D.C. USA. We estimated apparent survival and documented species-specific relationships with light and noise. Gray Catbird (Dumetella carolinensis) and House Wren (Thryothorus aedon) survival decreased and American Robin (Turdus migratorius) survival increased with greater amounts of anthropogenic light. Anthropogenic noise had no relationship with apparent survival for any of the seven species. Life-history trade-offs between survival and reproduction may account for differences in species-specific effects of light pollution. Future research should examine the availability of other fine scale environmental conditions, such as tree canopy cover, that might buffer avian exposure to noise and light pollution.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Although urban areas cover only about 3% of the Earth (Liu et al. 2014), urbanization can dramatically influence the abundance and distribution of birds at multiple scales (McKinney and Lockwood 1999; Marzluff et al. 2001). Projections indicate an increase of more than 3 billion people in urban settings between 2010 and 2050 (Buhaug and Urdal 2013). Increased urban populations drive concurrent increases in urban infrastructure, such as impervious surface, artificial lighting, and anthropogenic noise. Anthropogenic-dominated lightscapes and soundscapes can function as sensory pollution with pervasive ecological impacts, because virtually every organism on Earth relies on sensory systems to receive and respond to environmental cues (Dominoni et al. 2020; Oteiza and Baldwin 2021).
Rapid anthropogenic changes in noise and light pollution alter evolutionarily stable sensory environments. Avian species respond to artificial light and anthropogenic noise in a variety of ways that can affect reproduction (Dominoni et al. 2013; Senzaki et al. 2020), nestling growth and development (Rapp et al. 2016), individual health (i.e., body condition, Jiménez-Peñuela et al. 2019), communication (Nemeth and Brumm 2010), foraging behavior (Stracey et al. 2014), migration (Van Doren Benjamin et al. 2017) and mortality (La Sorte et al. 2017). Anthropogenic noise and light pollution together can cause changes in larger-scale patterns of community richness and abundance (Proppe et al. 2013; Francis 2015; Ware et al. 2015).
Several forms of behavioral plasticity may allow birds to mitigate potential costs from exposure to anthropogenic noise and artificial light, such as altering their vocalizations to reduce masking by anthropogenic noise (Slabbekoorn and Peet 2003; Nemeth and Brumm 2010; Da Silva et al. 2014), exploiting new night niches by consuming organisms that are attracted to artificial light (Lebbin et al. 2007), and seizing additional mating opportunities (Kempenaers et al. 2010). Additionally, the ability to adjust behaviorally to artificial light improves the fitness of some species in urban areas. For example, Northern Mockingbirds (Mimus polyglottos) were more likely to sing at night (Derrickson 1988) and were more likely to continue to feed nestlings past sunset in areas where the average light level around the nest was greater, further supporting the expectation that abundant urban species can exploit light pollution (Stracey et al. 2014).
Avian responses to artificial light and anthropogenic noise may vary based on species-specific natural history traits. For example, unlike cavity-nesting species, open-cup nesters lack sound-attenuating walls around nests. Thus, nestlings raised in cavities may experience less exposure to ambient noise than those raised in open-cup nests (Martin and Li 1992). Nest success and clutch sizes of forest-dwelling bird species declined in noisy environments (Senzaki et al. 2020). Additionally, traits such as vocalization frequency, nest location, and diet partly accounted for species-specific changes in reproductive timing and hatching success in response to noise levels (Senzaki et al. 2020). Noise may delay clutch initiation for species with lower-frequency vocalizations by interfering with male mate attraction songs (Swaddle et al. 2015), which can interfere with stimulation of females by males and delay female sexual receptivity (Huet des Aunay et al. 2017).
Anthropogenic noise and artificial light may directly or indirectly affect avian annual survival. Artificial light at night can interfere with the migration passages of nocturnal migrating birds, disorienting them and causing high mortality due to collisions with illuminated buildings and windows (Evans-Ogden 2022; Parkins et al. 2015). In areas with severe noise pollution, birds may modify predator vigilance (Ware et al. 2015, Kleist et al. 2016), which may increase predation risk or reduce the amount of time available for foraging. Additionally, noisy urban conditions can lead to changes in territorial behaviors, such as causing birds to approach more closely to be able to hear an acoustic signal. Closer interaction between conspecifics could eventually lead to aggressive territorial interactions that reduce individual fitness and potentially affect survival (Phillips and Derryberry 2018).
