
Abstract
Land-use and land-cover change strongly affect biodiversity patterns and are assumed to be growing threats in the future. Particularly increasing urbanisation may affect species turnover and functional composition of biological communities. This study aimed to assess the characteristics of land-cover change in a medium-sized urban municipality from 2011 to 2018, and the effects of urbanisation on avian species- and functional diversity. The study was performed in Trondheim (Norway), using local land-cover maps and GBIF bird species occurrence records. GLMMs were used to model species turnover as a function of urbanisation, and the probability of species appearance and disappearance based on urbanisation and species traits. The extent of bird species turnover within a municipality-wide 500 × 500m2 grid was not predicted by a changes in developed area, but the probability of disappearance and appearance of bird species varied with urbanisation and bird functional traits. Species associated with urban- or open areas showed a decreasing probability of disappearing and an increasing probability of appearing with increasing amount of developed area within grid cells. Similarly, granivorous species showed a decreasing probability of disappearing. Species feeding above ground-level showed positive responses to changes in land-cover. The probability of both appearance and disappearance, thus species turnover, increased with increasing longevity. Most functional groups respond negatively to increasing urbanisation, indicating a potential impoverishment of local avifauna with future land-cover modifications. Considering planned future land-cover changes within the municipality, the local avian communities are in danger of homogenisation. The recommendations for local management are to minimise conversion of vulnerable habitats, such as wetlands and woodlands, in particular if these are converted to developed area.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Changes in land-cover and land-use are some of the greatest threats to biodiversity worldwide (Díaz et al. 2019; Newbold et al. 2015; Sala et al. 2000). Increases in anthropogenic pressures, for example through urbanisation, intensification and expansion of agricultural areas, are the main culprits regarding range contractions, population declines and extinctions (IPBES 2018, 2019). Considering the growing human population and the subsequent increased urbanisation globally (United Nations, Department of Economic and Social Affairs 2019), understanding species’ responses to anthropogenic pressures are paramount for guiding conservation measures (Newbold et al. 2018).
It has been shown that land-cover and land-use determine species richness differently among different taxa, groups of conservation concern, and functional groups (Petersen et al. 2020, 2021a). In addition to differences in species richness, land-cover affects community composition; changes in land-cover are thus likely to cause species turnover. Urbanisation, a type of land-cover change of particular concern globally, greatly alters the physical environment (Kaye et al. 2006), and potentially creates new niches, allowing for a new suite of species to colonise areas. With changes in land-cover and physical structures, the structure of the local vegetation will be greatly affected, which will cascade through the food web, affecting all trophic levels and trophic interactions (Chace and Walsh 2006). Likewise, with increasing amounts of built infrastructure comes a change in the vertical structure of the landscape, affecting which feeding guilds might be more likely to succeed. During urbanisation, specialists are often replaced by generalist species (Hagen et al. 2017; MacLean et al. 2018), as the latter are often better equipped to thrive in a changing environment.
Urban areas are frequently identified as the point of entry of alien species (Gaston 2005; Padayachee et al. 2017). Species richness (α diversity) can therefore potentially increase following anthropogenic land-cover changes. However, urbanisation can also cause species homogenisation across large spatial scales (decreasing β diversity): as the urban environment across cities globally is relatively alike, cities across the world become more similar biologically (Blair 1996; Chace and Walsh 2006; McKinney 2006). To assess whether such a homogenisation is taking place, it can therefore be relevant to assess changes in community composition rather than changes in species richness as such, from a conservation point-of-view.
Several studies of biodiversity and land-cover/land-use have assessed the relation between species richness or species composition and current landscape features (e.g. Rittenhouse et al. 2012; Newbold et al. 2018; Petersen et al. (2020)). This approach potentially neglects the effects of past disturbances and biotic lag in the local community; species responses to changes in environmental variables can be time-lagged due to factors including species growth rates and/or resilience (Ernoult et al. 2006; Metzger et al. 2009). Several studies have used space-for-time substitution to infer the effects of land-cover changes and/or urbanisation (Bregman et al. 2016; La Sorte et al. 2018). Despite being a convenient approach in the paucity of time-series data, such substitution might misestimate the effects of habitat changes (Bonthoux et al. 2013; Johnson and Miyanishi 2008), and neglect the dynamics and history of the local community. Thus, using data from different points in time rather than inferring temporal relations extrapolated from spatial patterns is preferable. In the absence of structured time-series data, using species occurrence records from collated datasets from open databases, such as the Global Biodiversity Information Facility (GBIF) (GBIF.org 2019), is an alternative. This however comes with great trade-offs. Species occurrence records in GBIF are compiled from vastly different data contributors (Petersen et al. 2021b; Speed et al. 2018). This causes great variation in sampling effort in both time and space (Newbold 2010; Powney and Isaac 2015; Tiago et al. 2017), as well as issues with inaccurate identification. Nevertheless, such records are increasingly used in research, and continuous work on how to best account for the inherent caveats is underway (Amano et al. 2016; Gaiji et al. 2013). The vast majority of the species occurrence data available through GBIF are records of birds (Amano et al. 2016); this abundance of bird records is likely driven by societal interest and the increasing number of observational records from amateur ornithologists (Troudet et al. 2017). Birds serve important ecological functions (Sekercioglu 2006) and can be used as indicators/surrogates of overall biodiversity in use for conservation planning (Rodrigues and Brooks 2007). Considering their diverse roles in food webs and their large variety in use of microhabitats, assessing the effects of land-cover change on changes in bird biodiversity is a reasonable approach to elucidate changes in ecosystem functioning (Hausner et al. 2003).
Rather than assessing responses of single species, an ecologically meaningful approach is to assess the responses of different functional groups (Hausner et al. 2003; Ikin et al. 2012). Several species are registered infrequently on their own accord, but can potentially fulfil the same ecological function as other species; thus, investigating functional groups rather than species identities provides a mechanistic link between ecosystem characteristics and species communities (Ikin et al. 2012; Palacio et al. 2018). Multiple traits have been shown to affect species responses to land-cover characteristics and urbanisation, including diet, forage strata, body size and longevity (Bregman et al. 2016; Conole and Kirkpatrick 2011; Evans et al. 2018; Pinho et al. 2016). Overall, species responding positively to urban areas predominantly appear to be granivorous and/or omnivorous, forage in the understorey, mid-storey, canopy and in the air (in contrast to lower prevalence of species feeding near- or below the water surface) (Chace and Walsh 2006, Evans et al. 2018, La Sorte et al. 2018). Omnivorous species are versatile in their dietary choices, and thus capable of taking advange of the food sources provided by humans (Chace and Walsh 2006). In a similar fashion, granivores are more likely to thrive as they have been able to take advantage of anthropogenic wastes, such as dropped grain and seeds provided through feeding stations (Chace and Walsh 2006). The low prevalence of species feeding near- or below the water surface likely stem from a lack of suitable water bodies within the urban boundaries. Generalist species are better equipped for a changing urban habitat than are specialists (MacLean et al. 2018; Palacio et al. 2018). Some studies have found urban birds mostly having a medium body size (in contrast to species of more natural habitats, which tend to be either relatively small or large), which fits with a lower prevalence of specialists (Palacio et al. 2018). In contrast, another meta-study found no general patterns in body size (Sepp et al. 2018). Sepp et al. (2018) found urban birds to generally have a slower pace-of-life (and thus higher longevity). This might be explained by cities potentially being stochastic environments, with long-lived species being capable of having multiple reproduction attempts, thus not being as vulnerable to poor breeding conditions in single seasons (Kinnunen et al. 2022).
In this study, we aim to examine the effects of land-cover change in a northern boreal setting on local bird biodiversity by asking: (I) What characterises the land-cover changes which have occurred in a midsized urban municipality (Trondheim, Norway) within the studied time frame; (II) whether the degree of bird species composition change over time (β-diversity) is associated with the changes in built-up area (i.e. urbanisation); and (III) how bird species belonging to different functional groups respond to urbanisation. We expect β-diversity (species replacement) to increase with increasing amounts of urbanisation, and we expect contrasting responses from various functional groups; bird species with flexible habitat requirements or associated with urban areas to respond positively to urbanisation, whereas woodland specialists are expected to respond negatively. Likewise, species predominantly feeding on seeds should benefit from urbanisation, whereas insectivores are disadvantaged. Additionally, to put the results into local management context, we aim to assess the potential land-cover changes described in the municipal master plan for Trondheim municipality, and evaluate the potential consequences for local avian communities.
