
Abstract
Providing garden water sources (e.g., ponds, bird baths) has become a popular and strongly promoted form of wildlife-friendly gardening, yet evidence of their use by animals is scarce and limited to a few taxa and water source types. We examined the prevalence, variety and potential value to animal wildlife of supplementary water provided within gardens of Hertfordshire, United Kingdom, using an online questionnaire and field observations of wildlife visitation to urban water sources during summer 2021. Over 70% of 105 questionnaire respondents indicated the presence of at least one water source in their garden and almost 50% had two or more. Bird baths, ground water-bowls and ponds were the most common water source types provided. During 207 h of field observation, we recorded a total of 43 taxa (birds, insects, mammals, amphibians, reptiles) visiting urban lakes and garden birth baths, ponds and ground water-bowls. Taxa richness was similar at urban lakes (30) and garden water sources (27), although approximately 50% of the taxa recorded in each location were unique to that location. Visitation rates of smaller-bodied wildlife did not differ between lakes and gardens, nor among individual water source types. Multivariate analyses indicated insect assemblages visiting lakes did not differ from those visiting garden water sources, and small bird assemblages did not differ among each water source type. These results demonstrate garden water sources, especially for smaller-bodied animals, can supplement the wildlife values contributed by urban lake systems, and should continue to be promoted as an effective conservation action.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
As urbanised land area expands across the globe and human populations within them grow (Seto et al. 2012), natural habitats are becoming increasingly fragmented and degraded (Lambin et al. 2001; McKinney 2006; Burgin et al. 2016). While this has undoubtedly impacted the sustainability of both terrestrial (McKinney 2002) and water dependent (Hill et al. 2015, 2017) biodiversity, there is growing evidence that some wildlife including mammals, reptiles, birds and aquatic invertebrates can persist in both remnant patches of natural habitat (water, vegetation etc.) and highly modified habitats within the urban landscape (Koenig et al. 2001; Stokeld et al. 2014; Hill et al. 2017; Cooper et al. 2020; Van Helden et al. 2020a; 2020b). The capacity of these habitats to support animal wildlife, mostly due to species’ behavioural flexibility (Sol et al. 2013) and ability to exploit the novel resources on offer (Van Helden et al. 2021a; 2021b), has fuelled belief that urban landscapes, and particularly residential gardens, present a significant conservation opportunity for some wildlife (Goddard et al. 2010; Aronson et al. 2017; Soanes and Lentini 2019; Soanes et al. 2019).
One approach to maximise the conservation potential of residential gardens is through wildlife friendly gardening (WFG). Wildlife friendly gardening includes the provision of appropriate food, shelter and water resources in an attempt to support the needs of animal wildlife (Goddard et al. 2010). Associations between the presence of particular garden features (e.g., shelter, food and a lack of natural predators) and animals in gardens provide promising evidence that the manipulation of garden features could offer conservation benefit (e.g., Baker et al. 2003; Daniels and Kirkpatrick 2006; Fontaine et al. 2016; Threlfall et al. 2016; Van Helden et al. 2020b). As knowledge of the factors that influence wildlife presence in urban landscapes has increased, so too has the interest in and uptake of WFG (Gaston et al. 2007; Davies et al. 2009; Cox and Gaston 2018).
This growing interest in WFG is supported by strong promotional programs (see Goddard et al. 2010; Larson et al. 2022) that advocate for the protection, enhancement and addition of wildlife resources in gardens, including the supplementation of water sources. Supplementary provision of water within the residential landscape may particularly benefit conservation, as urban wetlands, lakes and rivers are often only seasonally available to animals (Gaston et al. 2005b), have been strongly altered from their natural state (e.g., Walsh et al. 2005), and the landscape-scale connectivity among them has been reduced by urban land use change (Burgin et al. 2016). In contrast, water sources in gardens are often permanently watered, can be numerous (Gaston et al. 2005b; Davies et al. 2009) and implemented by a substantial proportion of residents (Gaston 2007; Fardell et al. 2022). In the United Kingdom for example, there are an estimated 2.5–3.5 million private garden ponds (Davies et al. 2009) and multiple wildlife conservation programs encourage the public to make water available in their gardens as a means of supporting animal wildlife, especially through periods of hot, dry weather (e.g., RSPB 2021; The Wildlife Trusts 2021).
Animals use water for a multitude of reasons, including habitat, drinking, bathing and reproduction. As human populations within urban areas grow, natural aquatic habitats are increasingly becoming threatened (Walsh et al. 2005; Dudgeon et al. 2006; Vörösmarty et al. 2010; Reid et al. 2019), and their capacity to support biodiversity across multiple functional levels is reduced (Reid et al. 2019). In many regions, climate drying and warming compound these threats, further altering the capacity of natural habitats to support wildlife (Woodward et al. 2010). In areas experiencing both climate warming/drying and urbanisation, novel water sources including natural and constructed water bodies in public spaces and residential gardens have been identified as potentially important for supporting animal wildlife (Chester and Robson 2013; Cleary et al. 2016; Hill et al. 2017). While public blue spaces within urban areas are known to support a variety of animals (Lynn et al. 2006; Hill et al. 2017; Oertli and Parris 2019; Xie et al. 2022), knowledge of the potential value of garden water sources for particular animal taxa is comparatively scarce. The few studies that have explored the use of garden water sources by animal wildlife have concentrated on the use of baths by birds (although with some notable exceptions, see Beebee 1979; Gaston et al. 2005a). These studies demonstrate that a variety of both small and larger-bodied native birds utilise watered bird baths (e.g., Gehlbach 2012; Miller et al. 2015; Cleary et al. 2016), especially those that have adapted to human-dominated habitats (Cleary et al. 2016), and that supplementary water can enhance the value of other wildlife friendly gardening actions such as native plantings (Coetzee et al. 2018) or bird feeding stations (Miller et al. 2015).
