
Abstract
Animals’ adaptation to life in cities is a widely studied issue in ecology. One such adaptation is the adjustment of stress response to new conditions and anthropogenic disturbances. There are various examples of such adjustments in urban birds, but the stress response of non-avian species has rarely been investigated. We studied an urban and a rural population of a small mammal, the striped field mouse Apodemus agrarius. We compared baseline corticosterone (CORT) concentration in faecal samples from mice, using ELISA tests with antibodies for this hormone. We found that although urban specimens have more anthropogenic stressors than their rural equivalents, they do not have an elevated level of CORT. This confirms the hypothesis that urban animals undergo hormonal adjustment to urban conditions. Moreover, the CORT concentrations of urban mice were lower compared to their rural conspecifics, while for body weight we observed an inverse relationship. A CORT level allows urban mammals to redirect more energy to weight gain (body fat); this is probably an effect of the lower pressure of competitors in urban areas, as well as better food availability (access to anthropogenic food). The results of this work show that species that have managed to overcome fear of humans and adapt to anthropogenic stressors can therefore achieve measurable ecological benefits.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Rapid worldwide urbanization is causing a decline in the richness and abundance of many native species (Chace and Walsh 2006; McKinney 2008). On the other hand, some species are able to adapt to life in cities and create stable populations there (Møller 2009; Francis and Chadwick 2012). The adaptation of these animals to life in extremely man-made environments such as cities is a widely studied issue in ecology (Bonier 2012; Birnie-Gauvin et al. 2016; Vincze et al. 2016). It is generally believed that understanding the mechanisms responsible for animal synurbization will allow the prediction of further changes in wildlife in anticipation of further global urbanization (Fokidis et al. 2009; Bateman and Fleming 2012; Brearley et al. 2012; Birnie-Gauvin et al. 2016).
Recently many studies of animals in urban areas have focused on their behaviour, morphology, and physiology in comparison to non-urban populations of these species (Bonier 2012; Brearley et al. 2012; Giraudeau and McGraw 2014; Meillere et al. 2015; Vincze et al. 2016). It is considered that urban habitats can shape endocrine traits, and that these traits might contribute to adaptation to urban environments (Bonier 2012). Glucocorticoid hormones may play a major role in mediating responses to urbanization within wild species. Glucocorticoids are secreted by the adrenal glands (as part of the hypothalamic-pituitary-adrenal axis system) and are often referred to as the stress hormone. However, their role in physiology is much broader because they play a multifaceted role in energy balance and homeostasis (Lucas and French 2012; Fanson et al. 2017). Under baseline (nonstressed) conditions, when the energetic needs of an organism are perceived as stable, the level of glucocorticoids varies predictably across a circadian or circannual rhythm (Romero 2002, 2004; Sapolsky 2002; Malisch et al. 2007). In response to psychological and physical stressors, the level of glucocorticoids sometimes rises dramatically above baseline levels (Malisch et al. 2007; Lucas and French 2012).
Urbanization radically changes biotic and abiotic elements of environments and exposes animals to novel stimuli (novel challenges) that have not occurred in their evolutionary history. As a result, urban animals face an almost completely different set of physiological and behavioural challenges compared to their rural counterparts (Birnie-Gauvin et al. 2016). This should result in increased secretion of glucocorticoids in an effort to maintain homeostasis (Bonier et al. 2007; Rebolo-Ifran et al. 2015). Because chronic elevation of glucocorticoids is harmful for the organism (Sapolsky et al. 2000; Korte et al. 2005; Bonier et al. 2007), urban animals should adjust their stress response to new conditions. This reduced stress response is presumably necessary for all animals that thrive in ecosystems exposed to frequent anthropogenic disturbances, such as those in urban areas (Partecke et al. 2006; Fokidis and Deviche 2011; Rebolo-Ifran et al. 2015; Vincze et al. 2016; Tryjanowski et al. 2016). There are numerous examples of such adaptations in urban populations of birds (Chavez-Zichinelli et al. 2010; Fokidis and Deviche 2011; Zhang et al. 2011; Bokony et al. 2012; Meillere et al. 2015; Rebolo-Ifran et al. 2015; Angelier et al. 2016; Potvin et al. 2016). With a few exceptions, the stress responses of non-avian species in urban environments have rarely been investigated (Lucas and French 2012; Brearley et al. 2012; Francis et al. 2015; Siutz and Millesi 2012).