Materials and methods
Study area
The study was carried out within the administrative borders of Trondheim Municipality (Norway), located around 63.42°N, 10.38°E. Trondheim is a southern-boreal (Moen 1999), coastal municipality with an area of ca. 342 km2 (not counting marine areas) and a population of approximately 195,000 people (Statistics Norway 2020). Trondheim municipality was merged with Klæbu municipality pr. 01.01.2020; this paper deals with Trondheim as prior to this merger. The annual mean temperature and precipitation are approximately 4 °C and 1 123 mm (Climate-Data.org 2022). It is an administrative centre, dominated by education and service businesses and with limited industrial activities (Statistics Norway 2020). The municipality is well-sampled with regards to species occurrence records, in part due to the presence and activity of a university and a University Museum. Birds are particularly well-registered (Petersen et al. 2020, 2021a, b).
Land-cover data and geographical analyses
Data on land-cover within the municipality was based on the Norwegian AR5 maps from NIBIO (Ahlstrøm et al. 2014; Norwegian Institute of Bioeconomy Research 2018) (Table S1). The AR5 maps are periodically updated and provide the most complete data on national land-cover (Kartverket 2019). Shapefiles of the land-cover maps last updated in 2011 and 2018 respectively were provided by the Trondheim Municipality in April 2018.
The digital land-cover maps were cropped by the municipality border of Trondheim and overlaid with a 500 × 500 m2 grid. The entire municipality covered 2 246 grid cells, of which 278 were intersected by the municipality border and were thus not complete. The area of each land-cover category within the grid cells from both map layers were calculated, using the R-packages ‘sf’ (Pebesma 2018), ‘sp’ (Bivand et al. 2013; Pebesma and Bivand 2005) and ‘raster’ (Hijmans 2020). Only grid cells with complete land-cover within the municipality border were included. This includes marine grid cells. Grid cells partly in other municipalities were excluded. The final number of grid cells retained for analyses were 1 968 (87.6% of the initial grid cells). Differences in area of each land-cover category within each grid cell were calculated by subtracting the values from 2011 from the values from 2018 (Supplementary material, Fig. S1).
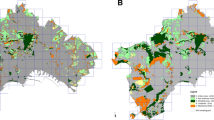
To avoid overfitting of the models, only one category of land-cover change was used in the following analyses. To assess the effects of urbanisation, the change in the land-cover categories “Developed area” and “Communication/traffic” (i.e. roads) (hereafter jointly called “Developed area”) within each of the grid cells was examined (Fig. 1a). See Table S1 for detailed descriptions of land-cover categories.

Change in Developed area and species composition dissimilarity in Trondheim. a Change in Developed area (m2) within 500 × 500 m2 grid cells between 2011 and 2018. b Logit-transformed turnover-component of the Jaccard β-diversity within each 500 × 500 m2 grid cell used in the analyses. Only grid cells included in the models are shown in (b)
All spatial data and maps were handled and drawn using the UTM (zone 32) projection, datum WGS84 (EPSG code 32,632).
Species occurrence data
All occurrence records without known geospatial issues from a bounding box around Trondheim Municipality (as the exact municipality border was too detailed to include in the process) were downloaded from GBIF on 26.04.2020. "Geospatial issues" refers to occurrence records which have been flagged as suspicious or missing content by GBIF (Buitrago 2020). Examples of these issues are mismatches between the written country and the given coordinates or an invalid geodetic datum. The data were subsequently spatially filtered according to the municipality border. The data was additionally filtered to include only records fulfilling the following criteria: (1) Occurrence status classified as “present”; as few datasets include information on absences, this step was needed to have common data structure. (2) Information on both genus and species epithet had to be present to ensure a consistent taxonomic resolution. (3) Coordinate uncertainty of ≤ 354 m, (1/2 length of the diagonal of 500 m × 500 m grid cells). (4) Records had to be registered in year 2010–2012 and 2017–2019 to ensure compatibility with the used land cover maps. (5) Records had to fall within the border of Trondheim Municipality.
The occurrence records were overlaid on the spatial grid, and community matrices for each time point (2010–2012 and 2017–2019) were constructed for all grid cells (species × grid cell). To avoid excessive influence of rare species, only species observed in at least five grid cells during each time step were included. To avoid disproportionate influence of grid cells with minimal sampling effort, only grid cells with at least three observed species were included in the further analyses. The community matrices were converted to presence/absence as unknown sampling effort within grid cells made abundance measures unreliable. The final species occurrence dataset included in the analyses included 151 species, belonging to 45 different families (Table S2). One outlier concerning body size (the whooper swan, Cygnus cygnus) was excluded from further analyses. The Lesser redpoll (Acanthis cabaret) only had data for habitat association; its recognition as a separate species rather than a subspecies of the Common redpoll (A. flammea) is too recent for trait data to have been made available (Billerman et al. 2020), and it was excluded from further analyses.
Statistical analyses
To assess the changes in diversity over time, the temporal β-diversity was calculated for each grid cell using the function beta.temp() from the ‘betapart’ package (Baselga and Orme 2012). Within the package, the options for diversity indices are restricted to the Jaccard and Sørensen families of indices, which have been shown to be roughly equivalent (Baselga 2012). The Jaccard index was used as it does not give extra weight to the number of shared species, as does the Sørensen index (see equations in Baselga & Orme (2012)). As the “nestedness” component of the β-diversity is likely highly influenced by sampling effort, and as the potential changes in functional composition was of interest, the turnover component of the index was used in the analyses. The turnover component (βturnover) of the Jaccard index is formulated as:
in which b indicates the number of species unique to the poorest site (here: time point), and a indicates the number of species shared between the sites (here: time points) (Baselga and Orme 2012).
This means that if species are only lost or gained (i.e. no replacement), the index will equal zero regardless of direction. In the case of species replacement with an equal number of species disappearing and appearing, the value of βturnover depends on the size of the species pool and the number of species being replaced. βturnover = 0.5 can be caused by the number of species lost or gained being exactly half of the number of species remaining in a site. 0 < βturnover < 0.5 can be caused by the number of species lost or gained being smaller than half the number of species remaining in a site. 0.5 < βturnover < 1 can be caused by the number of species lost or gained being larger than half the number of remaining species. The βturnover was logit-transformed to obtain normality.
To evaluate the potential relationship between βturnover and change in developed area, a Generalised Linear Mixed Effects Model (GLMM) predicting βturnover as a function of change in Developed area was constructed. To account for variation in sampling effort (spatially and temporally) additional terms were included: the total number of individual sampling events within the grid cell, and the difference in sampling effort between the time points (Isaac et al. 2014). The latter was calculated as:
Two grid cells were excluded from further analyses, as one proved to be an outlier regarding sampling effort (both in total and for the individual time points), and the other was an outlier regarding difference in sampling effort. The former grid cell is located at a locally popular bird-watching site, the latter is situated in central Trondheim, and the reason for the great difference is unknown. The total number of included grid cells was 356.
As preliminary models showed spatial autocorrelation in the model residuals (i.e. correlation of observations depending on distance, thus leading to fewer independent observations than assumed otherwise), a Matérn correlation function of coordinates (east–west and south-north) was used as a random effect (package ‘spaMM’ (Rousset and Ferdy 2014)). Stepwise backwards model selection was based on AIC. ∆AIC > 2 was used as the threshold for a significantly better model, as recommended by Burnham and Anderson (2004).
To compare the response of various functional groups, species disappearance (present in the first period, not present in the second period) vs. persistent presence (present in both periods), and appearance (not present in the first period, present in the second period) vs. persistent absence were assessed (limited to species in the regional species pool). Rather than assessing (dis-)appearance for each individual species, all species were included in a single model, and various traits of the species were included as covariates, thus evaluating the response of functional groups rather than species. The traits included in the models were adult body mass (g), longevity (years), main habitat association (categorical), main dietary component (categorical) and forage stratum (categorical). The data on traits were obtained from Billerman et al. (2020), Myhrvold et al. (2015), Wilman et al. (2014) and Norsk Ornitologisk Forening (Birdlife Norway) (2020) (Tables 1 and S2). Two binomial GLMMs of the form: (Dis-)appeareance ~ adult body mass + longevity + changeDeveloped area × habitat + changeDeveloped area × diet + changeDeveloped area × forage stratum were constructed; one predicting the log-odds of disappearance, one predicting the log-odds of appearance. To account for variance in sampling effort (spatially and temporally), the total number of individual sampling events within the grid cell, and the difference in sampling effort between the time points, were included as covariates. Grid cell ID was included as a random effect. A Matérn correlation function of distance based on the coordinates of grid cell centre points was included as a random effect to account for spatial autocorrelation. The interaction terms between the categorical trait variables and change in developed area was included to allow for contrasting responses of the functional groups. Stepwise backwards model selection was based on AIC (∆AIC > 2). To evaluate the effects of the linear predictors, the probability of (dis-)appearance were calculated for each variable, keeping all numerical variables constant (changeDeveloped area = 11 964.2 m2 (mean change in the grid cells included in the analyses), longevity = 15 years, no. sampling events = 10 000, difference in sampling events = 0). To evaluate the effects of the interaction terms, the predicted probability of (dis-)appearance were calculated for each factor level across a gradient of change in developed area ranging from min = -28 411.5 m2 to max = 115 540.5 m2 (corresponding the observed minimum- and maximum values), while keeping all other numerical variables constant.
