
Abstract
The settlement phase is a challenging period for meropelagic octopus, as they adapt to their new life in the sea bottom after a planktonic period. Their ecology and trophic interactions with the surrounding fauna in the wild are practically unknown. To understand their predatory role in the littoral zone, the diet of recently settled Octopus vulgaris from a Mediterranean sandy bottom was studied through molecular methods. Amphipods were present in all the analysed stomach contents, with a total of 20 amphipod taxa recorded as prey. Jassa slatteryi and Microprotopus maculatus were the most commonly found amphipod species. Hydrozoans, decapod crustaceans, cephalopods and bivalves followed amphipods in importance. Niche breadth index assigned to small recently settled octopus the role of a specialised predator, increasing the diversity of prey as they grow. Larger juveniles displayed a higher number of prey taxa suggesting a decreasing predatory specialisation with octopus size and an ontogenetic shift in trophic ecology from diet mainly based on amphipods to a richer diet. The considerable arm elongation and the strong muscular arm crown development after settlement probably allow the juvenile benthic octopus to develop ambush predation and speculative hunting, increasing the range of potential benthic prey species along its adaptation to the benthos.
Graphic abstract

Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The early life of many marine organisms occurs in the water column as planktonic larvae, when they passively dispersed with the movements of the water masses (McManus and Woodson 2012). At the end of this developmental stage, their morphologies change towards their adult forms. Benthic octopods have two development modes: (1) meropelagic, which are temporarily pelagic, producing thousands of eggs with planktonic hatchlings, and (2) holobenthic, that produce a relative smaller number of eggs with direct benthic hatchlings with similar ecology and habitat as their parents (Villanueva and Norman 2008). For meropelagic species (Lincoln et al. 1998), their ecological role is expected to suffer a drastic change, as they adapt from pelagic to benthic environments during a life-challenging step. While there are a number of studies dealing with the planktonic stage of octopus under laboratory conditions (e.g., Iglesias et al. 2014; Dan et al. 2019, 2021a, b; Uriarte et al. 2010), their study in natural environments is challenging, especially when considering advanced paralarval stages (Roura et al. 2019). Besides, few studies had succeeded in obtaining benthic individuals in the lab. The settlement phase is virtually unknown in the wild with the exception of a study on the population dynamics of Octopus bimaculatus in Southern California, where recently settled individuals were collected in kelp holdfasts along the year (Ambrose 1988). In the lab, the settlement of meropelagic octopuses starts when individuals begin to adhere to the tank surfaces with their suckers during increasing periods of time and search for shelters to hide (Villanueva 1995; Dan et al. 2021b; Roura et al. 2023a). During the settlement period, octopus should adapt their morphology to their new life at the sea bottom, which involves quick learning to change their behaviour, such as learning to avoid new predators and adapt to new foraging techniques. They also face important morphological changes, such as the strong positive allometric arm growth, the genesis of new chromatophores, iridophores and leucophores, the development of skin sculptural components and a horizontal pupillary response (Villanueva and Norman 2008; Dan et al. 2021b). They also loss several characters present during their planktonic life, such as the Kölliker organs (Villanueva et al. 2021), the lateral line system and the oral denticles of the beaks (Boletzky 1971; Lenz 1997; Villanueva and Norman 2008).
The common octopus Octopus vulgaris is a species with high commercial value that supports the most intense octopus fishery of the world (Sauer et al. 2021). Octopus vulgaris is a meropelagic species (Lincoln et al. 1998) whose paralarvae develop in the plankton for several weeks, depending on the water temperature (Villanueva et al. 2016). During laboratory rearings at 21 °C, settlement was observed from octopuses aged 47 days (Villanueva 1995; Iglesias and Fuentes 2014) with a transitional period to a full benthic life of around 10 days (Villanueva and Norman 2008). Studies on the development and feeding of O. vulgaris paralarvae in the wild show that the paralarvae are active predators of zooplankton, predating mainly on larvae of decapod crustaceans, and also feeding on other groups such as molluscs, ophiuroids, amphipods, cladocerans, copepods, chaetognaths and cnidarians (Roura et al. 2012, 2016, 2017; Olmos-Pérez et al. 2017). After settlement, benthic juvenile and subadult O. vulgaris individuals were collected during summer in the Azores Archipelago using PVC pipes with different diameters offered as shelter, collecting more individuals in sandy areas in comparison with rocky bottoms (Pham and Isidro 2010). Subadult and adult O. vulgaris diet is composed of a wide range of prey that reflect its opportunistic and plastic predatory behaviour, mostly composed of crustaceans, molluscs and teleost fish (see among others, Guerra 1978, b; Sánchez and Obarti 1993; Quetglas et al. 1998; Rosa et al. 2024).
Despite the fact that feeding habits of subadult and adult cephalopods in the wild are relatively well known in several species (e.g., Nixon 1987; Ibáñez et al. 2021; Ribas et al. 2021), including metabarcoding DNA studies of stomach contents in subadult and adult octopus (Ramos et al. 2023), the diet of early stages in the wild is far more mysterious. This is due to the small size of early ontogenetic stages and to the high degree of prey fragmentation. The use of DNA metabarcoding techniques is a good method for overcome this problem, as it allows the identification of digested unrecognizable food (e.g., Olmos-Pérez et al. 2017; Fernández-Álvarez et al. 2018; García-Mayoral et al. 2022). Contrasting with its ecological and ontogenetic importance, the diet of newly settled octopus is still unknown for any octopod species. The objective of the present study is to fill this knowledge gap, which is paramount for understanding the challenging transition from planktonic to benthic life in meropelagic octopus.
Material and methods
Sample collection and laboratory procedures
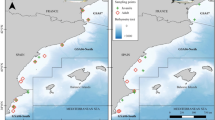
A total of 166 individuals of O. vulgaris collected between March 2020 and March 2021 through the artisanal fishing fleet of Vilanova i la Geltrú (NW Mediterranean, Fig. 1), during 52 sampling days, were used to extract the stomach contents. Stomach contents of individuals collected during spring (47%), summer (14%), autumn (27%) and winter (10%) were examined. The sampling was carried out between 05:00 and 11:00 h, on the sandy bottom of an artisanal octopus fishery ground and between 5 and 25 m depth. Individuals were obtained using experimental juvenile octopus collectors (Escolar et al. 2021) (JOC, n = 80), cuttlefish (Sepia officinalis) basket traps (CT, n = 68) and octopus (O. vulgaris) traps (OT, n = 10) and octopus pots (OP, n = 8). After collection, individuals were killed on board with an overdose of anaesthesia (5% MgCl2) following welfare recommendations and guidelines for cephalopods (Fiorito et al. 2015), and then stored at – 20 °C until lab processing.

Map of sample area in Vilanova i la Geltrú (Latitude: 41.2136611, Longitude: 1.736531) (A). Recently settled common octopus Octopus vulgaris of 327 mg in weight and 11.02 mm dorsal mantle length photographed in laboratory after collection (B)
In the laboratory, individuals were weighed after thawing using a microbalance. The stomach contents of a total of 166 individuals were extracted and visually assessed during dissections under a binocular stereoscope Leica S8AP0 (Leica Microsystems, Wetzlar, Germany). Individuals with scarce stomach content were excluded. For DNA analyses, 91 octopuses were selected. Selection of individuals was focused on the smaller size-class, looking for the more recently settled individuals. Moreover, different sampling gears were utilised in order to lower sampling bias of specific gears. Mean weight of the selected individuals was 3.55 ± 6.44 g, ranging from 0.17 to 37.45 g. Individuals were not sexed.
After finishing this sampling protocol and during a recent related study also on juvenile octopus, we have observed that 5% MgCl2 anaesthesia during 30 min used to euthanize the individuals, significantly reduced the weight of the juveniles. As far as we know, this is a previously unrecorded effect of MgCl2 in cephalopods. For this reason, the fresh, pre-anaesthesia octopus weight of the individuals used during the present study was estimated later. We obtained 35 additional O.vulgaris individuals 0.7 ± 1 g ranging from 0.2 to 4.4 g in weight, collected from the same sampling area during December 2023. Animals were weighted before and after 5% MgCl2 anaesthesia during 30 min, then frozen at -20 °C and thawed after a few days to obtain their thawed weight, following a similar protocol as the individuals used for stomach analysis. The loss of weight due to 30 min in 5% MgCl2 was 18 ± 5%, ranging from 8 to 34%. It was observed that weight loss was inversely related to the juvenile weight, being higher in smaller individuals (Online resource 1). Therefore, the estimation of the corresponding pre-anesthesia weight was carried out according to the different octopus size groups (see Table 1). For the largest group of juveniles (size class 5, see below), it was not possible to estimate their pre-anesthesia weight due to the lack of large individuals to test the degree of weight loss. When comparing post-anaesthesia and thawed weight, no significant differences were obtained (Student t-test, p = 0.96).
