
Abstract
The negative consequences of fragmentation, infrastructure development, and urbanization on biodiversity are well known. However, careful urban planning can provide viable habitat for some species and communities. Generally, r-selected species are more likely to persist in urbanizing landscapes, while long lived, mobile species, such as turtles, are likely to decline toward extinction. Understanding species responses across urban gradients is important for developing mitigation planning. We used VHF telemetry to quantify ranging behavior of three-toed box turtles in fragmented urban forest patches and a semi-contiguous large rural forest in Saint Louis, Missouri. We then simulated movement trajectories based on empirical data from the two turtles with the largest and smallest home ranges, overlaid on forest cover and road network maps of Missouri, to quantify the state-wide probability of turtles encountering roads. Home range (HR) size varied from 1–250Ha. The mean home range estimate of rural turtles was > 9 times larger than that of urban turtles. Simulations indicated that the least mobile turtle (HR 1Ha) would have a 22% likelihood of encountering a road if placed randomly in suitable habitat anywhere in the state. The likelihood increased to 90% for the most mobile turtle (HR 250Ha), with 10 road crossings per year. High turtle mortality, range restriction and population fragmentation are likely even in rural areas. In urban cityscapes, large, roadless forest patches offer the highest conservation potential for this species.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
The overwhelming impacts of wilderness fragmentation and loss, infrastructure development, urbanization, and other land use changes on biodiversity from local to global extents are well known (e.g. Forman and Alexander 1998, Pickett et al. 2001, McKinney 2002, Laurance et al. 2014, Torres et al. 2016). Roads are often the first infrastructural development into remote areas, a superstructure that paves the way for the tsunami of development that often follows (Soares-Filho et al. 2004; Laurance et al. 2006). Alteration of land cover under industrial scale exploitation for natural resource extraction and agriculture, proliferation of road and rail infrastructure, and growth of urban and suburban areas are viewed as essential for economic growth (Fan and Chan-Kang 2008; Percoco 2016) and are encouraged by national governments and international development agencies (Wilkie et al. 2000). Globally urban area is likely to nearly triple from 2000 levels by 2030 (Seto et al. 2012), and some 25 million km of new roads will be constructed between 2010 and 2050, a 60% increase since 2010 (Laurance et al. 2014).
While the growing intensity of the human footprint is often catastrophic for biodiversity, cities are novel habitats that can offer opportunities for biodiversity conservation (Goddard et al. 2010) and research on mechanisms of evolution (Lambert and Donihue 2020). How species and communities respond to urbanization is highly variable and depends in part on life history strategies, physiology, ecology, and the characteristics of the urban environment and surrounding matrix of land uses (Johnson and Munshi-South 2017; Jørgensen et al. 2019). Generally, long-lived species with long generation times, such as turtles, have lower adaptive potential to rapid change, thus as landscapes become urbanized these species face higher local extinction probability than r-selected species (McDonnell and Hahs 2015). Large mobile species that occur at low population density are often the hardest hit by anthropogenic change (e.g. Blake et al. 2008, Rytwinski and Fahrig 2012). However, populations of small animals may also be subjected to serious declines in abundance and increased risk of extinction (Oxley et al. 1974; Gibbs 1998; Andrews et al. 2015). The most vulnerable small animals include long-lived terrestrial species that have long generation times, low fecundity, variable recruitment rates and mobile movement patterns (Gibbs and Shriver 2002; Gibbs and Steen 2005). Many of the 356 species of turtles and tortoises (Rhodin et al. 2017) (Order: Testudines) share these life history traits, and it is no surprise that Testudines is the most threatened vertebrate order. Half of all known species are listed as “vulnerable” and 40 species as “critically endangered” by the International Union for the Conservation of Nature (IUCN. 2016).
Globally, the United States has the highest turtle species richness of any country, with 53 species (33 endemic) and 82 recognized taxa (57 endemic) (Mittermeier et al. 2015). Regionally, the Southeastern US contains 42 species of turtles and tortoises of which 11 are endemic, a species richness only surpassed by Indo Burma (Mittermeier et al. 2015). Thus, the second most important region globally for conservation of one of Earth’s most threatened vertebrate species groups is also that which is predicted to experience among the greatest levels of land use change on the North American continent in coming decades (Alig et al. 2004).
