
Abstract
Although large and medium-sized herbivorous mammals avoid urbanized areas, they have recently begun to colonize towns and cities. In general, ungulates continue to avoid the centres of urban areas, and utilize mainly their thinly built-up outskirts. While extension of urban development is preventing ungulates from penetrating the urban landscape, the influence of noise and light pollution on the occurrence of mammalian herbivores is still poorly understood. Hence, we investigated the hypothesis that habitat availability shapes the distribution of roe deer Capreolus capreolus and artificial lightening discourages them from penetrating the urban landscape. Roe deer was recorded on 37% of randomly selected sample plots (N = 60) located within the city of Kraków (S Poland). The occupied plots contained significantly more open habitats, woodland patches were larger in them, but proximity to rivers, and noise and light pollution were significantly lower. The logistic regression model revealed that an increasing area of open habitats was positively correlated with the probability of roe deer occurring. However, the artificial lighting at night was negatively correlated with the probability of the species occurring: the negative effect of light pollution was mitigated by the greater area of open habitats. Our study highlights the very considerable potential of light pollution as a predictor of the occurrence of large mammals in the urban landscape. We argue that urbanization and the related artificial lighting at night may be a factor preventing ungulates from penetrating potentially suitable habitats in urban areas.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The encroachment of urbanization on to natural terrain usually means a loss of species from the latter (Mcdonald et al. 2008; Bateman and Fleming 2012) or their adaptation to this new habitat (McCarthy et al. 1996; Acevedo et al. 2011). As a result, urban areas become an important habitat for certain groups of animals, in which these can achieve high population densities (Mcdonald et al. 2008; Ciach and Fröhlich 2017). Ungulates are animals which have only quite recently begun to colonize urban areas, but so far with rather moderate success (Kilpatrick and Spohr 2000; Acevedo et al. 2005; Underwood and Kilheffer 2016). In general, large mammalian herbivores avoid strongly urbanized areas (Underwood and Kilheffer 2016; Loro et al. 2016). Small populations of ungulates colonize mainly suburban areas (McCarthy et al. 1996; Kilpatrick and Spohr 2000; Mattila and Hadjigeorgiou 2015; Loro et al. 2016), with only small numbers of individuals entering densely built-up areas (Warren 2011). Moreover, the colonization of suburbs by ungulates is often of a transitional nature, resulting from overabundances in adjacent areas (Warren 2011; Mattila and Hadjigeorgiou 2015). However, the considerable behavioural plasticity of ungulates (Kilpatrick and Spohr 2000; Acevedo et al. 2011; Morellet et al. 2011; Kušta et al. 2017) may intensify their colonization of towns and cities. It is possible, therefore, that such areas will become significant habitats for this group of mammals.
Colonizing the urban landscape, however, is fraught with danger for terrestrial animals (Forman and Alexander 1998; Harrington and Conover 2006; Padié et al. 2015). Expansion of road networks in ungulate habitats hinders movements between habitat patches, which in turn restricts access to food and limits gene exchange, giving rise to a general deterioration in individual condition (Forman and Alexander 1998; Hewison et al. 2009; Seidler et al. 2015). Also, road traffic often leads to collisions with vehicles (Bruinderink and Hazebroek 1996; Forman and Alexander 1998; Zuberogoitia et al. 2014) and/or modifies the animals’ natural diurnal activity (Kušta et al. 2017). Traffic noise also interferes with the animals’ vocal communication and may impact on their courtship and vigilance (Nemeth et al. 2013; Klett-Mingo et al. 2016).
Human habitations pose a further threat to land animals (McCarthy et al. 1996; Hewison et al. 2001; Underwood and Kilheffer 2016): not only do they cause their habitats to shrink (Underwood and Kilheffer 2016; Loro et al. 2016), there is increased pressure on the part of humans and their dogs, which frequently flush out wild animals, raising stress levels and inflicting energy losses in the latter (Reimoser 2012; Padié et al. 2015). The encroachment of ungulates on to land used by humans often leads to conflicts (Putman and Moore 1998; Warren 2011; Mattila and Hadjigeorgiou 2015): as a consequence, habitats may be fenced off (Boone and Hobbs 2004; Harrington and Conover 2006) and the animals culled (Brown et al. 2000; Warren 2011). Yet another danger for terrestrial animals may be light pollution, which can interfere with their orientation abilities and thus with their movements (Longcore and Rich 2004; Beier 2006), while long-term exposure to artificial lighting can disrupt their biological rhythm (Yeates 1949; Lincoln and Guinness 1972; Barber-Meyer 2007); they may therefore tend to avoid strongly illuminated areas (Robert et al. 2015).
In the temperate zone, ungulates inhabit mainly woodland and open habitats (farmland, grassland) (Mattioli 2011; Morellet et al. 2011); at the same time, however, they demonstrate a number of adaptations to life in the neighbourhood of human habitations (Kilpatrick and Spohr 2000; Torres et al. 2011; Kušta et al. 2017). Adapting diurnal activity to road traffic helps to avoid collisions with vehicles (Kušta et al. 2017), and diminished activity during hunting periods may limit mortality (Pagon et al. 2013). Moreover, despite the potential threat from humans (hunters), ungulates are able to tolerate the presence of human habitations (Mysterud et al. 1999; Hewison et al. 2001) and the associated activities of people (Bonnot et al. 2013; Padié et al. 2015). In certain cases, ungulates can also take advantage of human-dependent food resources, which saves or helps to gain energy (Putman 1986; Kilpatrick and Spohr 2000). Milder temperatures are normal in large cities (Arnfield 2003), so theoretically ungulates may find favourable conditions there in winter, because the thinner snow cover is no obstacle to foraging (Mysterud et al. 1999; Ewald et al. 2014). Another important factor is that populations of the main natural enemies of ungulates – large and medium-sized predators like grey wolf Canis lupus and lynx Lynx lynx (Molinari-Jobin et al. 2002; Mattioli et al. 2004; Bateman and Fleming 2012) – are not present in urban areas. Predation by humans (hunting) is limited for reasons of safety; there is also greater awareness of / empathy for these animals on the part of the urban populace (Brown et al. 2000; Lee and Miller 2003; Warren 2011).
