
Abstract
Green roofs are potentially valuable habitats for plants and animals in urban areas. Wild bees are important pollinators for crops and wild plants and may be enhanced by anthropogenic structures, but little is known about wild bees on green roofs in cities. This study investigates the effects of green roof qualities (floral resources, substrate character and depth, roof height and age) on wild bee diversity, abundance and traits (nesting type, sociality, pollen specialisation, body size) on green roofs in Vienna. Nine green roofs were sampled monthly between March and September 2014 by a semi quantitative approach. Wild bees were collected in pre-defined sub-areas for the same amount of time and floral resources were recorded. Over all green roofs, 992 individuals belonging to 90 wild bee species were observed. Wild bee diversity and abundance was strongly positively affected by increasing forage availability and fine substrates. Wild bees on roofs were characteristically solitary, polylectic and 8.3–11.2 mm. Regarding nesting type, the percentage of above-ground nesting bees was higher compared to the common species composition in Middle Europe. Ground-nesting wild bees were mainly eusocial, smaller (6.4–9.6 mm) and positively affected by roofs with fine substrates. During June, when forage availability by wildflowers on roofs was “low” (5–15% flower coverage), flowering Sedum species were an important forage resource. We conclude that wild bee diversity and abundance on green roofs are enhanced by floral resources. Furthermore, the installations of areas with finer and deeper substrates benefit ground nesting and eusocial wild bees.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The degradation and loss of habitats are seriously threatening wild bees (Winfree et al. 2009; Le Féon et al. 2010; Potts et al. 2010; Kennedy et al. 2013), leading to declines in pollination services for crops (Kremen et al. 2002; Klein et al. 2007) and wild plants (Biesmeijer et al. 2006; Fontaine et al. 2006). As important pollinators in gardens, parks and other green spaces (Matteson et al. 2008; Frankie et al. 2009; Matteson and Langellotto 2010), wild bees play a crucial role for urban ecosystems. The availability of pollen and nectar as well as nesting sites within species-specific flying distances is important for wild bees to colonize habitats successfully (Westrich 1996; Gathmann and Tscharntke 2002; Zurbuchen et al. 2010; Wojcik and McBride 2012). Bees are ectothermic and thermophile organisms, thus the warmer (micro-) climate of cities enables favourable habitat conditions for many wild bee species (Cane 2005; Hennig and Ghazoul 2012). Research on urban sites likely to be populated by wild bees, provides important information for urban planning to enhance wild bee diversity (Hernandez et al. 2009).
Sustainable city planning considers green roofs as a vital measure to compensate negative effects of sealed surfaces. Many abiotic benefits have been reported (reviewed in Oberndorfer et al. 2007) like the improvement of city’s water run off management (Mentens et al. 2006; Hathaway et al. 2008; Berndtsson 2010), the reduction of heat islands (Susca et al. 2011) and the increase of energy efficiency by insulating the building’s indoor rooms against heat and cold (Castleton et al. 2010; Zinzi and Agnoli 2012). Further benefits are aesthetic and amenity for urban citizens (van den Berg et al. 2007). From an ecological point of view, green roofs have high potential to restore habitats for plants and animals in cities (Oberndorfer et al. 2007; Carter and Butler 2008; Dunnett and Kingsbury 2008). During the last decade, studies on plants (Landolt 2001; Köhler 2005), birds (Gedge 2003; Baumann 2006; Fernandez-Canero and Gonzalez-Redondo 2010) and various invertebrate taxa like Araneae (Brenneisen 2003; Kadas 2006), Collembola (Schrader and Böning 2006), Coleoptera (Kadas 2006) and Hymenoptera (Brenneisen 2003; Brenneisen 2005; Kadas 2006; Colla et al. 2009; MacIvor and Lundholm 2010; Tonietto et al. 2011; Ksiazek et al. 2014; MacIvor et al. 2014) underpinned the importance of green roofs to contribute to biodiversity and nature conservation in cities.
Green roofs are potentially important habitats for urban wild bees, as they provide pollen and nectar resources throughout the year (Tonietto et al. 2011) and incorporate different nesting habitats (Brenneisen 2005; MacIvor et al. 2014). In addition, the higher insulation and thus warmer micro-climate at roof level result in favourable habitat conditions (Matteson and Langellotto 2010). Beside, a continuous vegetation layer on green roofs is crucial to the abiotic benefits discussed above and linked to the pollination by wild bees (Dunnett and Kingsbury 2008).
Similar wild bee communities have been reported on green roofs and ground level habitats in Toronto (Colla et al. 2009) and Chicago (Tonietto et al. 2011) but higher wild bee diversity on ground sites was related to higher entomophilous plant diversity. The habitat quality of green roofs for wild bees was highlighted by observations of locally and/or nationally rare species (Kadas 2006) and of wild bee species newly recorded on green roofs (Ksiazek et al. 2014; MacIvor et al. 2014). Green roof types support wild bee diversity differently because of the type of substrate and its character and plant diversity (Brenneisen 2005). Furthermore, Sedum species are characteristic green roof plants and play an important role as a foraging resource for wild bees (MacIvor et al. 2014).
Whilst there has been research in Western Europe on green roofs and wild bees, knowledge about wild bees on green roofs in the eastern part of Europe, characterized by a warmer and dryer climate and therefore comprising a different wild bee species composition is scarce. In this paper, we report the wild bee diversity, abundance and traits in relation to green roof qualities on green roofs in Vienna. The main focus of this study was to determine:
-
(1)
The effect of temporal variable floral resources (forage availability and entomophilous plant diversity) and constant green roof qualities (substrate characteristics and depths, roof heights and ages) on wild bee communities.
-
(2)
Characteristic wild bee traits (pollen specialization, nesting type, sociality and body size) on green roofs and how these traits are affected by respective green roof qualities.
-
(3)
The importance of Sedum species compared to other wildflowers as foraging resource on green roofs.
Methods
Study sites
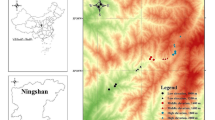
We studied wild bees on nine green roofs in Vienna (Fig. 1) during the vegetation period of 2014. In order to evaluate the wild bee diversity and abundance on roofs with different qualities (i.e. substrate characteristics and depths, roof height and age), the roofs varied in greening type (extensive, semi-extensive and intensive) leading to different planting possibilities (Dunnett and Kingsbury 2008) and floral resources (e.g. Sedum species and other wildflowers).

