
Abstract
Anadromous brown trout (Salmo trutta) in the northeastern Atlantic Ocean, and coastal cutthroat trout (Oncorhynchus clarkii clarkii) in the northeastern Pacific Ocean, are analogs in many ways. Both species display similar patterns of complex life histories and behavioral flexibility, are iteroparous and facultatively anadromous and occupy nearshore coastal marine habitats where numerous populations often mix. These characteristics create specific challenges for management and conservation that have been complicated by inadequate scientific attention in some areas. Both species are declining across their native range, and their ecology make them particularly vulnerable to habitat destruction, fishing, and climate change. Here, we review the available literature to compare the biology, ocean ecology, and management practices of these two species. We highlight ecological similarities and differences between the species and identify current knowledge gaps suggesting future research needs and management actions. Using a comparative approach, the review aims to promote and facilitate knowledge exchange between anadromous trout research communities from opposite sides of the globe to improve management and conservation of these species and stimulate the production of management plans specific to anadromous trout.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Anadromous salmonids present a paradox; they are highly valued in commercial and recreational fisheries, as cultural icons for indigenous people, and for their ecosystem benefits, yet they are also often depleted and in jeopardy from human activities (Waldman and Quinn 2022). Consequently, there is a pressing need to better understand their behavior and ecology to mitigate impacts of anthropogenic activities and refine management and recovery strategies. Anadromous brown trout (Salmo trutta) and coastal cutthroat trout (Oncorhynchus clarkii clarkii) have received less attention from managers, relative to their often larger bodied and more abundant congeners that are the focus of commercial and recreational fisheries management (i.e., Atlantic salmon, Salmo salar, and Pacific salmon, Oncorhynchus spp., including steelhead O. mykiss). Consequently, management and conservation plans associated with these trout are less developed and often lack extensive and reliable data (Trotter et al. 1993; Walker et al. 2004) or evaluation of management strategies currently being utilized. This lack of information is concerning, given their recreational value and role in the ecology of freshwater and nearshore ecosystems (Knutsen et al. 2001; Jonsson and Jonsson 2011; Quinn 2018).
The anadromous forms of brown trout and coastal cutthroat trout evolved independently, in the northeastern sides of their respective oceans, yet they inhabit similar ecological/trophic/behavioral niches in their respective ecosystems, making them ideal candidates for a comparison of the ecology and behavior of two similar but allopatric species. Across their ranges, these species are referred to with a variety of names. Anadromous brown trout are most commonly known as “sea trout” (not to be confused with Cynoscion nebulosus, a non-salmonid estuarine fish commonly known as spotted seatrout or speckled seatrout, from southeastern North America) but also sewin in Wales, finnock (for young sea trout from a Scottish Gaelic word meaning white) in Scotland and Ireland, peal and mort in England, and white trout and slob trout in Ireland (the latter term referring to a specific migratory variant using estuaries). In marine waters, anglers refer to cutthroat trout as sea-runs, bluebacks (though this term is also applied, in some areas, to sockeye salmon, O. nerka, and coho salmon, O. kisutch), harvest trout, sea-run cutthroat (SRC) and cutties. Both anadromous brown trout and cutthroat are concentrated in areas with sheltered coastlines, making them accessible to anglers in small boats and on shore. Achieving smaller average sizes than their anadromous congeners (i.e., Atlantic salmon and Pacific salmon and steelhead), both species express considerable phenotypic plasticity as a result of their iteroparous life history (Hutchings 2004), flexible freshwater habitat use patterns, and little genetic separation between anadromous and resident life forms within streams (Hindar et al. 1991; Johnson et al. 2010; Lemopoulos et al. 2018). Specifically, brown and cutthroat trout can display resident or anadromous life history pathways and partial migration regardless of those expressed by their parents (Claiborne et al. 2020; Duval et al. 2021).
While research is limited on anadromous trout globally, recent work on anadromous brown trout has accelerated understanding and provided guidance for research on the less-studied coastal cutthroat trout (Fig. 1) (Birnie-Gauvin and Aarestrup 2019; Kristensen et al. 2019a; Nevoux et al. 2019; Rohtla et al. 2020; Kennedy et al. 2022). By comparing the behavior and ecology of these species, the function and adaptive value of behaviors can be more clearly determined, and recent advances can stimulate communication and growth of current research programs and prioritize data gaps for these ecologically similar species. Together, this information should lead to comprehensive management plans designed specifically for anadromous trout which are lacking in their respective ranges.