In a citizen science study of resident birds in the greater Washington, DC area, Evans et al. (2015) discovered that the apparent survival of American Robin (Turdus migratorius), Song Sparrow (Melospiza melodia), Northern Cardinal (Cardinalis cardinalis), and Gray Catbird (Dumetella carolinensis) increased with urbanization (measured as amount of impervious surface). In contrast, the apparent survival of Carolina Wren (Thryothorus ludovicianus) and House Wren (Thryothorus aedon) decreased with increased urbanization. We sought to understand whether two major components of urbanization - artificial light, anthropogenic noise - could account for the complex patterns between annual survival and degree of urbanization. Thus, our goal was to examine annual survival rates of the same non-migratory populations of adult songbirds in the greater Washington, DC area in relation to artificial light, anthropogenic noise pollution, and their paired effects while controlling for the amount of impervious surface.
We used 20 years of band-resight data to estimate annual survival of the same seven bird species as Evans et al. (2015) in relation to light and noise. While recognizing the potential for behavioral plasticity and different natural history traits in mitigating or amplifying species responses, we predicted that even urban-adapted species experience reduced adult survival with increased light, noise, and their combination, irrespective of their associations with impervious surface. We investigated the effects of light and noise on apparent survivorship of seven species of birds most common across the development gradient within the study region using various combinations of model covariates of light, noise, and impervious surface cover.
Methods
We examined the response of apparent adult survivorship to light and noise. Staff and volunteers of the Neighborhood Nestwatch Program (NN), a citizen science project run by the Smithsonian Migratory Bird Center, provided avian count data from 2000 to 2020. With sampling predominately located at the homes of project participants, NN sampling sites were within a 100-km spatial scale encompassing a rural-to-urban land-use gradient in the greater Washington, D.C., USA, metropolitan region. Project participant sites were chosen based on their position along the rural-to-urban gradient as assessed by the proportion of impervious surface; sites were defined as a 100-m radius area surrounding each sampling location (described below). NN sites (n=242) included private residences, community centers, and schools, as well as sites within forested and agricultural land cover. By incorporating privately owned land within the study design, we captured portions of the urban and suburban matrix not normally monitored in avian survivorship studies. NN sites were representative of the urban-rural gradient within the study area, as documented by Evans et al. (2015).
NN technicians visited sites once annually during the avian breeding season (May-August). To mitigate between-site differences, visit dates were scheduled such that sites visited early in the breeding season each year were visited late on the following year. NN technicians captured adult birds with mist netting with a combination of territorial intrusion playback and mobbing call playback. Individuals of seven focal species were marked with a unique U.S. Fish and Wildlife Service aluminum band and a unique combination of colored plastic bands. During each banding visit, technicians spent one hour attempting to re-sight previously captured individuals within a 200-m radius of the banding station using the playback techniques as described above. Project participants searched for color-banded birds throughout the year and reported observations to NN. Participant resights accounted for the majority of re-encounters (55%, n = 2,263).
For each of the 242 NN sites, we extracted corresponding data from the georeferenced maps of light pollution, noise pollution, and impervious surface (Fig. 1). Using Pearson Correlation Coefficients, we determined that light, noise, and impervious surface were correlated (Table 1).

Inset maps and correlations of sampling sites of the Neighborhood Nestwatch Program in greater Washington, D.C., USA
Following Sensaki et al. (2020), we used data from the second world atlas of artificial night sky brightness converted to 270-m resolution (Falchi et al. 2016). Estimates of light pollution were the zenith anthropic sky brightness as a ratio to the natural background sky brightness. Light pollution estimates provided a single, average value across an entire pixel and all light data was considered equally. For example, an area with many houses or only one airport could emit similar levels of light.