One of the ecologically more flexible ungulate species is the roe deer Capreolus capreolus, which inhabits woodlands and open country, and also a mosaic of both (Putman 1986; Morellet et al. 2011; Ewald et al. 2014). The habitats utilized by roe deer are often situated in a landscape matrix that includes areas transformed by humans to varying extents (McCarthy et al. 1996; Acevedo et al. 2005; Torres et al. 2011; Loro et al. 2016). The results of studies done to date on the effects of urbanization on roe deer are equivocal: although this species seems to avoid areas with a moderate degree of urbanization (Hewison et al. 2001; Loro et al. 2016), it may occasionally prefer the proximity of thinly urbanized areas (Torres et al. 2011). The issue of roe deer encroaching into strongly urbanized areas has not been explored as yet. Furthermore, not many studies of the influence of urbanization on roe deer populations have been conducted during winter, when ungulates can forage on the supplementary food available in human settlements (Putman 1986; Kilpatrick and Spohr 2000; Torres et al. 2011). Apart from examining the influence of the urban landcover (Underwood and Kilheffer 2016; Loro et al. 2016) and the proximity of buildings (Hewison et al. 2001; Torres et al. 2011), studies to date have not looked at the effect of the noise and artificial light produced by towns and cities, which may discourages roe deer from entering them, even if potentially suitable habitats are available there. Understanding the effects limiting the exploration by roe deer of urban areas is important in the context of their frequent collisions with road vehicles (Bruinderink and Hazebroek 1996; Found and Boyce 2011), the important role they play in shaping the vegetation (Welch et al. 1991; Putman and Moore 1998) and the contribution they make to the diet of some medium and large predators (Molinari-Jobin et al. 2002; Mattioli et al. 2004; Bateman and Fleming 2012).
The aim of this study was to assess the factors governing the winter distribution of roe deer in the urban environment. We assessed the role of (a) habitat type within the urban landscape and (b) noise and light pollution in the shaping of the species’ distribution pattern. We hypothesized that the preferred habitat, which is woodland and/or open habitats in the case of roe deer (Mattioli 2011; Morellet et al. 2011), would be of primary importance. We predicted, however, that noise and light pollution – intense and ever-present in urban environments – would tend to discourage roe deer from entering them and would be a factor governing the probability of their occurring in the urban environment.
Methods
Study area
This study was carried out in the city of Kraków (southern Poland, 50°05’ N, 19°55′ E), which covers an area of 327 km2 and has a population density of 2321 persons/km2 (GUS 2016). Built-up areas cover around 6% of city’s overall area and are represented by an urbanization gradient – from the densely built-up city centre, through the suburbs with a moderate number of buildings to the scattered buildings typical of a rural landscape. Urban greenery is the predominant form of vegetation in the city (47%), consisting of gardens, squares, road verges and playgrounds, allotments and orchards, parks and cemeteries and other green areas. The next most common group of habitats consists of open areas (37%): arable land (14%), spontaneous vegetation on fallow land (13%), meadows and pastures (8%), wetland vegetation (2%). The remainder of the city’s green areas consists of forests and natural woodland (11%): natural and semi-natural scrub (5%), deciduous and mixed forest (4%) and damp, riparian forest and transformed tree stands (2%). Roads and railway lines make up 4% and surface waters just 1% of the city’s area (Dubiel and Szwagrzyk 2008). The principal waterway in Kraków is the River Wisła (Vistula); six medium-sized tributaries and numerous smaller watercourses flow into the Wisła within the city limits (MIIP 2016). The mean number of days with snow cover varies between 60 and 70 days and its mean thickness is 20 cm (maximum recorded was 65 cm); however, both these parameters are showing a long-term decrease indicative of systematic climate warming (Falarz 1998). The urban heat island of Kraków is estimated to cover the city area and surrounding regions: there is an up to 1 cm difference in the average depth of snow cover between the city centre and the outskirts, but no differences in the mean number of days with snow cover (Falarz 1998; Arnfield 2003). During the two study years, there was not much snow cover during the winter: it was limited to 12 of winter days in 2013/2014 (mean snow cover thickness = 0.9 cm ±2.5 SD, maximum = 9 cm) and 28 of winter days in 2014/2015 (mean snow cover thickness = 1.5 cm ±2.7 SD, maximum = 13 cm).
Selection of sample plots; roe deer counts
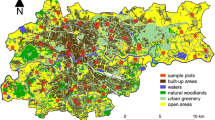
Sixty sample plots on which roe deer were to be counted (Fig. 1) were selected at random using Quantum GIS software (QGIS 2013). In the first step, Kraków was divided into 389 1 km × 1 km squares from which the sample plots were drawn. The grid of squares was based on a point with coordinates 50°N 20°E. Then, every square was subdivided into four smaller squares of sides 500 m × 500 m (25 ha); one of these four was chosen at random for the counts. On every sample plot two transects each 500 m long were marked out. The ideal transect runs longitudinally or latitudinally, but obstacles in the terrain (existing buildings, fences, walls) caused the real transects to deviate from the ideal.