To prepare the samples for the subsequent molecular analysis, special attention was paid to avoid ambient and cross contamination. In short, stomach contents were extracted carefully with sterilised tools. The sterilisation process consisted in several cleaning steps: (1) tools were cleaned with jets of bleach at 10% diluted in milli-Q water, (2) then with jets of milli-Q water, and (3) finally with jets of ethanol at 96%. After that, each surface of every tool was carefully treated with ultraviolet light for at least 20–30 min under the fume hood. After sterilisation, sampling tools were transported and kept carefully inside a plastic box also sterilised with the same methodology. The extraction of the stomach content of the individuals was executed inside a sterilised work area, new sterile latex gloves were used for each individual and sterile lab material was individually used for each dissection. Stomach contents were preserved in 96% ethanol and stored at – 20 °C until DNA extractions.
Blocking primer design and testing through mock communities of prey and stomach dissections
Co-amplification of predator DNA together with the prey is the main concern in dietary DNA metabarcoding studies, known as self-contamination (Fernández-Álvarez et al. 2018). To avoid the amplification of host cephalopod DNA in the stomach content analysis, a pilot analysis was carried out to test and design a blocking primer. The blocking primer (cytochrome c oxidase subunit 1 (COI)-intF_OctopusBP 5′ CGT TTA CCC RCC TCT TTC AAG AAA TTT AGC TC 3′) was produced following the methodology described by Vestheim and Jarman (2008). It was designed with Geneious 10.2.6 (Biomatters Ltd), based on its complementarity to O. vulgaris COI sequences downloaded from GenBank. A C3 CPG spacer was added to the 3’ end of the blocking primer to prevent elongation.
Mesofauna (size range 0.2–20 mm) from the sampled area was collected through juvenile octopus collectors, cuttlefish traps and octopus traps. The different sampling gears were sampled using a high-pressure seawater flux inside a 100 L plastic bucket with a 200 μm plankton net surrounding the bucket. The collected mesofauna was preserved at – 20 °C. After thawing in the laboratory, mesofauna was separated and identified to the lowest taxonomic level using taxonomic keys (Zariquiey 1968; Campbell 1984; Riedl 1986; Ortiz and Jimeno 2001; Handoo and Hunt 2016). Five pools with a total of 70 different potential prey species (Decapoda, Amphipoda, Isopoda, Leptostraca, Mysidacea, Gastropoda, Bivalvia and Cephalopoda) were used to test the effect of the blocking primers. DNA was extracted from each of the five pools following the same methodology as for the stomach content samples (see below).
A mix of DNA from the five pools of potential prey was used to develop a potential prey DNA solution, which was mixed with different amounts of O. vulgaris DNA for constructing the mock communities. Two mock communities were designed with different proportions of predator–prey DNA solution as follows: 50:50 and 90:10, respectively. Three recently settled octopus stomach samples and two mock communities were used to prepare four different treatments per sample by alternating the presence/absence of blocking primer, and using fixed (55 °C) or variable (60 and 55 °C) annealing temperature.
DNA metabarcoding
DNA was isolated from the samples using two different extraction kits according to sample size: the Quick-DNA Microprep Plus Kit (Zymo Research) was used for the smaller samples and the E.Z.N.A Tissue DNA kit (Omega Biotek) was used for the larger ones. Both kits were used following the manufacturer’s instructions and the DNA was resuspended in a final volume of 10 or 100 µL for the small and large samples, respectively. An extraction blank was included in every DNA extraction round and treated as a regular sample to check for cross-contamination. DNA was quantified using the Qubit High Sensitivity dsDNA Assay (Thermo Fisher Scientific).
In the pilot study, the addition of blocking primers and a fixed annealing temperature provided better results (Online Resource 2). Thus, these conditions were applied to the polymerase chain reaction (PCR) of the stomach content DNA extractions. A 313-base pair (bp) fragment of the mitochondrial gene COI was amplified using the primer pair COI-intF (5′ GGW ACW GGW TGA ACW GTW TAY CCY CC 3′) (Leray et al. 2013) and HCO2198 (5′ TAA ACT TCA GGG TGA CCA AAA AAT CA 3′) (Folmer et al. 1994). PCRs were carried out in a final volume of 12.5 µL, containing 1.25 µL of template DNA, 0.5 µM of each primer, 10 µM of the blocking primer, 6.25 µL of Supreme NZYTaq 2 × Green Master Mix (NZYTech), and ultrapure distilled water up to 12.5 µL. The reaction mixture was incubated as follows: an initial denaturation step at 95 °C for 5 min, followed by 35 cycles of 95 °C for 30 s, 55 °C for 45 s, 72 °C for 45 s, and a final extension step at 72 °C for 7 min. Illumina sequencing indexes were attached to these amplicons at their 5′ ends in 5 cycles following the same PCR conditions but with an annealing temperature of 60 °C.
De novo chimaera detection was performed using the UCHIME algorithm (Edgar et al. 2011) implemented in VSEARCH (Rognes et al. 2016). Sequences were clustered with the SWARM algorithm (Mahé et al. 2015) with a value of 13 following Bakker et al. (2019). The taxonomic identification was performed by querying the sequences resulting from clustering against the COI gene database build by Robeson et al. (2020) (updated on July 2020) based on the Barcode of Life Data System (BOLD) library reference database (Ratnasingham and Hebert 2007). To do so, the script feature-classifier classify-consensus-vsearch implemented in Qiime2 (Bokulich et al. 2018) and the VSEARCH algorithm with a sequence similarity threshold of 90% were used. Singletons (OTUs containing a single sequence in the whole dataset) were excluded from the OTU table. In DNA metabarcoding studies, the mistagging phenomenon has been reported (Esling et al. 2015, Bartram et al. 2016) in which a low percentage of the reads of a sample can be assigned to another as the result of the misassignment of the indices during library preparation, sequencing, and/or demultiplexing steps. To account for this error, sequences with less of 0.01% frequency in each sample were removed. Additional filters were used based on the taxonomic assignment: the Operational Taxonomic Units (OTUs) assigned to Mammalia and Fungi were removed from the OTU table, whereas the unidentified OTUs were kept for downstream analyses. Identification of those unassigned OTUs was performed through BLAST (Altschul et al. 1990) at the NCBI portal. Identification was considered successful only if the following parameters were fulfilled: percent identity of 90%, e-value of 1e-05, and a minimum hit coverage of 80%. The similarity threshold of 97% or higher was set for specific identifications, and between 90 and 97% for identifications at the family level. When BLAST hits were sequences identified as more than a single species or belonged to different taxonomic groups, they were assigned to the lowest taxonomic level shared among the original identifications. As PCR produces amplification bias that will skew abundance estimation (O’Rorke et al. 2012), the number of reads in the results of the genetic analysis has not been taken into account for downstream analyses and all inferences are based on presence/absence of prey OTUs.
Data and statistical analyses
Octopus were arranged in five groups according to their weight to assess possible size-related differences in feeding (Table 1). Trophic ecology data were analysed through several indexes. The Emptiness Index (EMI = Es/Ts × 100) evaluates the number of empty stomachs (Es) in relation to the total number of analysed stomachs (Ts). The taxa groups of Chromista, Fungi, Protozoa, Plantae, Insecta and Arachnida were excluded as prey intentionally ingested/selected for downstream analyses. The relative importance of a particular prey was studied through the frequency of occurrence index (FO = Si/Ts × 100), where Si is the number of octopus stomachs in which the prey was registered. Samples whose rarefaction curves did not tend towards saturation (n = 4), were excluded to statistical analyses and Cardona’s index estimation to avoid errors in the comparisons between groups. Cardona’s index (B’ = (SFO-s/100 × R) was used to estimate the trophic niche breadth using the frequency of occurrence data, where s represents the standard deviation of occurrence frequencies and R is the number of prey-species used by the whole guild (Cardona 1991). This index assesses the degree of specialisation with values ranging from 0.01 (highly specialist) to 1.0 (complete generalist).
