
Abstract
As urbanisation continues to reduce the available habitat for wildlife, some species, including the black-and-gold howler monkey (Alouatta caraya) in Pilar, southwest Paraguay, are making their homes in anthropogenic environments. Understanding an animal’s home range is an important step to understanding its ecological needs, and an essential requirement for the creation of robust conservation plans. In this study, we determined the home ranges and core areas of five groups of urban dwelling A. caraya using Minimum Convex Polygon (MCP) and Kernel Density Estimation (KDE) Analysis. We used a Spearman’s Correlation to explore the relationship between home range size and group size. All five groups had home ranges of less than 10 ha and used core areas of less than 1 ha. Group size had no significant relationship to home range size. We provide the first estimates of home range for A. caraya in an urban environment in Paraguay. Though the home ranges of the urban A. caraya in Pilar, Paraguay fall at the smaller end of the spectrum of range sizes in Alouatta, they are not abnormal for a species in this genus.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Urbanization, the process by which human populations move from rural to urban areas and the subsequent expansion of urban infrastructures (Berry 2008), has been accelerating in recent years. Today, it is estimated that more than half of the world’s population live in urban centres (Ritchie and Rosen 2018; Corrêa et al. 2018). Urbanization can result in the alteration, fragmentation or destruction of natural habitats (Pauchard et al. 2006; Liu et al. 2016; Šálek et al. 2015). As of 2017, 60% of non-human primates (hereafter primates) are threatened with extinction, making it increasingly important to understand the wide-ranging impacts of anthropogenic expansion on the ecology of wild primate populations (Estrada et al. 2017). Urban expansion can negatively impact the survival of primate populations directly and indirectly by decreasing available habitat size and quality (Estrada 2013; Heil et al. 2007; Fernandez-Juricic 2001; Spellerberg 1998). Primates must either increase dependence on a rapidly disappearing natural habitat or adapt to life in the anthropogenic environment (Mittermeier and Cheney 1987; Janzen 1983). Some primate species appear to have been able to adapt to highly anthropogenically altered areas (Jaman and Huffman 2013; Fehlmann et al. 2017; Kane and Smith 2020) and understanding the ecological needs of these populations is essential if successful long-term management and conservation measures are to implemented (Bicca-Marques et al. 2020).
One important characteristic of a species ecology, that the urban environment has the potential to alter significantly, is the home range. An animal’s home range is the area that an individual uses during its normal activities (Burt 1943). In natural environments, home range size is influenced by many ecological and environmental factors including body size (McNab 1963), food abundance (Hanya et al. 2006; López-Bao et al. 2010), climate and seasonal changes (Kuo and Lee 2012; Osborn 2003; Rautenstrauch and Krausman 1989), mating opportunities (Sugiyama et al. 1965), competition and territoriality (Garneau et al. 2008; Grant et al. 1992) and predation (Hankerson and Dietz 2014). Anthropogenic obstacles present in urban environments can change primate home range, daily path lengths or can even prevent effective dispersal (Duarte et al. 2011).
The urban environment can impact home range sizes in several ways. Urbanisation can lead to a reduction in availability of natural food resources, while anthropogenic food sources may increase in abundance or availability, affecting diet, activity budgets and ranging patterns (Altmann and Muruthi 1988; Bicca-Marques 2017; Sengupta and Radhakrishna 2018; Thatcher et al. 2018; Back et al. 2019). Urban-dwelling rhesus macaques (Macaca mulatta) have significantly smaller home range sizes than their rural counterparts, possibly because urban groups have access to high quality food and do not need to cover large distances to satisfy their metabolic requirements (Jaman and Huffman 2013). Long-tailed macaques (Macaca fascicularis) in Singapore and Barbary macaques (Macaca sylvanus) in Gibraltar that have access to anthropogenic food sources have reduced home range sizes compared with those that do not have access to human provisioned food sources (Klegarth et al. 2017).
Habitat fragmentation caused by the construction of roads, buildings or open gardens can present unique spatial use challenges to wild primates (Stevenson et al. 2015). To move between fragments, arboreal species may have to descend to the ground or incorporate anthropogenic structures such as powerlines into their travel paths (Corrêa et al. 2018; Lokschin, 2007). Descending to the ground to travel or using electric cables incurs novel risks that could impact home range such as electrocutions, traffic accidents or attacks by domestic dogs (Canis familiaris) and cats (Felis catus) (Boinski et al. 1998; Loskchin et al. 2006; Ram et al. 2015; Katsis et al. 2018; Balogh et al. 2011).