While these studies make good progress in evaluating the importance of bird baths, they do not consider the use of other water sources, such as ground water-bowls and ponds, nor the broader range of taxa known to inhabit residential gardens, including mammals (Van Helden et al. 2020b), insects (Jones and Leather 2012), amphibians (Beebee 1979) and reptiles (Koenig et al. 2001). Indeed, evidence that the number and variety of species present in gardens increases with water availability (Fardell et al. 2022) and that garden water sources are used by amphibians (Beebee 1979) and small mammals (Miller et al. 2015), suggests that supplementary water in gardens may benefit a broader array of animal wildlife than is already known. To ensure we are best providing for animal wildlife in the face of climate change in an increasingly urbanised world, it is important to better understand which species use garden water sources, the frequency of use, how these differ among water source types and whether garden water sources support different wildlife compared to larger public blue spaces; one urban water source for which biodiversity value has already been established (Hill et al. 2015, 2017; Chester and Robinson 2013).
We addressed these knowledge gaps using (i) an online questionnaire to establish the prevalence and variety of water sources provided within gardens and (ii) animal wildlife surveys to determine species richness, wildlife visitation, and species assemblages in gardens and small urban lakes in Hertfordshire in South-East England. This study provides evidence of the potential conservation value of garden water sources, and highlights future research required to fully explore and understand their potential use, conservation benefit and application in broader wildlife conservation strategies.
Materials and methods
Study area
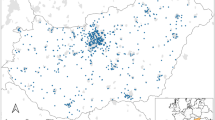
The study was conducted within a 20 km radius of St Albans city, Hertfordshire, South-East England, United Kingdom (Fig. 1). Hertfordshire covers 1643 km2, has a total population of approximately 1.2 million people and a population density of 728 people km− 2 (2020 estimates, ONS 2022). The study area included St Albans and nearby townships of Hemel Hempstead, Hatfield and Harpenden. St Albans supports a population of approximately 149,000 people within an area of 161 km2 (population density = 926 people km− 2) (2020 estimates, ONS 2022).
South-East England is one of the driest and warmest regions in the United Kingdom (Met Office 2016a, b). Both air temperature and precipitation vary seasonally; temperature is highest in July (mean maximum temperature = 22.1 °C) and lowest in January (mean maximum temperature = 7.1 °C), and rainfall is lowest in March (mean total rainfall = 42.7 mm) and highest in October (mean rainfall = 78.2 mm) (Rothamsted climate station; period of record 1991–2020; Met Office 2022a). The United Kingdom is predicted to experience significant climate drying and warming in summer, with rainfall declining by up to 47% and air temperature increasing by up to 5.4 °C by 2070 (Met Office 2022b).

Location of residential gardens (red dot •) and urban lakes (blue dot •) within Hertfordshire County (---), United Kingdom, where wildlife surveys were undertaken during June and July 2021. Shading on the main map indicates a 20 km radius from St Albans within which all survey sites and questionnaire respondents were located. Inset shows location of Hertfordshire (red dashed line ---) north of London (•) within the United Kingdom. Photos illustrate examples of garden water sources including (a) a bird bath, (b) a pond, (c) a ground water-bowl and urban lakes (d-f) where wildlife surveys were undertaken
Questionnaire
An online questionnaire (JISC software; https://www.jisc.ac.uk/about) was developed as part of a broader study (EKG unpublished data) to determine the prevalence and variety of garden water sources, how residents supplemented or modified these sources (i.e., addition of running water, animals and plants), the bird, mammal, amphibian and insect wildlife observed at garden water sources and the willingness of residents to participate in the wildlife observational study. We asked residents to report the number and dimensions of any ‘pond’ (water body constructed at or close to ground level), ‘ground water-bowl’ (bowl of water at ground level), ‘bird bath’ (bowl of water raised above ground level), ‘water feature’ (sculptural or artistic structure with flowing or standing water) and ‘other water source’ (any standing water source not previously defined) that was present in their garden. The survey was circulated via St Albans wildlife groups (Wilderhood Watch) and St Albans Facebook community pages (All Things St Albans, St Albans Eco, St Albans Friends of the Earth), for a period of 8 weeks between 14 June and 10 August 2021. Our release strategy intentionally targeted the ‘conservation-minded’ demographic to increase survey responses from residents who provided supplementary water in their gardens. Given the targeted release strategy, the proportion of respondents that provided supplementary water may not be representative of the broader demographic within the study area.
Wildlife surveys
We undertook timed animal wildlife (hereafter ‘wildlife’) surveys at 13 garden sites (selected from a total of 34 respondents indicating their willingness to undertake wildlife surveys) and six urban lakes within Hertfordshire (N = 19, Fig. 1). Garden sites were selected to include gardens with a pond (N = 5), ground water-bowl (N = 4) or bird bath (N = 4). Urban lakes (size range 200 m2 to 10,000 m2, N = 6) were selected to represent the variety of available urban waterbodies (small, large, natural or constructed). Each lake contained a variety of habitats including open water, reed beds, mud banks and overhanging vegetation. All urban lakes were located within 200 m of a garden site to reduce the potential for species’ geographical ranges to influence differences in taxa visitation among sites.
Wildlife surveys in residential gardens were undertaken by citizen scientists (i.e., the residents) and at urban lakes by an ecological researcher (EKG). At each site, we aimed to complete three, one-hour observations each week for four weeks in June-July 2021 (total = 12 observation hours/site). Citizen scientists were advised to undertake observations at one of their pre-existing garden water sources (bird bath, pond or ground water-bowl) for all observation periods. We were unable to account for variability in the number or type of water sources, additional WFG actions, or garden characteristics that may have also influenced wildlife observations at these sites. Wildlife surveys were undertaken at varying times during daylight hours between 0500 and 2100 h. To standardise methods employed, all citizen scientists were sent an information pack, pre-recorded video and an invitation to a live question and answer session to ensure their understanding of how to complete the wildlife survey. These resources also contained images of wildlife species likely to be encountered by participants during wildlife surveys to increase their accuracy of species identification (Mason and Arathi 2019).