North American common box turtles (Terrapene spp.) were once ubiquitous across a wide variety of habitats throughout the eastern and southern United States but are now in decline (Stickel 1978; Budischak et al. 2006; Dodd et al. 2006; Rees et al. 2009) and listed as vulnerable by the IUCN Red List (van Dijk 2016). Along with habitat loss to agriculture and increased risk from disease (Palmer et al. 2016; Agha et al. 2017), urban expansion likely plays a role in this decline. The three-toed box turtle (T. mexicana triunguis, TTBT) occurs west of the Mississippi River, and inhabits oak-hickory forests including the Ozark forests of Missouri and Arkansas. These states contain some of the largest patches of National Forest within the eastern United States. While predictions to the year 2050 suggest that these tracts of forest will remain relatively stable (Nowak and Walton 2005), there will be considerable urban expansion into less remote forested areas. In Missouri, the non-urban forest area is expected to decline by 3.4% (2,212km2), while forests situated in urban landscapes will increase from 0.9% to 1990 to 4.5% by 2050 (Nowak and Greenfield 2018). This growing urban-forest interface will present challenges to the survival of TTBT and other wildlife in forest fragments that remain in urban environments. Using simulated movement patterns Gibbs and Shriver (2002) estimated that mortality due to roads among a suite of turtle species was dependent on body size and movement patterns. Large land turtles experienced higher road mortality than smaller pond turtles due to their larger home ranges. Based on simulations, small pond turtles rarely encountered roads. Female pond turtles have larger home ranges than males and have higher mortality due to roads than males. Declines in pond turtle abundance as road density increases are likely due to road mortality effects on females (Patrick and Gibbs 2010). If home range size is a determinant of rates of road mortality, turtle species with small home ranges may be less vulnerable to the negative effects of roads. However, the abundance of box turtles, which have small home ranges of just several hectares (Dodd 2001) also declines with road density (Erb et al. 2015).
Here, we investigate the home range sizes of TTBT in an urban site in St. Louis City and a rural site in St. Louis County. We hypothesized that turtles living in large contiguous forest would have larger home ranges than turtles in small urban forest fragments. The basis for our prediction is that TTBTs are forest specialists and therefore size of forest fragment would influence home range size. We extrapolate the movement data to estimate how current habitat area and road infrastructure may impact turtle vulnerability at the state level. Finally, we discuss how projected land use change may impact box turtles in Missouri and throughout their range.
Methods
Study site
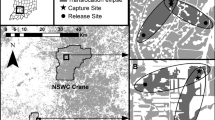
We selected two sites along the urban-rural gradient of the St. Louis metropolitan area in Missouri (Fig. 1). At 526 ha, Forest Park (Latitude N38.63906, Longitude W90.28494, Fig. 1C) is one of the largest urban parks in the US and is centrally located within the city. It comprises a mosaic of land uses including two museums, the Saint Louis Zoo, a theater, ice rink, golf courses and sports fields, numerous fragments of oak-hickory forest, prairies, and waterbodies in various states of restoration. A network of public roads, footpaths and bike trails traverse the unfenced park. Topography is relatively flat, and largely the result of landscaping. Forest fragments vary in size from several hundred square meters to the 24 ha Kennedy Forest. Over 13 million people visit Forest Park annually.