Avian communities in relation to planned land-cover changes
To relate the observed changes in functional composition of avian communities to potential future changes in land-cover, the municipal master plan was provided as shapefiles by Trondheim Municipality in September 2020. The maps describe the land-use element of the municipal master plan regarding areas within the municipality border in the period 2012 to 2024 (pers.comm.). The coding of the management units identified in the local development plan is not directly comparable with the classification used in the AR5 data; the AR5 categories comprise areas understood as land resources, while the planned categories are about land-use understood mostly as human (economic) activities. Thus, re-classifications of both data layers were needed to make the planned area management comparable to current land-cover. Five individual land-cover categories were identified (Table 2). The AR5 map from 2018 was overlaid with the local development plan, and intersect analysis performed, and potential differences in current land-cover and planned land-use were identified. A conservative approach was taken, assuming no planned changes in the case of ambiguous coding; as an example, the AR5-category “Open firm ground” covers land-cover types of various origins, both anthropogenic and non-anthropogenic (e.g. both mountainous bare rock and quarries are classified as “Open firm ground” (Ahlstrøm et al. 2014)). If an area classified as “Open firm ground” intersected an area classified as “Area for Agriculture, Nature, Outdoor-life and Reindeer husbandry”Footnote 1 (“LNFR”) in the municipal master plan, it was re-classified as “LNFR”. If an “Open firm ground” area intersected with an area classified as “Open area” in the master plan (e.g. quarries), it was re-classified as “Open area”. Marine polygons were excluded. The percentage of total terrestrial area within in each category of potential land-cover conversion was calculated (Table 2)Footnote 2.
The models of responses of different functional groups of birds were discussed in the light of the identified potential future land-cover changes to put the results in a local management context.
All data processing and analyses were performed in R version 3.6.3 (R Core Team 2019).
Results
Of the 1 968 grid cells with complete available land-cover data, 356 had sufficient bird occurrence records in both time periods to be included in the models. One hundred and fifty-one bird species belonging to 45 families were included in the calculations of the βturnover index. For the analyses of probability of (dis-)appearance of functional groups, 149 species were included in the models.
Changes in land-cover and planned land-cover changes
Only counting grid cells containing the developed area in question in 2011 or/and 2018, more grid cells showed an increase in developed area (n = 922; 88.1%), compared to grid cells showing decreases (n = 125; 11.9%) or no change (n = 0) between 2011 and 2018. Considering the potential land-cover changes specified in the local development plan, 39.7 km2 of the non-marine area of Trondheim municipality could be facing conversion of land-cover before year 2024 (11.5% of the total 344.6 km2 of non-marine area) (Fig. 2, Table S3). The greatest amount of land (14.6 km2) could potentially be converted to "LNFR" (4.2% of the non-marine area), followed by “Developed area” (3.3%) and “Green structures” (3.2%). Of the total 11.3 km2 potentially facing conversion to “Developed area”, 6.8 km2 are currently classified as “LNFR”.

Sankey diagram of planned land-cover changes. The width of the links are proportional to the area of the specified land-cover type. The left-hand nodes illustrate the land-cover type according to the latest AR5 map; the right-hand nodes illustrate the land-cover category according to the municipal master plan. The categories within the two datasets were not identical, hence the dissimilarities in naming. Colours indicate categories determined to be equivalent. See Table 2 for detailed descriptions of included categories
βturnover in relation to changes in developed area
The βturnover index for bird species (prior to logit-transformation) ranged from 0 to 1 (mean = 0.49, median = 0.47) (Fig. 1b). βturnover = 1 would indicate no species in common between the two time periods, not influenced by the number of species unique to each time period (14 grid cells). βturnover = 0 indicates no species unique to the poorest time point (12 grid cells). Only 11 grid cells had βturnover = 0; 7 caused by only species gain, 4 caused by only species loss. Relatively minor turnover (βturnover < 0.5) was observed in 173 grid cells, whereas relatively large species replacement (βturnover > 0.5) was observed in 133 grid cells. In 25 grid cells, the number of species unique to the poorest time period equalled half of the number of shared species (βturnover = 0.5).
For the models predicting βturnover (i.e. amount of species turnover), the model remaining after AIC-based stepwise backwards model selection (∆AIC < 2 as the threshold for a significantly better model) included only total number of sampling events and spatial dependency as predictors (marginal AIC = 1176.4; conditional AIC = -1753.2) (Table S4). A model including total number of sampling events and the change in Developed area as predictors had a lower marginal AIC (marginal AIC = 1174.9), but higher conditional AIC (conditional AIC = -1604.6). However, the marginal ∆AIC < 2, thus not supporting the model including change in Developed area as a reliable predictor of βturnover.
Responses of bird species functional groups to changes in land-cover
For the models predicting the log-odds of a species disappearing from a grid cell between the two time-points, the best models included change in Developed area, longevity, forage stratum and interactions between change in Developed area, habitat and main dietary component; both total number of sampling events and difference in sampling effort were retained as predictors (Table 3). The probability of a species disappearing decreased with both increasing number of sampling events (Fig. S.2), and with increasing difference in sampling effort (Fig. S.3). Respectively, 0.035% decrease in the odds of disappearance pr. additional sampling event in total, and 6% decrease in the odds of disappearance pr. unit increase in difference in sampling effort. The probability of disappearance decreased with increasing longevity (3% decrease in the odds of disappearance pr. year increase in longevity) (Fig. S.4). Comparisons of predicted probabilities of disappearance for the different forage strata (at change in Developed area = 11 964.2 m2, longevity = 15 years, no. sampling events = 10 000 and difference in sampling effort = 0) revealed no general pattern in differences between the different forage strata (Fig. 3). However, for ground-feeding species the probability of disappearance was higher for wetland-associated species (lowest and highest values of the C.I.'s for the included feeding guilds: 0.06 – 0.45) than for woodland- (0.01 – 0.1), open woodland- (0.007 – 0.07), open- (0.01 – 0.12), urban-associated species (0.01 – 0.1) and generalists (0.02 – 0.14). Likewise, for the species feeding at the mid-high stratum, wetland-associated species had a higher probability of disappearing (lowest and highest values of the C.I.'s for the included feeding guilds: 0.11 – 0.49) than species associated with woodland- (0.01 – 0.11), open woodland- (0.01 – 0.06), open habitats (0.02 – 0.14) and generalists (0.03 – 0.17).

Responses of the predicted probabilities of species disappearance. Predicted probabilities of species disappearance as a function of forage stratum and habitat association. Different colours indicate different main dietary components. Facets indicate different habitat associations. Ribbons indicate 0.95 confidence intervals around the predictions. Longevity was held constant at 15 years, total no. sampling events was fixed at 10 000, difference in sampling effort was fixed at zero, and change in Developed area was fixed at the mean area of change observed in the analysed grid cells: 11 964.2 m2. Spatial effects were removed
Only describing the combinations of variables with observational data available: the predictions across the observed range of change in Developed area, incorporating all levels of the factorial variables, revealed contrasting responses to change in developed area by the different functional groups (Fig. 4). Granivorous, urban species showed a slightly decreasing probability of disappearing with increasing land-cover change (probability of disappearance incl. 0.95 C.I. at the smallest change in developed area vs. the largest change: 0.039 [0.011;0.127] vs. 0.030 [0.034;0.217]). All generalist-, marine-, wetland-, scrub-, water- and woodland species were predicted to have an increasing probability of disappearing, regardless of feeding guild. Of species associated with open areas, insectivores showed a slightly increasing probability of disappearing change (0.046 [0.017;0.116] vs. 0.054 [0.013;0.193] for ground-feeding species, 0.054 [0.020;0.136] vs. 0.063 [0.016;0.223] for understorey feeding ones, 0.056 [0.021;0.139] vs 0.066 [0.016;0.223] for mid-high feeders, 0.059 [0.022;0.149] vs. 0.070 [0.017;0.245] for canopy feeding species and 0.063 [0.023;0.159] vs. 0.074 [0.018;0.258] for aerial feeders); all other feeding guilds showed a decreasing probability of disappearing change. Of species associated with open woodland, granivorous- (0.027 [0.010;0.071] vs. 0.020 [0.005;0.085] for ground feeding species, 0.031 [0.011;0.083] vs 0.024 [0.005;0.1] for understorey feeders) and omnivorous species (0.019 [0.007;0.051] vs. 0.018 [0.004;0.078] for ground feeders, 0.022 [0.008;0.060] vs. 0.022 [0.005;0.091] for understorey feeding species, 0.023 [0.008;0.062] vs. 0.22 [0.005;0.094] for mid-high feeding species) showed a decreasing probability of disappearing, whereas insectivorous (0.017 [0.006;0.045] vs. 0.09 [0.021;0.312] for ground feeding species, 0.02 [0.007;0.053] vs. 0.105 [0.025;0.351] for understorey feeders) and carnivorous/scavenging (0.024 [0.009;0.065] vs. 0.04 [0.008;0.166] for ground feeding species, 0.033 [0.012;0.091] vs. 0.055 [0.012;0.221] for aerial feeders) species showed an increasing probability of disappearing with increasing change in Developed area.