Locations of the nine green roofs in Vienna surveyed in 2014 (basic map: ViennaGIS 2016)
Detailed information about the roof’s qualities (Table 1) was gathered during wild bee sampling or by interviewing the people responsible for the buildings. Substrate characteristics were determined in the field. Coarse substrates consisted of high amounts of expanded clay and crashed brick and are typically used on extensive green roofs. Fine substrates consisted of a high amount of humus, earth and sand and are mainly used on intensive green roofs. Mixed substrate was present on roofs where coarse and fine substrates were used in different areas. Substrate depths, height and age were determined from planning documents or by interviewing people responsible for the building.
Vienna is characterised by a temperate Pannonian climate, with the potential for precipitation throughout the year (Auer and Böhm 2011). The average annual precipitation in 2014 was 756 mm and the average monthly temperature was 12.5 °C. The highest average daily temperature was recorded during July and early August and ranged between 21 and 23 °C (ZAMG - Zentralanstalt für Meterologie und Geodynamik 2015).
Wild bee sampling
Wild bee sampling was conducted by a semi-quantitative method, hand netting wild bees in each of the pre-defined sub-areas (Table 1) for the same period of time. The sub-areas were distinct patches characterized by dominating plants or vegetation types (e.g. wildflowers, Sedum, lawn, shrubs, Pannonian plants), structural characteristics (e.g. bare substrate, dead wood elements) or functionality (e.g. BBQ spot, unmaintained area). To assess the value of Sedum as foraging resource for wild bees, it was defined as a distinct sub-area. Patches with spontaneous flora and herbaceous garden plants, except for Sedum, were classified as “Wildflowers”. Sampling time was adapted to the size of the sub-areas and ranged between 3 min for small objects like dead wood elements and 15 min for large areas like lawns, shrubs, Sedum or wildflowers. Each roof was sampled monthly, seven times from March to September 2014 (Schindler et al. 2013). Sampling was conducted between 10 am and 4 pm on days with warm (22.8 ± 4.9 °C), windless and dry weather conditions. On each sampling date, information on floral resources was carried out. The forage availability (=flower coverage) of entomophilous plants, was assessed in each sub-area (Table 1) using five categories: 1 = “very low” (< 5%), 2 = “low” (5–15%), 3 = “medium” (15–25%), 4 = “high” (25–50%) and 5 = “very high” (> 50%). To assess the number of entomophilous plant species flowering at the time of sampling, plants were photographed and identified to species level. This was separately documented for each sub-area. On average (± SD), the roofs comprised “very low” to “medium” forage availability (2 ± 0.6) and 46.8 (± 39) entomophilous plant species. Evidence of nesting activity was gathered qualitatively during wild bee sampling. Bare substrate patches were observed for ground nesting activity and old plant stems, cavities in walls or dead wood branches for above-ground nesting activity. Wild bees were identified to species level (Ebmer 1969, 1970, 1971; Dathe 1980; Mauss 1994; Schmid-Egger and Scheuchl 1997; Amiet et al. 1999, 2001; Scheuchl 2000, 2006; Gokcezade et al. 2010) by the authors (SK, BP) and Karl Mazzucco was consulted for validation of some specimens mainly of the genus Lasioglossum and Hylaeus. All specimens are housed in the collection of the Institute for Integrative Nature Conservation Research at BOKU Vienna.
Wild bee traits (Table 2) were summarized by categories and determined by literature research (Westrich 1989a; Michener 2007; Scheuchl and Willner 2016). Information on body size was derived from identification literature (see above; bumblebees: von Hagen and Aichhorn 2003) which give the range of body size within a species. For trait analysis the average body size was calculated from the female and male values. The nesting type of Hylaeus imparilis remained unclear, but was attributed to the above-ground nesting group, like all other Hylaeus species (Falk 2015; Scheuchl and Willner 2016).
Data analyses
We excluded Apis mellifera from analysis, because it is a domestic species and the abundance could be biased by nearby hives (Kennedy et al. 2013). We only found a weak, insignificant positive relation between honey bee and wild bee abundance on the roofs (Spearman’s rank correlation; rho = 0.08; p = 0.8). Roof size was neither related to wild bee species richness (rho = −0.12; p = 0.7) nor to wild bee abundance (rho = −0.23; p = 0.5). Statistical analyses were performed in R 3.3.2 (R Core Team 2016) using R Studio V 0.99.903 (RStudio Team 2015).
The effects of temporal variable floral resources on wild bee species richness and abundance were analysed by generalized linear mixed models (GLMMs) with Poisson error distribution using the R-packages “lme4” (Bates et al. 2015) and “Matrix” (Bates and Maechler 2016). We formulated null and candidate models for each response variable with two random factors (“month” and “roof”) and either forage availability or entomophilous plant species richness as fixed factor. We did not formulate models containing both fixed factors because prior data exploration revealed them as collinear (cor = 0.6), which results in unreliable parameter estimation (Zuur et al. 2010). For GLMMs with forage availability as fixed factor, we used the category “very low” as baseline for parameter estimation. Model selection was carried out by the second order Akaike Information Criterion (AICc) using the R package “AICcmodavg” (Mazerolle 2016). The AICc is used for modelling data with small sample size (Motulsky and Christopoulos 2003). The cut-off to decide whether a model is more likely to be correct than the next one was set at ∆AICc < 2. We did not use p-values for interpretation because they are not reliable in GLMMs (Zuur et al. 2013).
The effects of uniform green roof qualities (substrate characteristics and depths, roof height, age) on wild bee species richness and abundance were fitted with Poisson generalized linear models (GLMs). For GLMs the wild bee species numbers and abundance were aggregated to total amounts per roof. Substrate depths were averaged if a range was indicated (Table 1) and the factor level “coarse” was the baseline for parameter estimation of substrate characteristics.
Characteristic traits on green roofs were examined by calculating community weighted means (CWM) with the R-package “FD” (Laliberté et al. 2014). To analyse how traits responded to green roof qualities, aggregated species numbers and abundance per trait (nesting type, sociality and pollen specialization) per roof were used as response variables in Poisson GLMs. The CWM of body size was used as response in GLMs with Gaussian error distribution. To reduce the number of GLMs for trait analysis, we chose the most interesting green roof qualities as explanatory variables for each trait (Table 2) based on our expertise. Because almost all explanatory variables were collinear (cor. ranged from 0.3 to 0.8), we only formulated GLMs with one explanatory variable. Model selection was done on AICc bases as described above. The explained deviance (R2GLM) was calculated for the most accurate GLMs to assess how much variation of the response is explained by the explanatory variable (Zuur et al. 2013).