The howler monkeys (Alouatta) are a widespread genus found from southern Mexico to northern Argentina and southern Brazil (Estrada 2015). They can occupy a variety of forest types at varying elevations and show a high degree of behavioural and ecological plasticity, tolerant of a certain degree of disturbance (Di Fiore 2007; Estrada 2015; Stevenson et al. 2015). This tolerance of degraded and fragmented habitat has allowed them to persist, at least in the short term, in areas of increased urbanisation (Bicca-Marques 2003; Di Fiore 2007; Estrada 2015). The black-and-gold howler monkey (A. caraya) is one of the most southernly distributed of the genus, being found throughout Paraguay, Brazil, Bolivia, Argentina, and potentially northwestern Uruguay (Villalba et al. 1995; Jardim et al. 2020). A frugivorous-folivore, this species has been observed to be very adaptable to different habitat types with varying levels of disturbance (Stavis et al. 2019). Groups can exist as male-female pairs or three individuals (Para La Tierra, unpublished data; Aguiar et al. 2003) to multi-male, multi-female groups of more than 20 individuals (Bravo and Sallenave 2003). Just like group size, home ranges in this species in different habitats are very variable, from as small as 1.3 ha (Muhle 2008) to as large as 111.9 ha (Agostini 2009) with a mean size of 11.7 ± 23.3 ha (Barbisan-Fortes et al. 2015). Core areas of the home range are typically very small ranging from 0.16 ha (Bicca-Marques 1993, 1994) to 27.03 ha (Barbisan-Fortes et al. 2015; Agostini 2009) and home ranges of different groups can show little to no overlap (Bravo and Sallenave 2003; Ludwig 2006) to overlaps of up to 70% (Kowalewski 2007).
The aim of this study was to determine the home ranges of five groups of black-and-gold howler monkeys living in the urban centre of Pilar, Ńeembucú, Paraguay, using Minimum Convex Polygon and Kernel Density Estimation analysis. 50% Kernel Density Estimation analysis in order to determine the size of core areas of the home range.
Methods
The authors declare no conflicts of interest. This research was approved by the Ministerio del Ambiente y Desarrollo (MADES) and Fundación Para La Tierra. Our research complied with all local laws. We collected data non-invasively and complied with the American Society of Primatologists Code of Best Practices for Field Primatology 2014.
Study site
We collected data from April to September 2018 in the city of Pilar (26° 52’ 04’’ S 58° 17’ 46’’W), Ñeembucú Department in southwest Paraguay (Fig. 1). Pilar has a population of around 33,000 people In this area of Paraguay this time period covers the last few weeks of Autumn, the full winter season (May-July) and most of Spring (August and September) (high, low and mean monthly temperatures Table 1). Our data were collected over mainly during colder and drier months when howler monkey home ranges typically are smaller due to energy minimising strategy and for a limited time of five and a half months.
Study subjects
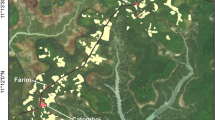
Though Pilar is home to a large population of black and gold howler monkeys (Fig. 1), our study focused on five groups of A. caraya living inside the city (Table 2).
The population of howler monkeys living inside Pilar consists of between 69 and 100 individuals living in 15 groups (Para La Tierra unpublished data). Interviews with local people indicated the population has resided in the city for over 50 years (Alesci 2018; Alesci et al. 2022). Immigrations and emigrations between groups (four dispersing individuals and two complete groups fusions) have been observed during the five year period that scientists from Paraguayan conservation NGO Fundación Para La Tierra have been monitoring the urban population (Para La Tierra unpublished data).
Data collection
Our team collected data from the 16th April 2018–27th September 2018 between 5:45am-10:45am and 13:45pm-17:45pm six days per week throughout the study period. We used focal animal sampling (Altmann 1974) to record the behaviour of each individual for a 45-minute period. At the start of every focal, we recorded the location of the focal individual using a handheld Garmin 64s GPS unit. Data were collated using the program Garmin BaseCamp Version 4.8.7.
Data analysis
The data were analysed using Quantum GIS 3.14 (QGIS 2020). Kernel Density Estimation (KDE) analysis (50% and 95%) were conducted using the Heatmap plugin while Minimum Convex Polygon (MCP) analysis was conducted using the Polygonize and Convex-Hull tools. Once a minimum-convex polygon was created for a group, it was manually edited to avoid overestimating home range size removing roads, open fields, rivers and car parks that had been confirmed as areas of non-use during direct observations. A 50-metre radius from all data point was used as the furthest estimate of home range.
Spearman’s rank correlation was carried out in R Studio version 1.3.1056 to determine whether group size had a relationship with the total home range, 50% core range use and 95% core range use. To avoid the possibility that the infant members of the groups were skewing our data for the entire group, we conducted Spearman’s rank correlation using the results of the full group, adults only and both adults and subadults.