Saint Louis Box Turtle Project study sites (A), illustrating details of turtle movements at Tyson Research Center (B), and Forest Park (C). Dots represent relocations of VHF-tagged three-toed box turtles, each color represents a different individual
Tyson Research Center (Tyson) (Latitude N38.52733, Longitude W90.56026) was chosen as the rural site (Fig. 1B). Covering 809 ha, Tyson is found on the southwest edge of greater St. Louis urban development and marks the nearest large expanse of forest cover to the southwest of the city. Located on the northeastern edge of the Ozark ecoregion, Tyson is topographically rugged and dominated by oak-hickory forest, but also contains several restored/reconstructed glades and prairies. During the early 1900s, the property was grazed and selectively logged. In 1942, it was taken over by the U.S. military for munitions storage and purchased in 1962 by Washington University in St. Louis for use as a field research station (Zimmerman and Wagner 1979). Tyson’s vegetation has been relatively undisturbed for ca. 80 years. Soil types include silty loams and silty clays that develop from shale limestone, limestone, cherty limestone and chert formations (Zimmerman and Wagner 1979). Ecological research, education and conservation are the primary activities at Tyson. Tyson is bisected by a single paved road with numerous gravel forest roads, mostly on ridge tops. A lightly trafficked paved road forms the northern boundary of Tyson, while Interstate 44 marks the southern boundary. County parks to the east and west extend the contiguous forest area to ca. 1220 ha.
Turtle movements
Turtle relocation data were collected between March 2012 and July 2015 (Fig. 1B, C). Turtles found during searches on foot at both sites were fitted with Very High Frequency (VHF) radio transmitters (Holohil Transmitters, Carp, Ontario, Canada, models RI-2B and RI-2 C). The transmitters, which weighed between 10 and 16 g, were glued to the carapace on the second vertebral scute using plumber’s epoxy. The weight of the transmitter plus epoxy was never more than 5% of turtle body weight. Tagged turtles were relocated every 1–2 weeks, using a TRX-16 radio receiver with a three element Yagi antenna (Wildlife Materials, Murpheysboro, Illinios, USA). The time encountered was recorded and a location coordinates were taken with a Garmin eTrex Venture (Garmin International Inc. Olathe, Kansas, USA). Details of habitat including forest type and understory characteristics were recorded including open or closed canopy oak hickory forest, open or closed understory, wooded prairie, and grassland.
Data analysis
GPS coordinates were projected into UTM shapefiles in ArcGIS 10.1. Turtle movement metrics, including Minimum Convex Polygon (MCP) home ranges, step lengths and turn angles were calculated using the Geospatial Modelling Environment (GME) (Beyer 2012). We plotted MCP area against number of relocations (in multiples of five) for each turtle and only selected those individuals for which the MCP areas reached an asymptote. For this sample of individuals we also calculated 50%, 75% and 95% Utilization Distributions using the function kernelUD in the adehabitatHR package in r (version 4.2.1) (R Development Core Team 2022).
Simulating turtle movements at site level and across Missouri
To determine the potential for road crossings and excursions out of the study areas at the site level we generated plausible simulations of turtle movements. We first selected all relocation data from the turtles with the largest and smallest three-year cumulative home ranges (Table 1) at each site. From the relocation data, step length and turn angles were calculated for each time step (usually but not always at weekly intervals during the active season). For each turtle, we then generated 100 unconstrained simple correlated random walks (CRW) each consisting of 100 steps using GME software (Beyer 2012). Each CRW started from the initial location of each turtle. For each turtle, we calculated the proportion of the 100 CRWs that crossed either a public road or the study site boundary using the “select by location” command of ArcGIS 10.1.
To assess the potential for turtles to cross roads throughout Missouri, we first identified suitable three-toed box turtle habitat, defined as forested and wooded habitats (including deciduous forest, evergreen forest, mixed forest, deciduous woody/herbaceous, evergreen woody/herbaceous, and mixed woody/herbaceous) as identified in the 2005 Missouri Land Use Land Classification shapefile available on the Missouri Spatial Data Information Service (http://msdis.missouri.edu/pub/lulc/lulc05). Within the resulting habitat shapefile we generated 100 random points, each of which served as an origin for 2 × 100 CRWs, generated from the movement data of the turtles with the largest and smallest three-year cumulative home ranges. As above, we calculated the proportion of CRWs that crossed roads or extended beyond suitable habitat with the select by location tool in ArcGIS 10.1, based the MO 2014 February MoDOT Roads shapefile ftp://msdis.missouri.edu/pub/Transportation_Networks/MO_2014_February_MoDOT_Roads_gdb.zip) and the lulc05.shp shapefile.