Model validation of GLMMs and GLMs was performed by diagnostic plots and dispersion values (Zuur et al. 2013). The package “effects” (Fox 2003) was used to visualize the effects of the most accurate models.
To assess whether Sedum and other wildflowers provided crucial floral resources, a data sub-set of the sub-areas “Sedum” and “Wildflowers” from five roofs (Table 1) was analysed. The other green roofs had to be omitted from this analysis because they did not include both sub-areas. Due to small sample size this was done descriptively by comparing the aggregated wild bee species richness, abundance and averaged forage availability along a timeline plot created with Microsoft Excel 2010.
Results
Excluding A. mellifera (1470 individuals), which was present on all roofs, we identified 90 wild bee species belonging to 19 genera. A total of 992 wild bee individuals were caught on the green roofs (see Table 6 in Appendix for species list). The most abundant species was Halictus subauratus with 94 individuals (9.5%), whereas 25 wild bee species (27.8%) were represented by only one individual.
On average (± SD), green roofs hosted 29 (± 16.2) wild bee species and 110 (± 95.3) individuals. The most diverse roof with 65 wild bee species (337 individuals) contrasted with a roof that had only six species and six individuals.
Green roof qualities
Wild bee species richness and abundance was best predicted by forage availability (Table 3).
Forage availability had a strong positive effect on wild bee species richness and abundance (Fig. 2a, b; Table 7 in Appendix). Species richness and abundance increased from “very low” to “low” and further to “medium” forage availability, but from “medium” to “high” the positive effect was minimal.

Effects of forage availability on (a) wild bee species and (b) abundance per monthly collection period (n = 7) on each roof (n = 9). “Very low” forage availability was used as baseline for parameter estimation. Error bars = 0.95 confidence intervals for fitted effects
The models with substrate characteristics as an explanatory variable were the most accurate for predicting wild bee species richness and abundance in relation to uniform green roof qualities (Table 4).
Roofs with fine substrates represented the highest mean (± SD) wild bee diversity (47.5 ± 24.7) and abundance (232.5 ± 147.8) and affected wild bee species richness (Fig. 3a) and abundance (Fig. 3b) distinctively positive compared to roofs with mixed or coarse substrates. Roofs with mixed substrates increased moderately and represented a higher mean (± SD) wild bee diversity (26 ± 9.2) and abundance (89.3 ± 42.8) compared to roofs with coarse substrates, which comprised the lowest mean (± SD) wild bee species richness (21.5 ± 11.3) and abundance (64.7 + 50.8).

Effects of substrate character on (a) wild bee species and (b) abundance. The factor level “coarse” was used as baseline for parameter estimation. Error bars = 0.95 confidence intervals for fitted effects
On average, floral resources were highest on roofs with fine substrates (forage availability: “medium”, flowering plant species: 87 ± 69.3), less on roofs with mixed substrates (forage availability: “low”, flowering plant species: 55.3 ± 20.6) and lowest on roofs with coarse substrate (forage availability: “very low” to “low”, flowering plant species: 20.3 ± 9.3).
Models including substrate depth, roof height and age as explanatory variables were less accurate in predicting the total wild bee species richness and abundance. However, substrate characteristics and depths were highly positive collinear (cor = 0.8) and it should be mentioned here, that roofs with deep substrates affected wild bees positively too (Fig. 5a, b in Appendix).
Wild bee traits on roofs
A majority of 81 polylectic wild bee species (97% individuals) contrasts with a minority of ten oligolectic species (3.4% individuals). As reflected by the CWM (Table 5), the wild bee community was composed of ground nesting (43.3% species; 43.3% individuals) and above-ground nesting wild bees (43.3% species; 53.5% individuals). Solitary species (62.2%) accounted for 54.7% of the individuals and eusocial species (24.4%) for 42.4% of the individuals. Parasitic species (13.3%) were represented by 3.1% of the total abundance and not included in the CWM results (Table 5). The CWM of body size from wild bees on the studied roofs ranged between 8.3 and 11.2 mm. Individuals of ground nesting species ranged in CWM of body size between 6.4 and 9.6 mm and were mainly eusocial (68%). On the other hand, individuals of above-ground nesting species were larger (8.4–14.5 mm) and predominantly solitary (74%).
Substrate character was the most important predictor for the species richness and abundance of ground nesting, above-ground nesting and solitary wild bees as well as for the abundance of eusocial species (Table 8 in Appendix). Fine substrate had the greatest positive effect on the abundance of ground nesting and eusocial species (R2GLM = 75.5% and 72.2%; Appendix Fig. 6c, g). The observations of Lasioglossum laticeps and Halictus subauratus nests on the roofs with fine substrate underpin that result. The positive effect of fine substrate on ground nesting wild bee diversity (Appendix Fig. 6a) was higher (R2GLM = 35.8%) compared to the effect of substrate depths (R2GLM = 9.3%; Appendix Fig. 6b). Substrate depths affected eusocial species richness positively (R2GLM = 42.4%; Appendix Fig. 6f). The positive effect of fine substrate was weaker on the abundance of above-ground nesting species and solitary species (R2GLM = 30.3%; Appendix Fig. 6e, i). The observation of a Bombus lapidarius colony in the fine substrate of a roof underlines its positive effect on above-ground nesting and eusocial species. The abundance of parasitic wild bees was slightly positively affected by substrate depth (R2GLM = 6.7%, Appendix Fig. 6k). Further, the effect of roof height on parasitic species richness (R2GLM = 4.14%) and abundance (R2GLM = 10.1%) was also weakly positive (Appendix Fig. 6j, l).
The species richness and abundance of polylectic wild bees was highly positively affected by the number of flowering plant species (R2GLM = 58.3%; 64.1%; Appendix Fig. 6m, n). Oligolectic wild bee species richness was enhanced by an increasing number of flowering plant species (R2GLM = 68.2%; Appendix Fig. 6o). Moreover, the abundance of oligolectic species (Appendix Fig. 6p, q) was even higher positively affected by forage availability (R2GLM = 74.6%) than by flowering plant species richness (R2GLM = 66.9%). Increasing roof height decreased the body size (Appendix Fig. 6r) of wild bees moderately (R2GLM = 23.9%).