During each observation period, all visits to the focal water source were recorded for birds, mammals, insects, amphibians and reptiles. A visit was defined as an individual interacting directly with the water (i.e., drinking, bathing, resting etc.). Consecutive visits by the same individual within 5 min were recorded as a single visit. Citizen scientists were instructed to observe their water source from approximately 5 m away to minimise disturbance to wildlife whilst allowing accurate identification and recording of species as small as a hoverfly (Syrphidae). At urban lakes, we restricted wildlife observations to a 200 m2 area to standardise wildlife detectability at different sized lakes. The observed area at each urban lake was representative of the diversity and abundance of available habitats. The observer was positioned less than 5 m from the water’s edge with unobstructed vision of the study area using binoculars.
Data analysis
From the full questionnaire developed as part of a broader study, we isolated information on the number, variety and dimensions of water sources, and any modifications that had been made to them, in respondents’ gardens. Any responses that were incomplete or from residences outside the study area (Fig. 1) were removed from analyses. Differences in the volume of each identified water source type was tested using a one-way analysis of variance with post-hoc Tukey multiple comparisons of means using a Bonferroni adjustment on log transformed water volumes. All analyses of questionnaire responses were performed in RStudio, R Statistical Software (V4.1.2; R Core Team 2021) and the significance level was set at alpha = 0.05.
Citizen-derived data on wildlife visitations to garden water sources were screened for accuracy and consistency among individual citizen scientists. As all citizen-derived wildlife survey data were based on focal observation, species identification could not be verified. We accepted most species data, because citizens have been shown to accurately identify morphospecies when given training (Mason and Arathi 2019). To increase reliability of data we made the following adjustments to some species records used for analysis. Records of resident species (e.g., pond snails Lymnaea stagnalis) or any taxa smaller than a hoverfly were omitted from analyses. Due to differences in the use of scientific and common names among citizen scientists, visitation records were recoded to higher taxonomic levels for some birds (e.g., ‘pigeon’ and ‘sparrow’) and some insects (e.g., ‘bees and wasps’, ‘hoverflies’, ‘damselflies’, ‘dragonflies’, and ‘butterflies and moths’). These taxonomic groups were subsequently applied to the visitation data recorded at urban lakes also.
Taxa richness and visitations were standardised by time and expressed as taxa hr− 1 and visits hr− 1 respectively. We also divided the full visitation data matrix (‘all wildlife’) into three data matrices to include ‘small wildlife’, ‘small birds’ and ‘insects’. For each of these subset data matrices, visitation rates of large water dependent birds (e.g., species of geese and ducks) were omitted on the basis that these taxa were unlikely to be able to access garden water sources due to their larger body size. ‘Small wildlife’ therefore included visitation rates for all taxa (except large birds), and ‘small birds’ and ‘insects’ only included visitation rates for small birds and insects respectively. Where possible, we compared wildlife visitation between existing public blue spaces (lakes or wetlands) and gardens with water sources (i.e., between ‘locations’), and between individual ‘water source types’ (i.e., urban lakes, bird baths, ground water-bowls and ponds) due to fundamental differences in size, depth, and position (e.g., elevated versus on ground).
We used a two-sample t-test with equal variance to test for differences in taxa richness and visitation rates among ‘locations’ and a one-way analysis of variance with post-hoc Tukey multiple comparisons of means using a Bonferroni adjustment to test for difference among water source types. For visitation rate, we ran these analyses on ‘all wildlife’ and ‘small wildlife’ data matrices. Both these analyses were undertaken in RStudio, R Statistical Software (V4.1.2; R Core Team 2021) on log-transformed data that met the assumptions of both data normality (Shapiro-Wilk’s p > 0.05) and homogeneity of variance (p > 0.05).
Differences in assemblages of ‘all wildlife’, ‘small wildlife’, ‘small birds’ and ‘insects’ among ‘locations’ and ‘water source types’ were examined using non-metric multidimensional scaling (nMDS) based on Bray Curtis similarity matrices of square-root transformed standardised visitation rates for all taxa in each assemblage type (Primer V7; Clarke and Gorley, 2015). The Analysis of Similarity (ANOSIM) routine in Primer V7 was used to determine significant differences in species assemblages between ‘locations’ and ‘water source types’, and the Similarity Percentages routine (SIMPER) was used to identify the species contributing most (> 5% contribution) to any significant dissimilarities among these groups.
Results
Questionnaire responses
Of the 105 questionnaire responses, 75 indicated the presence of at least one pre-existing water source in their garden. Of a total of 122 supplementary water sources reported, bird baths (n = 38), groundwater bowls (n = 35) and ponds (n = 33) were more common than garden water features (n = 14) and ‘other’ water source types (n = 2) (χ23, N=122 = 11.8, p = 0.008). The total number of all water sources in the gardens varied from one to six (mean = 1.16 ± 0.1 SE) and the proportion of gardens with one (n = 40) and more than one (n = 35) water source was similar. The total volume of supplementary water provided in gardens ranged from 0.45 to 726,000 L (mean = 12071.94 L ± 10072.01 L, N = 72) and the volume of each water source type (not including ‘other’) varied (F3, 99 = 75, p < 0.001); all source types differed from one another (all p < 0.01) except for ground water-bowls and bird baths. The mean volume of bird baths was 12.26 L (± 2.83 L, n = 37), garden water features was 1226.76 L (± 1026.93 L, = 13), ground water-bowls was 7.53 L (± 2.83 L, n = 34) and ponds was 27497.98 L (± 17107.22 L, n = 31).
Modifications were made by some residents to all water source types, except for ground water-bowls. The addition of running water was common for garden water features (78.57% of features; N = 14), although it was also added to some bird baths (7.89%; N = 38) and ponds (21.2%; N = 33). Plants were commonly added to ponds (90.91%) and garden water features (28.57%) whereas animals were only added to ponds (27.27%). Fish (goldfish Carassius auratus, koi carp Cyprinus rubrofuscus and chub Leuciscus cephalus) and frogs (including adults, tadpoles or spawn) were most commonly added to ponds (18.18% and 15.15% of ponds respectively), although newts (Lissotriton vulgaris) and snails (Gastropoda) had also been added to one pond each.