Results
For all groups, the MCP analysis resulted in the largest home range estimates, between 1.62 ha. and 7.86 ha (Table 3; Figs. 2, 3, 4, 5 and 6). The core areas (50% KDE) for all five groups were less than 1 hectare (Table 3; Figs. 2, 3, 4, 5 and 6). The locations of the home ranges in relation to each other is shown in Fig 7.

Location of Pilar in Paraguay and locations of known resident howler monkey groups within the city.

KDE (50% & 90%) and MCP home range maps of Factory Group Background from GoogleEarth 2020

KDE (50% & 90%) and MCP home range maps of Police Group Background from GoogleEarth 2020

KDE (50% & 90%) and MCP home range maps of New Group Background from GoogleEarth 2020

KDE (50% & 90%) and MCP home range maps of Crucecita Group Background from GoogleEarth 2020

KDE (50% & 90%) and MCP home range maps of Trio Group. Background from GoogleEarth 2020

KDE maps of the locations of the five groups in Pilar. Background from GoogleEarth 2020
The Spearmans’ correlation indicated there was no significant relationship between the home range size and group size (MCP: rs= -0.4, p = 0.50463. 50% KDE: rs= 0.2, p = 0.74706. 95% KDE: rs= 0.1, p = 0.87289). We conducted one Spearman correlation using only the adult members of the groups and another using both adult and subadult members. Neither case resulted in a strong correlation between group size and range size (rs= 0. p = 1, rs=-0.22361, p = 0.71769.).
Discussion
Our results showed that both MCP and KDE analysis resulted in small home range size estimates (all less than 10 ha) for all five of the urban howler monkey groups. The 50% KDE demonstrated that all groups had a core area within their home range of less than 1 ha. Home range sizes are extremely variable across the genus Alouatta and though these ranges are at the lower end of the home range size spectrum, they are not unusual for a species in this genus (Horwich & Gebhard 1983; Jardim 1997; Pinto 2002; Gómez-Posadas et al. 2007; Hopkins 2008; Agostini et al. 2010; Bolin 1981; Barbisan-Fortes et al. 2015). For the urban black-and-gold howler monkeys in this study, the MCP results showed home range sizes of between 1.62 and 7.86 ha. This is towards the lower end of the home range size spectrum (3.2–18.4 ha (Ludwig 2006)) for this species using MCP analysis. The 95% KDE analysis gave more extreme results with home range sizes of between 0.91 and 3.07 ha, whereas previous studies have found larger home range sizes in this species using 95% KDE of up to 111.9 ha (Agostini 2009).
The ability of these monkeys to survive in such small areas may be a contributing factor in their persistence in urban centre of Pilar (Alesci et al. 2022; Para La Tierra unpublished data). However, this by no means to say that the situation of A. caraya in this environment is ideal, or even sustainable in the long term as the city continues to develop. One potential reason that the Pilar monkeys are able to thrive in such limited spaces may be due to the presence of supplemental food resources – either by the abundance of mango and avocado trees providing a lot of high quality food or in some cases, direct provisioning by humans (observed frequently with the Police Group) (Fundación Para La Tierra unpublished data; Alesci et al. 2022). If these resources were to become unavailable in the future, potentially due to anthropogenic modification (urbanization is rarely a static process (Berry 2008)), it is possible that these groups could struggle in these environments or be forced to change their behaviour.
Our study found no relationship between group size and home range size. This finding is at odds with previous research where a positive correlation between group size and home range has been observed (González-Hernández et al. 2011), as greater numbers of group members increases food competition, requiring more space to forage and meet individuals energetic needs (Chapman et al. 2009). In contrast to these findings a comprehensive review that not only included group size but population density, fragment size and group biomass as factors in determining home range found that there are many variables at play that may determine an individual groups home range, and that group size alone is too simplistic a metric to use in assessing home range (Barbisan-Fortes et al. 2015). Our study groups in Pilar had access to high quality food and the group with the highest population density, the Police group, received direct human provisioning. Without further examination into other factors that may influence home range size and the limited time frame and season of our study, we are cautious to make any inferences about group size and home range from our current findings.
All our study groups home ranges included a matrix of roads, buildings and powerlines. Several of the groups included these into their daily paths and descended to the ground to move between fragmented sections of their range. This behavioural flexibility may have contributed to their persistence in the urban environment but these actions also come at a cost (Corrêa et al. 2018; Teixeira et al. 2013), that previous studies of the same groups in Pilar have demonstrated that the monkeys are not aware of (Wellian and Smith 2021).