Study area: distribution of sample plots in Kraków (S Poland) and the main habitat types used for assessing variables that influence the distribution pattern of roe deer Capreolus capreolus in an urban environment
Roe deer were surveyed during winter in 2014 and 2015 and two surveys were carried out each year: early winter (03.01–30.01) and late winter (01.02–28.02). A period of at least two weeks had to elapse between consecutive surveys in a given year. Walking along the transects, the observer scanned a terrain through binocular (10 × 42) recording the presence of roe deer. To enhance the efficiency of detection, observers also searched for tracks leaved on snow/wet soil (if present), which indicates presence of a herd or an individual. The surveys were conducted during the daytime, though only on rain-free, snow-free and windless days (0–1 Beaufort’s scale), between 08:00 and 15:00 h CET. The transects were walked at an average speed of 2 km/h and single survey of a sample plot took around an hour. Each of 60 sample plots was surveyed four times (two survey in each season). All roe deer individuals observed within a sample plot were counted and assigned to one of the distance zones delineated on either side of the transect: (a) 0–100 m and (b) >100 m. Sample plots were defined as occupied by roe deer if at least single individual was recorded during one of the four surveys within a plot limits.
Environmental variables
The habitat parameters were defined on the basis of existing spatial database resources using GIS tools and the fieldwork. The surface areas (ha) of the largest wooded habitat patch (WOOD_PATCH) and the total area of open habitats (OPEN_HABITAT) within a sample plot limits were calculated using the polygon vector layer of the atlas of the real vegetation of Kraków (UMK 2012), which is the effect of fieldwork done in 2006 (Dubiel and Szwagrzyk 2008). The area of largest wooded habitat patch (WOOD_PATCH) included deciduous forest, mixed woodland, naturally growing shrubs, parks and cemeteries. Open habitats (OPEN_HABITAT) included arable land, meadows, pastures, uncultivated and fallow land, rock vegetation, swards, heaths and the communities of trampled areas. Distance to rivers (RIVERS) was calculated by measuring the distance from the sample plot to the nearest line of the layer of rivers located within the study area (WODGiK 2015). The number of buildings (BUILDINGS) was calculated using the polygon vector layer of buildings (WODGiK 2015), summing the number of objects located within a sample plot limits.
The noise emission variable (NOISE) was determined from the road noise emission map (MIIP 2016). The mean noise class weighted by its range area was calculated for every sample plot. Average noise levels during the hours of darkness (22:00–06:00 h) were used for these calculations. Noise level during the nighttime is highly correlated with noise level during the daytime (rS = 0.97, p = 0.000). The map shows the noise level expressed in 9 classes of sound intensity (dB) (1 – <45, 2 – 45-50, 3 – 50.1-55, 4 – 55.1-60, 5 – 60.1-65, 6 – 65.1-70, 7 – 70.1-75, 8 – 75.1-80, 9 – >80). The map was compiled jointly by the Provincial Environmental Conservation Inspectorate in Kraków and the Kraków City Council in 2012 (MIIP 2016).
Light pollution (LIGHT) was determined on the basis of the Visible Infrared Imaging Radiometer Suite (VIIRS) supplied by The Earth Observations Group (EOG 2016). The raster layer contained average radiance using night-time data from the VIIRS during January 2015 and was expressed in nanowatts per square centimetre per steradian (nW/cm2 × SR). The map’s resolution (grid pixel size) was ~460 m (EOG 2016). Sample plots applied in our study do not fit the grid cells of light pollution map, overlapping with up to nine cells of light pollution map. Therefore, the value of this variable was calculated as the average of 100 points located in the regular grid covering each of sample plots.
Information on numbers of dogs (DOGS) was obtained during the transect counts. During each field survey all dogs seen within a sample plot limits were counted (free ranging, within enclosed properties or on a leash), and higher value recorded for two surveys (early winter and late winter) was taken as a number of dogs in a given season. Then, we calculated the mean number of dogs based on the values obtained for both seasons (2014 and 2015).
Data handling and analysis
In the first step, mean (±SD) and median (with quartiles) values of each environmental variable were calculated for plots occupied and unoccupied by roe deer, the differences between the two groups being analysed using Mann-Whitney’s U test. Then factors determining the probability of roe deer occurring in an urban environment were investigated using a generalized linear model with binomial error distribution (Bolker et al. 2009). The area of open habitats (OPEN_HABITAT), the area of the largest wooded habitat patch (WOOD_PATCH), proximity to rivers (RIVERS), the number of dogs (DOGS), the number of buildings (BUILDINGS), noise emission (NOISE) and light pollution (LIGHT) were taken to be explanatory variables potentially explaining the presence of roe deer. To control for multicollinearity between variables, a correlation matrix was plotted (the correlation was <0.7 for all variable pairs; Table 1S, see Supplementary materials).
An information theory approach was used to identify and rank the best-supported model, and Akaike’s information criterion (AIC) was used for model selection (Burnham and Anderson 2002). Because of the relatively small sample size (n/K ratio <40), a small-sample version of AIC with bias adjustment (AICc) was applied in the modelling. The resulting models were subsequently ranked in order of increasing AICc. The differences between the model and the lowest AICc were calculated (ΔAICc) for each of the resulting models.
In order to illustrate directions, estimates (±SE) and 95% confidence intervals (CI) of environmental variables, generalized linear model with all variables included in the best-supported models (ΔAICc ≤5) were calculated. Then, logistic regression models (with plots occupied/unoccupied as a dichotomous dependent variable) were run for the variables indicated in the linear model as being of major importance for the probability of roe deer occurring in order to detect threshold values determining the species’ presence.