Wildlife surveys
A total of 207 h of observation were undertaken, including 135 h in 12 gardens (mean = 11.25 h site− 1 ± 1.194 SE) and 72 h at six lakes (mean = 12 h site− 1). Bird baths and ground water-bowls were each observed for a total of 40 h and ponds for a total of 55 h (Table 1).
Across all sites we recorded a total of 43 taxa including 31 birds, 7 insects, 2 mammals, 2 amphibians and 1 reptile (Table 1). Of these, five are known exotic species, and two taxonomic groups potentially include exotic species (Table 1). Twenty-seven taxa were recorded in garden water sources; 16 birds, 7 insects, 2 mammal and 2 amphibian taxa. Thirty taxa were recorded at lakes; 23 birds, 4 insects and 1 mammal, amphibian and reptile. Approximately 50% of the species recorded in each location were unique to that location (gardens = 48.15%, N = 27; urban lakes = 53.33%, N = 30). Mean standardised taxa richness (species hr− 1) recorded at lakes was 0.89 species hr− 1 ± 0.08 SE (N = 6), at ponds was 0.52 species hr− 1 ± 0.07 SE (N = 5), at bird baths was 0.95 species hr− 1 ± 0.20 SE (N = 4) and at ground water-bowls was 0.64 species hr− 1 ± 0.12 SE (N = 4). Standardised taxa richness did not vary between locations (mean = 0.69 ± 0.09 SE, N = 13) (t17 = -1.770, p = 0.095) nor among water source types (all pairwise comparisons p > 0.05).
A total of 3505 animal visitations to water sources were recorded during the 207 h of observation. Of these, 2727 visitations (77.80%) were recorded at lakes and 778 visitations (22.20%) were recorded at garden water sources (Table 1). Mean visitation rate of ‘all wildlife’ was significantly higher at lakes (37.88 visits hr− 1± 12.66 SE) compared to gardens with water sources (5.32 visits hr− 1± 1.34 SE) (t17 = -4.791, p < 0.001) and to each garden water source type (F3, 15 = 8.445, p = 0.002; all pairwise comparisons p < 0.05) but did not vary among garden water source types (Fig. 2). Mean visitation rates for ‘small wildlife’ did not vary between locations (lakes: 6.43 visits hr− 1± 1.95 SE; gardens: 37.88 visits hr− 1± 12.66 SE) (t17 = 0.016, p = 0.988) or among water source types (F3, 15 = 0.475, p = 0.705) (Fig. 2).

Mean standardised visitation rate (visits hr-1) of all wildlife (grey bar) and ‘small wildlife’ (white bar) to water source types recorded during wildlife surveys undertaken in June and July 2021 in Hertfordshire, United Kingdom. Whiskers represent SE
Species assemblages at gardens and lakes
Multivariate analyses indicated no difference in insect assemblages between locations (Table 2; Fig. 3d), nor among the ‘small bird’ assemblages visiting each of the water source types (Table 2; Fig. 3c). In contrast, assemblages of ‘all wildlife’, ‘small wildlife’ and ‘small birds’ varied among locations, and assemblages of ‘all wildlife’, ‘small wildlife’ and ‘insects’ differed among water source types (Tables 2, Fig. 3a-d). For these latter wildlife groups, we detected no significant differences among source types in gardens (bird bath, ground water-bowl and pond); only differences between garden water sources and urban lakes.
The difference in the assemblage of ‘all wildlife’ among locations and among water source types (Table 2; Fig. 3a) was driven mostly by the absence of large aquatic birds visiting garden water sources, and the higher visitation rates of damselfly to urban lakes compared to garden water sources (Table 1). The difference in the assemblage of ‘small wildlife’ (Fig. 3b) among locations was influenced most by the visitation of house martin (Delichon urbicum) and black-headed gull (Chroicocephalus ridibundus) to urban lakes only, starlings (Sturnus vulgaris) to gardens only, higher visitation rates of insects (dragonfly and damselfly) to urban lakes and higher visitation rates of pigeons to gardens (Table 1). Differences in visitation of ‘small wildlife’ to urban lakes compared to bird baths and ground water-bowls (Fig. 3b, d) were influenced most by the variations in the visitation of starling, damselfly, black-headed gull, house martin, dragonfly, pigeon, robin and blackbird (Tables 1 and 2).
Differences in the ‘small bird’ assemblages between locations (Fig. 3c) were influenced most by the visitation of house martin and black-headed gull to urban lakes only, the visitation of starlings to gardens only and the higher visitation rates of all other bird species (contributing > 5% to the dissimilarity) to gardens (Table 1). For ‘insects’, differences in visitation to urban lakes and bird baths (Table 2; Fig. 3c) were influenced most by visitation of damselfly and dragonfly to lakes, the visitation of bees/wasps to bird baths only and the higher visitation rate of butterflies/moths to bird baths (Table 1). Differences in visitation of insects to urban lakes and ground water-bowls were influenced mostly by the visitation of damselfly and dragonfly to urban lakes only and the higher visitation rate of bumblebee to groundwater bowls (Table 1).

Non-metric multidimensional scaling (nMDS) plot showing variation in assemblages of (a) ‘all wildlife’ (b) ‘small wildlife’ (c) ‘small birds’ and (d) ‘insects’ between types of water sources in gardens (ground water-bowl (○), bird bath (∆) and pond (□)) and urban lake (•)
Discussion
Aquatic ecosystems are experiencing unprecedented pressure from land and climate change and their value in providing ecosystem and biodiversity services is becoming increasingly compromised (Dudgeon et al. 2006; Vörösmarty et al. 2010; Reid et al. 2019). The provision of supplementary water as a wildlife friendly garden feature may play an important conservation role through increasing the availability of water as well as the connectivity among watered habitats in regions experiencing climate warming and drying, and urban expansion. Beyond the already known prevalence of residential bird baths (Gaston et al. 2007) and ponds (Davies et al. 2009), our study indicates the likely prevalence of other water source types including ground water-bowls and water features that may also provide resources to wildlife. Our wildlife surveys demonstrated that while large-bodied water birds more frequently visited urban lakes, garden water sources were visited by small animal wildlife as frequently as urban lakes, but often by a different assemblage of species. Our results suggest that garden water sources can increase water dependant biodiversity across the urbanised landscape and, that for small-bodied taxa at least, garden water can supplement existing urban blue spaces, and support the diversity of species that use them. Together, our results contribute new evidence that supports the conservation potential of garden water sources.