The aim of this study was to investigate whether small mammals living in green city areas (subjected to anthropogenic stressors such as anthropogenic noise, vehicular traffic, light pollution, human presence, and high fragmentation of available habitats) differ in terms of their levels of glucocorticoids (corticosterone), compared to their conspecifics in areas subjected to much less human influence (rural areas). We studied an urban population of the striped field mouse Apodemus agrarius (Pallas, 1771). This species is known to be one of the most synurbic small mammal species within its geographical range (Andrzejewski et al. 1978; Łopucki et al. 2013). We hypothesized that since this species is considered to be well adapted to urban conditions, the corticosterone levels in individuals from urban populations would not be different from those typical for rural conspecifics.
Methods
Study area
The study was carried out in the city of Lublin (Poland; Europe; 51°15 N; 22°33 E) and a surrounding area of up to 10 km from the administrative boundaries of the city.
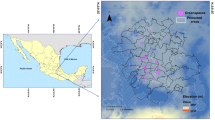
Lublin has an area of 147.5 km2 and a human population of about 350,000. Within the urban area, the study was conducted only in the most urbanized central part of the city (area about 2.87 km2), in 10 suitable green areas where the occurrence of small mammals was probable, i.e. patches with dense undergrowth or ground cover (Babińska-Werka et al. 1979). Suitable patches in this part of the city included: three wooded areas with a park-like character (sites 1, 2 and 3); one old neglected orchard (site 4); one complex of allotments (i.e. a complex of small plots of land cultivated by city inhabitants) (site 5); one wide strip of roadside trees (site 6); and four partly wooded unmanaged areas with ruderal vegetation (patches 7–10) (Fig. 1). It was known from previous studies (Łopucki et al. 2013; Łopucki and Kiersztyn 2015) that green areas in this part of the city are inhabited mainly by striped field mice.

Location of urban trapping sites in the city of Lublin
Near the city, studies were also conducted on 10 sites including fallow land in various stages of ecological succession (with herbs, grasses, segetal or ruderal plants and shrubs). These habitats are suitable for catching striped field mice and are optimal for obtaining samples from a rural population of this species (Łopucki et al. 2013).
Collection of faecal samples in the field
In order to collect faecal samples, live trapping of small mammals was carried out and samples were collected from the traps in which the targeted species were caught. Trapping was conducted for 10 days in both the urban and the rural sites, i.e. over a period of 20 days, alternately 1 day in urban and 1 day in rural sites. Each trapping session was carried out at a different location to eliminate or reduce the probability of catching the same individuals or disturbing the population by repetition of the trapping procedure. Rodents were captured in wooden live trap boxes containing food bait (oats). We usually used 60–100 traps per trapping session.
The field procedure of faecal sample collection for testing corticosterone level is described in the literature (Navarro-Castilla et al. 2014a, b; Łopucki et al. 2018): it requires collecting samples at specific times of the day and animals are held in the traps for a short period. We set traps at about midnight and checked after 5 h. This procedure (1) reduced the possible effect of capture on the corticosterone level as faecal corticosterone metabolites appear in faeces 8–12 h after a stressful episode (Touma et al. 2004), and (2) reduced the effect of daily differences in hormone excretion (Touma et al. 2003) as we only collected the fresh samples at the same times of the day, at the end of the nocturnal period of rodent activity (as in the works of Navarro-Castilla et al. 2014a, b). All captured animals were described in terms of species, sex, and body mass (±1 g), and were subsequently released at the site of capture. Faecal samples were collected only from traps containing the targeted species, i.e. the striped field mouse. From all such traps, we collected as many faeces pellets as possible that were not contaminated with urine. They were placed in an Eppendorf tube, immediately cooled in ice, and transported to the laboratory, where they were stored in a freezer at −20 °C until analysis. The traps in which animals were caught were taken to the laboratory, cleaned under running water, air dried and then reused.