A potential issue for the long-term survival of Pilar’s urban howler monkey population is the disconnected nature of their ranges. Smaller and more disconnected habitat fragments have been demonstrated to make effective dispersal harder for howler monkeys, but this has not been well explored in an urban environment (Arroyo-Rodríguez and Dias 2010). In a natural living population of brown howler monkeys (Alouatta guariba clamitans) in a fragmented, isolated habitat, group size and population density were large as dispersal was difficult and immigration rarely successful (Fortes and Bicca-Marques 2008). These difficulties led to an increase in inbreeding, a lack of genetic diversity and gave rise to the expression of rare alleles which gave some members of the group an abnormally lightened pelage (Fortes and Bicca-Marques 2008).
One solution that could allow for an expansion of home ranges or dispersal safely, is the construction of rope bridges. Rope bridges across roads may help to limit injuries and deaths due to collisions with traffic and provide an alternative to powerlines (Hernández-Pérez 2015; Teixeira et al. 2013). The insulation of power cables could also mitigate the danger associated with using them as crossings, although encouraging use of rope bridges in these areas simultaneously should be considered (Lokschin et al. 2007; Teixeira et al. 2013). The planting of trees to produce a “live fence” or natural canopy bridge has shown to be beneficial and increased home range size for Mexican howler monkeys (Alouatta palliata mexicana) living in edge and fragmented habitats (Asensio et al. 2009) and primates have been showed to prefer natural canopy bridges to artificial crossings (Lindshield 2016). A simple method of preventing monkeys from utilising powerlines is to clip the trees and hedges that provide access to them (Lindshield 2016), however doing so without providing another means of crossing may force monkeys to use terrestrial means to connect to part of their range, increasing the risk of traffic collisions or dog attacks.
As the human population of Pilar grows it is likely that the city will continue to expand into the surrounding Ñeembucú Wetland Complex, reducing available natural habitat for A. caraya. Although A. caraya show behavioural plasticity when faced with habitat fragmentation and anthropogenic disruption, it is still unclear to what degree can they adjust before their survival and genetic diversity is negatively influenced. By understanding the behaviour and ecology of those monkeys that have currently adapted to life in an anthropogenic environment, we can begin to develop long-term conservation strategies to help safeguard the survival of these unique primates into the future.
Data Availability
Data for this study is available at https://github.com/rebeccasmithPLT2020/Pilar-Howler-Monkeys.git.
References
Agostini I (2009) Ecology and behavior of two howler monkey species (Alouatta guariba clamitans and Alouatta caraya) living in sympatry in northeastern Argentina. PhD dissertation, Università degli Studi di Roma “La Sapienza”, Rome, Italy
Agostini I, Holzmann I, Di Bitetti MS (2010) Ranging patterns of two syntopic howler monkey species (Alouatta guariba and A. caraya) in northeastern Argentina. Inter J Primatol 31(3):363–381. https://doi.org/10.1007/s10764-010-9390-x
Aguiar LM, Reis NR, Ludwig G, Rocha VJ (2003) Dieta, área de vida e estimativas populacionais de Alouatta guariba em um remanescente florestal do norte do estado do Paraná. Neotropical Primates 11:78–85
Alesci M, Smith RL, Ayala Santacruz JD, Camperio Ciani A (2022) Attitudes towards urban black and gold howler monkeys (Alouatta caraya) in the city of Pilar, Ñeembucú, southwest Paraguay. Primates 63:161–171
Altmann J (1974) Observational study of behavior: sampling methods. Behav 49(3/4):227–267
Altmann J, Muruthi P (1988) Differences in daily life between semiprovisioned and wild-feeding baboons. Am J Primatol 15(3):213–221. https://doi.org/10.1002/ajp.1350150304
Arroyo-Rodríguez V, Dias PAD (2010) Effects of habitat fragmentation and disturbance on howler monkeys: a review. Am J Primatol 72(1):1–16. https://doi.org/10.1002/ajp.20753