Statistical analyses were carried out with the software PERMANOVA+ for PRIMER 6 (Anderson et al. 2008). To evaluate the data without taking into account the number of reads as abundance, the number of reads of each OTU corresponding to a prey category per sample was transformed to create a presence/absence matrix (i.e., “0” was considered as absence of a prey category, and any number of reads codified as “1” and considered as presence of that prey category). After the transformation of reads to a presence/absence matrix, data were standardised by the total of samples, and then a resemblance matrix using the Jaccard Index was created. For comparison, semiquantitative data was standardised, square root transformed and a resemblance matrix using the Bray–Curtis similarity method was applied. Non-metric multidimensional scaling (MDS) (Clarke and Warwick 2005) analyses with the superimposition of Cluster were applied. The MDS were represented by single samples, factors groups and by the average of size class and sampling gear groups combined (Online Resource 2). A permutational multivariate analysis of variance (PERMANOVA) with a Type III sum of squares (unrestricted permutation of raw data = 999) was used to test for significant dietary differences among size class, sampling gears and season. Additionally, a similarity percentage analysis (SIMPER) detected the intra-group percentage of mean diet dissimilarity for each size class, gear and season considering prey contribution of over 75%.
Results
A total of 121 out 166 dissected O. vulgaris showed food remains in their stomach content through visual assessment, resulting in an emptiness index (EMI) of 27%. A total of 91 octopuses were selected for DNA metabarcoding analysis, ranging in weight from 0.17 to 37.45 g. Particular effort was made to select the smaller size class (n = 32) individuals between 0.17 and 0.5 g, which are the more recently settled individuals.
Blocking primer performance
No substantial effect was detected in read counts using fixed or variable annealing temperature. There were not significative difference on the number of reads induced by the use or not of the blocking primers on octopus' prey, both in the mock communities (Student t-test, p > 0.64) and the stomach contents (p > 0.53). The presence of host (O. vulgaris) reads was markedly higher in PCRs without the blocking primer (Online Resource 3). For the mock communities in PCRs with blocking primers, O. vulgaris reads represented 0% of the total amount of reads for the mock community proportion 50:50, and 0.03% for 90:10. The PCRs without blocking primers showed larger proportions of O. vulgaris reads: 1.27–1.35% for the mock community 50:50 and 10.6–13.6% for the 90:10. For the recently settled octopus stomach content samples, host reads represented 0.0–0.3% with blocking primers and 2.4–39.7% without blocking primers.
DNA metabarcoding and prey profiles
A total of 5,399,108 reads were obtained, resulting in a mean of 59,330.9 ± 30,561.1 (range 889–125,387) reads per sample. From those, 4,304,850 reads and a mean of 47,306 ± 33,095 (range 5–120,799) reads per sample were identified, and a total of 1,050,915 and a mean of 11,548.5 ± 17,428.3 (range 0–72,721) remained taxonomically unassigned. DNA metabarcoding revealed a total of 352 OTUs on O. vulgaris stomach contents (Online Resource 4). Initially, with the use of the BOLD database in Qiime2 a total of 108 assigned and 260 unassigned taxa were classified. After subsequent BLAST searches at NCBI, data treatment and joining the reads belonging to the same taxa but over-splitted in several OTUs, the number of identified OTUs increased to 139, while 213 remained unassigned. Identified OTUs belonged to 18 phyla, 57 orders and 95 families. Mean self-contamination was 2.2 ± 8.7% (0–62.9%) of the reads. The number of OTUs present per stomach content, including several unassigned species, was 12.1 ± 8 (range: 3–54). The number of identified taxa present per stomach content was 6.2 ± 3.2 (range: 1–16). Species richness increased with the octopus size class (Table 1).
Among the identified taxa present in the stomach contents, the group Amphipoda was the order with the highest frequency of occurrence, which was present in all the analysed stomach contents (FO = 100%). Other prey groups by FO rank were: Hydrozoa (46%), Decapoda (31%), Ochrophyta (22%), Bivalvia (22%), Copepoda (13%), Gastropoda (12%), Sepiida (12%) and Chordata (11%). The rest of groups with FO > 2% are represented in Fig. 2.

Frequency of Occurrence (FO, %) and number of identified OTUs (orange line) in the stomach contents of O. vulgaris (n = 91). Groups with FO > 2% are shown. Chromista, Fungi, Protozoa, Plantae, Insecta, and Arachnida taxa were excluded. Recently settled common octopus Octopus vulgaris of 327 mg in weight and 11.02 mm dorsal mantle length photographed in laboratory after collection. Silhouette images from https://www.phylopic.org/
The species with higher FO were the amphipods Jassa slatteryi (90%), Microprotopus maculatus (49%), Amphipoda sp. (35%), Gammarus crinicornis (34%), Phtisica marina (25%) and Caprella scaura (23%). These six identified amphipods were six of the top seven OTUs with higher FO in the analysed stomach contents. The third taxon with the highest FO (47%) was not identified with the BOLD and NCBI databases. The identity of this species remained unassigned, as there were no DNA barcodes available. The next species with higher FO were from group Hydrozoa, Obelia dichotoma (22%), Bougainvillia muscus (20%) and Obelia geniculata (14%). Three species had the same FO of 12%: the cephalopod Sepia officinalis, the amphipod Leucothoe incisa and the clam Mactra stultorum. Interestingly, M. stultorum showed a remarkable increment in its FO from 0% by the smaller octopus size class 1 (0.2–0.4 g) to 41.7% with the larger size class 5 (9.1–37.5 g) (Table 2).
Decapod crustaceans were the following group in importance as prey of FO: Philocheras bispinosus (10%), Athanas nitescens (9%), Liocarcinus vernalis (9%) and Macropodia rostrata (5%). The species P. bispinosus and A. nitescens were more frequent in the stomach content of larger individuals from the size classes 4 (2.7–6.4 g) and 5 (9.1–37.5 g) (Table 2). Hydrozoans were more frequently found in the stomachs than decapods. However, the number of identified hydrozoans (n = 11) was lower than decapods (n = 15).
Octopus vulgaris juveniles also preyed on other cephalopods apart from Sepia officinalis, such as Sepiola affinis (4%), and exceptionally the small-spot octopus Callistoctopus macropus (1%). The class Bivalvia was represented mainly by Mactra stultorum (12%) and Chamelea gallina (5%). The presence of fish was low, with only five identified species: Sparus aurata (3%), Mullus barbatus (3%), Conger conger (2%), Spondyliosoma cantharus (1%) and Scyliorhinus canicula (1%).
There were several unassigned OTUs with important values of FO (range 1–47%) that might represent an important role in the diet of the benthic transition, but its identification was not possible at present (Table 3).
The amphipod Jassa slatteryi was the species with greater contribution for defining each group (size class, gear and season) as it represents from 40 to 100% of the contribution of similarity in the SIMPER analysis. Table 4 summarises the significant differences among size class, collector gear and season groups (P(Perm) < 0.05) using a PERMANOVA pairwise tests analysis. The MDS analysis showed for the assessment of single samples a value of two-dimensions stress of 0.22 with presence/absence assessment, and 0.21 with the use of semiquantitative data. In the case of the factors size class and collector gear combined, the value of two-dimensions stress was 0.19 with presence/absence assessment, and 0.18 with the use of semiquantitative data treatment (Online Resource 2). Respective values of two-dimensions stress near to 0.2 provided a poor representation that should not be placed on the detail of the plot (Clarke and Warwick 2001). Thus, differences in stress values are minimal between presence/absence and semiquantitative treatments.
As shown in Table 5, the factors size class, gear and season showed a significant difference (p < 0.05) with separated PERMANOVA analysis designs.