In total, 202 striped field mice were captured during the entire trapping period; 85 in urban (40 males and 45 females) and 117 in rural areas (60 males and 57 females). Within the city, the striped field mice accounted for 90.4% of all captured animals, whereas in non-urbanized areas this was 38.1%. The remaining mammals captured during the study belonged to the following species: (1) urban sites – yellow-necked mouse A. flavicollis (Melchior, 1834) (8.5%) and Eurasian harvest mouse Micromys minutus (Pallas, 1771) (1.1%); (2) rural sites – yellow-necked mouse and wood mouse A. sylvaticus (L., 1758) (22.2%), common vole Microtus arvalis (Pallas, 1778) (17.3%), bank vole Myodes glareolus (Schreber, 1780) (9.4%), Eurasian harvest mouse (8.4%), common shrew Sorex araneus (L., 1758) (2.5%), root vole M. oeconomus (Pallas, 1776) (1.6%) and field vole M. subterraneus (de Selys-Longchamps, 1836) (0.6%).
Trapping sessions were carried out in January 2017. We chose this time of year so as to conduct the research during the non-reproductive season, when all the individuals in the population have the same reproductive status (differences in reproductive status may affect the level of corticosterone Harper and Austad 2001; Harris and Saltzman 2013; Romero 2002). In Poland, the breeding season of mice and voles usually ends in September or October, but some papers indicate that it may last longer in urban populations (Andrzejewski et al. 1978). For our study we wanted to choose the period when the breeding season was over and the next had not yet not started. In the climate of Poland, January is the optimal month for this purpose. In the study region, January is the coldest month of the year (mean temperatures below −5 °C).
Measurement of faecal corticosterone
The laboratory procedure of measuring faecal corticosterone (CORT) has already been described by Łopucki et al. (2018). Briefly, each faecal sample was weighed with an accuracy of 0.001 g, and only subsamples of 0.1 g of wet mass were included in the analysis. The remaining samples were weighed, dried for 8 h at 45 °C, and weighed again to determine their dry mass content. For analysis of CORT concentrations in the faeces, we used a commercial test kit containing antibodies for this hormone (CORT ELISA Kit No. EU3108; Wuhan Fine Biological Technology Co.). The advertised sensitivity (minimal detectable amount) was lower than 46.87 pgCORT ml−1. Analysis was performed according to the manufacturer’s protocol (described in Łopucki et al. 2018). To validate the experiment procedure and the appropriateness of the sample dilution proportion, a pilot experiment was conducted with 8 samples using standards. The pilot experiment showed that the values obtained for samples with undiluted supernatant were within the calibration (standard) curve, therefore these undiluted supernatants were used in analysis. The range of the standard curve was 78.12 to 5000 pg ml−1. To account for the differences in the mass of the analysed samples, faecal CORT concentrations were expressed as nanograms of CORT per one gram of dry mass of faeces (ng g−1).
Statistical analyses
To analyse the drivers of the faecal CORT concentrations in the studied species, we applied a multivariate generalized linear model using a gamma distribution with a log link function. The data was not normally distributed and transformation to normality failed. Faecal CORT concentration was a response variable; location (urban or rural), sex, and interaction of sex and location were assessed as fixed factors, while body mass was estimated as a covariate. Due to similar limitations, the second model compared body mass using the same model type. In this model, the mass was a response variable, while location, sex, and interaction of sex and location were assessed as fixed factors. We compared various model types and a null model to achieve the best-fit model using the Akaike information criterion (AICc) in backward elimination. In both models, gamma distribution with a log link function presented the best values of AICc. The models were constructed with SPSS software (24.0).
We also compared distribution of CORT concentrations in the urban and rural populations of the studies species. We used a chi square test to find the differences between the populations based on the frequency of CORT concentration in the defined ranges. The urban population presented an empirical distribution, while the rural population presented a theoretical distribution. In this comparison, we used four ranges of CORT concentration: 0–3.999 ngCORT g−1 (low level); 4–7.999 ngCORT g−1 (medium); 8–11.999 ngCORT g−1 (high); >12 ngCORT g−1 (very high). In determining these ranges, we used the method proposed by Chavez-Zichinelli et al. (2013), who used the mean value + SD of CORT concentration to identify individuals with higher than expected CORT concentrations and to establish the physiological CORT thresholds that could affect the animals’ health. Such a level should be determined for a population living in the least disturbed habitat; in our case, this was the population from rural areas. The calculated threshold for our data was about 12 ng g−1.
Results
The best model for faecal CORT concentrations in the striped field mouse included only location and was highly statistically significant (χ2 = 18.89; df = 1; P = 0.000). Sex, interaction of sex and location, and body mass were not statistically significant (P > 0.05) and were excluded during the AICc procedure (Table 1). The mean level of faecal CORT concentrations in mice from urban areas was 4.22 ng g−1, which was over 30% lower than the mean CORT concentrations in mice from rural sites (6.59 ng g−1) (Fig. 2). The difference was highly statistically significant (χ2 = 20.07; df = 1; P = 0.000).