Response curves for the predicted probabilities of species disappearance. Predicted probabilities of species disappearance as a function of change in Developed area, main dietary component, and habitat association. Different colours indicate different forage strata. Ribbons indicate 0.95 confidence intervals around the predictions. Longevity was held constant at 15 years, total no. sampling events was fixed at 10 000 and difference in sampling effort was fixed at zero. Spatial effects were removed
For the models predicting the log-odds of a species appearing in a grid cell between the two time-points, the best models included change in Developed area, longevity, main dietary component and interactions between change in Developed area, habitat and forage stratum; both total number of sampling events and difference in sampling effort were retained as predictors (Table 4). The probability of a species appearing increased with both increasing number of sampling events, and with increasing difference in sampling effort. Respectively, 0.014% increase in the odds of appearance pr. additional sampling event in total (Fig. S.5), and 5.5% increase in the odds of appearance pr. unit increase in difference in sampling effort (Fig. S.6). The probability of appearance increased with increasing longevity (2.9% increase in the odds of appearance pr. year increase in longevity) (Fig. S.7). Comparisons of predicted probabilities of appearance for the different feeding guilds (at change in developed land-cover = 11 964.2 m2, longevity = 15 years, no. sampling events = 10 000 and difference in sampling effort = 0) revealed that the main difference among the various feeding guilds was a slightly higher probability of appearance of omnivorous species, compared to all other groups (Fig. 5).

Responses of the predicted probabilities of species appearance. Predicted probabilities of species appearance as a function of main dietary component and habitat association. Different colours indicate different forage strata. Facets indicate different habitat associations. Ribbons indicate 0.95 confidence intervals around the predictions. Longevity was held constant at 15 years, total no. sampling events was fixed at 10 000, difference in sampling effort was fixed at zero, and change in Developed area was fixed at the mean area of change observed in the analysed grid cells: 11 964.2 m2. Spatial effects were removed
Only describing the combinations of variables with observational data available: the predictions across the observed range of change in Developed area, incorporating all levels of the factorial variables, revealed contrasting responses to change in developed area by the different functional groups (Fig. 6). Open- (granivores: 0.122 [0.06;0.233] vs. 0.317 [0.139;0.571] for ground feeding species, 0.108 [0.051;0.214] vs. 0.662 [0.381;0.861] for mid-high feeders; insectivores: 0.122 [0.06;0.233] vs. 0.317 [0.14;0.57] for ground feeding species, 0.073 [0.033;0.154] vs. 0.421 [0.168;0.724] for understorey feeders, 0.108 [0.051;0.214] vs. 0.662 [0.382;0.861] for mid-high feeding species, 0.113 [0.047;0.248] vs. 0.143 [0.027;0.498] for canopy feeders and 0.097 [0.041;0.213] vs. 0.686 [0.334;0.905] for aerial feeders; omnivores: 0.163 [0.081;0.3] vs. 0.394 [0.185;0.651]; carnivores/scavengers: 0.129 [0.063;0.245] vs. 0.331 [0.147;0.587]), urban- (0.091 [0.034;0.219] vs. 0.553 [0.178;0.876]) and wetland-associated (insectivores: 0.046 [0.017;0.123] vs. 0.079 [0.012;0.371] for ground feeding species, 0.041 [0.014;0.11] vs. 0.266 [0.048;0.72] for mid-high feeders; omnivores: 0.064 [0.023;0.164] vs. 0.108 [0.017;0.453]; carnivores/scavengers: 0.049 [0.018;0.13] vs. 0.084 [0.013;0.387]) species showed increasing probabilities of appearing with increasing urbanisation. Among generalist species, all but ground-feeding species showed increasing probabilities of appearing (0.174 [0.076;0.0351] vs. 0.511 [0.181;0.831] for aerial carnivores/scavengers, 0.192 [0.09;0.366] vs. 0.482 [0.184;0.794] for mid-high feeding carnivores/scavengers, 0.215 [0.102;0.395] vs. 0.181 [0.053;0.467] for ground-feeding carnivores/scavengers, 0.248 [0.114;0.459] vs. 0.278 [0.071;0.662] for omnivores feeding around the water surface, 0.265 [0.131;0.463] vs. 0.226 [0.069;0.536] for ground-feeding omnivores). Among marine species, all but species feeding around the water surface showed decreasing probabilities of disappearance (0.057 [0.02;0.155] vs. 0.005 [0.0005;0.046] for insectivores feeding below the water surface, 0.051 [0.018;0.138] vs. 0.048 [0.006;0.302] for ground-feeding omnivores. For carnivores/scavengers: 0.061 [0.021;0.163] vs. 0.005 [0.0005;0.048] for species feeding below the water surface, 0.036 [0.012;0.1] vs. 0.048 [0.006;0.309] for species feeding around the surface, 0.04 [0.014;0.108] vs. 0.037 [0.004;0.247] for ground-feeders); the trends were however negligible in all cases. Species associated with scrubs (0.3 [0.048;0.782] vs. 0.0006 [0.0000008;0.288]), water (granivores: 0.107 [0.048;0.222] vs. 0.052 [0.013;0.188] for ground-feeding species, 0.1 [0.047;0.2] vs. 0.068 [0.022;0.189] for species feeding around the surface; insectivores: 0.16 [0.073;0.314] vs. 0.007 [0002.;0.035] for species feeding below the surface, 0.1 [0.047;0.198] vs. 0.068 [0.022;0.189] for species feeding around the surface, 0.107 [0.048;0.222] vs. 0.052 [0.013;0.188] for ground feeders; omnivores: 0.211 [0.099;0.392] vs. 0.01 [0.002;0.048] for species feeding below the surface, 0.134 [0.064;0.258] vs. 0.092 [0.031;0.247] for species feeding around the surface; carnivores/scavengers: 0.168 [0.077;0.329] vs. 0.008 [0.002;0.037] for species feeding below the surface, 0.105 [0.049;0.209] vs. 0.072 [0.023;0.199] for species feeding around the surface) or woodland (granivores: 0.382 [0.22;0.575] vs. 0.066 [0.023;0.171] for ground-feeders, 0.35 [0.197;0.542] vs. 0.228 [0.092;0.463] for mid-high feeders, 0.26 [0.134;0.444] vs. 0.099 [0.031;0.273] for understorey-feeders; insectivores: 0382. [0.183;0.589] vs. 0.066 [0.023;0.171] for ground-feeders, 0.26 [0.134;0.444] vs. 0.099 [0.031;0.274] for understorey-feeders, 0.35 [0.197;0.542] vs. 0.229 [0.093;0.463] for mid-high feeders, 0.362 [0.183;0.599] vs. 0.025 [0.004;0.125] for canopy feeders, 0.362 [0.189;0.581] vs. 0.085 [0.02;0.296] for species feeding around the water surface; omnivores: 0.464 [0.284;0.655] vs. 0.09 [0.033;0.224] for ground-feeders, 0.33 [0.178;0.529] vs. 0.134 [0.043;0.346] for understorey-feeders, 0.431 [0.256;0.624] vs. 0.294 [0.125;0.547] for mid-high feeders; carnivores/scavengers: 0.397 [0.231;0.591] vs. 0.07 [0.025;0.18]) all showed decreasing probabilities of appearing with increasing urbanisation. For species associated with open woodland, all species showed increasing probabilities of appearing, except for ground-feeding ones (granivores: 0.28 [0.149;0.463] vs. 0.234 [0.093;0.487] for ground-feeders, 0.181 [0.089;0.334] vs. 0.329 [0.129;0.619] for understorey feeders; insectivores: 0.28 [0.149;0.463] vs. 0.238 [0.093;0.488] for ground-feeders, 0.181 [0.089;0.335] vs. 0.329 [0.129;0.62] for understorey feeders; omnivores: 0.353 [0.198;0.548] vs. 0.305 [0.126;0.572] for ground-feeders, 0.237 [0.12;0.414] vs. 0.401 [0.172;0.696] for understorey feeders, 0.323 [0.173;0.521] vs. 0.649 [0.356;0.861] for mid-high feeders; carnivores/scavengers: 0.243 [0.113;0.446] vs. 0.61 [0.264;0.873] for aerial feeders, 0.293 [0.157;0.48] vs. 0.25 [0.099;0.504] for ground-feeders).