The provision of supplementary water in gardens has become a popular and strongly promoted form of WFG. It represents an important proportion of WFG actions undertaken by residents (Gaston et al. 2005b, 2007) and has resulted in the establishment of significant water resources across some urban landscapes (Davies et al. 2009). In the United Kingdom for example, up to 3.5 million ponds are estimated to contribute around 349 additional hectares of water surface area across the residential landscape (Davies et al. 2009). Our results suggest these figures likely underestimate the availability of supplementary garden water sources, as we found at least 50% of residents with a water source had more than one source, and residents provided water source types in addition to ponds, including bird baths, ground water-bowls, and water features that have not been incorporated into existing estimates. Given that the engagement in WFG, including water supplementation, varies demographically (Gaston et al. 2007), and the fact that our results are from a survey that targeted a relatively small cohort of conservation-minded participants, investigation of how much additional water may be provided through other water source types warrants further consideration.
The value of supplementary water provision lies in its ability to support wildlife, both locally at the point of source (e.g., Gaston et al. 2005a; Gehlbach 2012; Miller et al. 2015; Cleary et al. 2016), and more broadly through providing connectivity among the variety of water sources that occur across the urban landscape (sensu Goddard et al. 2010). Interestingly, despite strong advocacy for the provision of garden water sources (Goddard et al. 2010; Larson et al. 2022), and a growing body of evidence for the conservation value of garden habitats for terrestrial fauna (see for example Goddard et al. 2010; Maclagan et al. 2018; Van Helden et al. 2020b; 2021b; Gazzard et al. 2022), there remains a scarcity of quantitative evidence of the conservation value of garden water sources (Gaston et al. 2007; Davies et al. 2009). Until now, most studies have explored the use of bird baths by birds (Gehlbach 2012; Miller et al. 2015; Cleary et al. 2016), although some have also looked at the biodiversity value of ponds (Gaston et al. 2005a) or incidentally recorded the use of garden water sources by other taxa (Beebee 1979; Miller et al. 2015). Our data show that a variety of garden water sources are used by multiple taxa including birds, insects, amphibians, reptiles and mammals and that species richness and visitations are not influenced by source type, and also do not vary between gardens and lakes for small-bodied taxa at least. Our results suggest that garden water sources can elevate water dependant biodiversity across the urbanised landscape and provide additional resources to the suite of wildlife already occurring in established blue spaces.
Interestingly, respondents to our questionnaire reported making modifications to the water sources within their gardens including the addition of plants, running water and a variety of animal life. Habitat diversity and complexity strongly influences aquatic community structure across all levels of organisation (Soukup et al. 2022), and indirectly communities of terrestrial fauna that rely on aquatic food sources (e.g., Dahlin et al. 2021). Although it remains unexplored in the broader literature, whether modification of garden water sources provide additional biodiversity value is a worthy line of future research. Given the importance of behavioural flexibility to wildlife’s capacity to exploit novel resources within residential gardens (Sol et al. 2013; Van Helden et al. 2021b), understanding how different wildlife taxa utilise garden water sources may also provide insight into their value, as well as their design.
Despite significant growth in urban wildlife research over the last two decades, there remains a focus on few ‘fields of study’ and taxa, predominantly birds and mammals (Magle et al. 2012; Collins et al. 2021). In our study, we focussed on a seldom explored field of aquatic urban wildlife research and attempted to broaden the taxonomic focus to include ‘all wildlife’. Due to difficulties in accessing privately owned land, our approach was to engage citizen scientists for monitoring garden water sources, as their involvement has proven beneficial across a diversity of ecological research programs (Gallo-Cajiao et al. 2018; Paloniemi et al. 2018; Steven et al. 2019, 2021; Lloyd et al. 2020). We see several components of our approach that could be improved in future studies, specifically our reliance on observational wildlife surveys, undertaken by citizen scientists in one season, only during daylight hours and in a relatively small sample of garden and urban wetland sites. Despite residents being supplied with training materials and support, we found variation in survey ability among residents particularly for small, cryptic species. In relation to our study, this affected recording of some animal species (some birds and some insects) that we combined into higher taxonomic classifications due to inconsistencies in nomenclature and species discrimination. While focal observations can provide quantitative information on some taxa (e.g., birds), this approach is unlikely to provide comprehensive data on semi aquatic and aquatic fauna. Wildlife such as these often require sampling techniques other than observation, including animal handling rendering their utility in citizen-based sampling programs problematic due to animal welfare issues. While our focus on visitations of wildlife to water sources during summer, when wildlife demand for water is greatest, is ecologically relevant, exploring seasonal variability in the use of garden water sources (e.g., Van Helden et al. 2021a) would provide a more comprehensive understanding of their biodiversity value. All wildlife surveys were conducted during daylight hours and therefore potentially missed other nocturnal and crepuscular species. Future research could utilise additional sampling methods, such as motion triggered cameras, that have proven beneficial to other garden-based wildlife surveys (Van Helden et al. 2020b) and are suitable for deployment by citizen scientists.
Conclusions
This study represents one of very few quantitative studies demonstrating anthropogenic water sources in domestic gardens are utilised by a variety of animal taxa. We demonstrate that these water sources can both increase biodiversity within urban landscapes and provide additional water sources to support the suite of taxa already utilising urban blue spaces. Our results provide an important baseline on which future investigations can expand examination of the biodiversity value of garden water sources for all animal wildlife, how the use of these water sources varies temporally, the array of behaviours and requirements they support, and how they interact with other garden and landscape scale components of urban conservation strategies.