Finally, the percentage of deviance explained (100 × (deviancenull model – deviancemodel)/deviancenull model) was calculated for the three best supporting models (ΔAICc <5). Thereafter, deviance partitioning analysis (Borcard et al. 1992) was used to partition the deviance of each response variable included in the models into the independent effect of a particular predictor (pure effects) and the covarying effect of two or more predictors that cannot be disentangled (joint effects). This procedure allows one to calculate the percentage of the variation explained by each response variable and the joint effect of two or more variables not affected by the collinearity with other factor groups in the model (Legendre and Legendre 1998).
Since habitat characteristics could potentially affect the ability of researcher to detect roe deer, prior to analyses the effect of selected environmental variables on species detectability was tested. We assumed that increasing share of open habitats potentially increase detection distance due to higher visibility range, while increasing area of the largest wooded habitat patch and the number of buildings decrease detection distance due to reduced visibility. To check the influence of habitat characteristics on results of the surveys, we compared environmental variables between sample plots on which roe deer was detected in the different distance zones from the transect line. Based on the data combined from all surveys, sample plots were assigned into three groups: (a) plots on which roe deer was detected exclusively in the distance zone below 100 m, (b) plots on which roe deer was detected exclusively in the distance zone above 100 m, and (c) plots on which roe deer was detected in both zones. All analyzed environmental variables did not differ significantly between distinguished groups (OPEN_HABITAT: F2,19 = 0.05, p = 0.950; WOOD_PATCH: F2,19 = 0.31, p = 0.740; BUILDINGS: F2,19 = 0.79, p = 0.470), indicating that the distance in which animals were detected was not influenced by the habitat characteristics.
The statistical procedures were performed using Statistica 12 software (StatSoft Inc. 2014). Spatial autocorrelation of plots occupied by roe deer was assessed with Moran’s I tests (Rangel et al. 2010). We detected no evidence of spatial autocorrelation (Moran’s I was close to zero for all separation distances and semi-variance did not increase with lag distance).
Results
Roe deer were recorded on 36.7% of the randomly selected sample plots (N = 60). The median number of individuals recorded on a sample plot was 2.5 (quartile range 1–6, range 1–32) and the modal group size was 1. Occupied plots contained significantly more open habitats, proximity to rivers was significantly less there, and the area of the largest wooded habitat patch was greater (approaching significance level) (Table 1). The number of dogs did not differ significantly between occupied and unoccupied plots (Table 1). All variables that are attributes of urbanization differed significantly between plots: the number of buildings, the level of noise emission and light pollution were significantly higher on unoccupied than occupied plots (Table 1).
The best supported model (ΔAICc = 0.0) explaining the probability of roe deer occurring contained two variables: area of open habitats and light pollution (Table 2). Logistic regression models run separately for the two variables indicated in the best supported model revealed that the threshold values for a 0.5 occurrence probability were 8.8 ha of open habitats (corresponding to 35.2% of the plot area) and 10.3 nW/cm2 × SR of light pollution (Fig. 2). According to the logistic regression model (Table 3), a high occurrence probability (P ≥ 0.8) was recorded on both plots with high light pollution levels (~30 nW/cm2 × SR) but containing a large area of open habitats (~25 ha, corresponding to 100% of the plot area) and plots with a small area of open habitats (~5 ha, 20% of the area) but low light pollution levels (~0 nW/cm2 × SR) (Fig. 3). Two other, less-well supported models (ΔAICc = 0.4 and 3.6, see Table 2) indicated that the area of the largest wooded habitat patch was another potentially useful predictor for explaining the probability of roe deer occurring (Table 3).

Relationship between the area of open habitat (left; ha) and light pollution (right; nW/cm2 × SR) and the probability of roe deer Capreolus capreolus occurring in an urban environment (Kraków, S Poland; for variables, see Methods). The occurrence probability functions are: p = e(0.41y-3.62)/(1 + e(0.41y-3.62)) and p = e(−0.25x + 2.57)/(1 + e(−0.25x + 2.57)), where y and x represent the area of open habitat and light pollution, respectively. Grey shading indicates 95% confidence intervals

Logistic regression model predicting the probability of roe deer Capreolus capreolus occurring based on the area of open habitat (ha) and light pollution (nW/cm2 × SR) in an urban environment (Kraków, S Poland; for variables, see Methods). The occurrence probability function is p = e(0.37y-0.24x-0.61)/(1 + e(0.37y-0.24x-0.61)), where y and x represent the area of open habitat and light pollution, respectively
The best supported model (ΔAICc = 0.0) of the probability of roe deer occurring, including the area of open habitats and light pollution, explained 70.9% of the deviance in the dataset, a value that increased to 73.3% when the area of the largest wooded habitat patch was added. Deviance partitioning analysis showed that the area of open habitats had the greatest effect on the probability of roe deer occurring (Fig. 4), its pure effect accounting for 27% of the total deviance. The pure effect of light pollution was less important in explaining variability in occurrence probability (Fig. 4). However, the large deviance was explained by the joint effects of these variables: the probability of roe deer occurring increased with the area of open habitats not polluted by artificial light.