Following Mennitt and Fristrup (2016) we used a statistical approach to incorporate anthropogenic noise data from a georeferenced map of expected sound pressure levels. These data model natural sound levels from biotic and physiographic sources and compute anthropogenic noise exceedance levels. Geospatial sound models incorporate spatial representations of anthropogenic and physiographical factors to assess expected contributions to environmental sound levels from both anthropogenic and natural sources. A regression model is trained to find relationships between the explanatory variables and a response. The method we used can capture spatiotemporal patterns that may be exhibited by only a small portion of the available sample, as well as model interactions between variables and capture non-linear behavior effectively. We used anthropogenic daytime A-weighted L50 sound pressure levels as the estimate of anthropogenic noise.
We examined the influence of light and noise on the annual survival of seven species of birds most common across the development gradient within the study region: American Robin, Carolina Chickadee (Poecile carolinensis), Carolina Wren, Gray Catbird, House Wren, Northern Cardinal, and Song Sparrow. All these species were present in the study area year-round though individuals of some species (e.g., American Robin, Gray Catbird, and House Wren) may have been short-distance migrants. We investigated the effects of light and noise on apparent survivorship (Φ) and detection probability by fitting Cormack-Jolly-Seber survivorship models to these data. Detection probability was assumed to be constant across sites and years. Time-varying estimates of detection and survival were not achievable due to within-year sample size constraints. We used the R package marked (Laake et al. 2013) to build descriptive models and compare their fit to the data according to Akaike's Information Criterion (AIC; Akaike 1973) to estimate annual survival of the seven focal species at each site. We built models of adult survival that incorporated combinations of individual covariates (light, noise, and impervious surface) and ran species-specific models to determine the relative effect of our urbanization metrics on adult survival.
Because Evans et al. (2015) showed that avian adult survival in response to urbanization was often nonlinear, we included both linear (IMP) and quadratic (IMP2) impervious terms as model covariates (Table 2). Covariates used in model construction (see Table 2) were standardized as z-scores \(\left(\frac{x-\mu }{\sigma }\right)\). We used Akaike’s information criterion (AIC; Akaike 1973) to select the best models among a set of biologically plausible candidate models. We selected the model with the smallest ΔAICc as the best among all models being compared. Additionally, we used normalized AIC weights, the ratio of the likelihood of a given model relative to the sum of the likelihood across models, to evaluate the weight of evidence for a given model relative to the full set of candidate models. We used likelihood ratios (neg2lnl), the comparison of nested models, to evaluate the number of parameters (npar) of each model. When choosing the best model, we considered support based on ΔAICc, AIC weights, and neg2lnl (Johnson and Omland 2004).
To account for variation or potential bias in the estimates of detectability and survivorship when analyzing the effect of light pollution and noise pollution on apparent survival, we constructed an a priori model for each species that included sex. Territorial males may be more likely to be encountered than females due to behavioral differences (Amrhein et al. 2012); therefore, sex was included as a binary dummy covariate for estimating detectability. We then ranked the a priori model (sex) for each species based on AIC weights with combinations of models that included impervious surface and noise and light levels.
Because model selection and subsequent parameter estimates can become unstable (high variance) by over-fitting models (~10 parameters), especially when there is an insufficient sample size for an individual group variable (Breiman 1996, Burnham and Anderson 2002), we constrained our model set to those in which the parameter estimates were identifiable (Table 3). Here we are referring to extrinsic identifiability, where parameter estimates are at or near their boundary (0 or 1) or are otherwise unidentifiable because of insufficient sample size. Ultimately, we excluded these ‘‘over-parameterized’’ models that contained more parameters than could be accommodated by the data. Therefore, not all covariates could be included in a priori models for each species. We chose to present the model betas from the lowest ΔAICc of each a priori model for each species.
Results
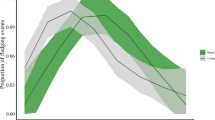
We included 7351 individuals of the seven focal species in the analysis (Table 4). Survival and detection estimates (Fig. 2) varied markedly between males and females and among species (see Table 4). Models that included light pollution and noise pollution variables received some support for five of the seven species (see Table 3).