The assessment of the trophic niche breadth of all 87 samples through Cardona’s index showed a value B’ = 0.05, which is indicative of specialised predation. A weak trend towards a more generalist diet was observed when data were studied separately by octopus size classes: B’ size class groups 1 (0.2–0.4 g), 2 (0.5–0.9 g) and 4 (2.7–6.4 g) had a slightly more specialised index (B’ = 0.05) than the other size groups. The size class 3 (1.0–2.4 g) showed a value of B’ = 0.06, while the largest size group 5 (9.1–37.5 g) displayed the highest value of B’ = 0.07.
Discussion
Most studies on cephalopod trophism are focused on adults and subadults, usually showing emptiness indexes higher than 45% that could be affected by variables such as temperature, season, sex and food availability, among others (see Ibáñez et al. 2021). Here, a lower emptiness index value of 27% was found during the present study, indicating that roughly only one quarter of the stomach contents visually examined in recently settled O. vulgaris did not have food remains. In subadult and adult O. vulgaris also visually assessed, the EMI values range from 45 to more than 70% (Sánchez and Obarti 1993; Diatta et al. 2001; Rosa et al. 2004; Ajana et al. 2018). The higher presence of stomach with food contents of the juveniles examined in comparison with the adults from previous studies could be related with several undetermined factors. However, this fact suggests that juvenile octopus maintain a notable predatory activity.
A total of 131 taxa belonging to 18 phyla, 57 orders and 95 families were molecularly identified. When Chromista, Fungi, Protozoa, Plantae, Insecta and Arachnida were excluded as plausible primary predation, the species richness (S) of potential diet in O. vulgaris juveniles reached S = 96 (Online resource 6). This value represents a higher number of taxa compared with previous studies from the same species based on DNA metabarcoding of paralarval diet, obtaining 54 (Olmos-Pérez et al. 2017) and 87 prey species (Roura et al. 2023b). Furthermore, results from visual analyses of subadult and adult individuals obtained lower richness values of S = 35 (Guerra 1978, b) and S = 23 (Rosa et al. 2004). However, it must be pointed out that molecular and visual assessments of prey are not directly comparable. Prey species richness by octopus juveniles ranged from 1 to 16 prey with a mean of 6 prey. Mean prey S increased with size class (Table 1). The high number of identified prey suggests that recently settled and early benthic octopus could predate on a wide spectrum of animals.
Amphipods as main prey for O. vulgaris at settlement and other secondary prey groups
The most important prey of O. vulgaris juveniles was a Mediterranean invasive amphipod species, Jassa slatteryi, present in 90% of the samples. It is an epibenthic species of intertidal vegetated habitats, living at depths lower than 30 m (Jeong et al. 2006). Conlan et al. (2021) pointed out that some species of Jassa had been transported by human vectors since at least the nineteenth century, which explains their wide distribution across the world. Amphipods are primary consumers, eating detritus and phytoplankton, and are in turn preyed by higher trophic level consumers, such as fish and large crustaceans (Jeong et al. 2006; Dauvin 2024). Jassa species has been used as successful prey for paralarval rearings of the octopus Robsonella fontaniana (González et al. 2011), O. vulgaris (Tur et al. 2020; Varó et al. 2022), hatchlings of the cuttlefish Sepia officinalis (Baeza-Rojano et al. 2010), and other marine gammarid amphipods showed to be suitable prey for Octopus maya hatchlings (Baeza-Rojano et al. 2013). The present study on the diet of wild juvenile octopus confirms amphipods as a food resource to develop laboratory rearing experiments on early stages of octopus. Different technologies for amphipod production have been published (see between others, Fernández-González et al. 2018; Jiménez-Prada et al. 2021; Vargas-Abúndez et al. 2021; Shahin et al. 2023) and probably can be adapted for octopus. The amphipod species Caprella scaura, the sixth species with more FO, is also considered a Mediterranean invasive species, first recorded in Roses Bay in 2005, 169 km north to the sampled area (Martínez & Adarraga 2008). The predation of juvenile octopus on invasive species is of interest, opening the role of recently settled octopus as a native controller on two Mediterranean invasive species. This fact furthers into feeding plasticity of octopus (e.g., Fernández-Álvarez et al. 2020), which are able to adapt to the use of new feeding resources available in the area.
The class Hydrozoa is the second group in terms of FO. It cannot be ascertained if the consumption of hydrozoans occurred over polyp or medusa stages. This would be an important difference, as the polyp stage is sessile and probably easier to catch than the swimming medusa stage. Obelia geniculata was also found in the diet of O. vulgaris and Alloteuthis media planktonic paralarvae diet study by Olmos-Pérez et al. (2017). Also, Obelia dichotoma was found in the diet of O. vulgaris paralarvae by Roura et al. (2023b). Surprisingly, present data suggest that hydrozoans constitute also an important feeding resource for recently settled octopus.
Eggs of the European cuttlefish Sepia officinalis are normally found in the fishing gears, which are a hard substrate where mature females lay eggs (Barrett et al. 2022). As cuttlefish egg capsule protect the embryo and its predation is usually limited by the mechanical power of the predator capture tools (Martins et al. 2018), it is likely octopus predated on hatchlings instead. Furthermore, analysing differences by octopus size classes, the bivalves were preyed preferably by larger juveniles. Thereby, M. stultorum increased its occurrence in the stomach contents of larger octopus and C. gallina and S. subtruncata were also more consumed for members of the largest octopus size class 5 (9.1–37.5 g) (Table 2). Octopus can consume bivalves through two possible methods: (1) drilling a hole on the shell then injecting cephalotoxin or (2) by pulling the shells using the strength of their suckers and arms (Fiorito and Gherardi 1999). The consumption of clams (Ruditapes phillipinarum) by Octopus sinensis juveniles under laboratory conditions increased gradually with octopus size and were not drilled (Dan et al. 2021b). The fact that bivalves were more abundant in stomach contents of the largest size class of O. vulgaris juveniles confirms the observations of Dan et al. (2021b), and are probably related with the rapid development of the arm crown in larger juveniles reaching enough arm strength to pull bivalve shells apart.
Decapods were the most consumed prey by O. vulgaris paralarvae in the water column (Olmos-Pérez et al. 2017; Roura et al. 2023b). In juveniles, this prey group increased their presence in the stomach contents of larger juveniles, as for P. bispinosus and A. nitescens, mostly predated by the largest juveniles from size class 5 (9.1–37.5 g). Furthermore, other Decapoda with an important FO were not consumed by the group of the recently settled size class 1 (0.2–0.4 g) (Table 2). These differences in FO among size class groups could be explained by the strong morphological development along the settlement period. Morphological adaptations as arm elongation would further enhance new hunting techniques, such as ambush predation and speculative hunting on the seafloor, predatory behaviours well known in subadults and adults (Hanlon and Messenger 2018) that may need a learning or adapting period after settlement in the new benthic environment (Fig. 3). Fish were represented by four species, with a maximum FO of 3%. Surprisingly, it included the small-spotted catshark Scyliorhinus canicula. This shark inhabits normally deeper waters and its size at hatching is far larger than O. vulgaris juveniles. Probably, this prey item might actually represent carrion consumption, likely coming from trawl fishery discards or artisanal fishery bait, which is used in the same fishery ground and/or secondary predation mediated by amphipods. Subadult and adult octopuses are known to be scavengers, as they are attracted by the dead bait inside traps (Sauer et al. 2021) or feeded by frozen prey and/or artificial food in the lab (Villanueva et al. 2017), suggesting that scavenger behaviour of juvenile octopuses in the wild also can be expected.

Schematic representation of prey diversity (black silouettes) of Octopus vulgaris along its life cycle. During planktonic stage most frequently detected prey are decapod zoea, followed by calanoid copepods, brittle stars and cladocerans (Olmos-Pérez et al. 2017; Roura et al. 2023a, b). After settlement, during juvenile stage amphipods are the prey with higher frequency of occurrence, followed by hydrozoans, decapod crustaceans and bivalves (present study). Subadult and adult stages prey mainly on crabs, molluscs and teleost fishes (Guerra 1978, b; Sánchez and Obarti 1993; Quetglas et al. 1998; Rosa et al. 2024), while brooding females stop feeding after spawning. Silhouette images from https://www.phylopic.org/. Central image of adult octopus by Ricardo Roberto Fernández
Benefits of the use of blocking primers
The use of blocking primers achieved an exceptionally low value of self-contamination (2.2 ± 8.7%), which contrast with the comparatively higher values (70–95%) commonly observed in other cephalopod molecular dietary studies (e.g., Olmos-Pérez et al. 2017; Fernández-Álvarez et al. 2018; García-Mayoral et al. 2022; Roura et al. 2023b). It is remarkable that the values of self-contamination obtained in this study before the use of blocking primers (Online Resource 2) were lower than the values observed in previous studies. Importantly, we were not able to identify any amplifying bias induced by the blocking primers on octopus prey, both in the mock communities and the stomach contents. While careful dissections are able to lower self-contamination values (e.g., Fernández-Álvarez et al. 2018), our results indicate that the use of blocking primers are especially useful to obtain better dietary assessments, especially for tiny animals difficult to dissect as paralarvae. However, blocking primers should be species-specific and therefore multispecies projects would need to use adequate species-specific primers for each predator, or assume possible amplification bias if a single blocking primer is designed for the whole set of predators.