Faecal corticosterone concentrations in 1 g of sample (mean ± SE) in the striped field mice with regard to location: within the city area (Urban), the control area (Rural), and pairwise comparison in GLM with Bonferroni adjustment (χ2 = 20.07; df = 1; P = 0.000; n = 85 for Urban, and n = 117 for Rural)
The faecal CORT concentrations in the striped field mice in the urban area were not similarly distributed compared to the rural sites (χ2 = 14.75; df = 3, P < 0.001). The most frequent CORT concentration in both the urban and rural populations was in the range 0–3.999 ng g−1, with a slightly higher proportion in the former than the latter. There was a similar decrease in the number of animals in each subsequent range, i.e. only slightly fewer samples were found in the ranges 4–7.999 and 8–11.999 ng g−1, but a very high CORT level of over 12 ng g−1 was found in only 15% of samples from the rural population (Fig. 3). Differences between the CORT values found in the city and rural populations were also observed in quartiles. Q1 presented similar values in both populations (2.85 for city and 2.47 for rural); the median (3.79 for city and 4.58 for rural) and Q3 values were more distant (5.03 for city and 7.88 for rural). The range of CORT level in the city population (0.32–9.74 ng g−1) was much narrower than in the rural population (0.86–32.31 ng g−1).

Distribution of faecal CORT concentrations in 1 g of sample in the striped field mice in the urban area (Urban) and the rural area (Rural); distributions statistically differ (χ2 = 14.75; df = 3, P < 0.001)
The best model for body mass in the study species included location and sex as explanatory variables (Table 2). The model was highly statistically significant (χ2 = 13.23; df = 2; P = 0.001). The AICc backward elimination procedure excluded non-significant interaction between these two factors. Individuals from the urban population were significantly heavier than mice from the rural population (19.69 g and 18.53 g respectively, χ2 = 8.75; df = 1; P = 0.003) (Fig. 4). Although the mean body mass differed only slightly between females and males (19.5 g and 18.7 g respectively), this difference was statistically significant (χ2 = 4.29; df = 1; P = 0.038).

Body mass (mean ± SE) of the striped field mice relating to a) location (Urban and Rural), b) sex (Females and Males), and pairwise comparison in GLM with Bonferroni adjustment (for location: χ2 = 8.75; df = 1; P = 0.003; n = 85 for Urban and n = 117 for Rural; for sex: χ2 = 4.29; df = 1; P = 0.038; n = 102 for Females, and n = 100 for Males)
Discussion
We expected that despite living in a stressful anthropogenic environment, the urban population of mice would show a similar CORT level as their rural conspecifics as an effect of physiological adjustment of their stress response to the urban environment. Such a phenomenon is often described in literature concerning the endocrine ecology of urban avian species (Chavez-Zichinelli et al. 2010; Fokidis and Deviche 2011; Zhang et al. 2011; Bokony et al. 2012; Meillere et al. 2015; Rebolo-Ifran et al. 2015; Angelier et al. 2016; Potvin et al. 2016). We found, however, that our results are not entirely consistent with the general pattern described for birds.
Differences in the CORT levels of the urban and rural populations
Although striped field mice living in urbanized areas have greater anthropogenic stressors than those in rural areas, they do not have an elevated corticosterone level. Such a result indicates that the expected reduction of stress response occurs in this species. However, the mean level of CORT concentrations in mice from urban areas was lower compared to their conspecifics from rural sites. One interpretation of this is that the hormonal response of animals living in the city is not only reduced to the level found in non-urbanized environments, but also that the urban conditions may be less stressful than rural conditions in certain situations. A similar reduction in urban hormone levels has also been described in other species. French et al. (2008) showed that urbanized tree lizards Urosaurus ornatus exhibited lower baseline and stress-induced CORT concentrations than rural ones. Lodjak and Maegi (2014) found that rural rooks Corvus frugilegus from urban colonies had lower baseline faecal CORT levels than their rural counterparts.