To test the hypothesis that Tyson (rural) turtles had larger home ranges than Forest Park (urban) turtles we used an Analysis of Variance (ANOVA) using the natural log of the home range metric (MCP, 50%UD, 75%UD, and 95%UD) of each turtle as the dependent variable, and sex and site as independent variables.
All statistical analyses were conducted using either Genstat Version 16 (VSN International, Hemel Hempstead, UK).
Results
Turtle home range size
A sample of 21 turtles was included in the study based on their MCP areas reaching asymptote over time. The MCP home range areas of box turtles varied over three orders of magnitude, from 1.0 to 249.9 ha (Table 1). Maximum linear displacement within these ranges was between 150 and 2504 m. An ANOVA strongly supported our hypothesis that home range areas would be larger at Tyson than Forest Park for all home range metrics (MCP: F(1,20) = 10.17, P = 0.005; 95%UD: F(1,20) = 8.15, P = 0.011; 75%UD: F(1,20) = 4.58, P = 0.046; 50%UD: F(1,20) = 6.56, P = 0.020). The largest difference in home range metrics between sites was the 50%UD (21.1 times larger at Tyson than Forest Park) (Table 1). Sex had no effect on turtle home range area for any home range metric considered (P > 0.2). Annual home range sizes at Tyson were consistently larger than Forest Park. Mean annual MCP areas in 2012, 2013, 2014 and 2015 were 13, 14, 4 and 3 times larger at Tyson than Forest Park respectively (Fig. 2).

Mean annual MCP home range estimates (± SE) of three-toed box turtles were consistently larger at Tyson Research Center than Forest Park
Movement simulations
Of 100 correlated random walks using movement data from the most mobile turtle (Tyson: Kevin) generated from each of the 100 random locations scattered across Missouri, some 89.5% involved road crossings. At 40% of starting locations, all 100 simulations crossed roads. Among simulations that crossed roads, the mean number of crossings was 10.6, with a maximum of 52 crossings. Simulations of the least mobile turtle (Forest Park: Kimi) crossed roads in 61% of simulations, with an average of 5.1 crossings per simulation that crossed a road at least once, and a maximum of 27 crossings for a single simulation. In both cases, the frequency of road crossings by road type was consistent; country roads were crossed most frequently (74% and 64% for most and least mobile respectively) followed by Missouri lettered routes (17% and 11%), and Missouri numbered routes (7% and 4%). The frequency of crossings involving other road types were less than 4%. Unfortunately, traffic volume data are not available for these road designations, and therefore estimating the likelihood of encounters between vehicles and turtles is not possible. The probability that simulated CRWs would extend beyond suitable habitat patches was also high. Of 10,000 CRWs generated from Kevin’s data, 98.5% extended beyond the suitable habitat boundary from which the CRWs were generated, while for Kimi’s CRWs 65.5% extended into unsuitable habitat.