Deviance partitioning analysis for the probability of roe deer Capreolus capreolus occurring in an urban environment (Kraków, S Poland); the percentage of variability in the probability of occurrence is explained by the pure (A – area of open habitat, B – light pollution, C – largest wooded habitat patch) and joint effects of habitat variables
Discussion
This study has shown that the probability of roe deer occurring in an urban matrix is positively correlated with an increasing area of open habitats (grassland and arable land). Roe deer, originally a woodland species (Mattioli 2011), exhibits considerable flexibility where habitat choice is concerned: it occurs in both woodland and in open terrain, and also in the ecotone, i.e. the boundary zone in between (Hewison et al. 2001; Morellet et al. 2011; Lovari et al. 2017). In predominantly open habitats, roe deer tend to remain close to clumps of trees (Hewison et al. 2001; Morellet et al. 2011; Lovari et al. 2017), the presence of which determines whether this species will inhabit farmland or not (Acevedo et al. 2005). Where patches of woodland are lacking, roe deer will avail themselves of shrubs and hedgerows (Morellet et al. 2011), which provide shelter/concealment, particularly in thickly populated urban areas (Mysterud et al. 1999; Bonnot et al. 2013). Outside the growing season, wooded areas are probably less important to roe deer, because leafless trees and shrubs do not provide effective camouflage (Putman 1986; Mysterud et al. 1999; Hewison et al. 2009; Bonnot et al. 2013). Moreover, during spells of bad weather in winter, roe deer have greater energy requirements, so they may spend more time foraging in farmland, which is less secure but richer in food (Putman 1986; Mysterud et al. 1999). During this season, therefore, farmland may be a key habitat, supporting the viability of a population.
Artificial light is often used as a predictor for the occurrence/density of animals, particularly birds and bats (Azam et al. 2016; Ciach and Fröhlich 2017; Froidevaux et al. 2017). Because they are frequently active at night, ungulates often enter areas that are affected by artificial lighting (Hewison et al. 2009; Pagon et al. 2013), so their populations are potentially affected by it (Beier 2006; Hölker et al. 2010). To the best of our knowledge, however, the present study is the first one to assess the correlation between artificial lighting at night and the distribution of ungulate population. The occurrence of roe deer was less probable in areas with high intensities of artificial lighting: this parameter explained the occurrence of this species better than the number of buildings or noise levels. Although the mechanism of avoiding artificial light has not been well documented, experimental modifications of the photoperiod have shown that artificial light can lead to hormonal changes in ungulates that alter their reproductive cycles as well as moult strategies for the maintenance of suitable body temperatures (Yeates 1949; Lincoln and Guinness 1972). The adverse effects of light pollution have also been demonstrated in other mammals: the extended photoperiod in urban areas lit up at night led to changes in their diurnal activity, thereby significantly affecting their time budget (Foster and Provencio 1999; Biebouw and Blumstein 2003; Barber-Meyer 2007) or causing them to avoid areas affected by artificial light (Robert et al. 2015). Our work has shown that light pollution could be used as an additional predictor of ungulate occurrence: there is therefore an urgent need for further studies of the influence of artificial light on the ecology of this group of animals.
The logistic regression model presented in this paper indicates that the probability of roe deer occurring on farmland not affected by artificial lighting is increasing (Fig. 5). Although the drop in quality of agricultural habitats exposed to artificial lighting has been confirmed with respect to some other species (De Molenaar and Sanders 2006; Bennie et al. 2017; Froidevaux et al. 2017), knowledge of this subject with regard to large mammals is still lacking. Since farmland is intensively utilized by humans during the growing season, ungulates tend to avoid this habitat at that time, but do make use of it in winter, when woodlands do not supply sufficient food resources (Putman 1986; Hewison et al. 2009). In the vicinity of human habitations, however, ungulates are mostly active at night, which is probably a strategy helping to avoid contact with humans (Mysterud et al. 1999; Bonnot et al. 2013; Pagon et al. 2013). Animals may equate the presence of artificial lighting in farming habitats with the presence of humans: perceiving this as a threat, they will tend to avoid illuminated areas (De Molenaar and Sanders 2006; Robert et al. 2015).

Examples of sample plots located within an urban environment illustrating potential habitats of roe deer Capreolus capreolus in an urban environment (Kraków, S Poland); plot selection based on threshold values for 0.5 occurrence probability – 8.8 ha for open habitats (corresponding to 35.2% of the plot area) and 10.3 nW/cm2 × SR for light pollution; satellite images taken from Google Earth, supplied by Digital Globe – the respective coordinates of the centres of the images are: 50°3′57.72″N 19°52′19.08″E, 50°4′30.54″N 19°51′53.58″E, 50°2′36.94″N 19°53′31.84″E, 50°1′47.93″N 19°51′51.01″E; eye altitudes 1.1 km
The results of our study indicate that there was a lower intensity of noise in the areas colonized by roe deer, although this was not a key factor in the population distribution model. Noise, the main source of which in urban areas is road traffic, interferes with the propagation of sound waves and seriously affects the distribution of animal populations in which individuals communicate vocally with one another (Nemeth et al. 2013; Ciach and Fröhlich 2017) and foraging (Siemers and Schaub 2011; Mason et al. 2016) relies on acoustic signals. However, the results of studies conducted on small mammals indicate that traffic noise is not considered a factor leading to avoidance of roads (McGregor and Bender 2008). Roads and the traffic they carry are more likely to bar the movements of terrestrial mammals (Forman and Alexander 1998; Seidler et al. 2015), which, moreover, often fall victim to collisions with vehicles (Bruinderink and Hazebroek 1996; Zuberogoitia et al. 2014). The presence of roads does also mean, however, that ungulates limit their activities near them during periods of heavy traffic (Kušta et al. 2017). This mechanism provides evidence that ungulates have to some extent adapted to inhabiting areas with a dense road network (McCarthy et al. 1996; Acevedo et al. 2005; Underwood and Kilheffer 2016; Loro et al. 2016; Kušta et al. 2017), and that noise in itself is probably not the most important factor governing the distribution of their populations (Figs. 5 and 6).