References
Aronson MFJ, Lepczyk CA, Evans KL, Goddard MA, Lerman SB, MacIvor JS, Nilon CH, Vargo T (2017) Biodiversity in the city: key challenges for urban green space management. Front Ecol Environ 15:189–196. https://doi.org/10.1002/fee.1480
Baker PJ, Ansell RJ, Dobbs PAA, Weber CE, Harris S (2003) Factors affecting the distribution of small mammals in an urban area. Mamm Rev 33:95–100. https://doi.org/10.1046/j.1365-2907.2003.00003.x
Beebee TJC (1979) Habitats of the british amphibians (2): suburban parks and gardens. Biol Conserv 15:241–257. https://doi.org/10.1016/0006-3207(79)90046-6
Burgin S, Franklin MJM, Hull L (2016) Wetland loss in the transition to urbanisation: a case study from western Sydney, Australia. Wetlands 36:985–994. https://doi.org/10.1007/s13157-016-0813-0
Chester ET, Robson BJ (2013) Anthropogenic refuges for freshwater biodiversity: their ecological characteristics and management. Biol Conserv 166:64–75. https://doi.org/10.1016/j.biocon.2013.06.016
Clarke KR, Gorley RN (2015) PRIMER v7: User manual/tutorial. PRIMER-EPlymouth
Cleary GP, Parsons H, Davis A, Coleman BR, Jones DN, Miller KK, Weston MA (2016) Avian assemblages at bird baths: a comparison of urban and rural bird baths in Australia. PLoS ONE 11:e0150899. https://doi.org/10.1371/journal.pone.0150899
Coetzee A, Barnard P, Pauw A (2018) Urban nectarivorous bird communities in Cape Town, South Africa, are structured by ecological generalisation and resource distribution. J Avian Biol 49:jav–01526. https://doi.org/10.1111/jav.01526
Collins MK, Magle SB, Gallo T (2021) Global trends in urban wildlife ecology and conservation. Biol Conserv 261:2021–109236. https://doi.org/10.1016/j.biocon.2021.109236
Cooper DS, Shultz AJ, Blumstein DT (2020) Temporally separated data sets reveal similar traits of birds persisting in a United States megacity. Front Ecol Evol. https://doi.org/10.3389/fevo.2020.00251
Cox DTC, Gaston KJ (2018) Human-nature interactions and the consequences and drivers of provisioning wildlife. Philos Trans R Soc Lond B Biol Sci. 2018 May 5;373(1745):20170092. https://doi.org/10.1098/rstb.2017.0092
Dahlin KM, Zarnetske PL, Read QD, Twardochleb LA, Kamoske AG, Cheruvelil KS, Soranno PA (2021) Linking terrestrial and aquatic biodiversity to ecosystem function across scales, trophic levels, and realms. Front Environ Scie 9:692401. https://doi.org/10.3389/fenvs.2021.692401
Daniels GD, Kirkpatrick JB (2006) Does variation in garden characteristics influence the conservation of birds in suburbia? Biol Cons 133:326–335. https://doi.org/10.1016/j.biocon.2006.06.011
Davies ZG, Fuller RA, Loram A, Irvine KN, Sims V, Gaston KJ (2009) A national scale inventory of resource provision for biodiversity within domestic gardens. Biol Conserv 142:761–771. https://doi.org/10.1016/j.biocon.2008.12.016
Dudgeon D, Arthington A, Gessner M, Kawabata Z, Knowler D, Lévêque C, Naiman RJ, Prieur-Richard A-H, Soto D, Stiassny MLJ, Sullivan CA (2006) Freshwater biodiversity: importance, threats, status and conservation challenges. Biol Rev 81:163–182. https://doi.org/10.1017/S1464793105006950
Fardell LL, Pavey CR, Dickman CR (2022) Backyard biomes: is anyone there? Improving public awareness of urban wildlife activity. Diversity 14:263. https://doi.org/10.3390/d14040263
Fontaine B, Bergerot B, Le Viol I, Julliard R (2016) Impact of urbanization and gardening practices on common butterfly communities in France. Ecol Evol 6:8174–8180. https://doi.org/10.1002/ece3.2526
Gallo-Cajiao E, Archibald C, Friedman R, Steven R, Fuller RA, Game ET, Morrison TH, Ritchie EG (2018) Crowdfunding biodiversity conservation. https://doi.org/10.1111/cobi.13144. Conserv biol doi
Gaston KJ, Smith RM, Thompson K, Warren PH (2005a) Urban domestic gardens (II): experimental tests of methods for increasing biodiversity. Biodivers Conserv 14:395. https://doi.org/10.1007/s10531-004-6066-x
Gaston KJ, Warren PH, Thompson K, Smith RM (2005b) Urban domestic gardens (IV): the extent of the resource and its associated features. Biodivers Conserv 14:3327–3349. https://doi.org/10.1007/s10531-004-9513-9
Gaston KJ, Fuller RA, Loram A, MacDonald C, Power S, Dempsey N (2007) Urban domestic gardens (XI): variation in urban wildlife gardening in the United Kingdom. Biodivers Conserv 16:3227–3238. https://doi.org/10.1007/s10531-007-9174-6
Gazzard A, Yarnell RW, Baker PJ (2022) Fine-scale habitat selection of a small mammalian urban adapter: the West European hedgehog (Erinaceus europaeus). Mamm Biol (2022). https://doi.org/10.1007/s42991-022-00251-5
Gehlbach FR (2012) Eastern screech-owl responses to suburban sprawl, warmer climate, and additional avian food in central Texas. Wilson J Ornithol 124:630–633. http://www.jstor.org/stable/23324575
Goddard MA, Dougill AJ, Benton TG (2010) Scaling up from gardens: biodiversity conservation in urban environments. Trends Ecol Evol 25:90–98. https://doi.org/10.1016/j.tree.2009.07.016
Hill MJ, Mathers KL, Wood PJ (2015) The aquatic macroinvertebrate biodiversity of urban ponds in a medium-sized european town (Loughborough, UK). Hydrobiol 760:225–238. https://doi.org/10.1007/s10750-015-2328-8
Hill MJ, Biggs J, Thornhill I, Briers RA, Gledhill DG, White JC, Wood PJ, Hassall C (2017) Urban ponds as an aquatic biodiversity resource in modified landscapes. Glob Change Biol 23:986–999. https://doi.org/10.1111/gcb.13401