Limitations of the study
As DNA metabarcoding involves a PCR, one of the main biases that occurs is the primers not hybridising with the whole spectrum of actual prey. Besides, they also might create amplification bias among the DNA of different species within the sample, as the primers might be more efficient amplifying the DNA of some species than others. Additionally, the high precision of recognition of DNA metabarcoding evokes the possibility that some identified taxa could have originated from secondary predation (Ibáñez et al. 2021, as “secondary digestion”), which means that the taxon was actually predated by the prey instead from a direct predation event from the predator. Moreover, other organisms living in association with the prey, such as epiphytes, symbionts or parasites, can also be sequenced. It is important to note that Clarke et al. (2020) showed that secondary digestion could be detected in fish diet samples, but the relative read abundance of these items would rarely be greater than 1%. Importantly, Nigmatullin et al. (2009) interpreted the presence of secondary digestion in 43.3% of the stomach content analysis from the neon flying squid Ommastrephes bartramii using morphological assessment methods. Fernández-Álvarez et al. (2018) identified 42% of reads of secondary digested prokaryotic algae and chloroplasts in subadult and adult flying squids, while this component was minimal in paralarvae. Moreover, it is expected that digestion time would be longer in species with hard structures. Thus, crustaceans should have longer digestion times than hydrozoans, for instance. This fact could provoke differences between the frequency of occurrence appeared with the analysis and their real contribution to the diet. The 91 analysed stomach contents were all from juveniles collected in a shallow (5–25 m depth) Mediterranean sandy bottom, during daytime, and mostly collected during spring and autumn. This environment and sampling characteristics obviously conditioned the results. It is possible that octopus juveniles settling in rocky beds might prey on different organisms, as the benthic communities would definitely differ. Future studies taking into account different environments and seasons will be necessary to understand the feeding dynamics of recently settled octopus.
In DNA metabarcoding studies, DNA for the host species is commonly regarded as self-contamination, but it can also be explained by cannibalism. The use of blocking primers for O. vulgaris prevents the possibility to assess cannibalism. According to Dan et al. (2021b), the incidence of cannibalism in O. sinensis under laboratory conditions intensifies after settlement in aquaria, and cannibalism has been reported in wild populations of octopus (Ibáñez and Keyl 2010). Thus, the existence of a cannibalistic component in the diet of O. vulgaris juveniles was not assessed here. Molecular methods can be used for cannibalism assessment, as it was studied in lionfish through microsatellites (Dahl et al. 2018).
Recently settled octopus as a specialist predator
During settlement, the young octopus probably lives a period of learning-dependent foraging and discovering and tasting new prey species. The movements and permanence in a specific area could be selected based on the quantity or quality of the available prey. According to Ibáñez et al. (2021), many studies confuse the terms opportunistic vs. generalist predator and selective vs. specialist predator. These authors also noted that although most cephalopods exhibit a generalised diet and wide trophic niche, their hunting strategy is selective and specialised at the individual level. During the present study, the value of Cardona’s index was low (B’ = 0.05). However, B’ increased slightly from 0.05 to 0.07 along octopus growth. These results show that the diet of the recently settled and early benthic O. vulgaris is typical of a specialist predator, strongly focused in amphipods, and that this specialised dietary habits blur with the increase in size. However, a mean of six identified prey or twelve OTUs and a range between 1 and 16 identified species or between 3 and 54 OTUs were found inside the stomachs, suggesting that the juveniles feed on a wide diversity of prey.
Anderson et al. (2008) obtained values very similar for adult Octopus americanus (identified as O. vulgaris) in the southern Caribbean with B’ of 0.08 analysing the middens of 38 octopus dens. A similar relationship was obtained by Alejo-Plata et al. (2018) for Octopus hubbsorum from NE Pacific, obtaining B' of 0.03 for small octopuses (40–80 mm ML) and B' of 0.09 for larger octopuses (> 80 mm ML). A slightly high index (B’ = 0.24) was observed for Octopus rubescens found in beer bottles on a sand/muddy habitat in the NE Pacific (Anderson et al. 1999) and a B’ of 0.28 was found for O. cyanea in Hawaii (Mather, unpublished data, cited in Anderson et al. 2008).
Some studies considered the planktonic stage of O. vulgaris paralarvae as a generalist predator, and others, as a specialist during their first days of life in the pelagic realm based on the prey species detected and their relative abundances in the zooplankton (Roura et al. 2012, 2023b; Nande et al. 2017; Olmos-Pérez et al. 2017). Nevertheless, several studies defined octopus as generalist feeders and Hanlon & Messenger (2018) assigned the term ‘opportunistic feeders’. However, there were some individuals that were specialising on certain taxa or even single species (Anderson et al. 2008). The results found in our study on a sandy bottom area showed recently settled octopus as specialist predators mainly focused on amphipods.
Conclusions
Amphipods are the prey with higher frequency of occurrence in the stomach contents of the recently settled and early benthic individuals of O. vulgaris in a littoral sandy bottom of the Mediterranean Sea. A total of 20 amphipod taxa were present in the analysed stomach contents and this group seems to play a key role as a feeding resource during the octopus settlement period in the studied area. Jassa slatteryi and Microprotopus maculatus are the amphipod species with higher occurrence, present in 90% and 50% of the stomach contents, respectively. As J. slatteryi (1st FO) and C. scaura (6th FO) are invasive species, it is possible that recently settled octopus are controllers of these non-native species. Hydrozoans, decapods, S. officinalis and M. stultorum were also important prey. Niche breadth index assigned to recently settled octopus the role of a specialised predator, increasing their prey diversity along the juvenile benthic growth. The analysed larger juvenile size class has a higher diversity of prey, suggesting a decreasing predatory specialisation. Importantly, the presence of bivalves and decapods increased with the weight of the juvenile octopuses and suggests that the strong muscular development of the arm and suckers after settlement provides the juveniles with the skills to increase their trophic niche breadth along its adaptation to the benthos, where they will remain for the rest of their lives. Less than one third of the stomach contents examined were empty, suggesting that juvenile octopus maintain a notable predatory activity.
Data availability
Genetic data underlying this article are available in the GenBank Nucleotide Database at https://www.ncbi.nlm.nih.gov/genbank/ and can be accessed with the GenBank accession numbers OR131649–OR131744. The FastQ files can be accessed within the GenBank Nucleotide Database with the BioProject accession number PRJNA984187. Additional material is available in Online Resources.
