
Abstract
Pond networks support high levels of biodiversity when compared to other freshwater ecosystems such as rivers, lakes and streams. The persistence of species in these small, sometimes ephemeral, aquatic habitats depends on the dispersal of individuals among ponds in the landscape. However, the number of ponds across the landscape is at a historical low as urbanisation and intensified agricultural practices have led to a substantial loss of ponds (nodes in the pond network) over more than a century. Here, we examine the extent and drivers of pond loss in a heavily urbanised landscape (Birmingham, UK) over 105 years and determine how pond loss influences key structural properties of the pond network using graph theoretic approaches. Specifically, we calculated minimum spanning trees (MST) and performed percolation analyses to determine changes in both the spatial configuration and resilience of the pond network through time. Pond numbers declined by 82% between ca1904 and 2009, such that pond density decreased from 7.1 km-2 to 1.3 km-2. The MST analyses revealed increased distance between ponds in the network (i.e. edge length increased) by up to 49% over the 105-year period, indicating that ponds in the modern landscape (2009) were considerably more isolated, with fewer neighbours. This study demonstrates that graph theory has an excellent potential to inform the management of pond networks in order to support ecological communities that are less vulnerable to environmental change.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Ponds are discrete aquatic habitats distributed across the terrestrial landscape to form a naturally fragmented network, or 'pondscape' (Boothby 1999) and many pond-dwelling organisms are effective dispersers that have the capacity to move long distances between pond habitats in order to acquire resources, avoid predators, competitors, and disturbance, and seek out conspecifics (Fahrig 2007). As a consequence, local populations in ponds become linked by the movement of individuals to form metapopulations sustained across the wider pond network (e.g. Jeffries 1994; Briers et al. 2004). Therefore, the ability of pond-dwelling organisms to disperse among ponds is especially pivotal in promoting species persistence in a dynamic habitat network (Gibbs 2000; Fortuna et al. 2006) in which individual ponds are gained and/or lost through time via a range of natural and anthropogenic processes (Jeffries 2012).
Land-use change such as urbanisation can limit the natural processes that create ponds such as erosional processes or floodplain dynamics (Indermuehle et al. 2008; Williams et al. 1998b, 2010) and accelerate the destruction of natural ponds (Sukopp 1981) or those used formerly for agriculture or industrial purposes (Wood and Barker 2000). This loss and destruction of pond habitat is common to many countries across the world (Fairchild et al. 2013; Hassall 2014). Across the United Kingdom (UK), 32% of ponds are estimated to have been lost over 120 years between 1880 and 2000: a rate of 0.27% per year (Biggs et al. 2005). Losses have occurred in both rural and urban areas, however, the greatest loss (>80%) has been estimated for urban areas such as London between 1870 and 1984 (Langton 1985) and the city of Cardiff (Rich 1998) or areas of intensive agriculture (Beresford and Wade 1982). These major declines are likely to mask a relatively high turnover of sites as ponds are lost and gained over time (Williams et al. 1998a). Some evidence has emerged to suggest that pond losses may have slowed or reversed recently (Biggs et al. 2005; Williams et al. 2010), potentially as pond creation has become imbedded within amenity developments (Jeffries 2012), or as a result of conservation action (e.g. the Million Ponds Project). Nevertheless, in many regions the number of ponds in the modern landscape is still likely to be the lowest in recorded history, with 80% of remaining ponds in the UK existing in a degraded state (Williams et al. 2010) consistent with other wetland habitats (Defra 2011).
The loss of ponds can threaten the persistence of metacommunities when distances between extant ponds begin to exceed the dispersal abilities of the species they support. Species populations that become isolated by pond loss are at a greater risk of local extinction when faced with environmental disturbances or pollution since they lack nearby habitats from which to source recolonists (Tischendorf and Fahrig 2000; Petersen and Masters 2004; Caquet et al. 2007). Within the network, connectivity to large ponds is important as these often support source populations (Van Geest et al. 2003; Sondergaard et al. 2005; Hill et al. 2015) consistent with source-sink island biogeography (MacArthur and Wilson 1967). Equally, connectivity to small ponds is also important since these are more likely to be fishless and serve as important reservoirs of aquatic invertebrates, amphibians and macrophytes (Oertli et al. 2002; Sondergaard et al. 2005; Scheffer et al. 2006). The overall spatial configuration and topology of the pond network (locations and distances between habitats) is thus a key consideration for freshwater biodiversity conservation (Biggs et al. 1994; Boothby 1997; Lundkvist et al. 2002; Jeffries 2005) and questions remain as to how the loss of ponds affects the metacommunity structure of the wider network. Through spatial analyses such as graph theory (Harary 1969), it is possible to determine the extent to which pond loss has fragmented the pond network, threatening species metapopulations.
Graph theory has recently emerged as a powerful tool to evaluate the connectivity of habitat networks and the movements of wildlife and genes (Garroway et al. 2008), and here we apply it to investigate the possible impacts of urbanisation on pond networks. In graph theory, networks are distilled into graphical form with nodes representing habitat patches, and edges indicating the existence of functioning connections or ‘ecological flux’ between node populations (Urban et al. 2009). Traditional applications of graph theory in the field of ecology have focused on modelling species networks, such as food webs, plant-pollinator mutualistic relationships or host-parasitoid webs (e.g. Proulx et al. 2005; Bascompte et al. 2006). To date, graph theory approaches have focussed on terrestrial habitat networks (e.g. Laita et al. 2010; Gurrutxaga et al. 2011; Decout et al. 2012) and application to aquatic systems has been largely confined to riverscapes (Erős et al. 2011; Segurado et al. 2013; Eros and Campbell Grant 2015), with scant application to lentic systems (Ishiyama et al. 2014).
Percolation theory, the science of clustering or clumping in random networks (Stauffer 1987), can be used to complement graph theoretical analyses in order to identify important network characteristics. Percolation analyses can elucidate network redundancy or robustness where, for example, apparently redundant nodes provide alternative dispersal pathways should any nodes be lost or impacted (Laita et al. 2011). Transposed into analyses of landscape connectivity, percolation theory is the quantitative analysis of connectivity in spatially structured systems (With 2002). Frequently, percolation analyses are undertaken to reflect known dispersal ability of a focal organism or organisms in order to gain an understanding of the relative connectedness of the network (O’Brien et al. 2006; Reunanen et al. 2012; Ishiyama et al. 2014).
Together, graph and percolation theory can be used to gain a strategic oversight of a habitat network (Galpern et al. 2011) and help identify areas of the network with high ecological flux for management planning or policy formation (Fall et al. 2007; Stewart-Koster et al. 2015). For urban areas this could yield better outcomes for nature conservation effort where resources may be limited. Without consideration of the spatial configuration of habitat there remains a risk that, notwithstanding the potential but largely unknown influence of garden ponds, as urban development continues pond networks could become increasingly fragmented and less resilient to multiple environmental stressors and climate change.