Source: Web of science search terms included “anadromous” or “sea run” and “cutthroat” and “sea trout” or “anadromous and brown trout”
Number of peer reviewed papers and reports published annually on anadromous brown (grey bars) and cutthroat trout (black bars) between 1950 and 2021.
Here we review the literature on anadromous brown trout and anadromous cutthroat trout and introduce new data to better understand the behavior and ecology of trout inhabiting marine waters. Specifically, our objectives are to describe 1) key biological characteristics including size, maximum age and growth, 2) marine ecology and behavior including diet, parasites, and migration patterns 3) and management approaches for these two species. Finally, common threats and data gaps for anadromous brown trout and cutthroat trout revealed from this work are discussed to provide a foundation for further research and production of management plans where none exist.
Origin and phylogeny
The genera Oncorhynchus and Salmo diverged from a common ancestor several millions of years ago. A fossil discovered in 1964 assigned to O. rastrosus, the saber-toothed salmon, was estimated to originate from the late Miocene to Pleistocene period (Sepkoski 1992), indicating that the Salmo genus diverged from Oncorhynchus well before the Pliocene, and as early as the Miocene (20 MYA). While cutthroat and brown trout are direct descendants of unique ancestors, O. rastrosus and S. salar, respectively, (McKay et al. 1996; Bernatchez et al. 2004; Trotter et al. 2018), their origin is the product of similar circumstances (glaciation, dispersal limitations, geographic barriers, and other climate-driven factors) in the Pleistocene era. Specifically, brown trout diverged from Atlantic salmon into several isolated geographic lineages (Bernatchez 2001; Crete-Lafreniere et al. 2012) that remained separate from each other for roughly 2 million years, and all retained the capacity for anadromy (Sloat et al. 2014). The anadromous form is thus not phylogenetically distinct from freshwater forms (Ferguson et al. 2019). Within these lineages, some authorities recognize multiple subspecies (Page 2008; Whiteley et al. 2019; Schöffmann 2021) and this is an area of rapidly developing science (Tougard 2022), but for the purpose of this paper these putative subspecies will be referred to as brown trout in the species trutta, given that they all have access to the marine environment and commonly express anadromy.
Outside the native range of brown trout in the northeastern Atlantic, cutthroat trout evolved in the northeastern Pacific (Smith and Stearley 2018). They occur on both sides of the Rocky Mountains (a major faunal divide for North American fishes) including the headwaters of the Rio Grande, Colorado, and Missouri rivers, and interior basins that do not currently flow into marine waters. Although many subspecies of cutthroat trout have been described (e.g. Yellowstone, Westslope, Bonneville, etc.), coastal cutthroat trout represent the oldest existing lineage (Behnke 1992), the only anadromous one, and the focus of this review. Coastal cutthroat trout demonstrate considerable life history diversity between nearby watersheds (Guy et al. 2008; Zimmerman et al. 2015; Losee et al. 2017b) and across their range on the west coast of North America. This life history diversity occurs despite a cline in genetic diversity; Griswold et al. (1997) described low genetic diversity for cutthroat trout that recently colonized streams in Alaska at the northern end of their range and higher genetic diversity in long-established populations farther south. In contrast to their ages as species, anadromous brown trout and coastal cutthroat trout currently occupy environments that are relatively young in the current post-glacial period, thus the fish must have colonized and been extirpated repeatedly between glacial retreats and advances.
At the end of the last glacial period about 13,500 years ago (Pielou 2008), many of the modern-day fjords began to form, converting glacial canyons to marine basins. This process took place in the Baltic Sea and fjords of Norway, bays in Ireland and Great Britain, numerous waterways of coastal Alaska and British Columbia (McPhail and Lindsey 1986), and hundreds of other bays, channels and sounds across the Northern Hemisphere. The Baltic Sea in northern Europe and Salish Sea along the west coast of North America exemplify the results of these geological events, with habitats occupied by anadromous brown and cutthroat, respectively (Huitfeldt-Kaas 1918; Trotter et al. 2018). Specifically, over the last ~ 10,000 years the Baltic and Salish seas transitioned from freshwater lakes to inland marine waters connected to the Atlantic and Pacific oceans as glaciers receded and sea levels rose and flooded glaciated valleys throughout the Northern Hemisphere (Alt and Hyndman 1995).