Jones LE, Leather RS (2012) Invertebrates in urban areas: a review. Eur J Entomol 109:463–478. https://doi.org/10.14411/eje.2012.060
Koenig J, Shine R, Shea G (2001) The ecology of an australian reptile icon: how do blue-tongued lizards (Tiliqua scincoides) survive in suburbia? Wildl Res 28:214–227. https://doi.org/10.1071/WR00068
Lambin EF, Turner BL, Geist HJ, Agbola SB, Angelsen A, Bruce JW, Coomes OT, Dirzo R, Fischer G, Folke C, George PS, Homewood, Imbernon J, Leemans R, Li X, Moran EF, Mortimore M, Ramakrishnan PS, Richards JF, Skånes H, Steffen W, Stone GD, Svedin U, Veldkamp TA, Vogel C, Xu J (2001) The causes of land-use and land-cover change: moving beyond the myths. Glob Enviro Change 11:261–269. https://doi.org/10.1016/S0959-3780(01)00007-3
Larson KL, Lerman SB, Nelson KC, Narango DL, Wheeler MM, Groffman PM, Hall SJ, Grove JM (2022) Examining the potential to expand wildlife-supporting residential yards and gardens. https://doi.org/10.1016/j.landurbplan.2022.104396. Landsc Urban Plan 222
Lloyd TJ, Fuller RA, Oliver JL, Tulloch AI, Barnes M, Steven R (2020) Estimating the spatial coverage of citizen science for monitoring threatened species. Glob Ecol Conserv. https://doi.org/10.1016/j.gecco.2020.e01048
Lynn JC, Chambers CL, Rosenstock SS (2006) Use of wildlife water developments by birds in southwest Arizona during migration. Wildl Soc Bull 34:592–601. https://doi.org/10.2193/0091-7648(2006)34[592:UOWWDB]2.0.CO;2
Maclagan SJ, Coates T, Ritchie EG (2018) Don’t judge habitat on its novelty: assessing the value of novel habitats for an endangered mammal in a peri-urban landscape. Biol Conserv 223:11–18. https://doi.org/10.1016/j.biocon.2018.04.022
Magle SB, Hunt VM, Vernon M, Crooks KR (2012) Urban wildlife research: past, present, and future. Biol Conser 155:23–32. https://doi.org/10.1016/j.biocon.2012.06.018
Mason L, Arathi HS (2019) Assessing the efficacy of citizen scientists monitoring native bees in urban areas. Glob Ecol Conserv 17:e00561
McKinney ML (2002) Urbanization, biodiversity, and conservation: the impacts of urbanization on native species are poorly studied, but educating a highly urbanized human population about these impacts can greatly improve species conservation in all ecosystems. Bioscience 52:883–890. https://doi.org/10.1641/0006-3568(2002)052[0883:UBAC]2.0.CO;2
McKinney ML (2006) Urbanization as a major cause of biotic homogenization. Biol Conserv 127:247–260. https://doi.org/10.1016/j.biocon.2005.09.005
Met Office (2016a) Eastern England: climate. https://www.metoffice.gov.uk/binaries/content/assets/metofficegovuk/pdf/weather/learn-about/uk-past-events/regional-climates/eastern-england_-climate---met-office.pdf. Accessed 4 May 2022
Met Office (2022a) UK climate averages: Rothamsted (Hertfordshire). https://www.metoffice.gov.uk/research/climate/maps-and-data/uk-climate-averages/gcpwxz1ey. Accessed 4 May 2022
Met Office (2016b) Southern England: climate https://www.metoffice.gov.uk/binaries/content/assets/metofficegovuk/pdf/weather/learn-about/uk-past-events/regional-climates/southern-england_-climate---met-office.pdf. Accessed 4 May 2022
Met Office (2022b) UKCP18 science overview, executive summary January 2019. https://www.metoffice.gov.uk/binaries/content/assets/metofficegovuk/pdf/research/ukcp/ukcp18-overview-summary.pdf. Accessed 4 May 2022
Miller KK, Blaszczynski VN, Weston MA (2015) Feeding wild birds in gardens: a test of water versus food. Ecol Manag Restor 16:156–158. https://doi.org/10.1111/emr.12157
Oertli B, Parris KM (2019) Review: toward management of urban ponds for freshwater biodiversity. Ecosphere 10:e02810. https://doi.org/10.1002/ecs2.2810
ONS (2022) Office of national statistics: population and migration. https://www.ons.gov.uk/peoplepopulationandcommunity/populationandmigration. Accessed 4 May 2022
Paloniemi R, Hujala T, Rantala S, Harlio A, Salomaa A, Primmer E, Arponen A (2018) Integrating social and ecological knowledge for targeting voluntary biodiversity conservation. Conserv Lett 11:e12340. https://doi.org/10.1111/conl.12340
R Core Team (2021) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Reid AJ, Carlson AK, Cree IF, Eliason EJ, Gell PA, Johnson PTJ, Kidd KA, MacCormack TJ, Olden JD, Ormerod SJ, Smol JP, Taylor WW, Tockner K, Vermaire JC, Dudgeon D, Cooke SJ (2019) Emerging threats and persistent conservation challenges for freshwater biodiversity. Biol Rev 94:849–873. https://doi.org/10.1111/brv.12480
RSPB (2021) How to provide water for wildlife. The Royal Society for the Protection of Birds. https://www.rspb.org.uk/birds-and-wildlife/advice/gardening-for-wildlife/water-for-wildlife/ Accessed 4 May 2022
Seto KC, Güneralp B, Hutyra LR (2012) Global forecasts of urban expansion to 2030 and direct impacts on biodiversity and carbon pools. Proc Natl Acad Sci 109:16083–16088. https://doi.org/10.1073/pnas.1211658109
Soanes K, Lentini PE (2019) When cities are the last chance for saving species. Front Ecol Environ 17:225–231. https://doi.org/10.1002/fee.2032
Soanes K, Sievers M, Chee YE, Williams NSG, Bhardwaj M, Marshall AJ, Parris KM (2019) Correcting common misconceptions to inspire conservation action in urban environments. Cons Biol 33:300–306. https://doi.org/10.1111/cobi.13193
Sol D, Lapiedra O, González-Lagos C (2013) Behavioural adjustments for a life in the city. Anim Behav 85:1101–1112. https://doi.org/10.1016/j.anbehav.2013.01.023