Ponds are a good candidate for graph theory analysis (Moilanen 2011), being discrete habitats linked by dispersal of aquatic biota (e.g. invertebrates), many of which live exclusively within pond networks (Céréghino et al. 2007). In this study, major changes in the structure and connectivity of a pond network within Birmingham, a heavily urbanised region of the UK, were identified for a 105 year period (ca1904 – 2009). The structure of the pond network was assessed over time by digitising ponds present on historical Ordnance Survey mapping and the resilience of the network determined by analysing changes in pond distribution, area and number in relation to shifts in land-use. We tested three hypotheses: 1) that considerable pond loss would be observed over the 105 year sequence, 2) pond losses would be strongly associated with urbanisation, and 3) the structural robustness of the Birmingham pond network would decline as the number of ponds in the network decreased.
Materials and methods
Study area
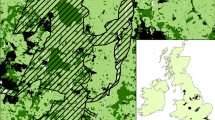
Birmingham (268 km2, 1.1M inhabitants) is located within the Midlands region of the UK (Fig. 1). The area has a rich industrial history of mining and manufacturing, however land-use within contemporary Birmingham comprises mostly of built-up areas and gardens (75% cover), improved grassland, including public parks and gardens (12%), arable and horticulture (8%), mixed, broadleaved or coniferous woodland (4%) and other habitats (1%) (data derived from Land Cover Map 2007). Water bodies (<1% by area) found in the area include rivers, streams and canals as well as lakes, reservoirs and ponds. Ponds with surface area up to 2ha (Biggs et al. 1998), the focus here, are widely dispersed across the region and range from small garden ponds, storm water basins, shallow, naturalised wetlands and ex- marl pits to concrete-lined ornamental ponds within parks.

The Birmingham city administration area (study extent) showing historical pond locations, indicative land-use (Land Cover Map 2007) and United Kingdom context (inset)
Pond digitisation
Data for historical analyses of the pond network in the Birmingham administrative area were derived from three historical map layers, accessed and downloaded in individual 1km2 raster tiles from EDINA Digimap and processed in ArcGIS 9.3 (ESRI, Redlands, CA, USA). First, the County Series 1st revision maps of Birmingham (Edina Historic Digimap Service 2012a) were published 1903-1905 (hereafter ca1904). Second, the National Grid overhaul and re-survey (Edition A) (Edina Historic Digimap Service 2012b) were published 1943-1995 (hereafter as ca1962). Both ca1904 and ca1962 were mapped at 1:2500 scale which enabled landscape features with an area of 16m2 or larger to be identified, although smaller, isolated or significant features may also be mapped (Oliver 2005). The third and most contemporary dataset (2009) was derived from Ordnance Survey MasterMap (EDINA Digimap Ordnance Survey Service 2009). A drawback of each of these mapping methods is that the number of temporary and garden ponds are likely to be underestimated (Jeffries 2012).
Ponds were drawn digitally as individual polygons using ESRI’s ArcScan tool pack extension which permitted surface area calculation. Where possible, curvilinear features without OS map annotation were cross-checked against the other data layers to elucidate pond presence (Thornhill 2013).
Land-use
Land use surrounding each pond was determined for each time period by fitting to Land Cover Map 2007 (LCM2007; Morton et al. 2011) Broad Habitat classifications and grouped accordingly; Farmland (arable and horticulture), Grassland (improved grassland, neutral grassland), Open/scrub (scrub), Suburban and Urban (differentiated by the continuity of urban component; Fuller et al. 2002), Woodland (broadleaved and coniferous woodland). For ca1904 and ca1962 this was achieved by examining map annotations and symbols, and for 2009 by extracting the land-use for each pond directly from LCM2007 coverage data. Using the same method, subsequent land-use was also recorded where ponds were lost (i.e. not drawn) or created over time.
Graph analyses (minimum spanning trees)
Within a graph, edges are considered binary where nodes are connected or not, or they can be quantitative based on probability of connection relative to the distance between them (Dale and Fortin 2010). A path is an unbranched route across a graph in which no node is revisited (Fig. 2a). Multiple connected paths, provided that no closed circuits are created, result in a tree (Fig. 2b), and a tree that connects all nodes within the graph is called a spanning tree. The minimum spanning tree (MST) is the tree that accumulates the least cost (e.g. distance) in connecting all nodes within a graph (Fig. 2c). Urban and Keitt (2001) recommended that conservation efforts should concentrate on the MST as it allows for dispersal across the entire network. Whilst more complex aspects of graph theory can be applied, the MST is likely to indicate the key corridors for the movement and exchange of organisms, or 'backbone' of connectivity (Bunn et al. 2000; Urban and Keitt 2001; Fall et al. 2007) and can be weighted by edge or node features such as surface area. In any graph, metrics such as the betweenness centrality (BCk, Freeman 1977) can also be calculated in order to identify the likely importance of nodes as hubs of connectivity or stepping stones (Minor and Urban 2007) for the wider network. For a full review of definitions and terms see (Urban et al. 2009; Kivelä et al. 2014).

Hypothetical graph arrangements within a pond network. Solid circles indicate connected nodes (ponds) and lines indicate edges (functional connections) where (a) is a path, (b) a tree, (c) a minimum spanning tree (MST) and d) a flux-weighted MST which deviates from the MST due to the increased surface area of node i, grey arrows indicate the direction of net ecological flux
MST metrics were calculated based on distance and pond surface area for each historical pond network as determined by Urban and Keitt (2001). First, a Euclidean distance matrix is calculate for all ponds within the network. A probability matrix is then constructed by applying a negative-exponential decay function, within which the steepness of the distance to probability of dispersal relationship is determined by a distance decay conversion factor (Loro et al. 2015). Finally in this factor, a reference distance is used that has ecological relevance (Ribeiro et al. 2011). Here, a distance of 1km was selected, broadly consistent with knowledge of aquatic organism dispersal including insects (e.g. Conrad et al. 1999; Angelibert and Giani 2003) and amphibians such as the newt Triturus cristatus (Boothby 1999; Kupfer and Kneitz 2000).
Calculation of a flux-weighted MSTf is achieved with the inclusion of relative patch size within the network where larger nodes contribute a higher degree of flux based on the assumption that they contribute larger and more stable populations (Erős et al. 2011; Segurado et al. 2013; Fig. 2d). Minimum spanning trees were derived using the package 'igraph' (Csardi and Nepusz 2006) for R Statistical Software (R Core Team 2017). The igraph package also allowed the computation of a number of diagnostics including average edge length, number of sub components (or clusters) and betweeness centrality (BCk) (Freeman 1977). The value of BCk for a given node is the number of shortest pathways between all pairs of nodes in a graph that require it as a stepping stone, which can also be expressed as a proportion to allow for comparison under different scenarios where there may be differing numbers of nodes (e.g. different historical pond networks).