Our results also allowed a more detailed insight into the described phenomenon. It should be noted that the higher mean level of CORT in the rural population is caused by some mice which displayed an atypical elevated level of corticosterone. About 15% of the population had this very high (sensu Chavez-Zichinelli et al. 2013) level of CORT; this phenomenon is related to neither the sex nor the body mass of the individuals. Therefore, two questions arise: (1) What kind of stress did we observe, acute or chronic?; (2) What was the stressor? In our opinion, the results show that the effect of acute rather than chronic stress was observed because the latter should also induce a weight loss effect (Wang et al. 2011; Chavez-Zichinelli et al. 2013). At such a critical time of the year, i.e. winter, chronic exposure to highly stressful factors would be reflected in body mass since energy needs are greatly increased at that time (Sipari et al. 2016). For these reasons, it is more likely that individuals with a very high level of CORT had rather stressful episodes in the last dozen or so hours and were therefore suffering from acute rather than chronic stress. The paper by Touma et al. (2004) showed that peak CORT concentrations in mice faeces can be observed with a delay of about 8–12 h after the onset of a stressful episode. What could this acute stressor be? From the groups of abiotic and biotic stressors presented in by Sheriff and Thaler (2014), it is most probable that the stressor is predators or competitors. If the stressors were abiotic factors, they should have an impact on more than only 15% of the population, whereas if stressors were biotic factors such as lack of food or disease, they should also have a significant impact on body weight and have a chronic rather than acute character.
A predator-dependent hypothesis seems to be the most suitable option to explain our results because contact with predators or with traces of their scent (in the case of mammalian predators) may cause a large increase in CORT concentration (Harris and Saltzman 2013). However, we intentionally did not investigate the pressure of predators in the studied areas and the proposed explanation should only be considered as a hypothesis (as Romero 2004 suggests). In the rural study area we observed at least four wild predatory species that significantly affect rodents: the least weasel Mustela nivalis; the red fox Vulpes vulpes; the common buzzard Buteo buteo; and an unidentified owl species, as well as domesticated predators such as cats and dogs. On the other hand, wild predators can also be found in urban areas; their species composition is reduced but those occur at far higher densities than in rural areas (Crooks 2002; Luniak 2004; Bateman and Fleming 2012). Furthermore, the abundance of domestic predators (mainly cats) in cities can be extremely high, although their overall predation pressure during winter is relatively low (Krauze-Gryz et al. 2017). Without metric data about the pressure of predators in our study sites, it is difficult to determine whether this factor was responsible for the fact that 15% of studied rodents from the rural population had elevated levels of CORT.
Other than the stressful effect of predators, another explanation of our results is antagonistic contact between the studied rodent and other species of small mammals, including resource competitors (other species of mice or voles) and nest predators, i.e. shrews (Liesenjohann et al. 2013). In the rural areas the pressure from competitive species is relatively high because the studied species accounted for only 38% of all captured small mammals. Aggressive interactions often play an important role in the ecology of closely related species and affect their coexistence (Simeonovska-Nikolova 2007). The striped field mouse may display superiority over some species or be subordinate to other species (Frynta et al. 1995; Simeonovska-Nikolova 2007). These considerations suggest that such antagonistic interspecific episodes can potentially increase corticosterone levels in the studied animals living in multispecies communities. In contrast to rural areas, in urban sites the pressure of competitors is much lower, as is evident in our results and in the literature (Andrzejewski et al. 1978; Łopucki et al. 2013; Łopucki and Kitowski 2017), therefore there are fewer antagonistic interspecific episodes. An unanswered question relates to the competitive relationship between the striped field mouse and brown rats Rattus norvegicus in urban areas, in which the mouse is apparently in a subordinate position. We did not catch any rats during our winter trapping sessions, therefore it seems that in this case a division of spatial niches occurs: the mouse lives in green areas, while the rat is more associated with buildings and places of gathering of anthropogenic waste (Sacchi et al. 2008).
Differences in the body mass of the urban and rural population
The individuals from urban areas had a significantly higher body mass than their rural conspecifics; this may indicate that lower levels of corticosterone allow them to manage their energy better. More energy can be redirected to weight gain (body fat) because they do not expend energy on interspecific competition (as described above). Differences in diet could also explain the body mass differences between urban and rural mice. Babińska-Werka (1981) found that the diet of urban mice contained more calories and (apart from seeds and insect larvae) droplets of fat probably derived from human food. This means that urban mice with access to anthropogenic food sources (food scraps – a practically unlimited source of food) gain a measurable benefit compared to their conspecifics in rural areas, where the pool of available food is limited and its quantity and quality decrease during the winter. Urban areas, on the other hand, tend to offer predictable and abundant food resources all year round (Chavez-Zichinelli et al. 2013). The higher calorific value of the food and the lack of need to devote energy to anti-predator and competitive behaviour mean that urban mice may expend more energy on gaining weight, which would explain our results concerning body mass and correspond with results concerning lower CORT concentrations.