Response curves for the predicted probabilities of species appearance. Predicted probabilities of species appearance as a function of change in Developed area, forage stratum and habitat association. Different colours indicate different main dietary components. Ribbons indicate 0.95 confidence intervals around the predictions. Longevity was held constant at 15 years, total no. sampling events was fixed at 10 000 and difference in sampling effort was fixed at zero. Spatial effects were removed
Discussion
Changes in land-cover, particularly urbanisation, are known to pose threats to biodiversity worldwide. Given the increasing urbanisation globally (United Nations, Department of Economic and Social Affairs 2019), understanding species’ responses to such pressures are essential for guiding conservation measures (Newbold et al. 2018).
In this study we investigated (I) what characterised the land-cover changes within the study area between 2011 and 2018; (II) whether bird species composition change over time could be predicted by the amount of land-cover change; and (III) how various avian functional groups respond to changes in land-cover.
The predominant type of land-cover change within the municipality has been an increase in built-up area, which can be interpreted as extensive urbanisation. Different functional groups respond contrastingly to urbanisation, causing a species turnover over time. βturnover could not be predicted by the change in area classified as “developed” (built-up areas and roads). However, the probability of a bird species either disappearing or appearing in a grid cell depended on both the change in Developed area within the grid cell and on the traits of the species – specifically longevity, habitat association, main dietary component, and forage stratum. Additionally, sampling effort affects the observed changes in community composition. Thus, urbanisation affects which species are either appearing or disappearing. The results thus indicate how urbanisation function as an ecological filter, favouring or impeding different functional groups.
βturnover for birds in relation to changes in land-cover
The (spatially explicit) null-model outperformed the model including change in Developed area as a predictor of βturnover. Thus, contrary to expectation, the amount of change in Developed area does not directly correlate with the extent of bird species turnover on the investigated spatiotemporal scale. This is in concordance with the results of MacLean et al. (2018), who found bird species richness to be relatively stable over time, despite considerable changes in land-cover. However, our model only assesses the extent of species turnover, not the characteristics of the species being replaced; as was also pointed out by MacLean et al. (2018), stability in community level metrics can cover deviations in species composition. An important note is that if the patterns in βturnover were truly random, one would not find any spatial structure in the residuals; the appropriate predictors have simply not been identified in this study at the investigated spatial scale. Direct effects of land-cover change on species turnover is also affected by the degree of spatial autocorrelation identified in the models. Changing the spatial resolution of the study could potentially decrease this effect. This approach is nevertheless complicated by the home ranges of the individual species (and variations therein, see Godet et al. (2018)) – as the different species have large differences in their requirements, finding a single spatial scale appropriate for all species is highly unlikely (Concepción et al. 2015).
The timespan covered in this study is relatively short (approximately seven years). This might be insufficient time for the full effects of urbanisation to have influenced the avian communities, as a considerable biotic lag is expected (Brooks et al. 1999).
Species’ appearance or disappearance
In the probabilistic models of appearance and disappearance, the same predictors were retained in both models: change in developed area, habitat association, main dietary component, forage stratum and longevity. Additionally, variables concerning sampling effort (total number of sampling events and the difference in sampling effort between time periods). However, the interaction terms were different for the two models: disappearance was predicted by an interaction between change in developed area and diet type, while appearance was predicted by an interaction between change in developed area and forage stratum.
The effects of sampling effort are not surprising. The probability of species disappearance decreased with increasing number of sampling events in total, whereas the probability of appearance increased. Both effects are highly intuitive. The same pattern was seen for the difference in sampling effort (here illustrated by the proportional difference in number of sampling events compared to the first time period). This result highlights the importance of including measures of sampling effort in biodiversity modelling, and in particular the importance of accounting for differences in sampling effort (whether that be spatially or temporally) when using open-source, compiled species occurrence records (Isaac et al. 2014; Petersen et al. 2021b).
The models and predictions show that the probability of species disappearance differs somewhat for the different functional groups. Overall, bird species with decreasing probability of disappearing are generally granivorous, urban-associated species or species associated with open habitats. Species with an increasing probability of appearing are generally urban-associated or generalist species, or species associated with open areas or open woodland; however the probability of appearance decreased for ground-feeding species, for generalists and species from open woodland. This trend towards urban- and generalist species is similar to the results of MacLean et al. (2018), who showed an increase in occupancy for species associated with human settlements within landscapes modified over time. In contrast, they found a decrease in occupancy of species associated with open habitat. These urban-associated species responded positively to cities in other studies as well: House Sparrow (Passer domesticus), Rock Pigeon (Columba livia) and Collared Dove (Streptopelia decaocto) (Conole and Kirkpatrick 2011; Evans et al. 2009; Husté and Boulinier 2011). Aronson et al. (2014) found House Sparrow and Rock Pigeon to be cosmopolitan and appearing in more than 80% of their investigated cities globally. The observed effects of feeding guild are somewhat in concordance with the results of Evans et al. (2011), who found birds with plant-based diets to have higher densities in urban- relative to rural areas, and Pinho et al. (2016), who found granivorous species to be associated with urban areas. This is likely related to the relatively low availability of invertebrates in urban- compared to rural areas, and in increase in plant/seed material due to supplementary feeding (Chace and Walsh 2006). Omnivores are more likely to find suitable food items in a heterogeneous landscape. However, the models presented here show that a clear distinction only based on feeding guild is inadequate, as this may interact with other traits.
Forage stratum interacted with the effects of change in developed area on the probability of appearance. Generally, ground-feeding species have decreasing probability of appearing (unless they are associated with urban- or open habitats), whereas above-ground feeding species have positive response to increasing land-cover change more frequently. La Sorte et al. (2018) found that urban bird assemblages had a higher mean percentage use of understorey, mid-storey, canopy and aerial strata for foraging. This is in line with the observed response by species from different forage strata in this study. This likely relates to direct conversion of substrate during urbanisation.
The probability of disappearance decreased with increasing longevity, whereas the probability of appearance increased with increasing longevity. In the first case, the relation can potentially be explained if the species included in the analyses display a higher degree of site fidelity. Long-lived species are less likely to have disappeared within the study period, whereas short-lived ones might have died, and new individuals have not recolonised due to the unfavourable habitat (i.e. species with short lifespans are more dependent on recruitment). The persistence of long-lived species within areas exhibiting great extents of land-cover change and/or urbanisation can thus represent a community not in equilibrium with the environment, with a pending extinction debt to pay, rather than species capable of adapting to disturbance and new conditions (Ramalho and Hobbs 2012). To determine this, longer time-series are needed (Dornelas et al. 2018; Magurran et al. 2019). Secondly, the positive effect of longevity on the probability of appearance is less intuitive. A potential explanation is related to the reasoning behind the relationship seen in the model of disappearance: long-lived species will likely show a greater time-lag in their response to environmental changes (Metzger et al. 2009). Coupled with site fidelity and spatial autocorrelation (individuals originating in nearby grid cells are likely to be observed within the focal grid cell regardless of the environmental suitability), the increasing probability of appearing with increasing life-span might be an artefact rather than a genuine, biological response.
Based on the retention of interaction terms in the models, our results agree with the conclusions of Croci et al. (2008) and Kark et al. (2007) – only combinations of traits indicates whether a bird species is (pre-)adapted for urban life and changing land-cover, not any single traits. Croci et al. (2008) found that urbanisation functions as a biological filter on bird functional traits, but only on a regional (not local) scale; they found traits to differ between urban avoiders and –adapters at a regional (city-wide) scale, but not between very- and moderately urbanophilic species at a local (0.5 – 2 ha) scale. For comparison, the spatial resolution of this study is intermediate (0.25 km2 = 25 ha).