Percolation analysis
Percolation analyses were carried out in order to consider connectivity across the pond network with respect to observed dispersal abilities of a range of pond dwelling organisms (see Supplementary material T1). Within percolation analysis, the degree of connectivity in a habitat network can be assessed by deriving an order parameter (e.g. the relative size of greatest connected component (GCC) to the whole network) which defines the transition from a connected system to a disconnected one (Kivelä et al. 2014). The distance at which the GCC is no longer evident is called the percolation threshold and represents a critical fraction of links that needs to be removed to break the network into disconnected components (Kivelä et al. 2014). In a pond network with a greater degree of robustness where redundant nodes are abundant, the percolation threshold will occur at a lower distance as the GCC is preserved for longer. The proximity of the percolation threshold to known dispersal abilities can therefore reveal information about the relative accessibility of the network to a given organism. Percolation analyses were carried out using EDENetworks (Kivelä et al. 2014).
Results
Pond habitat loss and turnover through time
Between ca1904 and 2009 there was an 82% decline in the number of ponds across Birmingham (Table 1), reflecting a net loss of 1573 ponds. Most (73%) ponds were lost between ca1904 and ca1962 reflecting an average loss of 15 ponds per year. Pond density declined from 7.1 km-2 in ca1904 to 2.0 km-2 by ca1962 and to 1.3 km-2 in 2009. Over time, the mean surface area of ponds increased (by 204%) as many small waterbodies were lost and larger ones retained (Table 2). This shift was statistically significant early in the sequence (1904 and ca1962, Mann-Whitney, P < 0.05), but not later (ca1962 and 2009, Mann-Whitney, P = 0.12). The total area of all habitat within the Birmingham pond network declined by 46% between ca1904 and 2009, although the retention of larger ponds offset the rapid loss of individual sites. As a proportion of the entire Birmingham administrative area, pond coverage was 0.36% in ca1904, 0.22% in ca1962 and 0.20% in 2009.
Changes in the total number of ponds masks a considerable amount of turnover within the pond stock over time (Table 3). Half (50.1%, 171 sites) of the ponds reviewed in 2009 were over 105 years old (present at ca1904 census) and half (49.9%, 170) were younger, created and not subsequently lost between ca1904 and ca1962 (17%, 58 sites) or between ca1962 and 2009 (32.3%, 112). The 171 ponds that persisted throughout the study represented just 8.9% of the number that were present in ca1904. Whilst overall mean surface area of ponds increased, the mean surface area of ponds that were present throughout the study period decreased over time from 2413m2 (ca1904) to 1935m2 (ca1962) to 1919m2 (2009) which represents an overall 21.5% decrease in average surface area.
Many ponds were originally associated with farmland, however, the number of such ponds decreased rapidly from 71% of total pond numbers in ca1904 to just 15% by 2009 (Table 4). Much pond loss was attributable to suburban expansion, which was the land-use identified to have replaced lost ponds in 875 (3.26 /km2) and 106 (0.40 /km2) cases between ca1904 and ca1962, and ca1962 and 2009 respectively (Table 3). Net losses within farmland were the second highest. The number of ponds associated with, or enveloped by, suburban areas and areas of grassland (including public parkland and golf courses) increased by 2009 to account for a total of 57% of those present, from just 8% in ca1904. The number of ponds associated with urban / industrial land-use was consistently low, at no point accounting for more than 8.5% (0.11 /km2) of ponds present. Though suffering an overall loss in numbers, the number of ponds associated with woodlands relative to all those within the network consistently increased through time from 9.6% (ca1904) to 17.6% (2009).
In total, 2195 geographically distinct ponds were present at any one time throughout the 105 year study period. A total of 283 were created and 1856 lost (Table 3). The study revealed the origins and use of a subset (334 ponds, 15%) of Birmingham's ponds. Most, often small (mean surface area 493m2), were built for purposes of landscaping (101 ponds, 30%) as either ornamental features (67 ponds) or moats (39 ponds). The minerals industry was the second most frequent contributor (81 ponds, 24%), of which just over half were as a result of brickworks (42 ponds, 52%). The third most important use was for recreational purposes, predominately for fishing, accounting for 18% (60 ponds) whilst other industrial uses (e.g. disused reservoirs, sludge lagoons, mill ponds) comprised a further 16%. Ponds created as a result of other industrial uses or for recreational purposes (e.g. fishing) were typically large relative to this audit (mean surface area 4129m2). Very few ponds (20, 6%) appeared to be created through natural processes; the majority (84%) of the subset were thus artificial, having been created to support human activities although it was not possible to ascertain the origins of 87% of ponds that were already present in ca1904.
Pond network resilience through time
Changes in the pond network were characterized by a 49% increase in MSTf mean edge length from 462m to 687m, meaning that ponds within the 2009 network were considerably more isolated with fewer neighbours. Mean centrality values (BCk, normalised for each time period) showed an increase of 210%, suggesting that biodiversity movement across the network was increasingly reliant upon fewer ponds (Table 2).
In the ca1904 and ca1962 landscapes the MSTf routed outside of the central area of Birmingham through areas of higher pond density to the south and to the east (Fig. 3b, c). In ca1904, three distinct patches of high pond density were apparent in the northeast, east and south of Birmingham centre. However, the numbers of ponds in these areas were much diminished by 2009 (Fig. 3a) where up to 30 ponds km-2 were lost between ca1904 and ca1962. Although much overshadowed by the impact of urban expansion affecting the wider pond network, several new ponds have been created as part of modern developments since ca1962 that marginally improve pond density within some central areas of Birmingham. Due to the loss of ponds to the south and east in particular, and a slight gain in the city centre, the 2009 MSTf re-routes through the central area (Fig. 3a).

Flux weighted minimum spanning trees for the Birmingham pond network for (a) ca1904, (b) ca1962 and (c) 2009. Nodes represent ponds, larger nodes indicate higher BCk relative to the network. Lines represent edges. Grey circles represent the location of the contemporary Birmingham city centre
Analysis of the distribution of centrality values across different size classes of pond within the MSTf demonstrated decreasing levels of redundancy in the pond network through time (Fig. 4). Across the ca1904 network the largest ponds (>5000m2) are also the most central typically with BCk values of approximately 0.5. Since ca1904 however, despite the retention of larger ponds, they are much less central (lower BCk values) indicating an inadequate network of stepping stones to allow the MSTf to route through habitats more likely to provide source populations.