“Sedum” vs. “Wildflowers” sub-areas
The five roofs with “Sedum” as well as “Wildflowers” sub-areas comprised 55 wild bee species (16.7 ± 7) and 421 individuals (42.2 ± 28.3). On average, the sub-areas ranged from “very low” to “medium” forage availability (1.7 ± 0.6 vs. 2.5 ± 0.5) and differed in flowering plant species richness (3.2 ± 0.8 vs. 21.4 ± 7.1). Wild bee species richness (17.6 ± 5.6) and abundance (44.8 ± 26.0) was slightly higher in “Wildflowers” sub-areas than in “Sedum” (15.8 ± 8.7; 39.6 ± 33.4).
The picture becomes clearer by looking at the whole sampling period: Except for May, the wild bee occurrence and diversity harmonized well with the forage availability in the sub-areas (Fig. 4a-c). For example, in April “Sedum” comprised on average “very low” forage availability and only one wild bee species (3 individuals) was sampled, whereas “Wildflowers” offered higher floral resources and 3 to 9 wild bee species (5 to 14 individuals) were observed. Further, “Sedum” reached the main flowering time in June with “medium” forage availability and 21 wild bee species (62 individuals), but “Wildflowers” decreased to “low” forage availability and only 13 wild bee species (23 individuals) were sampled. In July conditions in the sub-areas were reversed: “Sedum” decreased to “low” forage availability, whereas “Wildflowers” increased to “medium” forage availability and 26 wild bee species (62 individuals) exceeded the 21 wild bee species (42 individuals) in “Sedum” sub-areas.

Timelines to compare (a) total wild bee species richness, (b) total abundance and (c) mean forage availabilities in “Sedum” and “Wildflowers” sub-areas on green roofs during the sampling period 2014; Lines: full = “Sedum”, dashed = “Wildflowers”, dotted square = highlights Sedum main flowering time
The data sub-set of “Sedum” and “Wildflowers” sub-areas revealed that some species were only observed in “Sedum” sub-areas (Table 6 in Appendix) but these species (e.g. Bombus pascuorum or Megachile lagopoda) were also documented on other roofs and in other sub-areas. Therefore, no species was exclusively observed in “Sedum” sub-areas. The highly abundant group of Halictidae showed no preference for a distinct sub-area (Table 6 in Appendix).
Discussion
The studied green roofs hosted 90 wild bee species, which represent 20% of Vienna’s 456 recorded wild bees (Zettel et al. 2016). Compared to other studies, the roofs showed a considerable wild bee diversity: Depending on the number of sampled roofs, the amount of study years and the study’s scope, other authors documented between 17 and 77 wild bee species on green roofs (Brenneisen 2005; Kadas 2006; Tonietto et al. 2011; Ksiazek et al. 2012, 2014; MacIvor et al. 2014; MacIvor 2015). Due to its geographical position, Vienna is influenced by the Pannonian climate, which could be one reason that the studied green roofs emerged with higher species numbers compared to the studies mentioned above. The authors in Switzerland (Brenneisen 2005) and London (Kadas 2006) used yellow-pan traps for wild bee sampling, which may be another reason for the difference in wild bee diversity found on the roofs and makes a comparison of the studies difficult.
The roofs showed lower wild bee diversity compared to ground level habitats in Vienna, where recently 119 to 144 species were reported from the Danube Island (Pachinger and Hölzler 2006), the Viennese Botanical Garden (Hölzler 2004) and the Danube Park (Zettel et al. 2013). Similarly, authors from Chicago reported higher wild bee diversity and abundance on ground level habitats than on green roofs (Tonietto et al. 2011). Green roofs and ground level habitats in Toronto (Colla et al. 2009) and Switzerland (Brenneisen 2005) comprised similar wild bee diversity. However, these comparisons have little informative value because of the different sample sizes and study designs realised in these studies.
The results of our study are interpreted in the light of the small sample size and heterogeneous green roof qualities. This and the assessment of model quality by diagnostic plots reveal that the presented results are difficult to generalise. However, by interpreting and discussing the results carefully we would also like to point out open questions regarding wild bees on green roofs.
Green roof qualities
Wild bee diversity and abundance on green roofs are positively affected by enhanced floral resources and fine substrates. Although the most accurate GLMM indicated that forage availability affected wild bee diversity and abundance primarily, the importance of entomophilous plant species richness was evident by the relationship of these variables. The positive effect of high and ongoing forage availability through a high plant diversity on wild bee communities is known from other ecosystems (e.g. Potts et al. 2003; Zurbuchen and Müller 2012; Braun-Reichert 2013) and was found on green roofs in Switzerland (Brenneisen 2005) and Toronto (Tonietto et al. 2011).
The finding that roofs with fine substrates enhanced wild bee diversity and abundance has to be interpreted with caution, because only two of the nine roofs represented this factor level. Furthermore, planting possibilities on green roofs are limited to substrate properties (Dunnett and Kingsbury 2008). The reason for the positive effect of fine substrates on the overall wild bee diversity and abundance in this study could be the relation of higher floral resources on roofs with fine substrates. But wild bee traits, especially the occurrence of ground nesting and eusocial species may better explain this effect.
Wild bee traits
The studied green roofs mainly attracted pollen generalists. Although polylectic, these wild bees were affected by flowering plant diversity because some of them showed distinct preferences to certain plant taxa. For example, the benefit of Sedum for polylectic species with pollen preferences for Sedum (Anthidium oblongatum, A. strigatum, Hylaeus punctatus, Megachile leachella, M. pilidens, M. rotundata, M. willughbiella) (Westrich 1989b) was pointed out by their higher abundance on roofs with “Sedum” sub-areas compared to other roofs (Appendix Table 6). Oligolectic wild bee species richness was enhanced by increasing floral diversity, because it could raise the probability that different host plants occur on a green roof for species with different pollen specializations. Also, increasing forage availability could lead to higher flower coverage of distinct host plants and enhance the abundance of the respective oligolectic species. However, compared to studies carried out on ground level habitats in Vienna, recording 16 to 24% of oligolectic wild bee species (Hölzler 2004; Pachinger and Hölzler 2006; Zettel et al. 2013), we observed only 11% oligolectic species.