Estimates (mean symbol) of A annual survival and B detection by species and sex along an urbanization gradient in greater Washington, D.C., USA (2000-2020). See Table 1 for species names by code
There was weak model support for variation in apparent survivorship along a light pollution gradient for American Robin, Gray Catbird, and House Wren. Using beta (Φ) values, we estimated that annual survival probabilities (Fig. 3) increased for American Robin (0.324; se=0.165) and decreased for both Gray Catbird (-0.224; se=0.102) and House Wren (-0.505; se=0.163), as light increased (see Fig. 4).

Predicted annual apparent survival (Φ) probabilities of American Robin, Gray Catbird, and House Wren across a light (lux) pollution gradient in greater Washington, D.C., USA (2000-2020)

Apparent survivorship (Φ) values showing the influence of light on Apparent Survival. Confidence intervals that do not cross zero are considered significant
None of the seven species received model support for variation in apparent survivorship along a sound pollution gradient.
The most competitive model included the quadratic effect of impervious surface for Carolina Chickadee, Carolina Wren, Gray Catbird, House Wren, and Song Sparrow, indicating that survival was greatest at intermediate levels of impervious cover (see Table 3).
Discussion
We identified variation in adult songbird survivorship along the light pollution gradient, with greater apparent survival for one of seven focal species, and lower apparent survival for two of seven species. However, noise pollution does not appear to account for the avian responses to urbanization detected by Evans et al. (2015). Light pollution might account for increased survival of American Robins and decreased survival for House Wrens. Gray Catbird survival was negatively affected by light yet not by impervious surface.
Differences in apparent survival among species along a light pollution gradient may relate to variation in natural history traits among species. For example, light pollution has been determined to affect the singing behaviors of several species (Miller 2006; Francis et al. 2009; Ortega 2012). In areas with light pollution, American Robins initiate their morning chorus earlier (Miller 2006), and the increased time for daily activities such as courtship and foraging may have overall positive effects on robin survival, as we documented in this study. Although our study focused on seven bird species present year-round in the study area, Gray Catbird and House Wren are generally considered short-distance migrants (Butler 2003). Because these two species were the only of the seven to demonstrate reduced apparent survival as light pollution increased, it is possible that migration behaviors of some individuals contributed to elevated vulnerability to light pollution.
Birds that reside in urban areas face numerous human-related threats to survival. In some species, light pollution may lead to decreases in survival by indirectly reducing individual health due to an increase in stress (Gil and Brumm 2013). There is also the possibility of more light increasing the risk of predation at night (Longcore and Rich 2004; Gaston and Bennie 2014). Collisions with windows is a common source of bird mortality, particularly for migrant species, with rough estimates of between 100 million and 1 billion birds killed annually in the United States (Hager and Craig 2014). Buildings are a globally ubiquitous obstacle to avian flight, and collisions with buildings, especially their glass windows, are thought to be a major anthropogenic threat to North American birds (Machtans et al. 2013). Gray Catbirds are vulnerable to window collisions (Loss et al. 2014), which might explain the negative response to light in our study.
The lack of relationship between urban noise and survival may be linked to the generalist nature of the seven focal species. Urban‐adapted species may have greater behavioral, physiological, and ecological flexibility than non‐urban species (Bonier et al. 2007), allowing individuals to survive equally well along the noise gradient. For instance, some species modify or use higher minimum frequency alarm calls in places with higher background noise levels to avoid the concealing effects of urban noise (Lowry et al. 2012; Courter et al. 2020); this sort of phenotypic plasticity in behaviors may allow urban-adapted species to adjust to additional mortality risks associated with noisier environments.