Distribution of betweeness centrality measures between ponds (nodes) of different size classes within the flux-weighted MSTf for each historical pond network
In the ca1904 network, more nodes are retained within Greatest Connected Component (GCC) as the distance threshold is decreased resulting in a percolation threshold of 811m (Fig. 5). However, as ponds are lost over time, the percolation threshold increased to 1559m by ca1962 to 2361m by 2009 (Table 2). If a dispersal threshold of 1km was applied to the networks, most (92.4%) of ponds would comprise the GCC in the ca1904 pondscape whilst it would only be comprised of 148 (28.2%) and 63 (18.5%) ponds by ca1962 and 2009 networks respectively. Similarly, the total number of sub-components created by applying a 1km threshold proportionally increases over time, thus in ca1904, 42 sub-components were created from a possible 1914 (0.4%), whilst in ca1962 and 2009 33 (6.3%) and 7 (12.3%) sub components were created respectively (Fig. 6).

Percolation graphs for Birmingham’s historical pond networks demonstrating the impact of a decreasing threshold distance upon the greatest connected component (GCC) relative to the number of nodes within the pond network. Percolation thresholds also shown as solid vertical lines for (a) ca1904 - 811m, (b) ca1962 – 1559m and (c) 2009 – 2361m

Fragmentation of Birmingham’s historical pond networks demonstrated by the number of sub-components relative to the maximum number possible (i.e.the total number of nodes), generated as a result of a decreasing threshold distance
Discussion
This study aimed to assess how Birmingham's network of pond habitats has altered in response to increasing urbanisation over a 105 year period and consider changes in network robustness and implications for biodiversity. A rate of pond loss in Birmingham between ca1904 and 2009 of 0.78% per annum is comparable to that of London (0.79%) between 1870 and 1984 (Table 1), and an 82% total loss in total pond numbers between ca1904 and 2009 ranks Birmingham second highest in the UK (accept hypothesis one), behind urban London and comparable to losses in Bedfordshire's intensively agricultural landscape (Beresford and Wade 1982). Whilst there are few records of ponds loss outside of the UK, losses in Birmingham appear greater than nationwide losses observed in Sweden (Bjureke et al. 1976) and comparable to those in the Netherlands (Weinreich and Musters 1994). Since ca1962, the rate of annual pond loss has declined to 0.1%, consistent with the nationwide disappearance of ponds reported by Biggs et al. (2005). Therefore, pond-dwelling organisms within Birmingham are likely to rely upon fewer ponds, potentially rendering their metacommunities less resilient to stochastic events such as pollution or deterministic changes such as global climate change. Ponds that have remained throughout the study (171 ponds) have a notably reduced surface area, which may suggest the occurrence of natural successional processes e.g. vegetation encroachment, or development pressures.
Pond resource turnover, loss and creation
As reported by a number of other authors (Williams et al. 1998a; Jeffries 2012), the raw numbers mask a high turnover in pond resource. The turnover in stock is clearly linked to land-use change and strongly driven by the process of urbanisation (accept hypothesis two) not dissimilar to the impact of coastal urbanisation upon an estuarine wetland network (Dou and Cui 2014). Here however, former farmland field ponds were either lost to, or enveloped by, suburban development as others (few by comparison) were built as part of those developments. These findings accord with several studies of Birmingham's demography and changing landscape, where the population of Birmingham has increased from approximately 500,000 in 1900 to 1M by the early 2000s (Haynes 2008; University of Portsmouth 2017), which coincided with the expansion of Birmingham city centre throughout the 20th century as villages and hamlets coalesced into suburbs through industrial and residential development on former agricultural land (Axinte 2015).
The vast majority of ponds lost within Birmingham were probably artificial in nature, however in a highly altered landscape they are likely to act as surrogates for natural habitats and studies have shown artificial ponds to have high conservation value (Vermonden et al. 2009; Hill et al. 2017; Thornhill et al. 2017). Nevertheless, it is apparent that many ponds are more isolated from their neighbouring habitats and there is a large body of evidence in the published literature that this degeneration of the pond network with less connected nodes has large implications for local and regional biodiversity (e.g. Table 1).
A recent reduction in the rate of pond loss within Birmingham may be due to the retention of larger ponds, which are frequently located in public green spaces or used for recreation (e.g. boating and fishing). Such cultural landmarks may receive protection through local authority planning policies or legislation. Applying conventional island biogeography (MacArthur and Wilson 1967), the retention of high quality larger ponds could help to preserve some of the network's source populations and reduce the overall impact of pond loss. However, a number of studies have also shown that a cluster of small ponds are key contributors to regional invertebrate biodiversity (Wood et al. 2001; Scheffer et al. 2006; Boix et al. 2012). Reasons for this are complex, however it may be due to an increased fish and waterfowl presence in larger ponds resulting in the exclusion of some invertebrate species and reduction in vegetation complexity (Oertli et al. 2002; Sondergaard et al. 2005; Schilling et al. 2009) or to an increase in the number of habitat niches available across several small ponds (Williams et al. 2004). Nevertheless, the potential ecological value in retaining larger ponds for biodiversity may be compromised in Birmingham due to the loss of smaller stepping stone habitats that would otherwise connect larger ponds to the network by facilitating species dispersal.
Possible implications for the biota
As a result of pond loss, the 2009 percolation threshold (2.36km) suggests that for many aquatic biota the Birmingham pond network is comprised of a series of sub-components, the number of which can be inferred through statistical thresholding (Fig. 6). A percolation threshold of 811m and average MSTf edge length of 462m indicated that more frequent exchange of biota between ponds was likely in the ca1904 pond network, which would have historically allowed more rapid recovery of local populations from stochastic events. For invertebrates, the exception may be a small percentile of species populations which make long distance movements (Conrad et al. 1999) which may be sufficient to maintain genetic diversity (Lowe and Allendorf 2010) but not population recovery.
The majority of macroinvertebrate dispersal studies have focused on Odonates, though nearly all suggest that dispersal, particularly of Zygopterans, beyond 1km is rare and due to high levels of philopatry the majority of movements are constrained to less than 100m (Rouquette and Thompson 2007; Supplementary Material Table T1). Though potentially less severe due to stronger dispersal tendencies, the scenario is likely to be similar for some Hemipterans (Briers 1998) and Diptera (Service 1997). Less clear is the impact that pond loss is likely to have had on non-winged (i.e. passive) invertebrate dispersal (e.g. Gastropods and leeches (Hirudinea)) which are largely incapable of self-dispersal between habitats and rely on vectors (Bilton et al. 2001).