The positive effect of fine substrate on the abundance of ground nesting as well as eusocial species is reflected in the traits of the most abundant wild bee on the roofs: Halictus subauratus. This Sweat bee is a ground nesting and eusocial species, requires fine and sandy substrates and nests are established at 10–15 cm below the surface (Scheuchl and Willner 2016). Therefore, the substrate conditions that positively affected wild bees in this study are beneficial for ground nesting species, which require fine substrates but build their nests shallowly. Different ground nesting species require various substrate properties (e.g. compaction, content of sand or humus, Scheuchl and Willner 2016) and build nests in different depths (Cane and Neff 2011). This was also revealed in our study, because fine substrates as well as substrate depth, affected the diversity of ground nesting wild bees on green roofs. Like in other studies (Brenneisen 2005; Colla et al. 2009; Tonietto et al. 2011), we found Lasioglossum and Halictus species in high abundance (340 individuals) and species richness (20 species) compared to Andrena species with only few or single individuals per species. Two explanations seem reasonable for this observation: Firstly, Andrena species occurrence is more scattered because the females breed solitary. Secondly, Andrena was represented by large species, which probably require deeper substrates for nesting than present on the studied green roofs and just forage here. Similarly, Tonietto et al. (2011) concluded that large ground nesting species nested in adjacent ground level habitats and used green roofs for foraging. Since we did not sample wild bees in adjacent ground level habitats, our data cannot support these findings directly. However, more Andrena species where documented on ground level habitats in Vienna (Pachinger and Hölzler 2006; Zettel et al. 2013) than on the studied green roofs.
Based on the CWM, solitary wild bees were determined to be characteristically on the studied green roofs. It was difficult to determine whether ground nesting or above-ground nesting was the most typical nesting trait, but the latter were more abundant. Above-ground nesting wild bee species are typical of urban areas (reviewed in Hernandez et al. 2009), because the high density of vertical structures offers many potential nesting sites (Cane 2005). Further, we reported above-ground nesting wild bees in higher diversity (43.3% of species) compared to ground level habitats in Vienna, comprising 20 to 32% of above-ground nesting species (Hölzler 2004; Pachinger 2008; Zettel et al. 2013). This could be explained by differences in site features because the mentioned studies were conducted in Vienna’s botanical garden or big recreational areas (e.g. Danube Island) where the density of vertical structures within species-specific flying distances maybe lower than on roofs in urban areas. Further research has to be carried out to identify the effect of green roofs’ surrounding structures on above-ground nesting wild bees.
In contrast to MacIvor (2015), we found no strong evidence, that increasing roof height negatively altered the wild bee community. The CWM of body size from wild bees tended to decrease with increasing roof height. A possible explanation is that smaller species populate high green roofs without utilising ground level habitats. This trend is probably reflected by the weak positive effect of roof height on parasitic wild bees, because the occurrence of brood parasitic species indicates a vital host population (Hudson et al. 2006). For example, the appearance of four Coelioxys species was in line with the occurrence of their host, namely Megachile species, which were recorded in high abundance on the roofs (Table 6 in Appendix).
“Sedum” vs. “Wildflowers”
Based on the study of five roofs, Sedum is an important temporal floral resource because during its main flowering period in June higher wild bee diversity and abundance was observed in this sub-area. Similar observations were reported by MacIvor et al. (2014) who found high proportions of Sedum pollen in palynological samples from wild bees gathered on a green roof during the main flowering period of Sedum. However, in our study qualitative comparison showed no big difference of forage availability between these sub-areas, which may suggest complementary resource availability by Sedum and wildflowers. The concurrent trend of forage availability, wild bee diversity and abundance in the sub-areas over the season underpins the results discussed earlier that forage availability enhances the wild bee community on green roofs. This is supported by similar findings in other studies (Brenneisen 2005; Kadas 2006; Tonietto et al. 2011). The opposite trend in May is explained by observations during fieldwork that Sedum was already flowering and favourably visited by wild bees compared to other flowering plants. All Hylaeus species were exclusively sampled on “Sedum” sub-areas during this month, probably because of a preference for Sedum as forage resource, which is already known for some Hylaeus species (Westrich 1989b). Sedum promoted mainly generalist species because the abundance peaks in June and August (Fig. 4b) are dominated by a few generalist species (e.g. Anthidium oblongatum, Halictus subauratus, Hylaeus punctatus, H. hyalinatus, Megachile rotundata, M. willughbiella). During spring (March, April), early occurring wild bees (Andrena, Anthophora and Osmia) were exclusively sampled in “Wildflowers” sub-areas (Table 6 Appendix), and floral resources were only present in this sub-area (Fig. 4c).
Conclusion
It was demonstrated that the wild bee community on nine green roofs in Vienna was strongly positively affected by floral resources and substrate characteristics. We conclude that fine substrates enhance ground nesting and eusocial wild bee species. This attributes to some guidance (Gedge et al. 2008, 2012) and the proposition by Brenneisen (2006) that the creation of small areas of mounds, consisting of finer and deeper substrates should be a design consideration in the planning processes of (extensive) green roofs in order to support ground nesting wild bee species. Above-ground nesting and solitary species are characteristic in urban environments, but further research is needed to assess, which parameters surrounding green roofs affect them primarily.
Honey bees were observed on all roofs, but their abundance did not interfere with the occurrence of wild bees. Oligolectic wild bee species occurrence was low, but strongly positively affected by increasing floral diversity. Sedum that is typically planted on green roofs promotes polylectic wild bee species, of which some preferably forage on this plant. The study of “Sedum” and “Wildflower” sub-areas on five roofs indicated that Sedum species can compensate temporal lacks of resources on green roofs during its main flowering period. On the other hand, various wildflowers were important foraging resources in spring (March, April) and summer (July, August).
We conclude that increasing floral resources (flower abundance and floral diversity) and the installation of patches with fine and deeper substrates should be considered during the planning process to enhance the wild bee diversity and abundance on green roofs.