Changes in land-cover
“LNFR” was a rather ambiguous category covering most of the terrestrial area of Trondheim, including both forests, cultivated land, mires and open areas; thus, specific planned land-cover changes for this category were unavailable. Additionally, conversion between “LNFR” and “Green structures” is unlikely to lead to significant changes in actual land-cover, as the land-use element of the municipal master plan refers to human (economic) activities (Kommunal- og moderniseringsdepartementet 2020). However, transition from “LNFR” or “Green structures” to “Developed area” or anthropogenic “Open areas” entail the possibility of vegetation removal and urban development. As the land-cover index used in models of bird’s responses to land-cover change mainly correlated with changes in built-up area (urbanisation), the potential land-cover conversion from “LNFR” or “Green structures” towards either “Developed area” or anthropogenic “Open areas” are of primary concern.
Consequences for conservation and management
Unsurprisingly, the vast majority of the examined functional groups show negative responses to increasing urbanisation (i.e. increasing probability of disappearance or decreasing probability of appearance); urbanisation thus have the potential to impoverish local avian biodiversity. Particularly species associated with wetlands, woodlands and marine areas are at risk of disappearing with increasing urbanisation. Petersen et al. (2020) predicted coastal areas and open mires in Trondheim to harbour relatively high numbers of threatened species, and mires and bogs have been a habitat strongly affected by land-use changes since the 1950s (IPBES 2018). In combination, this warrants particular focus on the protection of these areas.
Given the planned potential land-cover changes within the municipality in the near future, this calls for concern (Trondheim 2013). Ca. forty km2 (11.5%) of the municipality’s non-marine area potentially face changes in land-cover within the current management plan, which lasts until 2024; of these, approximately 11 km2 (3%) could be converted to built-up areas. In comparison, ca. 15 km2 (4%) changed land-cover category between 2011 and 2018 (approximately 10.9 km2 changed category to Developed area). The second largest change category is conversion to developed/built-up area, which will direly affect the local avian communities. Specifically, ca. 7 km2 are conversions from forest, cultivated land, open (natural) areas, mires and grasslands to developed area. If these potential land-cover changes are realised, it will likely affect local community composition direly. A large area can potentially be converted from developed area to land-cover types with a smaller anthropogenic pressure and thus reflect maintenance of “blue-green” areas; in the suggested plan program of the municipal sector plan for biodiversity 2021–2032 (Miljøenheten 2020), the concept of “area neutrality” has been suggested as a further management tool. “Area neutrality” refers to the practice of ecological compensation, and thus requires the restoration of an equal area as is destroyed otherwise. However, given the inherent disturbance and biotic lag following such a conversion, this is unlikely to compensate within a foreseeable future. It is important to note that the local development plan covers the period from 2012 to 2024, and thus includes the time periods used in the analyses, complicating the interpretations. Nevertheless, as the potential changes in land-cover management identified here are relative to the land-cover maps latest updated in 2018, the conclusions are still valid.
Biodiversity dynamics in urban areas are found to be complex and maintenance complicated (Elmqvist et al. 2013, 2016); but as the city expands, wood- and wetland specialists disappear, and urban species and generalists take over. Based on these results, a recommendation for local management to maintain bird biodiversity is to minimise the anthropogenic pressure on vulnerable habitats and –species.
Availability of data and material
Species occurrence data is available from public repository (GBIF Occurrence Download 10 .15468/dl.nxxuv6 accessed via GBIF.org on 2020–04-26).
Code availability
All R code written to perform the data download and analyses is available through GitHub repository at https://github.com/tanjakofodpetersen/Landcover_change.git
Notes
Translated from the original Norwegian classification: “Landbruks-, natur- og friluftsformål samt reindrift”.
Agricultural-, Nature- and Outdoor purposes together with Reindeer husbandry (LNFR).
References
Ahlstrøm AP, Bjørkelo K, Frydenlund J (2014) AR5 klassifikasjonssystem. Klassifikasjon av arealressurser. Norsk institutt for skog og landskap report 06/2014. ISBN: 978–82–311–0211–3
Amano T, Lamming JDL, Sutherland WJ (2016) Spatial gaps in global biodiversity information and the role of citizen science. Bioscience 66:393–400. https://doi.org/10.1093/biosci/biw022
Aronson MFJ, La Sorte FA, Nilon CH, Katti M, Goddard MA, Lepczyk CA, Warren PS, Williams NSG, Cilliers S, Clarkson B, Dobbs C, Dolan R, Hedblom M, Klotz S, Kooijmans JL, Kühn I, MacGregor-Fors I, McDonnel M, Mörtberg U, Pyšek P, Siebert S, Sushinsky J, Werner P, Winter M (2014) A global analysis of the impacts of urbanization on bird and plant diversity reveals key anthropogenic drivers. Proc R Soc b: Biol Sci 281:20133330. https://doi.org/10.1098/rspb.2013.3330
Baselga A (2012) The relationship between species replacement, dissimilarity derived from nestedness, and nestedness. Glob Ecol and Biogeogr 21:1223–1232. https://doi.org/10.1111/j.1466-8238.2011.00756.x
Baselga A, Orme CDL (2012) Betapart: An R package for the study of beta diversity. Methods Ecol Evol 3:808–812. https://doi.org/10.1111/j.2041-210X.2012.00224.x
Billerman SM, Keeney BK, Rodewald PG, Schulenberg TS (2020) Birds of the World. Cornell Laboratory of Ornithology, Ithaca, NY, USA. https://birdsoftheworld.org/bow/home
Bivand RS, Pebesma E, Gómez-Rubio V (2013) Applied spatial data analysis with R (Second edi). Springer, New York
Blair RB (1996) Land use and avian species diversity along an urban gradient. Ecol Appl 6:506–519. https://doi.org/10.2307/2269387
Bonthoux S, Barnagaud J, Goulard M, Balent G (2013) Contrasting spatial and temporal responses of bird communities to landscape changes. Oecologia 172:563–574. https://doi.org/10.1007/s00442-012-2498-2
Bregman TP, Lees AC, MacGregor HEA, Darski B, de Moura NG, Aleixo A, Barlow J, Tobias JA (2016) Using avian functional traits to assess the impact of land-cover change on ecosystem processes linked to resilience in tropical forests. Proc R Soc b: Biol Sci 283:20161289. https://doi.org/10.1098/rspb.2016.1289
Brooks TM, Pimm SL, Oyugi JO (1999) Time lag between deforestation and bird extinction in tropical forest fragments. Conserv Biol 13:1140–1150. http://www.jstor.org/stable/2641747
Buitrago L (2020) GBIF issues & flags. GBIF data blog. https://data-blog.gbif.org/post/issues-and-flags/. Accessed on 06 Jun 2022
Burnham KP, Anderson DR (2004) Multimodel inference: Understanding AIC and BIC in model selection. Sociol Method Res 33:261–304. https://doi.org/10.1177/2F0049124104268644
Chace JF, Walsh JJ (2006) Urban effects on native avifauna: a review. Landsc Urb Plan 74:46–69. https://doi.org/10.1016/j.landurbplan.2004.08.007
Climate-Data.org (2022) Klima Trondheim (Norge). https://no.climate-data.org/europa/norge/sør-trøndelag-fylke/trondheim-707/. Accessed 30 Apr 2022
Concepción ED, Moretti M, Altermatt F, Nobis MP, Obrist MK (2015) Impacts of urbanisation on biodiversity: The role of species mobility, degree of specialisation and spatial scale. Oikos 124:1571–1582. https://doi.org/10.1111/oik.02166