Species-specific responses to urbanization may represent a tradeoff between survival and reproduction (Goodman 1974; Stearns 1992; Charlesworth 1994). If artificial light is a stressor that negatively affects reproduction, it is possible that bird populations compensate with greater survival. If artificial light is a stressor that negatively affects survival, then it is possible that birds compensate with greater reproduction. Senzaki et al. (2020) reported on reproductive success of three of our seven focal species (American Robin, Northern Cardinal, and House Wren). Considering our results in relation to Senzaki et al. (2020), we note that the negative consequences of light emerge within a life-history tradeoff. Thus, positive and negative consequences of light pollution may be obscured from detection within narrowly focused studies due to broader life-history tradeoffs.
Additionally, light pollution, particularly skyglow, extends into rural landscapes, which were underrepresented in this study. Hence, we might have detected a stronger influence of light pollution had the spatial extent of NN sites included more rural locations. Species-specific responses to noise and light pollution need additional study to determine whether the presence or absence of survival costs from noise and light pollution are linked to costs to reproduction.
The relationships between avian survival and characteristics of the urban environment are complex, so teasing out the mechanisms driving the relationships is a challenge. As in our study, impervious surface cover and the measures of noise and light pollution typically are correlated, so controlled experiments (i.e., in a laboratory setting) may be needed to decouple the unique influences of each factor. Additionally, future research on avian responses to artificial light and anthropogenic sound would benefit from finer-scale data that more precisely characterize individual bird exposures to artificial light and anthropogenic sound from location-based sampling. We used relatively coarse-scale geospatial maps of anthropogenic noise and urban light maps produced from models based on land-use and satellite data. Finally, survival analyses could be expanded by including other mapped covariates such as vegetation composition and structure or tree cover. These variables might play key roles in buffering exposure to noise and light pollution.
Anthropogenic accelerations of noise and light pollution are pervasive in that they propagate over all ecosystems on Earth. However, these changes are not geographically or temporally uniform. Urbanization patterns and accompanying transportation routes result in spatial and temporal patterns of noise and light pollution disturbance at multiple scales, including micro-scale pockets of disturbance or refuge, analogous to natural and built structures producing microclimates. Our findings of a correlation between light pollution and adult avian survival for three of seven focal species indicates that responses of different species to urbanization may be related to how easily that species can adjust to local conditions, which can depend on life history traits and/or behavioral plasticity.
Data availability
The data that support the findings of this study are available from co-author Brian Evans; EvansBr@si.edu, upon reasonable request.
References
Akaike H (1973) Information theory and an extension of the maximum likelihood principle. Pages 267–281 in F. Csaki, editor. Second International Symposium on Information Theory. Akamemiai Kiado, Budapest, Hungary
Amrhein V, Scaar B, Baumann M, Minery N, Binnert JP, Korner-Nievergelt F (2012) Estimating adult sex ratios from bird mist netting data. Methods Ecol Evol 3:713–720
Bonier F, Martin PR, Wingfield JC (2007) Urban birds have broader environmental tolerance. Biol Lett 3:670–673.https://doi.org/10.1098/rsbl.2007.0349
Breiman L (1996) Heuristics of instability and stabilization in model selection. Ann Stat 24:2350–2383
Buhaug H and Urdal H (2013) An urbanization bomb? Population growth and social disorder in cities. Glob Environ Chang 23(1):1–10