References
Amiet F, Müller A, Neumeyer R (1999) Apidae 2: Colletes, Dufourea, Hyleaus, Nomia, Nomioides, Rhophitoides, Rophites, Sphecodes, Systropha. Schweizerische Entomologische Gesellschaft, Neuachtel
Amiet F, Herrmann M, Müller A, Neumeyer R (2001) Apidae 3: Halictus, Lasioglossum. Schweizerische Entomologische Gesellschaft, Neuachtel
Auer I, Böhm R (2011) Wetter und Klima in Wien. Vielfalt auf engstem Raum. In: Berger R, Ehrendorfer F (eds) Ökosystem Wien: Die Naturgeschichte einer Stadt. Böhlau Verlag GesmbH&Co.KG, Wien, pp 88–105
Bates D, Maechler M (2016) Matrix: sparse and dense matrix classes and methods. R package version 1.2-8. https://CRAN.R-project.org/package=Matrix. Accessed 10 Nov 2016
Bates D, Maechler M, Bolker B, Walker S (2015) Fitting Linear Mixed-Effect Models Using lme4. J Stat Softw 67:1–48
Baumann N (2006) Ground-Nesting Birds on Green Roofs in Switzerland: Preliminary Observations. Urban Habitats 4:37–50
van den Berg AE, Hartig T, Staats H (2007) Preference for Nature in Urbanized Societies : Stress, Restoration, and the Pursuit of Sustainability. J Soc Issues 63:79–96
Berndtsson JC (2010) Green roof performance towards management of runoff water quantity and quality: A review. Ecol Eng 36:351–360. https://doi.org/10.1016/j.ecoleng.2009.12.014
Biesmeijer JC, Roberts SPM, Reemer M et al (2006) Parallel Declines in Pollinators and Insect-Pollinated Plants in Britain and the Netherlands. Science 313:351–354. https://doi.org/10.1126/science.1127863
Braun-Reichert R (2013) Der Einfluss unterschiedlicher Beweidung auf die Wildbienen- und Wespenfauna von Kalkmagerrasen Zeitpunkt, Frequenz und Kontinuität der Beweidung. Galathea-Berichte des Kr Nürnberger Entomolgen 29:7–22
Brenneisen S (2003) Ökologisches Ausgleichspotenzial von Extensiven Dachbegrünungen. Bedeutung des Ersatz-Ökotops für den Arten- und Naturschutz und die Stadtentwicklungsplanung. University of Basel, Basel
Brenneisen S (2005) The Natural Roof (NADA): Research Project Report on the use of Extensive Green Roofs by Wild Bees. Wädenswil
Brenneisen S (2006) Space for Urban Wildlife: Designing Green Roofs as Habitats in Switzerland. Urban Habitats 4:27–36
Cane JH (2005) Bees, pollination, and the challanges of sprawl. In: Johnson EA, Klemens MW (eds) Nature in Fragments: The legacy of sprawl. Columbia University Press, New York, pp 109–124
Cane JH, Neff JL (2011) Predicted fates of ground-nesting bees in soil heated by wildfire: Thermal tolerances of life stages and a survey of nesting depths. Biol Conserv 144:2631–2636. https://doi.org/10.1016/j.biocon.2011.07.019
Carter T, Butler C (2008) Ecological impacts of replacing traditional roofs with green roofs in two urban areas. Cities Environ 1:1–17
Castleton HF, Stovin V, Beck SBM, Davison JB (2010) Green roofs; building energy savings and the potential for retrofit. Energy Build 42:1582–1591. https://doi.org/10.1016/j.enbuild.2010.05.004
Colla SR, Willis E, Packer L (2009) Can green roofs provide habitat for urban bees (Hymenoptera: Apidae). Cities Environ 2:1–12
Dathe HH (1980) Die Arten der Gattung Hylaeus F. in Europa (Hymenoptera: Apoidea, Colletidae). Mitt Zool Mus Berl 56:207–294
Dunnett N, Kingsbury N (2008) Planting green roofs and living walls. Timber Press Inc., Oregon
Ebmer PAW (1969) Die Bienen des Genus Halictus LATR. S. L. im Großraum von Linz (Hymenoptera, Apidae): Systematik, Biogeographie, Ökologie und Biologie mit Berücksichtigung aller bisher aus Mitteleuropa bekannten Arten. Teil I. Mit neun Bildtafeln. Natkdl Jb Stadt Linz 1969:133–183
Ebmer PAW (1970) Die Bienen des Genus Halictus LATR.S.L. im Großraum von Linz (Hymenoptera, Apidae): Teil II Mit neun Bildtafeln. Natkdl Jb Stadt Linz 1970:19–82
Ebmer PAW (1971) Die Bienen des Genus Halictus LATR.S.L. im Großraum von Linz (Hymenoptera, Apidae): Teil III mit 19 Bildtafeln. Natkdl Jb Stadt Linz 1971:63–156
Falk S (2015) Field Guide to the Bees of Great Britain and Ireland. British Wildlife Publishing Bloomsbury, London
Fernandez-Canero R, Gonzalez-Redondo P (2010) Green Roofs as a habitat for birds: A review. J Anim Vet Adv 9:2041–2052
Fontaine C, Dajoz I, Meriguet J, Loreau M (2006) Functional Diversity of Plant–Pollinator Interaction Webs Enhances the Persistence of Plant Communities. PLoS Biol 4:129–135. https://doi.org/10.1371/journal.pbio.0040001
Fox J (2003) Effect Displays in R for Generalised Linear Models. J Stat Softw 8(15):1–27. http://www.jstatsoft.org/v08/i15/. Accessed 21 Feb 2017
Frankie GW, Thorp RW, Hernandez J et al (2009) Native bees are a rich natural resource in urban California gardens. Calif Agric 63:113–120. https://doi.org/10.3733/ca.v063n03p113
Gathmann A, Tscharntke T (2002) Foraging ranges of solitary bees. J Anim Ecol 71:757–764
Gedge D (2003) From rubble to redstars. Green Rooftops for Sustainable Communities. Report. Chicago
Gedge D, Newton J, Cradick K, et al (2008) Living Roofs and Walls. Technical Report for the Greater London Authority’s London Plan and Environment Teams. London
Gedge D, Grant G, Kadas G, Dinham C (2012) Creating Green Roofs for Invertebrates. Buglife, Peterborough, pp 1–29
Gokcezade JF, Gereben-Krenn BA, Neumayer J, Krenn HW (2010) Feldbestimmungsschlüssel für die Hummeln Österreichs, Deutschlands und der Schweiz (Hymenoptera, Apidae). Linzer Biol Beitr 42:5–42
Gusenleitner F, Schwarz M, Mazzucco K (2012) Apidae (Insecta: Hymenoptera). In: Schuster R (ed) Biosystematics and Ecology Series No. 29: Checkliste der Fauna Österreichs, No. 6. ÖAW-Verlag der Österreichischen Akademie der Wissenschaften, Vienna, pp 1–129