Conclusions
As has previously been described for bird species, the studied mammal species also did not have an elevated level of corticosterone in urban conditions. This could confirm the hypothesis concerning the hormonal adjustment of urban populations of animals to urban conditions. In addition, the lower pressure of competitors as well as better food conditions (access to anthropogenic food) mean that individuals in the urban population have a reduced level of corticosterone relative to their rural conspecifics. Better living conditions in the city in winter could also be a direct cause of the higher weight of individuals in the urban population. The results of this work are another example showing that species that have managed to overcome fear of humans and adapt to anthropogenic stressors can therefore achieve measurable evolutionary benefits.
References
Andrzejewski R, Babińska-Werka J, Gliwicz J, Goszczyński J (1978) Synurbization process in population of Apodemus agrarius. I. Characteristics of populations in urbanization gradient. Acta Theriol 23:341–358
Angelier F, Meillère A, Grace JK, Trouvé C, Brischoux F (2016) No evidence for an effect of traffic noise on the development of the corticosterone stress response in an urban exploiter. Gen Comp Endocrinol 232:43–50
Babińska-Werka J (1981) Food of the striped field mouse in different types of urban green areas. Acta Theriol 26:285–299
Babińska-Werka J, Gliwicz J, Goszczyński J (1979) Synurbization processes in an urban population of Apodemus agrarius. II. Habitats of the striped field mouse in town. Acta Theriol 26:405–415
Bateman PW, Fleming PA (2012) Big city life: carnivores in urban environments. J Zool 287:1–23
Birnie-Gauvin K, Peiman KS, Gallagher AJ, de Bruijn R, Cooke SJ (2016) Sublethal consequences of urban life for wild vertebrates. Environ Rev 24:416–425
Bokony V, Seress G, Sz N et al (2012) Multiple indices of body condition reveal no negative effect of urbanization in adult house sparrows. Landsc Urban Plan 104:75–84
Bonier F (2012) Hormones in the city: endocrine ecology of urban birds. Horm Behav 61:763–772
Bonier F, Martin PR, Sheldon KS, Jensen JP, Foltz SL, Wingfield JC (2007) Sex-specific consequences of life in the city. Behav Ecol 18:121–129
Brearley G, McAlpine C, Bell S, Bradley A (2012) Influence of urban edges on stress in an arboreal mammal: a case study of squirrel gliders in Southeast Queensland, Australia. Landsc Ecol 27:1407–1419
Chace JF, Walsh JJ (2006) Urban effects on native avifauna: a review. Landsc Urban Plan 74:46–69
Chavez-Zichinelli CA, MacGregor-Fors I, Talamas RP et al (2010) Stress responses of the House Sparrow (Passer domesticus) to different urban land uses. Landsc Urban Plan 98:183–189
Chavez-Zichinelli CA, MacGregor-Fors I, Quesada J et al (2013) How stressed are birds in an urbanizing landscape? Relationships between the physiology of birds and three levels of habitat alteration. Condor 115:84–92
Crooks KR (2002) Relative sensitivities of mammalian carnivores to habitat fragmentation. Conserv Biol 16:488–502
Fanson KV, Best EC, Bunce A, Fanson BG, Hogan LA, Keeley T, Narayan EJ, Palme R, Parrott ML, Sharp TM, Skogvold K, Tuthill L, Webster KN, Bashaw M (2017) One size does not fit all: monitoring faecal glucocorticoid metabolites in marsupials. Gen Comp Endocrinol 244:146–156
Fokidis HB, Deviche P (2011) Plasma corticosterone of city and desert curve-billed thrashers, Toxostoma curvirostre, in response to stress-related peptide administration. Comp Biochem Phys A 159:32–38
Fokidis HB, Orchinik M, Deviche P (2009) Corticosterone and corticosteroid binding globulin in birds: relation to urbanization in a desert city. Gen Comp Endocrinol 160:259–270
Francis RA, Chadwick MA (2012) What makes a species synurbic? Appl Geogr 32:514–521
Francis MJ, Spooner PG, Matthews A (2015) The influence of urban encroachment on squirrel gliders (Petaurus norfolcensis): effects of road density, light and noise pollution. Wildl Res 42:324–333