References
Ajana R, Techetach M, Saoud Y (2018) Diet of Octopus vulgaris from the Moroccan Mediterranean Coast. Thalassas 34:415–420. https://doi.org/10.1007/s41208-018-0084-z
Alejo-Plata MC, Ahumada-Sempoal MA, Guzmán SSL, Herrera-Galindo JE, García-Madrigal MS (2018) Diet of Octopus hubbsorum (Cephalopoda: Octopodidae) from the Coast of Oaxaca, Mexico. Am Malacol Bull 36:109–118. https://doi.org/10.4003/006.036.0111
Ambrose RF (1988) Population dynamics of Octopus bimaculatus: influence of life history patterns, synchronous reproduction and recruitment. Malacologia 29:23–39
Anderson RC, Hughes PD, Mather JA, Steele CW (1999) Determination of the diet of Octopus rubescens through examination of its beer bottle dens in Puget Sound. Malacologia 41:455–460
Anderson RC, Wood JB, Mather JA (2008) Octopus vulgaris in the Caribbean is a specializing generalist. Mar Ecol Prog Ser 371:199–202
Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ (1990) Basic local alignment search tool. J Mol Biol 215:403–410
Baeza-Rojano E, García S, Garrido D, Guerra-García JM, Domingues P (2010) Use of amphipods as alternative prey to culture cuttlefish (Sepia officinalis) hatchlings. Aquaculture 300:243–246
Baeza-Rojano E, Domingues P, Guerra-García JM, Capella S, Noreña-Barroso E, Caamal-Monsreal C, Rosas C (2013) Marine gammarids (Crustacea: Amphipoda): a new live prey to culture Octopus maya hatchlings. Aquac Res 44:1602–1612. https://doi.org/10.1111/j.1365-2109.2012.03169.x
Bakker J, Wangensteen O, Baillie C, Buddo D, Chapman D, Gallagher A, Guttridge T, Hertler H, Mariani S (2019) Biodiversity assessment of tropical shelf eukaryotic communities via pelagic eDNA metabarcoding. Ecol Evol 9:14341–14355. https://doi.org/10.1002/ece3.5871
Barrett CJ, Bensbai J, Broadhurst MK et al (2022) Cuttlefish conservation: a global review of methods to ameliorate unwanted fishing mortality and other anthropogenic threats to sustainability. ICES J Mar Sci 79:2579–2596. https://doi.org/10.1093/icesjms/fsac200
Bartram J, Mountjoy E, Brooks T, Hancock J, Williamson H, Wright G, Moppett J, Goulden N, Hubank M (2016) Accurate sample assignment in a multiplexed, ultrasensitive, high-throughput sequencing assay for minimal residual disease. J Mol Diagn 18:494–506. https://doi.org/10.1016/j.jmoldx.2016.02.008
Bokulich NA, Kaehler BD, Rideout JR, Dillon M, Bolyen E, Knight R, Huttley GA, Caporaso JG (2018) Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 22̆019s q2-feature-classifier plugin. Microbiome 6:90. https://doi.org/10.1186/s40168-018-0470-z
Boletzky SV (1971) Mandibules denticulées chez les larves des Teuthoidés et des Octopodes (Mollusca, Cephalopoda). C R Acad Sci Paris 272:2904–2906
Campbell CA (1984) Guía de la flora y fauna del mar mediterráneo. Omega, Barcelona, p 336p
Cardona L (1991) Measurement of trophic niche breadth using occurrence frequencies. J Fish Biol 39:901–903
Clarke KR, Warwick RM (2001) Change in marine communities, 2nd edn. PRIMER-E Ltd, Plymouth
Clarke KR, Warwick RM (2005) Primer-6 Computer Program. Council, N.E.R. (Ed.), Plymouth
Clarke LJ, Trebilco R, Walters A, Polanowski AM, Deagle BE (2020) DNA-based diet analysis of mesopelagic fish from the southern Kerguelen Axis. Deep Sea Res. Part II Top. Stud. Oceanogr Vol. 174
Conlan KE, Desiderato A, Beermann J (2021) Jassa (Crustacea: Amphipoda): a new morphological and molecular assessment of the genus. Zootaxa 4939:1–191. https://doi.org/10.11646/zootaxa.4939.1.1
Dan S, Iwasaki H, Takasugi A, Shibasaki S, Yamazaki H, Oka M, Hamasaki K (2019) Effects of co-supply ratios of swimming crab Portunus trituberculatus zoeae and Artemia on survival and growth of East Asian common octopus Octopus sinensis paralarvae under an upwelling culture system. Aquac Res 50:1361–1370. https://doi.org/10.1111/are.14013
Dan S, Kamei Y, Takeshima S, Yamashita K, Hamasaki K (2021a) Stepwise changes in morphology during the settlement process in a merobenthic octopus, Octopus sinensis, raised in the laboratory. Invertebr Biol 141:e12358. https://doi.org/10.1111/ivb.12358
Dan S, Shibasaki S, Takasugi A, Takeshima S, Yamazaki H, Ito A, Hamasaki K (2021b) Changes in behavioural patterns from swimming to clinging, shelter utilization and prey preference of East Asian common octopus Octopus sinensis during the settlement process under laboratory conditions. J Exp Mar Biol Ecol 539:151537. https://doi.org/10.1016/j.jembe.2021.151537
Dauvin JC (2024) Overview of predation by birds, cephalopods, fish and marine mammals on marine benthic amphipods. J Mar Sci Eng 12:403. https://doi.org/10.3390/jmse12030403
Diatta Y, Clotilde-Ba FL, Capape C (2001) Le régime alimentaire du poulpe commun, Octopus vulgaris Cuvier, 1797 (Cephalopoda, Octopodidae) de la côte du Sénégal (Atlantique Oriental Tropical). Bull Inst Natl Des Sci Technol La Mer Salambó 28:65–67
Edgar RC, Haas BJ, Clemente JC, Quince C, Knight R (2011) UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 27:2194–2200. https://doi.org/10.1093/bioinformatics/btr381
Escolar O, Villanueva R, Fernández-Álvarez FA (2021) Captador de pulpos juveniles. Utility model number U202131210, Oficina Española de Patentes y Marcas. Country for priority: Spain; Titularity: CSIC. https://digital.csic.es/handle/10261/271830. Accessed 26 Jun 2023
Esling P, Lejzerowicz F, Pawlowski J (2015) Accurate multiplexing and filtering for high-throughput amplicon-sequencing. Nucleic Acids Res 43:2513–2524. https://doi.org/10.1093/nar/gkv107
Fernández-Álvarez FÁ, Farré M, Sánchez-Márquez A, Villanueva R, Escolar O, Navarro J (2020) Potentially handicapped but otherwise functional: Malformations in prey capture tools show no impacts on octopus life. Ecol Evol 10:12685–12689. https://doi.org/10.1002/ece3.69
Fernández-Álvarez F, Machordom A, García-Jiménez R, Salinas-Zavala CA, Villanueva R (2018) Predatory flying squids are detritivores during their early planktonic life. Sci Rep 8:3440. https://doi.org/10.1038/s41598-018-21501-y
Fernández-González V, Toledo-Guedes K, Valero-Rodriguez JM, Agraso MM, Sanchez-Jerez P (2018) Harvesting amphipods applying the integrated multitrophic aquaculture (IMTA) concept in off-shore areas. Aquaculture 489:62–69. https://doi.org/10.1016/j.aquaculture.2018.02.008
Fiorito G, Gherardi F (1999) Prey-handling behaviour of Octopus vulgaris (Mollusca, Cephalopoda) on Bivalve preys. Behav Processes 46(1):75–88. https://doi.org/10.1016/S0376-6357(99)00020-0
Fiorito G, Affuso A, Basil J et al (2015) Guidelines for the care and welfare of cephalopods in research: a consensus based on an initiative by CephRes, FELASA and the Boyd Group. Lab Anim 49:1–90. https://doi.org/10.1177/0023677215580006
Folmer O, Hoeh W, Black M, Vrijenhoek R (1994) DNA primers for amplification of mitochondrial Cytochrome C oxidase subunit I from diverse metazoan invertebrates. Mol Mar Biol Biotechnol 3:294–299
García-Mayoral E, Roura Á, Moreno A, González FA (2022) Diet composition of wild Loligo vulgaris paralarvae along the West Iberian Peninsula coast. Mar Ecol Prog Ser 681:71–85. https://doi.org/10.3354/meps13915
González ML, Perez-Schultheiss J, Lopez DA (2011) Exotic amphipods in aquaculture systems: presence and potential use. Crustaceana 84:769–775
Guerra A (1978) Sobre la alimentación y el comportamiento alimentario de Octopus vulgaris. Invest Pesq 42:351–364