Conole LE, Kirkpatrick JB (2011) Functional and spatial differentiation of urban bird assemblages at the landscape scale. Landsc Urb Plan 100:11–23. https://doi.org/10.1016/j.landurbplan.2010.11.007
Croci S, Butet A, Clergau P (2008) Does urbanization filter birds on the basis of their biological traits? The Condor 110:223–240. https://doi.org/10.1525/cond.2008.8409
Díaz S, Settele J, Brondízio ES, Ngo HT, Agard J, Arneth A, Balvanera P, Brauman KA, Butchart SHM, Chan KMA, Garibaldi LA, Ichii K, Liu J, Subramanian SM, Midgley GF, Miloslavich P, Molnár Z, Obura D, Pfaff A, Polasky S, Purvis A, Razzaque J, Reyers B, Chowdhury RR, Shin YJ, Visseren-Hamakers I, Willis KJ, Zayas CN (2019) Pervasive human-driven decline of life on Earth points to the need for transformative change. Science 13:366. https://doi.org/10.1126/science.aax3100
Dornelas M et al (2018) BioTIME: A database of biodiversity time series for the Anthropocene. Glob Ecol Biogeogr 27:760–786. https://doi.org/10.1111/geb.12729
Elmqvist T, Fragkias M, Goodness J, Güneralp B, Marcotullio PJ, McDonald RI, Parnell S, Schewenius M, Sendestad M, Seto KC, Wilkinson K (2013) Urbanization, biodiversity and ecosystem services: challenges and opportunities: a global assessment. Springer, Dordrecht Heidelberg
Elmqvist T, Gómez-Baggethun E, Langemeyer J (2016) Ecosystem services provided by urban green infrastructure. In: Potschin M, Haines-Young R, Fish R, Kerry Turner R (2016) Routledge Handbook of Ecosystem Services, pp 452–468
Ernoult A, Tremauville Y, Cellier C, Margerie P, Langlois E, Alar D (2006) Potential landscape drivers of biodiversity components in a flood plain: Past or present patterns? Biol Conserv 127:1–17. https://doi.org/10.1016/j.biocon.2005.07.008
Evans BS, Reitsma R, Hurlbert AH, Marra PP (2018) Environmental filtering of avian communities along a rural-to-urban gradient in Greater Washington, D.C., USA. Ecosphere 9: e02402. https://doi.org/10.1002/ecs2.2402
Evans KL, Newson SE, Gaston KJ (2009) Habitat influences on urban avian assemblages. Ibis 151:19–39. https://doi.org/10.1111/j.1474-919X.2008.00898.x
Evans KL, Chamberlain DE, Hatchwell BJ, Gregory RD, Gaston KJ (2011) What makes an urban bird? Glob Change Biol 17:32–44. https://doi.org/10.1111/j.1365-2486.2010.02247.x
Gaiji S, Chavan V, Ariño AH, Otegui J, Hobern D, Sood R, Robles E (2013) Content assessment of the primary biodiversity data published through GBIF network: Status, challenges and potentials. Biodiversity Informatics 8:94–172. https://doi.org/10.17161/bi.v8i2.4124
Gaston KJ (2005) Biodiversity and extinction: species and people. Prog Phys Geogr 29:239–247. https://doi.org/10.1191/2F0309133305pp445pr
GBIF.org (2019) GBIF home page. https://www.gbif.org/. Accessed on 26 Apr 2020
Godet L, Harmange C, Marquet M, Joyeux E, Fournier J (2018) Differences in home-range sizes of a bird species in its original, refuge and substitution habitats: challenges to conservation in anthropogenic habitats. Biodivers Conserv 27:719–732. https://doi.org/10.1007/s10531-017-1460-3
Hagen EO, Hagen O, Ibáñez-Álamo JDm Petchey OL, Evans KL, (2017) Impacts of urban areas and their characteristics on avian functional diversity. Front Ecol Evol 5:1–15. https://doi.org/10.3389/fevo.2017.00084
Hausner VH, Yoccoz NG, Ims RA (2003) Selecting indicator traits for monitoring land use impacts: birds in northern coastal birch forests. Ecol Appl, 13: 999–1012. https://www.jstor.org/stable/4134738
Hijmans RJ (2020) raster: geographic data analysis and modeling. R package. https://CRAN.R-project.org/package=raster
Husté A, Boulinier T (2011) Determinants of bird community composition on patches in the suburbs of Paris, France. Biol Conserv 144:243–252. https://doi.org/10.1016/j.biocon.2010.08.022
Ikin K, Knight E, Lindenmayer DB, Fischer J, Manning AD (2012) Linking bird species traits to vegetation characteristics in a future urban development zone: Implications for urban planning. Urb Ecosyst 15:961–977. https://doi.org/10.1007/s11252-012-0247-2
IPBES (2018) The IPBES regional assessment report on biodiversity and ecosystem services for Europe and Central Asia. Rounsevell M, Fischer M, Torre-Marin Rando A, Mader A. (eds.). Secretariat of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services, Bonn, Germany
IPBES (2019) Summary for policymakers of the global assessment report on biodiversity and ecosystem services of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services. Díaz S, Settele J, Brondízio ES, Ngo HT, Guèze M, Agard J, Arneth A, Balvanera P, Brauman KA, Butchart SHM, Chan KMA, Garibaldi LA, Ichii K, Liu J, Subramanian SM, Midgley GF, Miloslavich P, Molnár Z, Obura D, Pfaff A, Polasky S, Purvis A, Razzaque J, Reyers B, Roy ChowdhuryR, Shin YJ, Visseren-Hamakers IJ, Willis KJ, Zayas CN (eds.). IPBES secretariat, Bonn, Germany
Isaac NJB, van Strien AJ, August TA, de Zeeuw MP, Roy DB (2014) Statistics for citizen science: Extracting signals of change from noisy ecological data. Methods Ecol Evol 5:1052–1060. https://doi.org/10.1111/2041-210X.12254
Johnson EA, Miyanishi K (2008) Testing the assumptions of chronosequences in succession. Ecol Lett 11:419–431. https://doi.org/10.1111/j.1461-0248.2008.01173.x
Kark S, Iwaniuk A, Schalimtzek A, Banker E (2007) Living in the City : Can anyone become an ’Urban Exploiter’? J Biogeogr 34:638–651. https://doi.org/10.1111/j.1365-2699.2006.01638.x
Kartverket (2019) SOSI-standarder. https://kartverket.no/geodataarbeid/standarder/sosi/. Accessed on 04 May 2022
Kaye JP, Groffman PM, Grimm NB, Baker LA, Pouyat RV (2006) A distinct urban biogeochemistry? Trends Ecol Evol 21:192–199. https://doi.org/10.1016/j.tree.2005.12.006
Kinnunen RP, Fraser KC, Schmidt C, Garroway CJ (2022) The socioeconomic status of cities covaries with avian life-history strategies. Ecosphere 13:e3918. https://doi.org/10.1002/ecs2.3918
Kommunal- og moderniseringsdepartementet (2020) Planlegging for spredt bolig- , fritids- og næringsbebyggelse i landbruks- , natur- , frilufts- og reindriftsområder. https://www.regjeringen.no/no/dokumenter/planlegging-for-spredt-bolig--fritids--og-naringsbebyggelse-i-landbruks--natur--frilufts--og-reindriftsomrader-lnfr-omrader/id2696910/?ch=1. Accessed 04 May 2022
La Sorte FA, Lepczyk CA, Aronson MFJ, Goddard MA, Hedblom M, Katti M, MacGregor-Fors I, Mörtberg U, Nilon CH, Warren PS, Williams NSG, Yang J (2018) The phylogenetic and functional diversity of regional breeding bird assemblages is reduced and constricted through urbanization. Divers Distrib 24:928–938. https://doi.org/10.1111/ddi.12738
MacLean SA, Rios Dominguez AF, de Valpine P, Beissinger SR (2018) A century of climate and land-use change cause species turnover without loss of beta diversity in California’s Central Valley. Glob Change Biol 24:5882–5894. https://doi.org/10.1111/gcb.14458
Magurran AE, Dornelas M, Moyes F, Henderson PA (2019) Temporal β diversity—A macroecological perspective. Glob Ecol Biogeogr 28:1949–1960. https://doi.org/10.1111/geb.13026
McKinney ML (2006) Urbanization as a major cause of biotic homogenization. Biol Conserv 127:247–260. https://doi.org/10.1016/j.biocon.2005.09.005
Metzger JP, Martensen AC, Dixo M, Bernacci LC, Ribeiro MC, Teixeira AMG, Pardini R (2009) Time-lag in biological responses to landscape changes in a highly dynamic Atlantic forest region. Biol Conserv 142:1166–1177. https://doi.org/10.1016/j.biocon.2009.01.033
Miljøenheten (2020) Kommunedelplan for naturmangfold 2021–2032. Forslag til planprogram. https://www.trondheim.kommune.no/globalassets/10-bilder-og-filer/10-byutvikling/miljoenheten/klima-og-energi/kdp-naturmangfold_forslag-til-planprogram_horing-2020.pdf. Accessed 04 May 2022