French SS, Fokidis HB, Moore MC (2008) Variation in stress and innate immunity in the tree lizard (Urosaurus ornatus) across an urban-rural gradient. J Comp Physiol B 178:997–1005
Frynta D, Exnerová A, Nováková A (1995) Intraspecific behaviour interactions in the striped field-mouse (Apodemus agrarius) and its interspecific relationships to the wood mouse (Apodemus sylvaticus): dyadic encounters in a neutral cage. Acta Soc Zool Bohem 59:53–62
Giraudeau M, McGraw KJ (2014) Physiological correlates of urbanization in a desert songbird. Integr Comp Biol 54:622–632
Harper JM, Austad SN (2001) Effect of capture and season on fecal glucocorticoid levels in deer mice (Peromyscus maniculatus) and red-backed voles (Clethrionomys gapperi). Gen Comp Endocrinol 123:337–344
Harris BN, Saltzman W (2013) Effect of reproductive status on hypothalamic–pituitary–adrenal (HPA) activity and reactivity in male California mice (Peromyscus californicus). Physiol Behav 112–113:70–76
Korte SM, Koolhaas JM, Wingfield JC, McEwen BS (2005) The Darwinian concept of stress: benefits of allostasis and costs of allostatic load and the trade-offs in health and disease. Neurosci Biobehav Rev 29:3–38
Krauze-Gryz D, Zmihorski M, Gryz J (2017) Annual variation in prey composition of domestic cats in rural and urban environment. Urban Ecosyst 20:945–952
Liesenjohann M, Liesenjohann T, Palme R, Eccard JA (2013) Differential behavioural and endocrine responses of common voles (Microtus arvalis) to nest predators and resource competitors. BMC Ecol 13:33
Lodjak J, Maegi M (2014) Differences in nest site selection and baseline faecal corticosterone levels between urban and rural rooks (Corvus frugilegus). Hirundo 27:33–53
Łopucki R, Kiersztyn A (2015) Urban green space conservation and management based on biodiversity of terrestrial fauna – a decision support tool. Urban For Urban Gree 14:508–518
Łopucki R, Kitowski I (2017) How small cities affect the biodiversity of ground-dwelling mammals and the relevance of this knowledge in planning urban land expansion in terms of urban wildlife. Urban Ecosyst 20:933–943
Łopucki R, Mróz I, Berliński Ł, Burzych M (2013) Effects of urbanization on small-mammal communities and the population structure of synurbic species: an example of a medium-sized city. Can J Zool 91:554–561
Łopucki R, Klich D, Ścibior A, Gołębiowska D, Perzanowski K (2018) Living in habitats affected by wind turbines may result in an increase in corticosterone levels in ground dwelling animals. Ecol Indic 84:165–171
Lucas LD, French SS (2012) Stress-induced tradeoffs in a free-living lizard across a variable landscape: consequences for individuals and populations. PLoS ONE 7:e49895
Luniak M (2004) Synurbization—adaptation of animal wildlife to urban development. In: Shaw WW, Harris KL, VanDruff L (eds) Proceedings of the 4th international symposium on urban wildlife conservation, Tucson, Ariz., 1–5 May 1999. University of Arizona, Tucson, pp 50–55
Malisch JL, Saltzman W, Gomes FR, Rezende EL, Jeske DR, Garland Jr T (2007) Baseline and stress-induced plasma corticosterone concentrations of mice selectively bred for high voluntary wheel running. Physiol Biochem Zool 80:146–156
McKinney ML (2008) Effects of urbanization on species richness: a review of plants and animals. Urban Ecosyst 11:161–176
Meillere A, Brischoux F, Parenteau C, Angelier F (2015) Influence of urbanization on body size, condition, and physiology in an urban exploiter: a multi-component approach. PLoS ONE 10:e0135685
Møller AP (2009) Successful city dwellers: a comparative study of the ecological characteristics of urban birds in the Western Palearctic. Oecologia 159:849–858
Navarro-Castilla A, Barja I, Olea PO, Piñeiro A et al (2014a) Are degraded habitats from agricultural crops associated with elevated faecal glucocorticoids in a wild population of common vole (Microtus arvalis)? Mamm Biol 79:36–43