Hathaway AM, Hunt WF, Jennings GD (2008) A field study of green roof hydrologic and wanter quality performance. Trans ASABE 51:37–44
Hennig EI, Ghazoul J (2012) Pollinating animals in the urban environment. Urban Ecosyst 15:149–166. https://doi.org/10.1007/s11252-011-0202-7
Hernandez JL, Frankie GW, Thorp RW (2009) Ecology of Urban Bees : A Review of Current Knowledge and Directions for Future Study. Cities Environ 2:1–15
Hölzler G (2004) Die Wildbienen des Botanischen Gartens der Universität Wien. In: Pernstich A, Krenn HW (eds) Die Tierwelt des Botanischen Gartens der Universität Wien. Eigenverlag Institut für Angewandte Biologie und Umweltbildung, Vienna, pp 141–163
Hudson PJ, Dobson AP, Lafferty KD (2006) Is a healthy ecosystem one that is rich in parasites? Trends Ecol Evol 21:381–385. https://doi.org/10.1016/j.tree.2006.04.007
Kadas G (2006) Rare Invertebrates Colonizing Green Roofs in London. Urban Habitats 4:66–86
Kennedy CM, Lonsdorf E, Neel MC et al (2013) A global quantitative synthesis of local and landscape effects on wild bee pollinators in agroecosystems. Ecol Lett 16:584–599. https://doi.org/10.1111/ele.12082
Klein A-M, Vaissière BE, Cane JH et al (2007) Importance of pollinators in changing landscapes for world crops. Proc Biol Sci 274:303–313. https://doi.org/10.1098/rspb.2006.3721
Köhler M (2005) Long-Term Vegetation Research on Two Extensive Green Roofs in Berlin. Urban Habitats 4:3–26
Kremen C, Williams NM, Thorp RW (2002) Crop pollination from native bees at risk from agricultural intensification. Proc Natl Acad Sci U S A 99:16812–16816. https://doi.org/10.1073/pnas.262413599
Ksiazek K, Fant J, Skogen K (2012) An assessment of pollen limitation on Chicago green roofs. Landsc Urban Plan 107:401–408
Ksiazek K, Tonietto R, Ascher J (2014) Ten bee species new to green roofs in the Chicago area. Gt Lakes Entomol 47:87–92
Laliberté E, Legendre P, Shipley B (2014) FD: measuring functional diversity from multiple traits, and other tools for functional ecology. R package version 1.0-12.
Landolt E (2001) Orchideen-Wiese in Wollishofen (Zürich) - ein erstaunliches Relikt aus dem Anfang des 20. Jahrhunderts. Vierteljahrsschrift der Naturforschenden Gesellschaft Zürich 146:41–51
Le Féon V, Schermann-Legionnet A, Delettre Y et al (2010) Intensification of agriculture, landscape composition and wild bee communities: A large scale study in four European countries. Agric Ecosyst Environ 137:143–150. https://doi.org/10.1016/j.agee.2010.01.015
MacIvor JS (2015) Building height matters: nesting activity of bees and wasps on vegetated roofs. Isr J Ecol Evol 9801:1–9. https://doi.org/10.1080/15659801.2015.1052635
MacIvor JS, Lundholm J (2010) Insect species composition and diversity on intensive green roofs and adjacent level-ground habitats. Urban Ecosyst 14:225–241. https://doi.org/10.1007/s11252-010-0149-0
MacIvor JS, Ruttan A, Salehi B (2014) Exotics on exotics: Pollen analysis of urban bees visiting Sedum on a green roof. Urban Ecosyst 18:419–430. https://doi.org/10.1007/s11252-014-0408-6
Matteson KC, Langellotto GA (2010) Determinates of inner city butterfly and bee species richness. Urban Ecosyst 13:333–347. https://doi.org/10.1007/s11252-010-0122-y
Matteson KC, Ascher JS, Langellotto GA (2008) Bee Richness and Abundance in New York City Urban Gardens Bee Richness and Abundance in New York City Urban Gardens. Ann Entomol Soc Am 101:140–150
Mauss V (1994) Bestimmungsschlüssel für Hummeln. In: Deutscher Jugendbund für Naturbeobachtungen (ed), 5th edn. Hamburg, p 51
Mazerolle MJ (2016) AICcmodavg: model selection and multimodel inference based on (Q)AIC(c). R package version 2.1-0. https://cran.r-project.org/package=AICcmodavg. Accessed 13 Feb 2017
Mentens J, Raes D, Hermy M (2006) Green roofs as a tool for solving the rainwater runoff problem in the urbanized 21st century ? Landsc Urban Plan 77:217–226. https://doi.org/10.1016/j.landurbplan.2005.02.010
Michener CD (2007) The Bees of the World, 2nd edn. The Johns Hopkins University Press, Baltimore
Motulsky H, Christopoulos A (2003) Fitting Models to Biological Data using Linear and Nonlinear Regression: A practical guide to curve fittin, 4th edn. GraphPad Software, Inc, San Diego
Oberndorfer E, Lundholm J, Bass B et al (2007) Green Roofs as Urban Ecosystems: Ecological Structures, Functions, and Services. Bioscience 57:823–833
Pachinger B (2008) Der Hohlweg am Johannesberg (Wien, Unterlaa) Lebensraum und Trittstein für Wildbienen (Hymenoptera:Apidae). Beiträge zur Entomofaunistik 8:69–83
Pachinger B, Hölzler G (2006) Die Wildbienen (Hymenoptera, Apidae) der Wiener Donauinsel. Beiträge zur Entomofaunistik 7:119–148
Potts SG, Vulliamy B, Dafni A et al (2003) Linking bees and flowers: How do floral communities structure pollinator communities? Ecology 84:2628–2642. https://doi.org/10.1890/02-0136
Potts SG, Biesmeijer JC, Kremen C et al (2010) Global pollinator declines: trends, impacts and drivers. Trends Ecol Evol 25:345–353. https://doi.org/10.1016/j.tree.2010.01.007
R Core Development Team (2016) R: A Language and Environment for Statistical Computing. V 3.3.2. R Foundationi for Statistical Computing, Vienna
RStudio Team (2015) RStudio: Integrated Development for R. V 0.99.903. RStudio Inc., Boston
Scheuchl E (2000) Illustrierte Bestimmungstabellen der Widbienen Deutschlands und Österreichs Band 1: Schlüssel der Gattungen und der Arten der Familie Anthophoridae. Eigenverlag, Velden/Vils
Scheuchl E (2006) Illustrierte Bestimmungstabellen der Wildbienen Deutschlands und Österreichs Band 2: Megachilidae - Melittidae, 2nd edn. Eigenverlag, Velden/Vils