Winter habitats of roe deer Capreolus capreolus in an urban environment (Kraków, S Poland)
Our study has shown that the areas colonized by roe deer lie close to rivers. Rivers and their associated riparian forests act as migration corridors (Romanowski 2007; Rodriguez-Iturbe et al. 2009). Despite the widespread and far-reaching transformation of riparian environments, watercourses in highly urbanized environments are often the only corridors along which animals can penetrate them (May 2006; Ignatieva et al. 2011). Maintaining urban watercourses in as natural a state as possible will therefore probably favour the colonization of towns and cities by large mammals.
This research has also shown that the number of dogs was unimportant as a factor affecting the probability of roe deer occurring. The dog is most closely related to the wolf, the natural predator of medium-sized and large ungulates (Mattioli et al. 2004). The barking of dogs scares off ungulates, which can affect the time budget of the latter and increase their energy expenditure (Reimoser 2012; Padié et al. 2015). Most dogs in urban areas are kept indoors or within enclosed properties, while those moving about the city are on a leash. Since there are no feral and/or free-ranging dogs, ungulates to some extent simply ignore the presence of these animals (Padié et al. 2015); again, this is probably another adaptation to living near human habitations, where pet dogs are ubiquitous (McCarthy et al. 1996; Kilpatrick and Spohr 2000; Loro et al. 2016).
We have shown that the probability of roe deer occurring in urban environments is correlated with the presence of open habitats and light pollution. To date, studies of the influence of urbanization on ungulates analysed only the area built up or the distance to buildings (Hewison et al. 2001; Torres et al. 2011; Underwood and Kilheffer 2016; Loro et al. 2016). This paper is the first to demonstrate that light pollution can substantially affect the probability of roe deer occurring; consequently, artificial lighting is likely to be an additional factor discouraging ungulates from colonizing potentially suitable habitats in urban areas. The roe deer is one of the commonest grazing animals in the temperate zone and its presence/absence may therefore affect the vegetation structure. This strongly suggests, in turn, an indirect effect of light pollution on environment, where changes in species distributions can give rise to changes in the habitats that are utilized or avoided. Our results demonstrate that light pollution may be an urban effect shaping the spatial distribution of large mammals.
References
Acevedo P, Delibes-Mateos M, Escudero MA et al (2005) Environmental constraints in the colonization sequence of roe deer (Capreolus capreolus Linnaeus, 1758) across the Iberian Mountains, Spain. J Biogeogr 32:1671–1680
Acevedo P, Farfán MÁ, Márquez AL et al (2011) Past, present and future of wild ungulates in relation to changes in land use. Landsc Ecol 26:19–31
Arnfield AJ (2003) Two decades of urban climate research: a review of turbulence, exchanges of energy and water, and the urban heat island. Int J Climatol 23:1–26
Azam C, Le Viol I, Julien J-F et al (2016) Disentangling the relative effect of light pollution, impervious surfaces and intensive agriculture on bat activity with a national-scale monitoring program. Landsc Ecol 31:2471–2483
Barber-Meyer SM (2007) Photopollution impacts on the nocturnal behaviour of the sugar glider (Petaurus breviceps). Pacific Conserv Biol 13:171–176
Bateman PW, Fleming PA (2012) Big city life: carnivores in urban environments. J Zool 287:1–23
Beier P (2006) Effects of artificial night lighting on terrestrial mammals. In: Rich C, Longcore T (eds) Ecological consequences of artificial night lighting. Island press, Washington. Covelo, London, pp 19–42
Bennie J, Davies TW, Cruse D et al (2017) Artificial light at night alters grassland vegetation species composition and phenology. J Appl Ecol. https://doi.org/10.1111/1365-2664.12927
Biebouw K, Blumstein DT (2003) Tammar wallabies ( Macropus eugenii ) associate safety with higher levels of nocturnal illumination. Ethol Ecol Evol 15:159–172
Bolker BM, Brooks ME, Clark CJ et al (2009) Generalized linear mixed models: a practical guide for ecology and evolution. Trends Ecol Evol 24:127–135
Bonnot N, Morellet N, Verheyden H (2013) Habitat use under predation risk: hunting, roads and human dwellings influence the spatial behaviour of roe deer. Eur J Wildl Res 59:185–193
Boone RB, Hobbs NT (2004) Lines around fragments: effects of fencing on large herbivores. African J Range Forage Sci 21:147–158
Borcard D, Legendre P, Drapeau P (1992) Partialling out the spatial component of ecological variation. Ecology 73:1045–1055
Brown T, Decker D, Riley S, Enck J (2000) The future of hunting as a mechanism to control white-tailed deer populations. Wildl Soc 28:797–807
Bruinderink GWTAG, Hazebroek E (1996) Ungulate traffic collisions in Europe. Conserv Biol 10:1059–1067
Burnham KP, Anderson DR (2002) Model selection and multi-model inference: a practical-theoretical approach. Springer, New York