Soukup PR, Näslund J, Höjesjö J, Boukal DS (2022) From individuals to communities: Habitat complexity affects all levels of organization in aquatic environments. Wiley Interdisciplinary Reviews: Water 9:e1575. https://doi.org/10.1002/wat2.1575
Steven R, Barnes M, Garnett ST, Garrard G, O’Connor J, Oliver JL, Fuller RA (2019) Aligning citizen science with best practice: threatened species conservation in Australia. Conserv Sci Pract 1:e100. https://doi.org/10.1111/csp2.100
Steven R, Van Helden BE, Tulloch A, Barnes M, Close PG, Fuller RA (2021) Exploring the ability of urban householders to correctly identify nocturnal mammals. Urban Ecosyst 24:1359–1369. https://doi.org/10.1007/s11252-021-01118-2
Stokeld D, Hamer AJ, van der Ree R, Pettigrove V, Gillespie G (2014) Factors influencing occurrence of a freshwater turtle in an urban landscape: a resilient species? Wildl Res 41:163–171. https://doi.org/10.1071/WR13205
The Wildlife Trusts (2021) How to provide water for wildlife. https://www.wildlifetrusts.org/actions/how-provide-water-wildlife Accessed 4 May 2022
Threlfall CG, Williams NSG, Hahs AK, Livesley SJ (2016) Approaches to urban vegetation management and the impacts on urban bird and bat assemblages. Landsc Urban Plann 153:28–39. https://doi.org/10.1016/j.landurbplan.2016.04.011
Van Helden BE, Close PG, Steven R (2020a) Mammal conservation in a changing world: can urban gardens play a role? Urban Ecosyst 23:555–567. https://doi.org/10.1007/s11252-020-00935-1
Van Helden BE, Close PG, Stewart BA, Speldewinde PC, Comer SJ (2020b) An underrated habitat: residential gardens support similar mammal assemblages to urban remnant vegetation. Biol Conserv 250:108760. https://doi.org/10.1016/j.biocon.2020.108760
Van Helden BE, Close PG, Stewart BA, Speldewinde PC (2021a) Managing gardens for wildlife: garden features that predict mammal presence and abundance in gardens vary seasonally. Ecosphere 12:e03453. https://doi.org/10.1002/ecs2.3453
Van Helden BE, Close PG, Stewart BA, Speldewinde PC, Comer SJ (2021b) Critically endangered marsupial calls residential gardens home. Anim Conser 24:445–456
Vörösmarty CJ, McIntyre PB, Gessner MO, Dudgeon D, Prusevich A, Green P, Glidden S, Bunn SE, Sullivan CA, Reidy Liermann C, Davies PM (2010) Global threats to human water security and river biodiversity. Nature 467:555–561. https://doi.org/10.1038/nature09440
Walsh CJ, Roy AH, Feminella JW, Cottingham PD, Groffman PM, Morgan IIRP (2005) The urban stream syndrome: current knowledge and the search for a cure. J North Am Benthol Soc 24:706–723. https://doi.org/10.1899/04-028.1
Woodward G, Perkins DM, Brown LE (2010) Climate change and freshwater ecosystems: impacts across multiple levels of organization. Philos Trans R Soc Lond B Biol Sci 365:2093–2106. https://doi.org/10.1098/rstb.2010.0055PMID: 20513717; PMCID: PMC2880135
Xie S, Marzluff JM, Su Y, Wang Y, Meng N, Wu T, Gong C, Lu F, Xian C, Zhang Y, Ouyang Z (2022) The role of urban waterbodies in maintaining bird species diversity within built area of Beijing. Sci Total Environ 806:150430. https://doi.org/10.1016/j.scitotenv.2021.150430
Acknowledgements
We thank residents of Hertfordshire for their enthusiastic participation in our research as citizen scientists. We thank Wilderhood Watch, All Things St Albans, St Albans Eco and St Albans Friends of the Earth community groups for promoting our project, particularly the questionnaire to the residents of Hertfordshire. We thank Dr Erik Kiviat and an additional anonymous reviewer for their helpful suggestions on an earlier version of this manuscript.
Funding
Open Access funding enabled and organized by CAUL and its Member Institutions. This work received administrative research support from The University of Western Australia and Bristol University. BVH was supported through the Australian Government’s Inspiring Australia Science Engagement Program.
Author information
Authors and Affiliations
Contributions
Conceived and designed the research: EKG, PGC, BVH, NJR. Collected the data: EKG. Prepared the data for analysis: PGC, EKG. Performed statistical analysis: PGC. Wrote the manuscript: PGC, EKG. Reviewed and revised the manuscript: PGC, EKG, BVH, NJR. Approved final manuscript: PGC, EKG, BVH, NJR. Supervised the project: PGC, BVH, NJR.
Corresponding author
Ethics declarations
Ethics approval
This research was approved by the Human Ethics Committees of Bristol University and The University of Western Australia (2021/ET000595), and the Animal Ethics Committees of Bristol University (UIN/21/020) and The University of Western Australia (F18979).
Consent to participate
Participation of citizen scientists in this research was voluntary. Participation was supported by a Participant Information Form for the questionnaire and training information and opportunities for the wildlife survey. Completion of each component was considered evidence of consent to participate.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Gibbons, E.K., Close, P.G., Van Helden, B.E. et al. Water in the city: visitation of animal wildlife to garden water sources and urban lakes. Urban Ecosyst 26, 1413–1425 (2023). https://doi.org/10.1007/s11252-023-01391-3
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11252-023-01391-3