Burnham KP, Anderson DR (2002) Model selection and multimodel inference: a practical information-theoretic approach, 2nd edn. Springer-Verlag, New York, New York, USA
Butler CJ (2003) The disproportionate effect of global warming on the arrival dates of short-distance migratory birds in North America. Ibis 145(3):484–495
Charlesworth B (1994) Evolution in age-structured populations. Cambridge University Press, Cambridge, UK
Courter JR, Perruci RJ, McGinnis KJ, Rainieri JK (2020) Black-capped chickadees (Poecile atricapillus) alter alarm call duration and peak frequency in response to traffic noise. PLoS One 15(10)
Da Silva A, Samplonius JM, Schlicht E, Valcu M, Kempenaers B (2014) Artificial night lighting rather than traffic noise affects the daily timing of dawn and dusk singing in common European songbirds. Behav Ecol 25(5):1037–1047
Derrickson KC (1988) Condor 90(3):592–606
Dominoni D, Quetting M, Partecke J (2013) Artificial light at night advances avian reproductive physiology. Proc R Soc B Biol Sci 280
Dominoni DM, Halfwerk W, Baird E, Buxton RT, Fernández-Juricic E, Fristrup KM, Barber JR (2020) Why conservation biology can benefit from sensory ecology. Nat Ecol Evol 4(4):502–511
Evans BS, Ryder TB, Reitsma R, Hurlbert AH, Marra PP (2015) Characterizing avian survival along a rural-to-urban land use gradient. Ecology 96(6):1631–1640
Evans-Ogden LJ (2022) Summary report on the bird friendly building program: effect of light reduction on collision of migratory birds. A special report for the Fatal Light Awareness Program (FLAP)
Falchi F, Cinzano P, Duriscoe D, Kyba CC, Elvidge CD, Baugh K, Furgoni R (2016) The new world atlas of artificial night sky brightness. Sci Adv 2(6)
Francis CD (2015) Vocal traits and diet explain avian sensitivities to anthropogenic noise. Glob Chang Biol 21(5):1809–1820
Francis CD, Ortega CP, Cruz A (2009) Noise pollution changes avian communities and species interactions. Curr Biol 19(16):1415–1419
Gaston KJ, Bennie J (2014) Demographic effects of artificial nighttime lighting on animal populations. Environ Rev 22(4):323–330
Gil D, Brumm H (Eds.) (2013) Avian Urban Ecol. OUP Oxford
Goodman D (1974) Natural selection and a cost ceiling on reproductive effort. Am Nat 108:247–268. https://doi.org/10.1086/282906
Hager SB, Craig ME (2014) Bird-window collisions in the summer breeding season. PeerJ 2
Huet des Aunay G et al (2017) Negative impact of urban noise on sexual receptivity and clutch size in female domestic canaries. Ethology 123:843–853
Jiménez-Peñuela J et al (2019) "Urbanization and blood parasite infections affect the body condition of wild birds." Sci Total Environ 651:3015–3022
Johnson JB, Omland KS (2004) Model selection in ecology and evolution. Trends Ecol Evol 19(2):101–108
Kempenaers B, Borgström P, Loës P, Schlicht E, Valcu M (2010) Artificial night lighting affects dawn song, extra-pair siring success, and lay date in songbirds. Curr Biol 20(19):1735–1739
Kleist NJ, Guralnick RP, Cruz A, Francis CD (2016) Anthropogenic noise weakens territorial response to intruder’s songs. Ecosphere 7:1–11
Laake J, Johnson D, Conn P (2013) Marked: An R package for maximum-likelihood and MCMC analysis of capture-recapture data. Methods Ecol Evol 4(9):885–890
La Sorte FA, Fink D, Buler JJ, Farnsworth A, Cabrera-Cruz SA (2017) Seasonal associations with urban light pollution for nocturnally migrating bird populations. Glob Chang Biol 23(11):4609–4619
Lebbin DJ, Harvey MG, Lenz TC, Andersen MJ, Ellis JM (2007) Wilson J Ornithol 119(3):506–508
Liu Z, He C, Zhou Y, Wu J (2014) How much of the world’s land has been urbanized, really? A hierarchical framework for avoiding confusion. Landsc Ecol 29:763–771
Longcore T, Rich C (2004) Ecological light pollution. Front Ecol Environ 2(4):191–198
Loss SR, Will T, Loss SS, Marra PP (2014) Bird–building collisions in the United States: Estimates of annual mortality and species vulnerability. Condor 116(1):8–23
Lowry H, Lill A, Wong BB (2012) How noisy does a noisy miner have to be? Amplitude adjustments of alarm calls in an avian urban ‘adapter.’ PLoS One 7(1)
Machtans CS, Wedeles CHR, Bayne EM (2013) A first estimate for Canada of the number of birds killed by colliding with buildings. Avian Conserv Ecol 8(2):6