Asensio N, Arroyo-Rodríguez V, Dunn JC, Cristóbal-Azkarate J (2009) Conservation value of landscape supplementation for howler monkeys living in forest patches. Biotropica 41(6):768–773. https://doi.org/10.1111/j.1744-7429.2009.00533.x
Back JP, Bicca-Marques JC (2019) Supplemented howler monkeys eat less wild fruits, but do not change their activity budgets. Am J Primatol 81(9):e23051. https://doi.org/10.1002/ajp.23051
Balogh AL, Ryder TB, Marra PP (2011) Population demography of gray catbirds in the suburban matrix: sources, sinks and domestic cats. J Orinthol 152:717–726. https://doi.org/10.1007/s10336-011-0648-7
Barbisan-Fortes VB, Bicca-Marques JC, Urbani B, Fernández VA, da Silva Pereira T (2015) Ranging behavior and spatial cognition of howler monkeys. In: Kowalewski M, Garber P, Cortés-Oritz L, Urbani B, Youlatos D (eds) Howler monkeys. Developments in primatology: Progress and prospects. Springer, New York. https://doi.org/10.1007/978-1-4939-1960-4_9
Berry BJ (2008) Urbanization. In: Marzluff JM et al (eds) Urban Ecology. Springer, Boston, MA. https://doi.org/10.1007/978-0-387-73412-5_3
Bicca-Marques JC (1993) Padrão de atividades diárias do bugio-preto Alouatta caraya (Primates, Cebidae): Uma análise temporal e bioenergética. In: Yamamoto ME, Sousa MBC (eds) A Primatologia no Brasil, vol 4. Editora Universitária UFRN, Natal, pp 35–49
Bicca-Marques JC (1994) Padrão de utilização de uma ilha de mata por Alouatta caraya (Primates: Cebidae). Rev Brasil Biol 54:161–171
Bicca-Marques JC (2017) Urbanization (and primate conservation). In: Fuentes A, Bezanson M, Campbell CJ, Di Fiore AF, Elton S, Estrada A, Yamagiwa J (eds) The international encyclopedia of primatology. Wiley, New York, pp 1–5. https://doi.org/10.1002/9781119179313.wbprim0153
Bicca-Marques JC (2003) How do howler monkeys cope with habitat fragmentation? In: Marsh LK (ed) Primates in fragments: Ecology and conservation. Springer US, Boston, MA, pp 283–303. https://doi.org/10.1007/978-1-4757-3770-7_18
Bicca-Marques JC, Chaves ÓM, Hass GP (2020) Howler monkey tolerance to habitat shrinking: Lifetime warranty or death sentence? Am J Primatol 82:e23089. https://doi.org/10.1002/ajp.23089
Boinski S, Jack K, Lamarsh C, Coltrane JA (1998) Squirrel monkeys in Costa Rica: Drifting to extinction. Oryx 32:45–58. https://doi.org/10.1046/j.1365-3008.1998.00017.x
Bolin I (1981) Male parental behavior in black howler monkeys (Alouatta palliata pigra) in Belize and Guatemala. Primates 22:349–360. https://doi.org/10.1007/BF02381575
Bravo SP, Sallenave A (2003) Foraging behavior and activity patterns of Alouatta caraya in the Northeastern Argentinean flooded forest. Int J Primatol 24:825–846
Burt WH (1943) Territoriality and home range concepts as applied to mammals. J Mammal 24(3):346–352. https://doi.org/10.2307/1374834
Chapman CA, Rothman JM, Hodder SA (2009) Can parasite infections be a selective force influencing primate group size? A test with red colobus. In: Chapman CA, Huffman MA (eds) Primate parasite ecology. The dynamics and study of host-parasite relationships. Cambridge University, Cambridge, pp 423–440
Corrêa FM, Chaves ÓM, Printes RC, Romanowski HP (2018) Surviving in the urban–rural interface: Feeding and ranging behavior of brown howlers (Alouatta guariba clamitans) in an urban fragment in southern Brazil. Am J Primatol 80:e22865. https://doi.org/10.1002/ajp.22865
Di Fiore A, Campbell CJ (2007) The atelines: Variation in ecology, behavior, and social organization. In: Campbell CJ, Fuentes A, MacKinnon KC, Panger M, Beader SK (eds) Primates in perspective. Oxford University Press, New York, pp 155–185
Duarte MH, Vecci MA, Hirsch A, Young RJ (2011) Noisy human neighbours affect where urban monkeys live. Biol lett 7:840–842. https://doi.org/10.1098/rsbl.2011.0529
Estrada A (2015) Conservation of alouatta: Social and economic drivers of habitat loss, information vacuum, and mitigating population declines. In: Kowalewski MM, Garber PA, Cortés-Ortiz L, Urbani B, Youlatos D (eds) Howler monkeys: Behavior, ecology, and conservation. Springer New York, New York, pp 383–409
Estrada A, Garber PA, Rylands AB, Roos C, Fernandez-Duque E, Di Fiore A et al (2017) Impending extinction crisis of the world’s primates: Why primates matter. Sci Adv 3(1):e1600946. https://doi.org/10.1126/sciadv.1600946