Discussion
Three-toed box turtles at Tyson had larger home ranges than turtles in Forest Park. Specifically, mean MCP area was nine times larger at Tyson (68.2Ha compared to 7.4Ha at Forest Park). This pattern was consistent for 95%, 75%, and 50%UD areas, with the largest difference between metrics being the 50%UD. The 50%UD is often considered the “core” of an animal’s home range (Seaman and Powell 1990; White and Garrott 1990), within which key resources are concentrated (Ewert et al. 1994). We do not know details of resource requirements of TTBT nor the behaviors of individual turtles in this study, however the enormous size difference in core areas, and all home range metrics, implies very different energy expenditure of turtles between the urban and rural area. The mean MCP area of Tyson turtles was also larger than any previously reported population level home range estimates: From a total of 23 published studies involving ornate turtles (Terrapene ornata ornata) and TTBT, Habeck et al. (2019) found a mean MCP area of 6.06Ha (figure obtained from calculating the mean of values presented in their results), over 10 times smaller than the mean Tyson turtle MCP areas and slightly smaller than the mean MCP area from Forest Park. Why turtles at Tyson are apparently so mobile is unclear but may be because most previous turtle studies selected animals in forest fragments rather than large contiguous blocks, though for some studies the local habitat geography was difficult to assess (e.g. Bernstein et al. 2007, Iglay et al. 2007, Rittenhouse et al. 2007, Refsnider et al. 2012). Relative to other box turtle species, there are few movement studies of TTBT and this sub-species may be more mobile than eastern and ornate box turtles. That mobility is related to forest fragment size is suggested by our data which demonstrated that TTBT in the small habitat fragments of Forest Park rarely left forested habitats. In an urban park setting the consequence of doing so can be severe for notoriously slow moving and cryptic turtles (Gibbs and Shriver 2002; Converse et al. 2005; Gibbs and Steen 2005; Dodd et al. 2006; Beaudry et al. 2008; Turtle Conservation Coalition 2011; Ferronato et al. 2016). Previous research on our study populations documented the odds of annual survival was 3.5 times lower in Forest Park turtles compared to Tyson turtles with most Forest Park turtle deaths due to unspecified “winter kill.” (Palmer et al. 2019) Of 23 Forest Park turtles monitored over seven years, three were killed by lawnmowers on leaving their forest fragments (Palmer et al. 2019). If ranging patterns and habitat fidelity are heritable traits, positive selection pressure for small home range and high preference for forest habitat is likely to be high in urban settings.
Given the detrimental impacts of anthropogenic factors on turtles (Gibbs and Shriver 2002; Gibbs and Steen 2005; Beaudry et al. 2008; Turtle Conservation Coalition 2011; Refsnider et al. 2012; Baxter-Gilbert et al. 2015) it is intriguing that box turtles continue to exist in Forest Park. Over 13 million people visit the park each year, road traffic is intense during summer months when turtles are active, and the tradition of turtle collection for the pet trade probably all combine to threaten Forest Park turtles. These movement data do not allow us to effectively evaluate the turtle conservation potential of forest patches in an urban park, but they do suggest that the “single large” forest fragment will have higher conservation value than “several small” fragments when planning urban forests (though several large would be better still (Diamond and May 1981)).
While we expected smaller turtle home ranges at the urban versus rural site, we did not predict the trends revealed by the spatial analysis across the state of Missouri. Road density in Missouri is moderate, with a mean density of 1.18 km per square kilometer of land surface and high concentration of roads in urban and sub-urban areas (https://www.fhwa.dot.gov/policyinformation/statistics/2019/) (contiguous US mean by state = 1.13kmkm− 2, max. = 6.25 km− 2, min. = 0.191 kmkm− 2, Rhode Island and Wyoming respectively). Simulations based on field data revealed that when originating from 100 random points distributed in suitable box turtle habitat anywhere in Missouri, correlated random walks generated from the turtle with the smallest MCP from this study would cross a road from 61% of locations. Simulated movement data based on the turtle with the largest home range (254 ha) involved road crossings at 100% of locations (Table 2). Turtles are threatened with morbidity and mortality associated with road crossings throughout the state, and few turtles range outside of the direct influence of roads in Missouri. Similarly, despite the Missouri Ozarks containing some of the largest contiguous forest blocks east of the Rockies, our data and simulations suggest that TTBT movements are restricted across most of their range by habitat fragmentation. It is problematic to extrapolate from two small study sites to the state or species range extent, however our projections are likely to be conservative. We sampled turtle movements in a rural-urban interface at the edge of greater Saint Louis, where forest blocks are relatively small compared to more rural sites, thus our movement data are likely to under-estimate mean state-wide home range characteristics.
In conclusion, TTBT MCP home range size varied from 1 to 249.9 ha, and range size among individuals was positively correlated with habitat fragment size. Turtles in an urban park may live for extended periods (at least four years based on our data) in home ranges of less than 2 ha, thus small urban forest fragments may have some conservation value for box turtles. Simulations suggest that mortality of even highly sedentary box turtles from roads is high throughout Missouri, a state of moderate road density and with large swathes of contiguous forest. The usual suite of conservation recommendations, specifically reducing the negative impacts of roads and fragmentation, must be applied in rural areas as well as densely populated cityscapes if box turtle populations are to avoid further declines.