Analysis of the spatial configuration of pond networks alone suggests that the majority of the 2009 Birmingham pond network has too few ponds that are typically too far apart to sustain populations of the European protected amphibian, Triturus cristatus. T. cristatus is generally considered to disperse up to 250m (Langton et al. 2001), with few studies reporting movements up to 1km (Kupfer and Kneitz 2000) and optimum pond density for a T. cristatus metapopulation is considered to be 4km-2 (Oldham et al. 2000), which is seldom achieved by 2009 in this study (mean pond density 1.3km-2). These findings may substantiate the suggestion that T. cristatus populations across Birmingham are generally thought to have experienced a decline, though limited data exist (The Wildlife Trust 2000).
Although the overall robustness of the Birmingham pond network has clearly declined with probable implications for many pond-dwelling organisms, further study is required to understand the relative significance of impacts to biota with different dispersal modes and strengths (partially accept hypothesis 3). In addition, this analysis does not represent probable losses in temporary ponds (Jeffries 2012) and or the occurrence small garden ponds which are estimated to be present within 10% of UK gardens (Davies et al. 2009). However, whilst valuable as temporary refuges the biodiversity supported by garden ponds has been shown to be a nested subset of field ponds (Hill and Wood 2014) and potentially unlikely to offset their loss.
Future directions
The present study provides the first application of graph theory to a pond network. However, research in three areas would improve the ecological grounding of such spatial models. First, dispersal across the urban landscape is highly unlikely to be uniform as it is comprised of many obstacles such as roads (Parris 2006) and artificial lighting (Bilton et al. 2001; Smith et al. 2009). Small aquatic invertebrates in particular are difficult to track and efforts have focused primarily on rare species (e.g. Purse et al. 2003; Hassall and Thompson 2012). Second, within habitat quality could not be assessed, yet national studies have identified a decline in the quality of ponds (Williams et al. 2010), which could suggest that many included here may not be suitable for colonisation for pollution sensitive taxa. To this end, public participation in data generation (i.e. citizen science; Thornhill et al. 2016) and improvements in remote-sensing (Palmer et al. 2015) are promising avenues. Thirdly, we used a 1km threshold as a broad average of the dispersal ability of the community, however, a more in depth analysis could apply shorter and longer thresholds to better represent the varied abilities of the ecological community to disperse (Galpern et al. 2011), whilst being careful not carry out analysis without sufficient evidence base (Moilanen 2011); thus referring back to point one above.
Conclusion
The identification of the backbone of an urban pond network by using graph theory and the concept of minimum spanning trees (MSTs) is the beginning of a landscape-scale strategy for the conservation of pond fauna and flora rather than traditional single site management. An extended analysis should be carried out to include the wider pond network such that study boundaries are reflective of natural boundaries. However, this study highlights important clusters and pathways within the current pond stock as well as evidence of a need to improve the networks spatial resilience.
This study finds that ponds have become increasingly scarce in the urban landscape over more than 100 years as they are lost to the process of urbanisation. The loss of ponds since ca1904 is considerable, but the rate of pond loss has slowed in recent times. This may be reflective of the types of ponds which are being retained as they are often in the public eye and form part of amenity parkland.
The manner in which this study has been carried out is stepwise and intuitive and can be undertaken with widely available software such that it may be readily repeated across regions and adapted for other landscapes. Landscape managers should ensure that the ponds that remain are of good quality and could use the analytical approach presented here to strategically create ponds in order to reduce the vulnerability of the pond network to further habitat loss. However, opportunities remain to further refine the approach by incorporating inter-habitat resistance to dispersal due to unfavourable land-use.
References
Angelibert S, Giani N (2003) Dispersal characteristics of three odonate species in a patchy habitat. Ecography 1:13–20
Axinte A (2015) Birmingham Historic Landscape Characterisation (HLC).
Bailey-Watts T, Lyle A, Battarbee R et al (2000) Lakes and ponds. In: Acreman M (ed) The hydrology of the UK: a study of change. Routledge, London, pp 180–203
Bascompte J, Jordano P, Olesen JM (2006) Asymmetric coevolutionary networks facilitate biodiversity maintenance. Science 312:431–433. https://doi.org/10.1126/science.1123412
Beebee TJC (1997) Changes in dewpond numbers and amphibian diversity over 20 years on chalk downland in Sussex, England. Biol Conserv 81:215–219. https://doi.org/10.1016/S0006-3207(97)00002-5
Beresford JE, Wade PM (1982) Field ponds in North Leicestershire: their characteristics, aquatic flora and decline. Trans Leicester Lit Philos Soc 76:25–34
Biggs J, Corfield A, Walker D (1994) New approaches to the management of ponds. Br Wildl 5:273–287
Biggs J, Fox G, Nicolet P, et al (1998) A guide to the methods of the National Pond Survey.
Biggs J, Williams P, Whitfield M et al (2005) 15 Years of pond assessment in Britain: Results and lessons learned from the work of Pond Conservation. Aquat Conserv Mar Freshw Ecosyst 15:693–714. https://doi.org/10.1002/aqc.745
Bilton DT, Freeland JR, Okamura B (2001) Dispersal in freshwater invertebrates. Annu Rev Ecol Syst 32:159–181
Bjureke K, Dahlgren U, Fronaeus M et al (1976) Margel I Lundabygden. Lund, Sweden
Boix D, Biggs J, Céréghino R et al (2012) Pond research and management in Europe: “Small is Beautiful.”. Hydrobiologia 689:1–9. https://doi.org/10.1007/s10750-012-1015-2
Boothby J (1997) Pond conservation: Towards a delineation of pondscape. Aquat Cons Mar Freshw Ecosyst 7:127–132
Boothby J (1999) Framing a strategy for pond landscape conservation: aims, objectives and issues. Landsc Res 24:67–83. https://doi.org/10.1080/01426399908706551
Boothby J, Hull AP (1997) A census of ponds in Cheshire. North West England. 7:75–79
Bressi N, Stoch F (1998) Karstic ponds and pools: history, biodiversity and conservation. In: Ponds and pond landscapes of Europe. Proceedings of the International Conference of the Pond Life Project. Maastricht, The Netherlands,
Briers RA (1998) Metapopulation ecology of Notonecta in small ponds. University of Sheffield
Briers RA, Gee JHR, Cariss HM, Geoghegan R (2004) Inter-population dispersal by adult stoneflies detected by stable isotope enrichment. Freshw Biol 49:425–431. https://doi.org/10.1111/j.1365-2427.2004.01198.x
Bunn AG, Urban DL, Keitt TH (2000) Landscape connectivity: a conservation application of graph theory. J Environ Manage 59:265–278
Caquet T, Hanson ML, Roucaute M et al (2007) Influence of isolation on the recovery of pond mesocosms from the application of an insecticide. II. Benthic macroinvertebrate responses. Environ Toxicol Chem 26:1280–1290
Céréghino R, Biggs J, Oertli B, Declerck S (2007) The ecology of European ponds: defining the characteristics of a neglected freshwater habitat. Hydrobiologia 597:1–6. https://doi.org/10.1007/s10750-007-9225-8
Conrad KF, Willson KH, Harvey IF et al (1999) Dispersal characteristics of seven odonate species in an agricultural landscape. Ecography (Cop) 22:524–531. https://doi.org/10.1111/j.1600-0587.1999.tb00541.x
Csardi G, Nepusz T (2006) The igraph software package for complex network research. InterJournal Complex Sy:1695.