Martin TE, Li P (1992) Life history traits of open‐vs. cavity‐nesting birds. Ecology 73(2):579–592
Marzluff JM, Bowman R, Dennelly R (eds) (2001) Avian ecology and conservation in an urbanizing world. Kluwer Academic Publishers, Boston
McKinney ML, Lockwood JL (1999) Biotic homogenization: a few winners replacing many losers in the next mass extinction. Trends Ecol Evol 14:450–453
Mennitt DJ, Fristrup KM (2016) Influence factors and spatiotemporal patterns of environmental sound levels in the contiguous United States. Noise Control Eng J 64(3):342–353
Miller MW (2006) Apparent effects of light pollution on singing behavior of American robins. Condor 108(1):130–139
Nemeth E, Brumm H (2010) Birds and anthropogenic noise: are urban songs adaptive? Am Nat 176(4):465–475
Ortega CP (2012) Chapter 2: Effects of noise pollution on birds: A brief review of our knowledge. Ornithol Monogr 74(1):6–22
Oteiza P, Baldwin MW (2021) Evolution of sensory systems. Curr Opin Neurobiol. 71:52–59. https://doi.org/10.1016/j.conb.2021.08.005. Epub 2021 Sep 29 PMID:34600187
Parkins KL, Elbin SB, Barnes E (2015) Light, glass, and bird—building collisions in an urban park. Northeast Nat 22(1):84–94
Phillips JN, Derryberry EP (2018) Urban sparrows respond to a sexually selected trait with increased aggression in noise. Sci Rep 8:7505
Proppe DS, Sturdy CB, St. Clair CC (2013) Anthropogenic noise decreases urban songbird diversity and may contribute to homogenization. Glob Chang Biol 19(4):1075–1084
Raap T, Casasole G, Costantini D, AbdElgawad H, Asard H, Pinxten R, Eens M (2016) Artificial light at night affects body mass but not oxidative status in free-living nestling songbirds: an experimental study. Sci Rep 6(1):1–8
Senzaki M, Barber JR, Phillips JN, Carter NH, Cooper CB, Ditmer MA, Vukomanovic J (2020) Sensory pollutants alter bird phenology and fitness across a continent. Nature 1–5
Slabbekoorn H, Peet M (2003) Birds sing at a higher pitch in urban noise. Nature 424(6946):267–267
Stearns CC (1992) The evolution of life histories. Oxford University Press, Oxford, UK
Stracey CM, Wynn B, Robinson SK (2014) Light pollution allows the northern mockingbird (Mimus polyglottos) to feed nestlings after dark. Wilson J Ornithol 126(2):366–369
Swaddle JP et al (2015) A framework to assess evolutionary responses to anthropogenic light and sound. Trends Ecol Evol 30:550–560
Van Doren Benjamin M et al (2017) High-intensity urban light installation dramatically alters nocturnal bird migration. Proc Natl Acad Sci 114(42):11175–11180
Ware HE, McClure CJ, Carlisle JD, Barber JR (2015) A phantom road experiment reveals traffic noise is an invisible source of habitat degradation. Proc Natl Acad Sci 112(39):12105–12109
Acknowledgements
We thank The North Carolina Wildlife Federation and the Alongside Wildlife Foundation for providing financial support for this study. We also thank Robert Reitsma, Neighborhood Nestwatch project manager, as well as Neighborhood Nestwatch participants for their commitments to this project.
Funding
This work was supported by the Alongside Wildlife Foundation and The North Carolina Wildlife Federation.
Author information
Authors and Affiliations
Contributions
Lauren D. Pharr, Caren B. Cooper, Brian Evans, and Peter Marra conceived the study. Material preparation, data collection and analysis were performed by Lauren D. Pharr, Caren B. Cooper, Brian Evans, and Jelena Vukomanovic. The first draft of the manuscript was written by Lauren D. Pharr and all authors commented on previous versions of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflicts of interest
The authors have no conflicts of interest to declare that are relevant to the content of this article.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Pharr, L.D., Cooper, C.B., Evans, B. et al. Using citizen science data to investigate annual survival rates of resident birds in relation to noise and light pollution. Urban Ecosyst 26, 1629–1637 (2023). https://doi.org/10.1007/s11252-023-01403-2
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11252-023-01403-2