Overview of potential three-toed box turtle habitat in Missouri, roads and their potential impact; (A) suitable three-toed box turtle habitat, (B) the distribution of state highways in Missouri, (C) locations of 100 origin points for simulated box turtle movements using correlated random walks generated from turtle movement data, (D) close up view of one origin point showing 100 simulated correlated random walks based on the movement data from a highly mobile (red) and a highly sedentary (blue) box turtle
Data Availability
Movement data are freely available on movebank.org under the study “St. Louis Box Turtle Project”.
Change history
13 May 2023
A Correction to this paper has been published: https://doi.org/10.1007/s11252-023-01367-3
References
Agha M, Price SJ, Nowakowski AJ, Augustine B, Todd BD (2017) Mass mortality of eastern box turtles with upper respiratory disease following atypical cold weather. Dis Aquat Organ 124:91–100
Alig RJ, Kline JD, Lichtenstein M (2004) Urbanization on the US landscape: looking ahead in the 21st century. Landsc Urban Plann 69:219–234
Andrews KM, Priya N, Riley SPD (2015) Roads and ecological infrastructure: comcepts and applications for small animals. Johns Hopkins University Press, Baltimore, Maryland, USA
Baxter-Gilbert JH, Riley JL, Lesbarrères D, Litzgus JD (2015) Mitigating Reptile Road Mortality: fence failures compromise ecopassage effectiveness. PLoS ONE 10:e0120537
Beaudry F, deMaynadier PG, Hunter ML Jr (2008) Identifying road mortality threat at multiple spatial scales for semi-aquatic turtles. Biol Conserv 141:2550–2563
Bernstein NP, Richtsmeier RJ, Black RW, Montgomery BR (2007) Home range and philopatry in the ornate box turtle, Terrapene ornata ornata, in Iowa. Am Midl Nat 157:162–174
Beyer HL (2012) Geospatial Modelling Environment (Version 0.7. 2.1). Spatial Ecology, LLC
Blake S, Deem. S L, Strindberg S, Maisels F, Momont L, Bila-Isla I, Douglas-Hamilton I, Karesh WB, Kock MD (2008) Roadless wilderness area determines Forest Elephant Movements in the Congo Basin. PLoS ONE 3:e3546
Budischak SA, Hester JM, Price SJ, Dorcas ME (2006) Natural history of Terrapene carolina (Box Turtles) in an urbanized landscape. Southeast Nat 5:191–204
Converse SJ, Iverson JB, Savidge JA (2005) Demographics of an ornate box turtle population experiencing minimal human-induced disturbances. Ecol Appl 15:2171–2179
Diamond J, May R (1981) Island biogeography and the design of natural reserves. In” Theoretical Ecology: Principles and Applications, “(RM May, ed.), Ch. 10. Blackwell, Oxford and Sinauer, Sunderland, Massachusetts
Dodd K, Ozgul A, Oli MK (2006) The influence of disturbance events on Survival and Dispersal Rates of Florida Box Turtles. Ecol Appl 16:1936–1944
Ewert MA, Jackson DR, Nelson CE (1994) Patterns of temperature-dependent sex determination in turtles. J Exp Zool 270:3–15
Fan SG, Chan-Kang C (2008) Regional road development, rural and urban poverty: evidence from China. Transp Policy 15:305–314
Ferronato BO, Roe JH, Georges A (2016) Urban hazards: spatial ecology and survivorship of a turtle in an expanding suburban environment. Urban Ecosyst 19:415–428
Forman RTT, Alexander LE (1998) Roads and their major ecological effects. Annu Rev Ecol Syst 29:207–231
Gibbs JP (1998) Amphibian movements in response to forest edges, roads, and streambeds in southern New England. J Wildl Manage 62:584–589