Dale MRT, Fortin M-J (2010) From Graphs to Spatial Graphs. Annu Rev Ecol Evol Syst 41:21–38. https://doi.org/10.1146/annurev-ecolsys-102209-144718
Davies ZG, Fuller RA, Loram A et al (2009) A national scale inventory of resource provision for biodiversity within domestic gardens. Biol Conserv 142:761–771. https://doi.org/10.1016/j.biocon.2008.12.016
Decout S, Manel S, Miaud C, Luque S (2012) Integrative approach for landscape-based graph connectivity analysis: a case study with the common frog (Rana temporaria) in human-dominated landscapes. Landsc Ecol 27:267–279. https://doi.org/10.1007/s10980-011-9694-z
Defra (2011) Water for Life. Department for Environment and Rural Affairs, London
Dou P, Cui B (2014) Dynamics and integrity of wetland network in estuary. Ecol Inform 24:1–10. https://doi.org/10.1016/j.ecoinf.2014.06.002
EDINA Digimap Ordnance Survey Service (2009) OS MasterMap Topography Layer [GML geospatial data], Coverage: Birmingham, Updated Jan 2009. Ordnance Survey, GB
Edina Historic Digimap Service (2012a) Birmingham, Sandwell, Walsall, Wolverhampton and Dudley areas. Scale 1:2,500. Ordnance Survey County Series 1st Revision [TIFF geospatial data] Published 1902-1905. Landmark Information Group, UK
Edina Historic Digimap Service (2012b) Birmingham, Sandwell, Walsall, Wolverhampton and Dudley areas. Scale 1:2,500. National Grid Series overhaul and resurvey (Edition A) [TIFF geospatial data] Published 1933-1977. Landmark Information Group, UK
Eros T, Campbell Grant EH (2015) Unifying research on the fragmentation of terrestrial and aquatic habitats: patches, connectivity and the matrix in riverscapes. Freshw Biol 60:1487–1501. https://doi.org/10.1111/fwb.12596
Erős T, Olden JD, Schick RS et al (2011) Characterizing connectivity relationships in freshwaters using patch-based graphs. Landsc Ecol 27:303–317. https://doi.org/10.1007/s10980-011-9659-2
Fahrig L (2007) Non-optimal animal movement in human-altered landscapes. Funct Ecol 21:1003–1015. https://doi.org/10.1111/j.1365-2435.2007.01326.x
Fairchild GW, Robinson C, Brainard AS, Coutu GW (2013) Historical Changes in the Distribution and Abundance of Constructed Ponds in Response to Changing Population Density and Land Use. Landsc Res 38:593–606. https://doi.org/10.1080/01426397.2012.672640
Fall A, Fortin MJ, Manseau M, O’Brien D (2007) Spatial graphs: Principles and applications for habitat connectivity. Ecosystems 10:448–461. https://doi.org/10.1007/s10021-007-9038-7
Fortuna MA, Gómez-Rodríguez C, Bascompte J (2006) Spatial network structure and amphibian persistence in stochastic environments. Proc Biol Sci 273:1429–1434. https://doi.org/10.1098/rspb.2005.3448
Freeman L (1977) A set of measures of centrality based on betweenness. Sociometry 40:35–41
Fuller RM, Smith GM, Sanderson JM, et al (2002) Countryside Survey 2000 Module 7 Land Cover Map 2000. Dorchester
Galpern P, Manseau M, Fall A (2011) Patch-based graphs of landscape connectivity: A guide to construction, analysis and application for conservation. Biol Conserv 144:44–55. https://doi.org/10.1016/j.biocon.2010.09.002
Garroway CJ, Bowman J, Carr D, Wilson PJ (2008) Applications of graph theory to landscape genetics. Evol Appl 1:620–630. https://doi.org/10.1111/j.1752-4571.2008.00047.x
Gibbs JP (2000) Wetland Loss and Biodiversity Conservation. Conserv Biol 14:314–317. https://doi.org/10.1046/j.1523-1739.2000.98608.x
Glindt D (1993) Situation, plege unt Neuanlage kleiner Stillgewasser im Flachland Norwestdeutschlands. Metelen, Germany
Globevnik L, Kirn T (2009) Small water bodies - Assessment of status and threats of standing small water bodies.
Guadagnin DL, Peter ÂS, Perello LFC, Maltchik L (2005) Spatial and Temporal Patterns of Waterbird Assemblages in Fragmented Wetlands of Southern Brazil. Waterbirds 28:261–272. https://doi.org/10.1675/1524-4695(2005)028[0261:SATPOW]2.0.CO;2
Gurrutxaga M, Rubio L, Saura S (2011) Key connectors in protected forest area networks and the impact of highways: A transnational case study from the Cantabrian Range to the Western Alps (SW Europe). Landsc Urban Plan 101:310–320. https://doi.org/10.1016/j.landurbplan.2011.02.036
Harary F (1969) Graph theory. Addison-Wesley, Reading
Hassall C (2014) The ecology and biodiversity of urban ponds. Wiley Interdiscip Rev Water 1:187–206. https://doi.org/10.1002/wat2.1014
Hassall C, Thompson DJDJ (2012) Study design and mark-recapture estimates of dispersal: A case study with the endangered damselfly Coenagrion mercuriale. J Insect Conserv 16:111–120. https://doi.org/10.1007/s10841-011-9399-2
Haynes M (2008) The Evolution of the Economy of the West Midlands 1700-2007. Wolverhampton
Heath DJ, Whitehead A (1992) A survey of pond loss in Essex, south-east England.