Geologically, the Baltic Sea is very young, forming when the Scandinavian ice sheet retreated after the last glaciation in Europe approximately 12,000 years ago. However, since that time the Baltic Sea has experienced a variety of salinity changes and transitionary phases of marine/brackish water (Harff and Hoth 2011). The Baltic Sea as we know it today, with brackish water and narrow connection to the North Sea, has been in place for approximately 2,000 years (Emeis et al. 2003; Leppäranta and Myrberg 2009) with a surface salinity range from 0.3% to 0.9% (Viktorsson 2018). Similarly, the Salish Sea’s connection and estuary-like salinity stabilized in the last 4,000 years, providing a transitional environment between the rivers of the Pacific Northwest and the Pacific Ocean (Burns 1985; Alt and Hyndman 1995). These two habitats (the Baltic and Salish seas) and others like them promoted diversification and phenotypic plasticity of anadromous forms of brown and cutthroat trout (Campton and Utter 1987; Bernatchez 2001) under similar circumstances.
Distribution
Coastal cutthroat trout occur from northern California to south-central Alaska (Trotter 2008), though the other subspecies are more widely distributed. Brown trout is much more widely distributed than coastal cutthroat trout, having historically occupied much of Europe and a large area in Asia (Snoj et al. 2021) (Fig. 2). For both species, the greatest abundance of anadromous forms exists along temperate latitudes where sheltered coastlines are common. As a result of stocking by Europeans, beginning in the mid to late 1800’s, naturalized populations of brown trout (often anadromous) exist in every continent except Antarctica, and in many island groups in the southern hemisphere (MacCrimmon and Marshall 1968; MacCrimmon et al. 1970; Klemetsen et al. 2003).

Brown trout rarely go beyond the fjord of their home river (Jonsson and Jonsson 2011) but longer distance migrations have been documented (Bekkevold et al. 2021). Additionally, anadromous brown trout inhabit regions without large, fjord-like inlets in both their native ranges such as the coastlines of North Africa (Lobón-Cerviá et al. 2019) and France (Nevoux et al. 2019), and outside their native range such as Japan (Honda et al. 2012), South America (O'Neal and Stanford 2011; Minett et al. 2021), Newfoundland (Westley and Fleming 2011), and the Kerguelen Islands in the southern Indian ocean (Lecomte et al. 2013).
Off the coast of Iceland, brown trout spent the majority of time (72–93%) near the shore in water < 5 m deep. However, they have been detected > 70 m below the surface (Sturlaugsson 2017; Kristensen et al. 2019c), suggesting that offshore distributions to deeper water are possible. Similarly, spawning populations of anadromous cutthroat are present in numerous small independent tributaries of the Pacific Ocean in Oregon and California that lack protected fjord-like environments (Johnson et al. 1999). The depths occupied by cutthroat in the ocean have not been reported but they have been observed in offshore waters. For instance, Loch and Miller (1988) captured juvenile and adult cutthroat 31.5 km offshore near the Columbia River plume, and Pearcy et al. (1990) captured them up to 66 km offshore. In the latter study, an individual cutthroat was captured and released in Yaquina Bay, Oregon and recaptured 290 km to the south, near the mouth of the Umpqua River, thus demonstrating the potential for long-distance migrations, though this seems to be rare. Together, this information highlights the broad spatial distribution of anadromous brown and cutthroat trout within their respective ranges, but reliance on nearshore habitats at all marine life stages generally characterizes both species and distinguishes them from most other anadromous salmonids. Indeed, the failure to colonize the western shores of their respective oceans seems consistent with limited oceanic distributions rather than specialized habitat requirements in fresh water.