Fehlmann G, O’Riain MJ, Kerr-Smith C et al (2017) Extreme behavioural shifts by baboons exploiting risky, resource-rich, human-modified environments. Sci Rep 7:15057. https://doi.org/10.1038/s41598-017-14871-2
Fortes VB (2008) Ecologia e comportamento do bugio-ruivo (Alouatta guariba clamitans Cabrera, 1940) em fragmentos florestais na Depressão Central do Rio Grande do Sul, Brasil. PhD dissertation, Pontifícia Universidade Católica do Rio Grande do Sul, Porto Alegre, Brazil
Fortes VB, Bicca-Marques JC (2008) Abnormal pelage color in an isolated population of Alouatta guariba clamitans Cabrera, 1940 in South Brazil. Intern J Primatol 29(3):717. https://doi.org/10.1007/s10764-008-9264-7
Garneau DE, Boudreau T, Keech M, Post E (2008) Habitat use by black bears in relation to conspecifics and competitors. Mammalian Biol 73(1):48–57. https://doi.org/10.1016/j.mambio.2006.11.004https://doi.org/10.1016/j.biocon.2006.10.048
Gómez-Posadas C, Martínez J, Giraldo P, Kattan GH (2007) Density, habitat use, and ranging patterns of red howler monkeys in a Colombian Andean forest. Neotrop Primates 14:2–10. https://doi.org/10.1896/044.014.0102
González-Hernández M, Dias PAD, Romero-Salas D, Canales-Espinosa D (2011) Does home range use explain the relationship between group size and parasitism? A test with two sympatric species of howler monkeys. Primates 52(3):211–216. https://doi.org/10.1007/s10329-011-0252-8
Grant JWA, Chapman CA, Richardson KS (1992) Defended versus undefended home range size of carnivores, ungulates and primates. Behav Ecol Sociobiol 31(3):149–161. https://doi.org/10.1007/BF00168642
Hankerson S, Dietz J (2014) Predation rate and future reproductive potential explain home range size in golden lion tamarins. Anim Behav 96:87–95. https://doi.org/10.1016/j.anbehav.2014.07.026
Hanya G, Kiyono M, Yamada A, Suzuki K, Furukawa M, Yoshida Y, Chijiiwa A (2006) Not only annual food abundance but also fallback food quality determines the Japanese macaque density: Evidence from seasonal variations in home range size. Primates 47(3):275–278. https://doi.org/10.1007/s10329-005-0176-2
Hernández-Pérez E (2015) Rope bridges: A strategy for enhancing habitat connectivity of the black howler monkey (Alouatta pigra). Neotrop Primates 22(2):94
Hopkins ME (2008) Spatial foraging patterns and ranging behavior of mantled howler monkeys (Alouatta palliata), Barro Colorado Island, Panama. PhD dissertation, University of California, Berkeley, USA
Horwich RH, Gebhard K (1983) Roaring rhythms in black howler monkeys (Alouatta pigra) in Central America. Primates 24:290–296. https://doi.org/10.1007/BF02381093
Jaman MF, Huffman MA (2013) The effect of urban and rural habitats and resource type on activity budgets of commensal rhesus macaques (Macaca mulatta) in Bangladesh. Primates 54(1):49–59. https://doi.org/10.1007/s10329-012-0330-6
Jardim MMA (1997) Estratégias de forrageamento e uso do espaço por Alouatta belzebul (Primates, Cebidae) na Estação Científica Ferreira Penna, Melgaço, Pará. MSc dissertation, Universidade Federal do Rio Grande do Sul, Porto Alegre, Brazil
Jardim MMA, Queirolo D, Peters FB, Mazim FD, Favarini MO, Tirelli FP, Trindade RA, Bonatto SL, Bicca-Marques JC, Mourthe I (2020) Southern extension of the geographic range of black-and-gold howler monkeys (Alouatta caraya). Mammalia 84(1):102–106. https://doi.org/10.1515/mammalia-2018-0127
Kane J, Smith RL (2020) Bertiella sp. (Meyner, 1895) infection of Alouatta caraya (Humboldt, 1812) in urban and natural environments in Ñeembucú, southwest Paraguay. Am J Primatol 82(9):e23166. https://doi.org/10.1002/ajp.23166
Katsis L, Cunneyworth PMK, Turner KME, Presotto A (2018) Spatial patterns of primate electrocutions in Diani, Kenya. Int J Primatol 39:492–510. https://doi.org/10.1007/s10764-018-0046-6
Kowalewski MM (2007) Patterns of affiliation and co-operation in howler monkeys: An alternative model to explain social organization in non-human primates. PhD dissertation, University of Illinois, Urbana
Klegarth AR, Hollocher H, Jones-Engel L, Shaw E, Lee BPH, Feeney T et al (2017) Urban primate ranging patterns: GPS‐collar deployments for Macaca fascicularis and M. sylvanus. Am J Primatol 79(5):e22633. https://doi.org/10.1002/ajp.22633