Scheuchl E, Willner W (2016) Taschenlexikon der Wildbienen Mitteleuropas: Alle Arten im Porträt. Quelle & Meyer Verlag, Wiebelsheim
Schindler M, Diestelhorst O, Härtel S et al (2013) Monitoring agricultural ecosystems by using wild bees as environmental indicators. BioRisk 71:53–71. https://doi.org/10.3897/biorisk.8.3600
Schmid-Egger C, Scheuchl E (1997) Illustrierte Bestimmungstabellen der Wildbienen Deutschlands und Österreichs unter Berücksichtigung der Arten der Schweiz Band 3: Schlüssel der Arten der Familie Andrenidae. Eigenverlag, Velden/Vils
Schrader S, Böning M (2006) Soil formation on green roofs and its contribution to urban biodiversity with emphasis on Collembolans. Pedobiologia (Jena) 50:347–356. https://doi.org/10.1016/j.pedobi.2006.06.003
Stadt Wien Vienna GIS (2005) Data: wiener umweltschutzabteilung – MA 22. Basic data: mehrzweckkarte, stadtvermessung wien – MA 41. Further Information: Themenplan Wien Umweltgut. Vienna: Stadt Wien
Susca T, Gaffin SR, Dell’Osso GR (2011) Positive effects of vegetation: Urban heat island and green roofs. Environ Pollut 159:2119–2126. https://doi.org/10.1016/j.envpol.2011.03.007
Tonietto R, Fant J, Ascher J et al (2011) A comparison of bee communities of Chicago green roofs, parks and prairies. Landsc Urban Plan 103:102–108. https://doi.org/10.1016/j.landurbplan.2011.07.004
ViennaGIS (2016) Vienna City Map. https://www.wien.gv.at/stadtplan/. Accessed 5 Dec 2016
von Hagen E, Aichhorn A (2003) Hummeln: bestimmen, ansiedeln, vermehren, schützen, 5. überarb. Fauna Verlag, Nottuln
Westrich P (1989a) Die Wildbienen Baden-Württembergs: Spezieller Teil. Die Gattungen und Arten. Eugen Ulmer GmbH&Co., Stuttgart
Westrich P (1989b) Die Wildbienen Baden-Württembergs: Allgemeiner Teil. Lebensräume, Verhalten, Ökologie und Schutz. Eugen Ulmer GmbH&Co., Stuttgart
Westrich P (1996) Habitat requirements of central European bees and the problems of partial habitats. In: Matheson A, Buchmann SL, O’Toole C, Westrich P and Williams IH (eds) The conservation of bees. Academic Press, London pp 1–16
Winfree R, Abuilar R, Vázquez DP et al (2009) A meta-analysis of bees’ response to anthropogenic disturbance. Ecology 90:2068–2076. https://doi.org/10.1890/07-1861.1
Wojcik VA, McBride JR (2012) Common factors influence bee foraging in urban and wildland landscapes. Urban Ecosyst 15:581–598. https://doi.org/10.1007/s11252-011-0211-6
ZAMG - Zentralanstalt für Meterologie und Geodynamik (2015) Klimaspiegel Wien Hohe Warte für 2014. http://www.zamg.ac.at/cms/de/klima/klima-aktuell/klimaspiegel/jahr/wien_hohe_warte/?jahr=2014. Accessed 5 Dec 2016
Zettel H, Zimmermann D, Wiesbauer H (2013) Die Bienen und Grabwespen (Hymenoptera: Apoidea) im Donaupark in Wien (Österreich). Sabulosi 3:1–23
Zettel H, Zimmermann D, Wiesbauer H (2016) Ergänzungen zur Bienenfauna (Hymenoptera: Apidae) von Wien, Österreich. Beiträge zur Entomofaunistik 17:85–107
Zinzi M, Agnoli S (2012) Cool and green roofs. An energy and comfort comparison between passive cooling and mitigation urban heat island techniques for residential buildings in the Mediterranean region. Energy Build 55:66–76. https://doi.org/10.1016/j.enbuild.2011.09.024
Zurbuchen A, Müller A (2012) Wildbienenschutz - von der Wissenschaft zur Praxis. Bristol-Stifung, Bern
Zurbuchen A, Landert L, Klaiber J et al (2010) Maximum foraging ranges in solitary bees: only few individuals have the capability to cover long foraging distances. Biol Conserv 143:669–676. https://doi.org/10.1016/j.biocon.2009.12.003
Zuur AF, Ieno EN, Elphick CS (2010) A protocol for data exploration to avoid common statistical problems. Methods Ecol Evol 1:3–14. https://doi.org/10.1111/j.2041-210X.2009.00001.x
Zuur AF, Hilbe JM, Ieno EN (2013) A Beginner’s Guide to GLM and GLMM with R: A frequentist and Bayesian perspective for ecologists. Highland Statistics Ltd., Newburgh
Acknowledgements
Open access funding provided by University of Natural Resources and Life Sciences, Vienna (BOKU). We would like to thank the responsible people of the buildings (Mrs. Arlt, Mrs. Ehs, Mrs. Haimer, Mrs. Kapui, Mrs. Leidinger, Mr. Möseler, Mr. Schatovits, Mr. Steinbauer, Mr. Ziemak,) and the municipal council 22 – Environmental Protection in Vienna (especially Mrs. Doppler and Mr. Preiss) for obtaining flexible access to the study roofs. Further, we thank Vera Enzi for her helpful suggestions of green roofs during the selection process of study sites. Special thanks go to Karl Mazzucco for his help with bee species identification, to Luise Kratschmer and Martin Wittner for help with bee preparation, to Matthias Kropf and Michael Kopetzky for help with plant identification, to Dusty Gedge for proof reading and to Bernhard Kratschmer for financial support. The baseline of this work was done in the context of the first author’s master thesis (“Summen auf den Dächern Wiens”) approved by the University of Natural Resources and Life Sciences, Vienna (BOKU) in May 2015.
Author information
Authors and Affiliations
Corresponding author
Appendix
Appendix

Effects of substrate depths (cm) on (a) wild bee species and (b) abundance. Grey band = 0.95 confidence band for fitted effects


Effects of green roof qualities on wild bee species richness and abundance per traits. Error bars and grey bars = 0.95 confidence intervals for fitted effects
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Kratschmer, S., Kriechbaum, M. & Pachinger, B. Buzzing on top: Linking wild bee diversity, abundance and traits with green roof qualities. Urban Ecosyst 21, 429–446 (2018). https://doi.org/10.1007/s11252-017-0726-6
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11252-017-0726-6