Handoo Z, Hunt DJ (2016) Taxonomy, identification and principal species. Root-Knot Nematodes 50:1–53
Hanlon RT, Messenger JB (2018) Cephalopod Behaviour, 2nd edn. University Press, Cambridge. https://doi.org/10.1017/9780511843600
Ibáñez CM, Keyl F (2010) Cannibalism in cephalopods. Rev Fish Biol Fish 20:123–136
Ibáñez CM, Riera R, Leite T, Díaz-Santana-Iturrios M, Rosa R, Pardo-Gandarillas MC (2021) Stomach content analysis in cephalopods: past research, current challenges, and future directions. Rev Fish Biol Fish 31:505–522. https://doi.org/10.1007/s11160-021-09653-z
Iglesias J, Fuentes L (2014) Octopus vulgaris. Paralarval culture. In: Iglesias J, Fuentes L, Villanueva R (eds) Cephalopod culture. Springer, London, pp 427–450
Iglesias J, Fuentes L, Villanueva R (2014) Cephalopod culture. Springer, Berlin, p 494. https://doi.org/10.1007/978-94-017-8648-5
Jiménez-Prada P, Hachero-Cruzado I, Guerra-García JM (2021) Aquaculture waste as food for amphipods: the case of Gammarus insensibilis in marsh ponds from southern Spain. Aquac Int 29:139–153. https://doi.org/10.1007/s10499-020-00615-z
Jeong S, Yu OH, Suh HL (2006) Secondary production of Jassa slatteryi (Amphipoda, Ischyroceridae) on a Zostera marina seagrass bed in southern Korea. Mar Ecol Prog Ser 309:205–211
Lenz S (1997) Cilia in the epidermis of late embryonic stages and paralarvae of Octopus vulgaris (Mollusca: Cephalopoda). Vie Milieu 47:143–147
Leray M, Yang JY, Meyer CP, Mills SC, Agudelo N, Ranwez V, Boehm JT, Machida RJ (2013) A new versatile primer set targeting a short fragment of the mitochondrial coi region for metabarcoding metazoan diversity: application for characterizing coral reef fish gut contents. Front Zool 10:1–14. https://doi.org/10.1186/1742-9994-10-34
Lincoln R, Boxshall G, Clarke P (1998) A dictionary of ecology, evolution and systematics, 2nd edn. Cambridge University Press, Cambridge
Martins CPP, Fernández-Álvarez FÁ, Villanueva R (2018) Invertebrate predation on egg masses of the European cuttlefish, Sepia officinalis: an experimental approach. Estuar Coast Shelf Sci 200:437–448. https://doi.org/10.1016/j.ecss.2017.11.016
Martínez J, Adarraga I (2008) First record of invasive caprellid Caprella scaura Templeton, 1836 sensu lato (Crustacea: Amphipoda: Caprellidae) from the Iberian Peninsula. Aquat Invasions 3(2):165–171. https://doi.org/10.3391/ai.2008.3.2.6
McManus MA, Woodson CB (2012) Plankton distribution and ocean dispersal. J Exp Biol 215:1008–1016
Nande M, Presa P, Roura Á, Andrews PLR, Pérez M (2017) Prey capture, ingestion, and digestion dynamics of Octopus vulgaris paralarvae fed live zooplankton. Front Physiol 8:573. https://doi.org/10.3389/fphys.2017.00573
Nigmatullin CM, Shchetinnikov AS, Shukhgalter OA (2009) On feeding and helminth fauna of neon flying squid Ommastrephes bartramii (Lesueur, 1821) (Cephalopoda: Ommastrephidae) in the southeastern Pacific. Rev Biol Mar Oceano 44:227–235
Nixon M (1987) Cephalopod diets. In: Boyle PR (ed) Cephalopod life cycles: comparative reviews, vol 2. Academic Press, London, pp 201–219
Olmos-Pérez L, Roura Á, Pierce GJ, Boyer S, González ÁF (2017) Diet composition and variability of wild Octopus vulgaris and Alloteuthis media (Cephalopoda) paralarvae through a metagenomic lens. Front Physiol 8:321. https://doi.org/10.3389/fphys.2017.00321
O’Rorke R, Lavery S, Chow S et al (2012) Determining the diet of larvae of Western rock lobster (Panulirus cygnus) using High-Throughput DNA sequencing techniques. PLoS ONE 7:e42757. https://doi.org/10.1371/journal.pone.0042757
Ortiz M, Jimeno A (2001) Guía para la identificación de familias y los géneros de los anfípodos del suborden Gammaridea de la península ibérica. Graellsia 57:1–76
Pham CK, Isidro E (2010) Experimental harvesting of juvenile common octopus Octopus vulgaris, for commercial ongrowing in the Azores. Arquipelago 27:41–47
Quetglas A, Alemany F, Carbonell A, Merella P, Sánchez P (1998) Biology and fishery of Octopus vulgaris Cuvier, 1797, caught by trawlers in Mallorca (Balearic Sea, western Mediterranean). Fish Res 36:237–249
Ramos JE, Roura Á, Strugnell JM, Moltschaniwskyj NA, Bargiela R, Pecl G (2023) Stomach content characterisation of the marine range-shifting Octopus tetricus using DNA metabarcoding. Mar Ecol Prog Ser 717:67–83
Ratnasingham S, Hebert PD (2007) BOLD: the barcode of life data system (http://www.barcodinglife.org). Mol Ecol Notes 7:355–364. https://doi.org/10.1111/j.1471-8286.2007.01678.x
Ribas T, Sales JBL, Boer HJ et al (2021) Unexpected diversity in the diet of Doryteuthis sanpaulensis (Brakoniecki, 1984) (Mollusca:Cephalopoda) from the southern Brazilian sardine fishery identified by metabarcoding. Fish Res 239:105936. https://doi.org/10.1016/j.fishres.2021.105936
Riedl R (1986) Fauna y Flora del Mediterráneo. Omega, Barcelona, p 858p
Robeson MS, O’Rourke DR, Kaehler BD, Ziemski M, Dillon MR, Foster JT, Bokulich NA (2020) RESCRIPt: Reproducible sequence taxonomy reference database management for the masses. bioRxiv. https://doi.org/10.1101/2020.10.05.326504
Rognes T, Flouri T, Nichols B, Quince C, Mahé F (2016) VSEARCH: A versatile open source tool for metagenomics. PeerJ 4:e2584. https://doi.org/10.7717/peerj.2584
Rosa R, Marques AM, Nunes ML, Bandarra N, Reis CS (2004) Spatial-temporal changes in dimethyl acetal (octadecanal) levels of Octopus vulgaris (Mollusca: Cephalopoda): relation to feeding ecology. Sci Mar 68:227–236
Rosa R, Roura Á, Amor M, Gleadall IG, Guerra Á, González ÁF, Lopes VM, Pereira J, Pierce GJ, Sampaio E, Villanueva R (2024) Chapter 13: Octopus vulgaris, the common octopus. In: Rosa R, Gleadall IG, Pierce GJ, Villanueva R (eds) Octopus biology and ecology. Academic Press, London, pp 187–216. https://doi.org/10.1016/B978-0-12-820639-3.00020-0
Roura Á, González ÁF, Redd K, Guerra Á (2012) Molecular prey identification in wild Octopus vulgaris paralarvae. Mar Biol 159:1335–1345. https://doi.org/10.1007/s00227-012-1914-9
Roura Á, Álvarez-Salgado XA, González ÁF, Gregori M, Rosón G, Otero J, Guerra A (2016) Life strategies of cephalopod paralarvae in a coastal upwelling system (NW Iberian Peninsula): insights from zooplankton community and spatio-temporal analyses. Fish Oceanogr 25:241–258. https://doi.org/10.1111/fog.12151
Roura Á, Doyle SR, Nande M, Strugnell JM (2017) You are what you eat: a genomic analysis of the gut microbiome of captive and wild Octopus vulgaris paralarvae and their zooplankton prey. Front Physiol 8:362. https://doi.org/10.3389/fphys.2017.00362
Roura Á, Amor M, González ÁF, Guerra Á, Barton ED, Strugnell JM (2019) Oceanographic processes shape genetic signatures of planktonic cephalopod paralarvae in two upwelling regions. Prog Oceanogr 170:11–27. https://doi.org/10.1016/j.pocean.2018.10.005
Roura Á, Castro-Bugallo A, Martínez-Pérez M (2023a) The settlement stage in the common octopus Octopus vulgaris Cuvier, 1797: a complex transition between planktonic and benthic lifestyles. Mar Biol 170:53. https://doi.org/10.1007/s00227-023-04188-2
Roura Á, Doyle SR, Castro-Bugallo A, Hall NE, Gonzalez AF, Strugnell JM (2023b) Trophic ecology of Octopus vulgaris paralarvae along the Iberian Canary current eastern boundary upwelling system. Sci Rep 13:8744. https://doi.org/10.1038/s41598-023-35206-4