Ciach M, Fröhlich A (2017) Habitat type, food resources, noise and light pollution explain the species composition, abundance and stability of a winter bird assemblage in an urban environment. Urban Ecosyst 20:547–559
De Molenaar J, Sanders M (2006) Road lighting and grassland birds: local influence of road lighting on a black-tailed godwit population. In: Rich C, Longcore T (eds) Ecological Consequences of Artificial Night Lighting. Island Press, Washington D.C., pp 114–136
Dubiel E, Szwagrzyk J (2008) Atlas Roślinności Rzeczywistej Krakowa. In: Urząd Miasta Krakowa. Wydział Kształtowania Środowiska, Kraków
EOG (The Earth Observations Group) (2016) Nighttime VIIRS Day/Night Band Composites. http://ngdc.noaa.gov/eog/viirs/download_monthly.html. Accessed 28 Mar 2015
Ewald M, Dupke C, Heurich M et al (2014) LiDAR remote sensing of Forest structure and GPS telemetry data provide insights on winter habitat selection of European roe deer. Forests 5:1374–1390
Falarz M (1998) Long-term variability of snow cover in Cracow and suburban areas. Folia Geogr Phys 3:473–481
Forman RTT, Alexander LE (1998) Roadfs and their major ecological effects. Annu Rev Ecol Syst 29:207–231
Foster RG, Provencio I (1999) The regulation of vertebrate biological clocks by light. In: Djamgoz MBA, Loew ER et al (eds) Archer SN. Adaptive Mechanisms in the Ecology of Vision. Springer, Dordrecht, pp 223–243
Found R, Boyce MS (2011) Predicting deer–vehicle collisions in an urban area. J Environ Manag 92:2486–2493
Froidevaux JSP, Boughey KL, Barlow KE, Jones G (2017) Factors driving population recovery of the greater horseshoe bat (Rhinolophus ferrumequinum) in the UK: implications for conservation. Biodivers Conserv 26:1601–1621
GUS (Główny Urząd Statystyczny) (2016) Powierzchnia i ludność w przekroju terytorialnym w 2015 r. Główny Urząd Statystyczny, Warszawa
Harrington JL, Conover MR (2006) Characteristics of ungulate behavior and mortality associated with wire fences. Wildl Soc Bull 34:1295–1305
Hewison AM, Vincent JP, Joachim J et al (2001) The effects of woodland fragmentation and human activity on roe deer distribution in agricultural landscapes. Can J Zool 79:679–689
Hewison AJM, Morellet N, Verheyden H et al (2009) Landscape fragmentation influences winter body mass of roe deer. Ecography 32:1062–1070
Hölker F, Wolter C, Perkin EK, Tockner K (2010) Light pollution as a biodiversity threat. Trends Ecol Evol 25:681–682
Ignatieva M, Stewart GH, Meurk C (2011) Planning and design of ecological networks in urban areas. Landsc Ecol Eng 7:17–25
Kilpatrick H, Spohr S (2000) Spatial and temporal use of a suburban landscape by female white-tailed deer. Wildl Soc Bull 28:1023–1029
Klett-Mingo IJ, Pavón I, Gil D (2016) Great tits, Parus major, increase vigilance time and reduce feeding effort during peaks of aircraft noise. Anim Behav 115:29–34
Kušta T, Keken Z, Ježek M et al (2017) The effect of traffic intensity and animal activity on probability of ungulate-vehicle collisions in the Czech Republic. Saf Sci 91:105–113
Lee M, Miller R (2003) Managing elk in the wildland-urban interface: attitudes of Flagstaff. Arizona residents Wildl Soc Bull 31:185–191
Legendre P, Legendre L (1998) Numerical ecology. 3rd Edn. Elsevier publishers, Amsterdam, Holland
Lincoln GA, Guinness FE (1972) Effect of altered photoperiod on delayed implantation and moulting in roe deer. J Reprod Fertil 31:455–457
Longcore T, Rich C (2004) Ecological light pollution. Front Ecol Environ 2:191–198
Loro M, Ortega E, Arce RM, Geneletti D (2016) Assessing landscape resistance to roe deer dispersal using fuzzy set theory and multicriteria analysis: a case study in Central Spain. Landsc Ecol Eng 12:41–60
Lovari S, Serrao G, Mori E (2017) Woodland features determining home range size of roe deer. Behav Process 140:115–120
Mason TJ, McClure CJW, Barber JR (2016) Anthropogenic noise impairs owl hunting behavior. Biol Conserv 199:29–32
Mattila M, Hadjigeorgiou I (2015) Conservation and management of fallow deer (Dama dama dama L.) on Lemnos Island, Greece. Turkish J Vet Anim Sci 39:560–567
Mattioli S (2011) Family Cervidae (deer). In: Wilson DE, Mittermeier RA (eds) Handbook of the mammals of the world, vol 2. Hoofed Mammals. Lynx edicions, Barcelona, pp 350–443
Mattioli L, Capitani C, Avanzinelli E et al (2004) Predation by wolves (Canis lupus) on roe deer (Capreolus capreolus) in north-eastern Apennine, Italy. J Zool 264:249–258
May R (2006) “Connectivity” in urban rivers: conflict and convergence between ecology and design. Technol Soc 28:477–488
McCarthy A, Baker A, Rotherham I (1996) Urban-fringe deer management issues-a South Yorkshire case study. Br Wildl 8:12–19
Mcdonald RI, Kareiva P, Forman RTT (2008) The implications of current and future urbanization for global protected areas and biodiversity conservation. Biol Conserv 141:1695–1703
McGregor R, Bender D (2008) Do small mammals avoid roads because of the traffic? J Appl Ecol 45:117–123
MIIP (Małopolska Infrastruktura Informacji Przestrzennej) (2016) Geoportal MIIP. http://miip.geomalopolska.pl/imap/. Accessed 28 Mar 2016
Molinari-Jobin A, Molinari P, Breitenmoser-Würsten C, Breitenmoser Molinari-Jobin U (2002) Significance of lynx Lynx lynx predation for roe deer Capreolus capreolus and chamois Rupicapra rupicapra mortality in the Swiss Jura Mountains. Wildlife Biol 8:109–115