Gibbs JP, Shriver WG (2002) Estimating the effects of road mortality on turtle populations. Conserv Biol 16:1647–1652
Gibbs JP, Steen DA (2005) Trends in Sex Ratios of Turtles in the United States: implications of Road Mortality. Conserv Biol 19:552–556
Goddard MA, Dougill AJ, Benton TG (2010) Scaling up from gardens: biodiversity conservation in urban environments. Trends Ecol Evol 25:90–98
Habeck CW, Figueras MP, Deo JE, Burke RL (2019) A surfeit of studies: what have we learned from all the Box Turtle (Terrapene carolina and T. ornata. Home Range Studies? Diversity 11:68
Iglay RB, Bowman JL, Nazdrowicz NH (2007) Eastern Box Turtle (Terrapene carolina carolina) movements in a fragmented landscape. J Herpetology 41:102–106
Johnson MTJ, Munshi-South J (2017) Evolution of life in urban environments. Science 358:eaam8327
Jørgensen PS, Folke C, Carroll SP (2019) Evolution in the Anthropocene: informing governance and policy. Annual Review of Ecology. Evol Syst 50:527–546
Lambert MR, Donihue CM (2020) Urban biodiversity management using evolutionary tools. Nat Ecol Evol 4:903–910
Laurance WF, Cochrane MA, Bergen S, Fearnside PM, Delamonica P, Barber C, D’Angelo S, Fernandez T (2006) The future of the brazilian Amazon. Science 291:438–439
Laurance WF, Clements GR, Sloan S, O’Connell CS, Mueller ND, Goosem M, Venter O, Edwards DP, Phalan B, Balmford A, Van Der Ree R, Arrea IB (2014) A global strategy for road building. Nature 513:229–232
McDonnell MJ, Hahs AK (2015) Adaptation and Adaptedness of Organisms to Urban Environments. Annual Review of Ecology, Evolution, and Systematics 46:261–280
McKinney ML (2002) Urbanization, biodiversity, and conservation. Bioscience 52:883–890
Mittermeier RA, van Dijk PP, Rhodin AG, Nash SD (2015) Turtle hotspots: an analysis of the occurrence of tortoises and freshwater turtles in Biodiversity hotspots, High-Biodiversity wilderness areas, and Turtle Priority Areas. Chelonian Conserv Biology 14:2–10
Oxley DJ, Fenton MB, Carmody GR (1974) Effects of roads on populations of small mammals. J Appl Ecol 11:51–59
Palmer JL, Blake S, Wellehan JFX, Childress AL, Deem SL (2016) Clinical Mycoplasma sp infections in free-living three-toed Box Turtles (Terrapene carolina triunguis) in Missouri, USA. J Wildl Dis 52:378–382
Palmer JL, Brenn-White M, Blake S, Deem SL (2019) Mortality in Three-Toed Box Turtles (Terrapene mexicana triunguis) at two Sites in Missouri. Frontiers in Veterinary Science 6.
Patrick D, Gibbs J (2010) Population structure and movements of freshwater turtles across a road-density gradient. Landscape Ecol 25:791–801
Percoco M (2016) Highways, local economic structure and urban development. J Econ Geogr 16:1035–1054
Pickett STA, Cadenasso ML, Grove JM, Nilon CH, Pouyat RV, Zipperer WC, Costanza R (2001) Urban ecological systems: linking terrestrial ecological, physical, and socioeconomic components of metropolitan areas. Annu Rev Ecol Syst 32:127–157
Rees M, Roe JH, Georges A (2009) Life in the suburbs: Behavior and survival of a freshwater turtle in response to drought and urbanization. Biol Conserv 142:3172–3181
Refsnider JM, Strickland J, Janzen FJ (2012) Home Range and Site Fidelity of Imperiled Ornate Box Turtles (Terrapene ornata) in Northwestern Illinois. Chelonian Conserv Biology 11:78–83
Rhodin AGJ, Iverson JB, Bour R, Fritz U, Georges A, Shaffer HB (2017) and P. P. v. Dijk. Turtles of the World: Annotated Checklist and Atlas of Taxonomy, Synonymy, Distribution, and Conservation Status (8th Ed.)