Sánchez P, Obarti R (1993) The biology and fishery of Octopus vulgaris caught with clay pots on the Spanish Mediterranean coast. In: O’Dor RK, Kubodera T (eds) Recent Advances in Fisheries Biology Okutani T. Tokai Univ Press Tokyo, Japan, pp 477–487
Sauer WHH, Gleadall IA, Downey-Breedt N et al (2021) World Octopus Fisheries. Rev Fish Sci Aquac 29:279–429. https://doi.org/10.1080/23308249.2019.1680603
Shahin S, Okomoda VT, Ishak SD, Waiho K, Fazhan H, Azra MN et al (2023) Life history traits of the marine amphipod Cymadusa vadosa under laboratory conditions: insights on productivity and aquaculture potential. Aquat Sci 85:103. https://doi.org/10.1007/s00027-023-01000-7
Tur R, Domingues P, Almansa E, Lago-Rouco MJ, Garcia, P, Perez E (2020) Method for cultivating paralarvae of the common octopus, Octopus vulgaris. US 2020/0367476A1. https://patentimages.storage.googleapis.com/30/cb/c3/a6ffd92024b0ef/US20200367476A1.pdf. Accessed 26 Jun 2023
Uriarte I, Hernández J, Dorner J, Paschke K, Farias A, Crovetto E, Rosas C (2010) Rearing and growth of the octopus Robsonella fontaniana (Cephalopoda: Octopodidae) from planktonic hatchlings to benthic juveniles. Biol Bull 218:200–210
Vargas-Abundez JA, Martinez-Moreno GL, Simoes N, Norena-Barroso E, Mascaro M (2021) Marine amphipods (Parhyale hawaiensis) as an alternative feed for the lined seahorse (Hippocampus erectus, Perri 1810): nutritional value and feeding trial. PeerJ 9:e12288. https://doi.org/10.7717/peerj.12288
Varó I, Prado-Álvarez M, Ortea I et al (2022) Proteogenomic study of the effect of an improved mixed diet of live preys on the aquaculture of Octopus vulgaris paralarvae. Front Mar Sci 8:817701. https://doi.org/10.3389/fmars.2021.817701
Vestheim H, Jarman SN (2008) Blocking primers to enhance PCR amplification of rare sequences in mixed samples-a case study on prey DNA in Antarctic krill stomachs. Front Zool 5:12. https://doi.org/10.1186/1742-9994-5-12
Villanueva R (1995) Experimental rearing and growth of planktonic Octopus vulgaris from hatching to settlement. Can J Fish Aquat Sci 52:2639–2650. https://doi.org/10.1139/f95-853
Villanueva R, Norman MD (2008) Biology of the planktonic stages of benthic octopuses. Ocean Mar Biol Ann Rev 46:105–202
Villanueva R, Vidal EAG, Fernández-Álvarez F, Nabhitabhata J (2016) Early mode of life and hatchling size in cephalopod molluscs: influence on the species distributional ranges. PLoS ONE 11:e0165334. https://doi.org/10.1371/journal.pone.0165334
Villanueva R, Perricone V, Fiorito G (2017) Cephalopods as predators: a short journey among behavioral flexibilities, adaptions, and feeding habits. Front Physiol 8:598. https://doi.org/10.3389/fphys.2017.00598
Villanueva R, Coll-Lladó M, Bonnaud-Ponticelli L et al (2021) Born with bristles: new insights on the Kölliker’s organs of octopus skin. Front Mar Sci 8:645738. https://doi.org/10.3389/fmars.2021.645738
Zariquiey RA (1968) Crustáceos Decápodos Ibéricos. Inv Pesq 32:1–510
Acknowledgements
Special thanks to Eduardo Escolar, owner and captain of the artisanal fishing boat Nova Miriam. Thanks to him for all his work and marine knowledge shared along all the sampling days carried out to get the successful realization of this project. Thanks to Ainhoa Bernal (Instituto Español de Oceanografía, CSIC) for her support with statistical analyses. DNA metabarcoding analyses were carried out by AllGenetics & Biology SL (www.allgenetics.eu). Funding was provided by the Spanish Ministry of Science, Innovation and Universities (OCTOSET, Ref. RTI2018-097908-B-I00; ECOPHYN, Ref. PID2021-126824NB-C32; MCIU/AEI/FEDER, EU), and the Spanish government through the “Severo Ochoa Center of Excellence” accreditation (CEX2019-000928-S). F.Á.F.-Á. was supported by a JdC-I Postdoctoral Fellowship Grant (Ref. IJC2020-043170-I) awarded by MCIN/AEI /https://doi.org/10.13039/501100011033 and by a Beatriu de Pinós fellowship from Secretaria d´Universitats i Recerca del Departament de Recerca i Universitats of the Generalitat de Catalunya (Ref. BP 2021 00035). We thank the Associate Editor and anonymous reviewers for their helpful comments.
Funding
Open Access funding provided thanks to the CRUE-CSIC agreement with Springer Nature.
Author information
Authors and Affiliations
Contributions
RV defined the initial project and all authors contributed to the study conception and design. Sampling was performed by OE. Morphological data collection and analyses were performed by OE. Data and statistical analyses were performed by OE. Molecular data curation by OE and FÁF-Á. Funding acquisition by RV. The first draft of the manuscript was written by OE and all authors commented on subsequent versions of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
11160_2024_9875_MOESM1_ESM.xlsx
Supplementary file 1: Percentage of weight reduction after 5% MgCl2 anaesthesia during 30 min according to the different octopus size class groups. (XLSX 10 KB)
11160_2024_9875_MOESM2_ESM.pdf
Supplementary file 2: Non-metric multidimensional scaling (MDS) analyses with the superimposition of cluster, applied to the Jaccard index matrix based in presence/absence and to Bray-Curtis matrix based in semiquantitative data, both applied to single samples, grouped by size class, collector gear, season and the clustering of size class and gear factors. Abbreviations: 1–5, size class groups; CT, cuttlefish trap; JOC5, juvenile octopus collector at 5 m depth; JOC15, juvenile octopus collector at 15 m depth; OP, octopus pot; OT, octopus trap. (PDF 659 KB)
11160_2024_9875_MOESM3_ESM.pdf
Supplementary file 3: Percentage of O. vulgaris reads obtained per sample with and without blocking primers, and with fixed and variable annealing temperature. MC–50–50–OV, Mock community 50% and 50% O. vulgaris; MC–90–10–OV, Mock community 10% and 90% O. vulgaris; O. vulgaris 1–3, samples of O. vulgaris stomach content; T, temperature. (PDF 123 KB)
11160_2024_9875_MOESM4_ESM.xlsx
Supplementary file 4: OTU table of COI reads. Primary identifications were based on BOLD database through Qiime2. Some unassigned references were identified through BLAST. The Frecuency of Ocurrence (FO, %) of each prey item is indicated. (XLSX 130 KB)
11160_2024_9875_MOESM5_ESM.pdf
Supplementary file 5: Cluster analyses applied to the Jaccard index matrix based in presence/absence by size class, collector gear, season and the clustering of size class and gear groups. Abbreviations: 1–5, size class groups; CT, cuttlefish trap; JOC5, juvenile octopus collector at 5 m depth; JOC15, juvenile octopus collector at 15 m depth; OP, octopus pot; OT, octopus trap. (PDF 472 KB)
11160_2024_9875_MOESM6_ESM.xlsx
Supplementary file 6: Frequency of occurrence (FO) of all the identified species recorded from stomach contents of 91 samples of O. vulgaris in percentage (%), assorted by size class. Prey taxa are ordered by total FO (%). Taxa groups of Chromista, Fungi, Protozoa, Plantae, Insecta, Arachnida were excluded. The taxonomic criterion of WoRMS Editorial Board (2022) was applied. Asterisks designate species identified through BLAST at NCBI. Abbreviations: N/A, not available; SO, superorder. (XLSX 17 KB)
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Escolar, O., Fernández-Álvarez, F.Á. & Villanueva, R. Octopus diet during the settlement period using DNA metabarcoding. Rev Fish Biol Fisheries (2024). https://doi.org/10.1007/s11160-024-09875-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11160-024-09875-x