Self-sustaining populations of brown trout outside their native range threaten native species via predation and competition (Idyll 1942; Bisson et al. 1986; Sánchez-Hernández 2020) but also have considerable economic importance across the globe (Ahn et al. 2000; O’Reilly et al. 2006; O’Reilly 2015; Blicharska and Rönnbäck 2018). In contrast, spawning populations of coastal cutthroat trout have not been documented outside their native range, and efforts to transplant them have been limited (Crawford and Muir 2008) thus distribution patterns for coastal cutthroat have not changed significantly over the last 200 years (Metcalf et al. 2012). This difference in range expansion is likely due to the limited artificial breeding and stocking that has occurred for coastal cutthroat trout. This literature comparison with brown trout provides a glimpse into potential benefits and risks that could result from future efforts to artificially stock cutthroat outside their native range.
Life history and migratory patterns
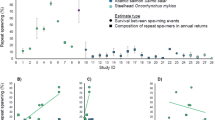
Brown and cutthroat trout exhibit four basic life history pathways: lacustrine (lake dwelling), fluvial (riverine), adfluvial (river and lake), and anadromous (marine) (Trotter 1989; Lobón-Cerviá and Sanz 2017; Arostegui and Quinn 2019; Jonsson et al. 2019). The anadromous forms are the focus of this report; however, individuals entering marine water can express multiple life history patterns during their life span, either before or after their initial freshwater emigration (Saiget et al. 2007; Thorstad 2016). Moreover, offspring from anadromous parents may remain in freshwater habitats their entire life (Rohtla et al. 2017; Claiborne et al. 2020), and non-anadromous parents may produce anadromous offspring (Fig. 3). For instance, approximately 12% of anadromous brown trout sampled in the Baltic Sea near Estonia were produced by non-anadromous mothers, based on otolith microchemistry analysis (Rohtla et al. 2020). Such partial migration, where individuals from the same population exhibit differential migratory behavior, has been well documented for salmonids including brown and cutthroat trout (Saiget et al. 2007; O'Neal and Stanford 2011; Sloat 2013; Rohtla et al. 2020; Duval et al. 2021) and reviewed across various systems (Vélez-Espino et al. 2013; Sloat et al. 2014; Kendall et al. 2015). The factors determining whether individuals remain in fresh water or migrate to sea are not fully understood but involve a combination of genetic and environment controls; regardless, this flexibility tends to increase the resilience of populations (Jonsson and Jonsson 1993; Chapman et al. 2011, 2012; Hayes et al. 2012; Lemopoulos et al. 2018; Ferguson et al. 2019; Nevoux et al. 2019).

Generalized life cycle, examples of common patterns and some of the major threats of anadromous brown and anadromous cutthroat trout; adapted from Nevoux et al. (2019)
The importance of the freshwater environment for both cutthroat and brown trout is evident in the extended and variable freshwater rearing period and the diversity of stream and lake habitats they use. Both species may rear for 1 to 6 years prior to ocean entry in spring but 2–4 years is typical (Sumner 1962; L'abée-Lund 1989; Trotter 1989). However latitudinal gradients in smolt age have been documented (Jonsson and L'Abee-Lund 1993) and recent studies have revealed the importance of rearing and migration patterns that were once assumed to be uncommon, including autumn downstream migration (Birnie-Gauvin and Aarestrup 2019; Jensen et al. 2022), use of non-natal streams (Taal et al. 2018; Källo et al. 2022), and dispersal of juvenile brown trout less than one year of age from natal waters into neighboring streams via the marine environment, reported by Taal et al. (2018). Degerman et al. (2012) hypothesized that this behavior could result from habitat limitations in the natal stream (e.g., density dependence, flow limitations etc.) and/or filling unused nearby rearing areas. However, the fate of individuals expressing such dispersal is unknown. No such pattern has been documented for cutthroat trout but they regularly spawn in small streams with limited flow (Pauley et al. 1989; Losee et al. 2016). These intermittent flows in the spring may support spawning, incubation, and emergence of fry but are not sufficient for larger juveniles in summer. In such cases, juveniles may seek suitable habitat in neighboring streams. The tools exist to clarify the role of these less understood life history patterns. Fisheries managers and scientists should prioritize this research, given the growing body of evidence that life history diversity is important in stabilizing and recovering anadromous fish populations (Moore et al. 2014; Schindler et al. 2015; Jonsson et al. 2019).