Hill MJ, Biggs J, Thornhill I et al (2017) Urban ponds as an aquatic biodiversity resource in modified landscapes. Glob Chang Biol. https://doi.org/10.1111/gcb.13401
Hill MJ, Mathers KL, Wood PJ (2015) The aquatic macroinvertebrate biodiversity of urban ponds in a medium-sized European town (Loughborough, UK). Hydrobiologia 760:225–238. https://doi.org/10.1007/s10750-015-2328-8
Hill MJ, Wood PJ (2014) The macroinvertebrate biodiversity and conservation value of garden and field ponds along a rural-urban gradient. Fundam Appl Limnol / Arch für Hydrobiol 185:107–119. https://doi.org/10.1127/fal/2014/0612
Hutinec BJ, Struna S (2007) A survey of ponds and their loss in Žumberak-Samoborsko Gorje Nature Park, northwest Croatia. Nat Croat 16:121–137
Indermuehle N, Oertli B, Biggs J, et al (2008) Pond conservation in Europe : the European Pond Conservation Network ( EPCN ). 30:446–448
Ishiyama N, Akasaka T, Nakamura F (2014) Mobility-dependent response of aquatic animal species richness to a wetland network in an agricultural landscape. Aquat Sci 76:437–449. https://doi.org/10.1007/s00027-014-0345-8
Jeffries MJ (1994) Invertebrate communities and turnover in wetlands ponds affected by drought. Freshw Biol 32:603–612
Jeffries M (2005) Small ponds and big landscapes: the challenge of invertebrate spatial and temporal dynamics for European pond conservation. Aquat Conserv Mar Freshw Ecosyst 15:541–547. https://doi.org/10.1002/aqc.753
Jeffries MJ (2012) Ponds and the importance of their history: an audit of pond numbers, turnover and the relationship between the origins of ponds and their contemporary plant communities in south-east Northumberland, UK. Hydrobiologia 689:11–21. https://doi.org/10.1007/s10750-011-0678-4
Jeffries MJ, Mills D (1990) Freshwater ecology: principles and applications. Wiley, Chichester
Kivelä M, Arnaud-Haond S, Saramäki J (2014) EDENetworks: A user-friendly software to build and analyse networks in biogeography, ecology and population genetics. Mol Ecol Resour:117–122. https://doi.org/10.1111/1755-0998.12290
Kupfer A, Kneitz S (2000) Population ecology of Great Crested Newts (Triturus cristatus) in an agricultural landscape: dynamics, pond fidelity and dispersal. Herpetol J 10:165–172
Laita A, Kotiaho JS, Mönkkönen M (2011) Graph-theoretic connectivity measures: what do they tell us about connectivity? Landsc Ecol 26:951–967. https://doi.org/10.1007/s10980-011-9620-4
Laita A, Mönkkönen M, Kotiaho JS (2010) Woodland key habitats evaluated as part of a functional reserve network. Biol Conserv 143:1212–1227. https://doi.org/10.1016/j.biocon.2010.02.029
Langton T (1985) The London pond survey. Oryx 19:163–166
Langton TES, Beckett CL, Foster JP (2001) Great Crested Newt Conservation Handbook. Froglife, Halesworth
Loro M, Ortega E, Arce RM, Geneletti D (2015) Ecological connectivity analysis to reduce the barrier effect of roads. An innovative graph-theory approach to define wildlife corridors with multiple paths and without bottlenecks. Landsc Urban Plan 139:149–162. https://doi.org/10.1016/j.landurbplan.2015.03.006
Lowe WH, Allendorf FW (2010) What can genetics tell us about population connectivity? Mol Ecol 19:3038–3051. https://doi.org/10.1111/j.1365-294X.2010.04688.x
Lundkvist E, Landin J, Karlsson F (2002) Dispersing diving beetles ( Dytiscidae ) in agricultural and urban landscapes in. Ann Zool Fennici 39:109–123
MacArthur RH, Wilson EO (1967) The Theory of Island Biogeography. Princeton University Press, Princeton
Minor ES, Urban DL (2007) Graph theory as a proxy for spatially explicit population models in conservation planning. Ecol Appl 17:1771–1782. https://doi.org/10.1890/06-1073.1
Moilanen A (2011) On the limitations of graph-theoretic connectivity in spatial ecology and conservation. J Appl Ecol 48:1543–1547. https://doi.org/10.1111/j.1365-2664.2011.02062.x
Morton D, Rowland C, Wood C, et al (2011) Final report for LCM2007 – the new UK land cover map. CS Technical Report No 11/07 NERC/Centre for Ecology & Hydrology 112pp. (CEH project number: C03259)
O’Brien D, Manseau M, Fall A, Fortin MJ (2006) Testing the importance of spatial configuration of winter habitat for woodland caribou: An application of graph theory. Biol Conserv 130:70–83. https://doi.org/10.1016/j.biocon.2005.12.014
Oertli B, Joye DA, Castella E et al (2002) Does size matter? The relationship between pond area and biodiversity. Biol Conserv 104:59–70. https://doi.org/10.1016/S0006-3207(01)00154-9
Oldham R, Keeble J, Swan M, Jeffcote M (2000) Evaluating the suitability of habitat for the great crested newt (Triturus cristatus). Herpetol J 10:143–155
Oliver R (2005) Ornance Survey maps: a concise guide for historians. The Charles Close Society, London
Palmer SCJ, Kutser T, Hunter PD (2015) Remote sensing of inland waters: challenges, progress and future directions. Remote Sens Environ. https://doi.org/10.1016/j.rse.2014.09.021
Parris KM (2006) Urban amphibian assemblages as metacommunities. J Anim Ecol 75:757–764. https://doi.org/10.1111/j.1365-2656.2006.01096.x
Petersen I, Masters Z (2004) Dispersal of adult aquatic insects in catchments of differing land use. :934–950
Pieńkowski P (2003) Disappearance of the mid-field ponds as a result of agriculture intensification
Proulx SR, Promislow DEL, Phillips PC (2005) Network thinking in ecology and evolution. Trends Ecol Evol 20:345–353. https://doi.org/10.1016/j.tree.2005.04.004
Purse BV, Hopkins GW, Day KJ, Thompson DJ (2003) Dispersal characteristics and management of a rare damselfly. J Appl Ecol 40:716–728. https://doi.org/10.1046/j.1365-2664.2003.00829.x
R Core Team (2017) R: A language and environment for statistical computing
Rackham O (1986) The history of the countryside: the classic history of Britain’s landscape, flora and fauna. Dent, London
Reunanen P, Fall A, Nikula A (2012) Spatial graphs as templates for habitat networks in boreal landscapes. Biodivers Conserv 21:3569–3584. https://doi.org/10.1007/s10531-012-0382-3
Ribeiro R, Carretero MA, Sillero N et al (2011) The pond network: Can structural connectivity reflect on (amphibian) biodiversity patterns? Landsc Ecol 26:673–682. https://doi.org/10.1007/s10980-011-9592-4
Rich T (1998) A comparison of the ponds in the County of Cardiff with the national statistics from the Lowlands Ponds Survey 1996.