Rittenhouse CD, Millspaugh JJ, Hubbard MW, Sheriff SL (2007) Movements of translocated and Resident Three-Toed Box Turtles. J Herpetology 41:115–121
Rytwinski T, Fahrig L (2012) Do species life history traits explain population responses to roads? A meta-analysis. Biol Conserv 147:87–98
Seaman DE, Powell RA (1990) Identifying Patterns and Intensity of Home Range Use. Bears: Their Biology and Management 8:243–249
Seto KC, Güneralp B, Hutyra LR (2012) Global forecasts of urban expansion to 2030 and direct impacts on biodiversity and carbon pools. Proceedings of the National Academy of Sciences 109:16083–16088
Soares-Filho B, Alencar A, Nepstad D, Cerqueira G, Vera Diaz MdC, Rivero S, Solórzano L, Voll E (2004) Simulating the response of land-cover changes to road paving and governance along a major Amazon highway: the Santarém–Cuiabá corridor. Glob Change Biol 10:745–764
Stickel LF (1978) Changes in a box turtle population during 3 decades. Copeia:221–225
Torres A, Jaeger JAG, Alonso JC (2016) Assessing large-scale wildlife responses to human infrastructure development. Proceedings of the National Academy of Sciences 113:8472–8477
Turtle Conservation Coalition (2011) IUCN/SSC Tortoise and Freshwater Turtle Specialist Group, Turtle Conservation Fund, Turtle Survival Alliance, Turtle Conservancy. Chelonian Research Foundation, Conservation International, Wildlife Conservation Society, and San Diego Zoo Global, Lunenburg, MA, Turtles in Trouble: The World’s 25 + Most Endangered Tortoises and Freshwater Turtles-2011
van Dijk PP (2016) Terrapene carolina, The IUCN Red List of Threatened Species 2016
White G, Garrott R (1990) Analysis of Wildlife Radio-tracking data. Academic Press, New York. USA
Wilkie D, Shaw E, Rotberg F, Morelli G, Auzel P (2000) Roads, development, and conservation in the Congo basin. Conserv Biol 14:1614–1622
Zimmerman M, Wagner WL (1979) A description of the woody vegetation of oak-hickory forest in the Northern Ozark Highlands. Bull Torrey Bot Club 106:117–122
IUCN (2016) The IUCN Red List of Threatened Species
R Development Core Team (2022) R. A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria
Acknowledgements
We thank the Saint Louis Zoo, Forest Park Forever, and the Tyson Research Center of Washington University in Saint Louis for supporting the Saint Louis Box Turtle Project (SLBTP). Numerous undergraduates from the Tyson Summer Internship program, and high school students from the Tyson Environmental Research Fellowship (TERF) and Shaw Institute for Field Training (SIFT) programs as well as student interns at the Institute for Conservation Medicine assisted with data collection. Two anonymous reviewers are thanked for their comments and their attention to detail which improved the paper considerably.
Funding
Open Access funding enabled and organized by Projekt DEAL. The Saint Louis Box Turtle Project was supported by the National Science Foundation (DEB 1258062) and by the Saint Louis Zoo.
Author information
Authors and Affiliations
Contributions
Stephen Blake and Sharon Deem designed the study. All authors contributed to data collection. Stephen Blake analyzed the data and wrote the first draft of the manuscript. All authors edited and developed the final draft. All authors approved the final manuscript.
Corresponding author
Ethics declarations
The authors declare no competing interests.
Ethics approval.
All turtle handling procedures were approved by the Saint Louis Zoo Institutional Animal Care and Use Committee, and handling of turtles in FP and TRC were covered by a Missouri Department of Conservation permit.
Competing interests.
The authors have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
The original online version of this article was revised: Several typographical errors were found in the initial online published version of this manuscript, which have been corrected in the current version.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Blake, S., Palmer, J., Brenn-White, M. et al. Home ranges of box turtles in a rural woodland and an urban park in Saint Louis, MO; implications for turtle conservation. Urban Ecosyst 26, 1181–1189 (2023). https://doi.org/10.1007/s11252-023-01354-8
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11252-023-01354-8