Morellet N, Van Moorter B, Cargnelutti B et al (2011) Landscape composition influences roe deer habitat selection at both home range and landscape scales. Landsc Ecol 26:999–1010
Mysterud A, Lian L-B, Hjermann DØ (1999) Scale-dependent trade-offs in foraging by European roe deer ( Capreolus capreolus ) during winter. Can J Zool 77:1486–1493
Nemeth E, Pieretti N, Zollinger SA et al (2013) Bird song and anthropogenic noise: vocal constraints may explain why birds sing higher-frequency songs in cities. Proc R Soc B Biol Sci 280:2012–2798
Padié S, Morellet N, Cargnelutti B et al (2015) Time to leave? Immediate response of roe deer to experimental disturbances using playbacks. Eur J Wildl Res 61:871–879
Pagon N, Grignolio S, Pipia A et al (2013) Seasonal variation of activity patterns in roe deer in a temperate forested area. Chronobiol Int 30:772–785
Putman RJ (1986) Foraging by roe deer in agricultural areas and impact on arable crops. J Appl Ecol 23:91–99
Putman RJ, Moore NP (1998) Impact of deer in lowland Britain on agriculture, forestry and conservation habitats. Mamm Rev 28:141–164
QGIS (Quantum GIS Development Team) (2013) Quantum GIS geographic information system. In: Open source Geospatial Foundation project
Rangel TF, Diniz-Filho JAF, Bini LM (2010) SAM: a comprehensive application for spatial analysis in macroecology. Ecography 33:46–50
Reimoser S (2012) Influence of anthropogenic disturbances on activity, behavior and heart rate of roe deer (Capreolus capreolus) and Red Deer (Cervus elaphus), in context of their daily and yearly patterns. In: Cahler AA, Marsten JP (eds) Deer: Habitat. Behavioral and Conservation. Nova Science Publisher, New York, pp 1–95
Robert KA, Lesku JA, Partecke J, Chambers B (2015) Artificial light at night desynchronizes strictly seasonal reproduction in a wild mammal. Proc Royal Soc B 282. https://doi.org/10.1098/rspb.2015.1745
Rodriguez-Iturbe I, Muneepeerakul R, Bertuzzo E et al (2009) River networks as ecological corridors: a complex systems perspective for integrating hydrologic, geomorphologic, and ecologic dynamics. Water Resour Res. https://doi.org/10.1029/2008WR007124
Romanowski J (2007) Vistula river valley as the ecological corridor for mammals. Polish J Ecol 55:805–819
Seidler RG, Long RA, Berger J et al (2015) Identifying impediments to long-distance mammal migrations. Conserv Biol 29:99–109
Siemers BM, Schaub A (2011) Hunting at the highway: traffic noise reduces foraging efficiency in acoustic predators. Proc Biol Sci 278:1646–1652
StatSoft Inc. (2014) Statistica (data analysis software system), version 10
Torres RT, Carvalho JC, Panzacchi M et al (2011) Comparative use of forest habitats by roe deer and moose in a human-modified landscape in southeastern Norway during winter. Ecol Res 26:781–789
UMK (Urząd Miasta Krakowa) (2012) Mapa roślinności rzeczywistej miasta Krakowa. Urząd Miasta Krakowa, Kraków
Underwood H, Kilheffer CR (2016) Modeling the effects of land cover and use on landscape capability for urban ungulate populations. In: Francis RA, Millington JDA, Chadwick MA (eds) Urban landscape ecology: science. Policy and Practice. Routledge, London and New York, pp 181–208
Warren R (2011) Deer overabundance in the USA: recent advances in population control. Anim Prod Sci 51:259–266
Welch D, Staines BW, Scott D et al (1991) Leader browsing by red and roe deer on young Sitka spruce trees in Western Scotland I. Damage rates and the influence of habitat factors. Forestry 64:61–82
WODGiK (Wojewódzki Ośrodek Dokumentacji Geodezyjnej i Kartograficznej) (2015) Baza danych obiektów topograficznych. http://www.geomalopolska.pl/. Accessed 28 Mar 2015
Yeates N (1949) The breeding season of the sheep with particular reference to its modification by artificial means using light. J Agric Sci 39:1–43
Zuberogoitia I, del Real J, Torres JJ et al (2014) Ungulate vehicle collisions in a Peri-urban environment: consequences of transportation infrastructures planned assuming the absence of ungulates. PLoS One 9:e107713
Acknowledgements
We wish to express our gratitude to members of the Foresters’ Scientific Club of the Faculty of Forestry in Kraków for their help with the fieldwork. Financial support for this study was provided by the Polish Ministry of Science and Higher Education by statutory grant (DS 3421).
Author contributions statement
AF conceived the idea and coordinated the collection of the data, AF and MC designed the study and participated in the fieldworks; MC analyzed the data; MC and AF wrote the manuscript.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethical approval
All applicable institutional and national guidelines for the care and use of animals were followed.
Electronic supplementary material
ESM 1
(DOCX 59 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Ciach, M., Fröhlich, A. Ungulates in the city: light pollution and open habitats predict the probability of roe deer occurring in an urban environment. Urban Ecosyst 22, 513–523 (2019). https://doi.org/10.1007/s11252-019-00840-2
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11252-019-00840-2