Rouquette JR, Thompson DJ (2007) Patterns of movement and dispersal in an endangered damselfly and the consequences for its management. J Appl Ecol 44:692–701. https://doi.org/10.1111/j.1365-2664.2007.01284.x
Ryszkowshi L, Balazy S (1995) Agricultural Landscapes in Wielkopolska: Threats and Protection. Poznan, Poland
Scheffer M, Van Geest GJ, Zimmer K et al (2006) Small habitat size and isolation can promote species richness: second-order effects on biodi v ersity in shallow lakes and ponds. Oikos 112:227–231
Schilling EG, Loftin CS, Huryn AD (2009) Macroinvertebrates as indicators of fish absence in naturally fishless lakes. Freshw Biol 54:181–202. https://doi.org/10.1111/j.1365-2427.2008.02096.x
Schneeweiss N, Beckmann H (1999) The ponds of the young-moraine-landscape: habitats and centres of distribution of amphibians in Brandenburg (NE-Germany). In: Boothy J (ed) Pond and Pond Landscapes of Europe. Proceedings of the International Conference of the Pond Life Project, 30 August – 2 September 1998. Maastricht, The Netherlands, pp 197–201
Scoccianti C (1999) Loss of ponds in three different areas of Tuscany: conservation plans, actions and restoration projects. In: Boothby J (ed) Pond and Pond Landscapes of Europe. Proceedings of the International Conference of the Pond Life Project, 30 August – 2 September 1998, Maastricht, The Netherlands. Colin Cross Printers Ltd, Garstang, Lancashire, UK, pp 203–210
Segurado P, Branco P, Ferreira MT (2013) Prioritizing restoration of structural connectivity in rivers: A graph based approach. Landsc Ecol 28:1231–1238. https://doi.org/10.1007/s10980-013-9883-z
Service MW (1997) Mosquito (Diptera: Culicidae) dispersal: the long and short of it. J Med Entomol 34:579–588
Smith RF, Alexander LC, Lamp WO (2009) Dispersal by terrestrial stages of stream insects in urban watersheds: a synthesis of current knowledge. J North Am Benthol Soc 28:1022–1037. https://doi.org/10.1899/08-176.1
Sondergaard M, Jeppesen E, Jensen J-P (2005) Pond or lake; does it make any difference? Arch für Hydrobiol 162:143–165
Stauffer D (1987) Introduction to percolation theory. Taylor & Francis, London
Stewart-Koster B, Olden JD, Johnson PTJ (2015) Integrating landscape connectivity and habitat suitability to guide offensive and defensive invasive species management. J Appl Ecol 52:366–378. https://doi.org/10.1111/1365-2664.12395
Sukopp H (1981) Grundwasserabsenkungen. Ursachen u. Auswirkungen auf Natur u. Landschaft Berlins. Wasser S:239–272.
The Wildlife Trust (2000) Species Action Plan: Great crested newt (Tristurus cristatus). In: Biodivers. Action Plan Birmingham Black Ctry. http://www.wildlifetrust.org.uk/urbanwt/ecorecord/bap/html/gcnewt.htm.
Thornhill I (2013) Water quality, biodiversity and ecosystem functioning in ponds across an urban land-use gradient in Birmingham. University of Birmingham, U.K.
Thornhill I, Batty L, Death RG et al (2017) Local and landscape scale determinants of macroinvertebrate assemblages and their conservation value in ponds across an urban land-use gradient. Biodivers Conserv 26:1065–1086. https://doi.org/10.1007/s10531-016-1286-4
Thornhill I, Loiselle S, Lind K, Ophof D (2016) The citizen science opportunity for researchers and agencies. Bioscience 66:720–721. https://doi.org/10.1093/biosci/biw089
Tischendorf L, Fahrig L (2000) On the usage and measurement of landscape connectivity. Oikos 90:7–19. https://doi.org/10.1034/j.1600-0706.2000.900102.x
University of Portsmouth (2017) Birmingham District through time, population statistics, total population. In: A Vis. Britain through time
Urban D, Keitt T (2001) Landscape connectivity: a graph-theoretic perspective. Ecology 82:1205–1218
Urban DL, Minor ES, Treml EA, Robert S (2009) Graph models of habitat mosaics. Ecol Lett 12:260–273. https://doi.org/10.1111/j.1461-0248.2008.01271.x
Van Geest GJ, Roozen FCJM, Coops H et al (2003) Vegetation abundance in lowland flood plain lakes determined by surface area, age and connectivity. Freshw Biol 48:440–454. https://doi.org/10.1046/j.1365-2427.2003.01022.x
Vermonden K, Leuven RSEW, Van Der Velde G et al (2009) Urban drainage systems : An undervalued habitat for aquatic macroinvertebrates. Biol Conserv 142:1105–1115. https://doi.org/10.1016/j.biocon.2009.01.026
Weinreich JA, Musters CJM (1994) The Situation of Nature in the Netherlands. Den Haag, Netherlands
Williams PJ, Biggs J, Barr CJ, et al (1998b) Lowland pond survey 1996. London
Williams PJ, Biggs J, Crowe A, et al (2010) CS Technical Report No. 7/07 Countryside Survey: Ponds Report from 2007. Lancaster
Williams PJ, Biggs J, Whitfield M, et al (1998a) Ancient ponds and modern landscapes.
Williams P, Whitfielda M, Biggs J et al (2004) Comparative biodiversity of rivers, streams, ditches and ponds in an agricultural landscape in Southern England. Biol Conserv 115:329–341. https://doi.org/10.1016/S0006-3207(03)00153-8
With KA (2002) Using Percolation Theory to Assess Landscape Connectivity and Effects of Habitat Fragmentation. In: Applying Landscape Ecology in Biological Conservation, pp 105–130
Wood PJ, Barker S (2000) Old industrial mill ponds: a neglected ecological resource. Appl Geogr 20:65–81. https://doi.org/10.1016/S0143-6228(99)00015-6
Wood PJ, Greenwood MT, Agnew MD (2003) Pond biodiversity and habitat loss in the UK. Area 35:206–216. https://doi.org/10.1111/1475-4762.00249
Wood PJ, Greenwood MT, Barker SA, Gunn J (2001) The effects of amenity management for angling on the conservation value of aquatic invertebrate communities in old industrial ponds. Biol Conserv 102:17–29. https://doi.org/10.1016/S0006-3207(01)00087-8
Acknowledgements
This work was supported by a studentship awarded by Natural Environment Research Council (NERC) and received CASE Studentship support from the Macauley Land-use Research (now The James Hutton Institute). Thanks also go to two anonymous reviewers for their helpful suggestions during the refinement of this manuscript.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
ESM 1
(DOCX 19 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Thornhill, I., Batty, L., Hewitt, M. et al. The application of graph theory and percolation analysis for assessing change in the spatial configuration of pond networks. Urban Ecosyst 21, 213–225 (2018). https://doi.org/10.1007/s11252-017-0724-8
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11252-017-0724-8