Kuo CC, Lee LL (2012) Home range and activity of the Indian giant flying squirrel (Petaurista philippensis) in Taiwan: Influence of diet, temperature, and rainfall. Acta Theriol 57(3):269–276. https://doi.org/10.1007/s13364-012-0076-y
Lindshield SM (2016) Protecting nonhuman primates in peri-urban environments: A case study of neotropical monkeys, corridor ecology, and coastal economy in the Caribe Sur of Costa Rica. In: Waller MT (ed) Ethnoprimatology: Primate conservation in the 21st century. Springer International Publishing, pp 351–369. https://doi.org/10.1007/978-3-319-30469-4_19
Liu Z, He C, Wu J (2016) The relationship between habitat loss and fragmentation during urbanization: An empirical evaluation from 16 world cities. PLoS ONE 11:e0154613. https://doi.org/10.1371/journal.pone.0154613
Lokschin LX, Rodrigo CP, Hallal Cabral JN, Buss G (2007) Power lines and howler monkey conservation in Porto Alegre, Rio Grande do Sul, Brazil. Neotrop Primates 14(2):76–80. https://doi.org/10.1896/044.014.0206
López-Bao JV, Palomares F, Rodríguez A, Delibes M (2010) Effects of food supplementation on home‐range size, reproductive success, productivity and recruitment in a small population of Iberian lynx. Anim Conserv 13(1):35–42. https://doi.org/10.1111/j.1469-1795.2009.00300.x
Ludwig G (2006) Área de vida e uso do espaço por Alouatta caraya (Humboldt, 1812) em ilha e continente do alto rio Paraná. MSc dissertation, Universidade Federal do Paraná, Curitiba, Brazil
McNab BK (1963) Bioenergetics and the determination of home range size. Am Nat 97(894):133–140. https://doi.org/10.1086/282264
Muhle CB (2008) Estratégias adaptativas de um grupo de bugios pretos, Alouatta caraya (Humboldt, 1812), em um bosque dominado por eucaliptos. MSc dissertation, Pontifícia Universidade Católica do Rio Grande do Sul, Porto Alegre, Brazil
Osborn F (2003) Seasonal influence of rainfall and crops on home-range expansion by bull elephants. IUCN 53. https://library.wur.nl/WebQuery/wurpubs/fulltext/43262#page=56 Accessed October 23rd, 2020
Para La Tierra. Colección Zoologica de Para La Tierra.Pilar, Paraguay
Pauchard A, Aguayo M, Peña E, Urrutia R (2006) Multiple effects of urbanization on the biodiversity of developing countries: The case of a fast-growing metropolitan area (Concepción, Chile). Biol Conserv 127:272–281. https://doi.org/10.1016/j.biocon.2005.05.015
Pinto LP (2002) Dieta, padrão de atividades e área de vida de Alouatta belzebul discolor (Primates, Atelidae) em Paranaíta, Norte de Mato Grosso. MSc dissertation, Universidade Estadual de Campinas, Campinas, Brazil
Ram C, Sharma G, Rajpurohit L (2015) Mortality and threats to hanuman langurs (Semnopithecus entellus entellus) in and around Jodhpur (Rajasthan). Indian For 141:1042–1045
Rautenstrauch KR, Krausman PR (1989) Influence of water availability and rainfall on movements of desert mule deer. J Mammal 70(1):197–201. https://doi.org/10.2307/1381689
Riley CM, Koenig BL, Gumert MD (2015) Observation of a fatal dog attack on a juvenile long-tailed macaque in a human-modified environment in Singapore. Nat Singap 8:57–63
Ritchie H, Roser M (2018) Urbanization. Our world in data. https://www.ourworldindata.org/urbanization Accessed 20th May 2021
Šálek M, Drahníková L, Tkadlec E (2015) Changes in home range sizes and population densities of carnivore species along the natural to urban habitat gradient. Mammal Rev 45:1–14. https://doi.org/10.1111/mam.12027
Scheun J, Greeff D, Nowack J (2019) Urbanisation as an important driver of nocturnal primate sociality. Primates 60:375–381. https://doi.org/10.1007/s10329-019-00725-0
Schlichte HJ (1978) A preliminary report on the habitat utilization of a group of howler monkeys (Alouatta villosa pigra) in the national park of Tikal, Guatemala. In: Montgomery GG (ed) The ecology of arboreal folivores. Smithsonian Institution Press, Washington, D.C, pp 551–559
Sengupta A, Radhakrishna S (2018) The hand that feeds the monkey: Mutual influence of humans and rhesus macaques (Macaca mulatta) in the context of provisioning. Int J Primatol 39:817–830. https://doi.org/10.1007/s10764-018-0014-1