Moen A (1999) National atlas of Norway: vegetation . Norwegian Mapping Authority, Hønefoss
Myhrvold NP, Baldridge E, Chan B, Sivam D, Freeman DL, Morgan Ernest SK (2015) An amniote life-history database to perform comparative analyses with birds, mammals, and reptiles. Ecol 96:3109–3109. https://doi.org/10.1890/15-0846R.1
Newbold T (2010) Applications and limitations of museum data for conservation and ecology, with particular attention to species distribution models. Progr Phys Geogr 34:3–22. https://doi.org/10.1177/2F0309133309355630
Newbold T, Hudson LN, Hill SLL, Contu S, Lysenko I, Senior RA, Börger L, Bennett DJ, Choimes A, Collen B, Day J, De Palma A, Díaz S, Echeverria-Londoño S, Edgar MJ, Feldman A, Garon M, Harrison MLK, Alhusseini T, Ingram DJ, Itescu Y, Kattge J, Kemp V, Kirkpatrick L, Kleyer M, Correia DLP, Martin CD, Meiri S, Novosolov M, Pan Y, Philips HRP, Purves DW, Robinson A, Simpson J, Tuck SL, Weiher E, White HJ, Ewers RM, Mace GM, Scharlemann JPW, Purvis A (2015) Global effects of land use on local terrestrial biodiversity. Nature 520:45–50. https://doi.org/10.1038/nature14324
Newbold T, Hudson LN, Contu S, Hill SLL, Beck J, Liu Y, Meyer C, Philips HRP, Scharlemann JPW, Purvis A (2018) Widespread winners and narrow-ranged losers: Land use homogenizes biodiversity in local assemblages worldwide. PLoS Biol 16:1–25. https://doi.org/10.1371/journal.pbio.2006841
Norsk Ornitologisk Forening (Birdlife Norway) (2020) Fuglekunnskap, Norske fugler. https://www.birdlife.no/fuglekunnskap/fugleatlas/. Accessed 04 May 2022
Norwegian Institute of Bioeconomy Research (2018) AR5. https://www.nibio.no/tema/jord/arealressurser/arealressurskart-ar5/. Accessed 04 May 2022
Padayachee AL, Irlich UM, Faulkner KT, Gaertner M, Proches S, Wilson JRU, Rouget M (2017) How do invasive species travel to and through urban environments? Biol Invasions 19:3557–3570. https://doi.org/10.1007/s10530-017-1596-9
Palacio FX, Ibañez LM, Maragliano RE, Montalti D (2018) Urbanization as a driver of taxonomic, functional, and phylogenetic diversity losses in bird communities. Can J Zool 96:1114–1121. https://doi.org/10.1139/cjz-2018-0008
Pebesma E (2018) Simple features for R: standardized support for spatial vector data. The R J 10:439–446. https://doi.org/10.32614/RJ-2018-009
Pebesma E, Bivand R (2005) Classes and methods for spatial data in R R News 5. https://cran.r-project.org/doc/Rnews/
Petersen TK, Speed JDM, Grøtan V, Austrheim G (2020) Urban aliens and threatened near-naturals : Land-cover affects the species richness of alien- and threatened species in an urban-rural setting. Sci Rep 10:1–14. https://doi.org/10.1038/s41598-020-65459-2
Petersen TK, Speed JDM, Grøtan V, Austrheim, (2021a) Competitors and ruderals go to town: plant community composition and function along an urbanisation gradient. N J Bot 39:1–14. https://doi.org/10.1111/njb.03026
Petersen TK, Speed JDM, Grøtan V, Austrheim G (2021b) Species data for understanding biodiversity dynamics : The what, where and when of species occurrence data collection. Ecol Solut Evid 2:1–17. https://doi.org/10.1002/2688-8319.12048
Pinho P, Correia O, Lecoq M, Munzi S, Vasconcelos S, Gonçalves P, Rebelo R, Antunes C, Silva P, Freitas C, Lopes N, Santos-Reis M, Branquinho C (2016) Evaluating green infrastructure in urban environments using a multi-taxa and functional diversity approach. Environ Res 147:601–610. https://doi.org/10.1016/j.envres.2015.12.025
Powney GD, Isaac NJB (2015) Beyond maps: A review of the applications of biological records. Biol J Linnean Soc 115:532–542. https://doi.org/10.1111/bij.12517
R Core Team (2019) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Ramalho CE, Hobbs RJ (2012) Time for a change: dynamic urban ecology. Trends Ecol Evol 27:179–188. https://doi.org/10.1016/j.tree.2011.10.008
Rittenhouse CD, Pidgeon AM, Albright TP, Culbert PD, Clayton MK, Flather CH, Masek JG, Radeloff VC (2012) Land-cover change and avian diversity in the conterminous United States. Conserv Biol 26:821–829. https://www.jstor.org/stable/23255335
Rodrigues ASL, Brooks TM (2007) Shortcuts for biodiversity conservation planning: the effectiveness of surrogates. Annu Rev Ecol Evol Syst 38:713–737. https://doi.org/10.1146/annurev.ecolsys.38.091206.095737
Rousset F, Ferdy JB (2014) Testing environmental and genetic effects in the presence of spatial autocorrelation. Ecography 37:781–790. https://doi.org/10.1111/ecog.00566
Sala OE, Chapin S, Armesto JJ, Berlow E, Bloomfield J, Dirzo R, Huber-Sanwald E, Huenneke LF, Jackson RB, Kinzig A, Leemans R, Lodge DM, Mooney HA, Oesterheld M, Poff NL, Sykes MT, Walker BH, Walker M, Wall DH (2000) Global biodiversity scenarios for the year 2100 global biodiversity scenarios for the year 2100. Science 287:1770–1774. https://doi.org/10.1126/science.287.5459.1770
Sekercioglu CH (2006) Increasing awareness of avian ecological function. Trends Ecol Evol 21:464–471. https://doi.org/10.1016/j.tree.2006.05.007
Sepp T, McGraw KJ, Kaasik A, Giraudeau M (2018) A review of urban impacts on avian life-history evolution: Does city living lead to slower pace of life? Glob Change Biol 24:1452–1469. https://doi.org/10.1111/gcb.13969
Speed JDM, Bendiksby M, Finstad AG, Hassel K, Kolstad AL, Prestø T (2018) Contrasting spatial, temporal and environmental patterns in observation and specimen based species occurrence data. PLoS Biol 13:1–17. https://doi.org/10.1371/journal.pone.0196417
Statistics Norway (2020) Statistisk Sentralbyrå. https://www.ssb.no/. Accessed 04 May 2022
Tiago P, Ceia-Hasse A, Marques TA, Capinha C, Pereira HM (2017) Spatial distribution of citizen science casuistic observations for different taxonomic groups. Sci Rep 7:1–9. https://doi.org/10.1038/s41598-017-13130-8
Trondheim K (2013) Temaplan for naturmiljøet i Trondheim. https://www.trondheim.kommune.no/tema/bygg-kart-og-eiendom/arealplaner/temaplaner-prosjekter-og-utredninger/markaplanen/temaplan-for-naturmiljoet/. Accessed 04 May 2022
Troudet J, Grandcolas P, Blin A, Vignes-Lebbe R, Legendre F (2017) Taxonomic bias in biodiversity data and societal preferences. Sci Rep 7:1–14. https://doi.org/10.1038/s41598-017-09084-6
United Nations, Department of Economic and Social Affairs, Population Division (2019) World urbanization prospects: the 2018 revision (ST/ESA/SER.A/420). New York: United Nations
Wilman H, Belmaker J, Simpson J, de la Rosa C, Rivadeneira MM, Jetz W (2014) EltonTraits 1.0: Species-level foraging attributes of the world’s birds and mammals. Ecology 95:E095-178. https://doi.org/10.1890/13-1917.1
Acknowledgements
We thank the personnel of Trondheim Municipality for providing data and descriptions of land-cover and local development plans.
Funding
Open access funding provided by NTNU Norwegian University of Science and Technology (incl St. Olavs Hospital - Trondheim University Hospital).
Author information
Authors and Affiliations
Contributions
TKP, GA, JDMS and VG conceived the idea and designed the methodology; TKP and YKF retrieved and analysed the data; TKP wrote the first draft of the manuscript. All authors contributed critically to the drafts and gave final approval for publication.
Corresponding author
Ethics declarations
Conflicts of interest
The authors declare no conflicts of interest.
Ethics approval
Not applicable.
Consent to participate
Not applicable.
Consent for publication
Not applicable.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Petersen, T.K., Speed, J.D.M., Grøtan, V. et al. Urbanisation and land-cover change affect functional, but not compositional turnover of bird communities. Urban Ecosyst 25, 1679–1698 (2022). https://doi.org/10.1007/s11252-022-01258-z
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11252-022-01258-z