Navarro-Castilla A, Mata C, Ruiz-Capillas P, Palme R, Malo JE, Barja I (2014b) Are motorways potential stressors of roadside wood mice (Apodemus sylvaticus) populations? PLoS ONE 9:e91942
Partecke J, Schwabl I, Gwinner E (2006) Stress and the city: urbanization and its effects on the stress physiology in European blackbirds. Ecology 87:1945–1952
Potvin DA, Curcio MT, Swaddle JP, MacDougall-Shackleton SA (2016) Experimental exposure to urban and pink noise affects brain development and song learning in zebra finches (Taenopygia guttata). PEERJ 4:e2287
Rebolo-Ifran N, Carrete M, Sanz-Aguilar A et al (2015) Links between fear of humans, stress and survival support a non-random distribution of birds among urban and rural habitats. Sci Rep-UK 5:13723
Romero LM (2002) Seasonal changes in plasma glucocorticoid concentrations in free-living vertebrates. Gen Comp Endocrinol 128:1–24
Romero ML (2004) Physiological stress in ecology: lessons from biomedical research. Trends Ecol Evol 19:249–255
Sacchi R, Gentilli A, Pilon N, Bernini F (2008) GIS-modelling the distribution of Rattus norvegicus in urban areas using non toxic attractive baits. Hystrix It J Mamm 19:13–22
Sapolsky RM (2002) Endocrinology of the stress-response. In: Becker JB, Breedlove SM, Crews D, McCarthy MM (eds) Behavioral endocrinology. MIT Press, Cambridge, pp 409–450
Sapolsky RM, Romero LM, Munck AU (2000) How do glucocorticoids influence stress responses? Integrating permissive, suppressive, stimulatory, and preparative actions. Endocr Rev 21:55–89
Sheriff MJ, Thaler JS (2014) Ecophysiological effects of predation risk; an integration across disciplines. Oecologia 176:607–611
Simeonovska-Nikolova D (2007) Interspecific social interactions and behavioral responses of Apodemus agrarius and Apodemus flavicollis to conspecific and heterospecific odors. J Ethol 25:41–48
Sipari S, Haapakoski M, Klemme I, Palme R, Sundell J, Ylönen H (2016) Population sex-ratio affecting behavior and physiology of overwintering bank voles (Myodes glareolus). Physiol Behav 159:45–51
Siutz C, Millesi E (2012) Effects of birth date and natal dispersal on faecal glucocorticoid concentrations in juvenile common hamsters. Gen Comp Endocrinol 178:323–329
Touma C, Sachser N, Möstl E, Palme R (2003) Effects of sex and time of day on metabolism and excretion of corticosterone in urine and faeces of mice. Gen Comp Endocrinol 130:267–278
Touma C, Palme R, Sachser N (2004) Analyzing corticosterone metabolites in fecal samples of mice: a noninvasive technique to monitor stress hormones. Horm Behav 45:10–22
Tryjanowski P, Moller AP, Morelli F et al (2016) Urbanization affects neophilia and risk-taking at bird-feeders. Sci Rep-UK 6:28575
Vincze E, Papp S, Preiszner B, Seress G, Bókony V, Liker A (2016) Habituation to human disturbance is faster in urban than rural house sparrows. Behav Ecol 27:1304–1313
Wang Z, Wang B, Lu J (2011) Behavioral and physiological responses of striped field mice (Apodemus agrarius) to predator odor. Integr Zool 6:334–340
Zhang S, Lei F, Liu S, Li D, Chen C, Wang P (2011) Variation in baseline corticosterone levels of Tree Sparrow (Passer montanus ) populations along an urban gradient in Beijing, China. J Ornithol 152:801–806
Acknowledgements
We would like to thank the anonymous reviewers for their suggestions that improved the quality of the paper. The authors gratefully acknowledge use of the services and facilities of the Centre for Interdisciplinary Research of The John Paul II Catholic University of Lublin, Lublin, Poland, co-funded by the European Union from the European Regional Development Fund in the frame of the Operational Programme Development of Eastern Poland 2007-2013 (POPW.01.03.00-06-003/09-00).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Łopucki, R., Klich, D., Ścibior, A. et al. Hormonal adjustments to urban conditions: stress hormone levels in urban and rural populations of Apodemus agrarius. Urban Ecosyst 22, 435–442 (2019). https://doi.org/10.1007/s11252-019-0832-8
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11252-019-0832-8