Sleeman JM, Meader LL, Mudakikwa AB, Foster JW, Patton S (2000) Gastrointestinal parasites of mountain gorillas (Gorilla gorilla beringei) in the parc national des Volcans, Rwanda. J Zoo Wildlife Medic 31:322–328. https://doi.org/10.1638/1042-7260(2000)031[0322:GPOMGG]2.0.CO;2
Stavis VK, Marques MR, Bordignon MO, Pereira W, da Cunha ER, Fina BG (2019) Area of use and space used by Alouatta caraya in the center-west of Brazil. Res J Life Sci Bioinform Pharm Chem Sci 5(1):787–805. https://doi.org/10.26479/2019.0501.66
Stevenson PR, Beltrán ML, Quiñones MJ, Ahumada JA(2015) Differences in home range, activity patterns and diet of red howler monkeys in a continuous forest and a forest fragment in Colombia. Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales 39(153): 503–513. https://doi.org/10.1002/ajp.1057
Sugiyama Y, Yoshiba K, Pathasarathy M (1965) Home range, mating season, male group and inter-troop relations in hanuman langurs (Presbytis entellus). Primates 6(1):73–106. https://doi.org/10.1007/BF01794459
Thatcher H, Downs C, Koyama NF (2018) Anthropogenic influences on the time budgets of urban vervet monkeys. Landsc Urban Plan 181:38–44. https://doi.org/10.1016/j.landurbplan.2018.09.014
Teixeira B, Hirsch A, Goulart VDLR, Passos L, Teixeira CP, James P, Young R (2016) Good neighbours: Distribution of black-tufted marmoset (Callithrix penicillata) in an urban environment. Wildl Res 42:579–589. https://doi.org/10.1071/WR14148
Teixeira FZ, Printes RC, Fagundes JCG, Alonso AC, Kindel A (2013) Canopy bridges as road overpasses for wildlife in urban fragmented landscapes. Biota Neotrop 13:117–123. https://doi.org/10.1590/s1676-06032013000100013
Villalba IJS, Prigioni CM, Sappa AC (1995) Sobre la posible presencia de Alouatta caraya em Uruguay. Neotrop Primates 3:173–174
Waters S, El Harrad A, Chetuan M, Bell S, Setchell JM (2017) Dogs disrupting wildlife: Domestic dogs harass and kill barbary macaques in Bouhachem forest, North Morocco. Afr Primates 12:55–58
Williams-Guilén(2003) The behavioral ecology of mantled howling monkeys Alouatta palliata living in a Nicaraguan shade coffee plantation. PhD dissertation, New York University, New York, USA
Wellian J, Smith RL (2021) Risk awareness of black-and-gold howler monkeys living in an urban environment in south-west Paraguay. J Urban Ecol 7:juab010. https://doi.org/10.1093/jue/juab010
Acknowledgements
X Duffy would like to thank Queens University School of Biological Sciences. All authors are grateful to the staff, interns and volunteers of Fundación Para La Tierra, the creative community behind the QGIS program. We express our immense gratitude to the Pilar community for their incredible hospitality and friendliness that made this research possible. RL. Smith is grateful to the PRONII program of Conacyt. Thank you to Susan Smith for proof reading this manuscript and to two anonymous reviewers for their helpful comments. Fundación Para La Tierra is grateful to the estate of Don Julio Contreras for their endless support of PLT’s activities.
Funding
The authors did not receive funding for this submitted work.
Author information
Authors and Affiliations
Contributions
Xander Duffy – Data analysis, writing; Jake Wellian – Project design, data collection, writing; Rebecca L. Smith – Project design, data collection, data analysis, writing.
Corresponding author
Ethics declarations
Conflicts of interest/Competing interests
The authors confirm that there are no conflicts of interest.
Ethics approval
This research was approved by the Ministerio del Ambiente y Desarrollo (MADEs) and Fundación Para La Tierra. The research complied with all local laws. All of our data was collected in a non-invasive manner and complied with the American Society of Primatologists Code of Best Practices for Field Primatology 2014.
Consent for publication
All authors consent to the submission of this work.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Duffy, P.X., Wellian, J. & Smith, R.L. Use of Space by black-and-gold howler monkeys (Alouatta caraya) in an urban environment in Paraguay. Urban Ecosyst 25, 1777–1786 (2022). https://doi.org/10.1007/s11252-022-01262-3
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11252-022-01262-3