While a comprehensive understanding of movement patterns for anadromous brown trout and cutthroat is lacking, general behaviors have been described and provide an important area for comparison between the two species (Table 1). For instance, both species occupy similar ecological niches in their respective marine habitats and exhibit repeatability in migratory behaviors for individuals at the juvenile (Bohlin et al. 1996; Goetz et al. 2013; Zydlewski et al. 2014) and adult (Losee et al. 2017b; Halttunen et al. 2018; Birnie‐Gauvin et al. 2021) life stages. Specifically, they rarely migrate more than 150 km from land with typical migrations < 80 km from natal rivers (Thorstad 2016; Quinn 2018; Schöffmann 2021). For instance, Berg and Berg (1987) documented that 93% of Norwegian trout remained within 80 km of their natal river mouth, and Kallio-Nyberg et al. (2002) reported that > 90% of tagged sea trout were caught within 200 km. For cutthroat, results are sparse but the majority of cutthroat were caught within 30 km of their natal stream in south Puget Sound based on genetic stock identification (Losee et al. 2017a, b) and 77 km from their tagging location in the Pacific Ocean (Pearcy et al. 1990). While some longer distance migrations have been documented for both species (Kallio-Nyberg et al. 2002; Pearcy et al. 2018; Kristensen et al. 2019a) quantitative estimates of the proportions of fish offshore vs. nearshore exist are limited for brown trout and non-existent for cutthroat. As mentioned previously, these limited distances traveled are markedly different from their congeners. This difference was revealed by Moore et al. (2010), who compared migratory patterns between cutthroat trout and their close relative, steelhead, and their naturally occurring hybrids in Hood Canal, a fjord of Puget Sound. Moore et al. (2010) showed that the long, surface-oriented migrations to offshore waters characteristic of steelhead were not exhibited by coastal cutthroat trout which remained in nearshore waters within Hood Canal, and the hybrids showed intermediate patterns between those of the two pure species. Similarly, Klemetsen et al. (2003) highlighted the significantly shorter migratory distances traveled by brown trout compared to Atlantic salmon.
Given their flexible life history strategy, it is not a surprise that variations to general patterns of movement have been identified. For instance, Ferguson (2006) reviewed molecular methods used to describe two variations of anadromy for brown trout: the typical “sea trout” that migrates to marine water to feed prior to returning to natal rivers to spawn and the “slob trout” that is restricted to the estuary. Similarly, acoustic tracking of cutthroat trout in Hood Canal, revealed two movement “clusters” based on a variety of migratory metrics with some overlap between clusters. These clusters included individuals that exhibited an extended estuary residence time and limited movement and some with a larger range and less time spent in the estuary (Goetz et al. 2013). These categories were generally consistent with the results of Losee et al. (2017b) in three other fjords of south Puget Sound. Subsequently, Losee et al. (2018) observed high site fidelity of cutthroat, remaining within or returning to small sections of coastline for more than 12 months with short absences during the late winter, presumably moving into nearby streams to spawn.
This pattern of high site fidelity to marine beaches was observed for cutthroat trout as young as age 2, suggesting that individuals may disperse to locate favorable habitat (e.g., productive feeding areas, low predation risk, optimal temperature etc.) at a young age and then remain there for much of their lives. While site fidelity of trout in freshwater has been documented at various life stages (Bachman 1984; Heggenes and Slaveit 1990; Heggenes et al. 1991; Kuliskova et al. 2009; Slavík and Horký 2019) this behavior has not been described for anadromous brown trout in marine water. However, similar behavior was detected for anadromous bull trout in North Puget Sound, USA (Hayes et al. 2011), and anadromous grayling (Thymallus thymallus) in the Baltic Sea (Hellström et al. 2023), and it may also occur in anadromous brown trout in Europe. Identifying marine areas of high use by trout would provide managers an opportunity to protect vulnerable or threatened populations through refined fishery regulations or focused habitat improvement projects.
Considerable research has been focused on the factors affecting migratory patterns of anadromous trout. For instance, Jonsson and Jonsson (2014) reported that larger anadromous brown trout traveled farther from natal spawning locations than smaller individuals leaving streams of southern Norway, though this pattern has not been observed consistently (Thorstad 2016). For anadromous cutthroat, Goetz et al. (2013) showed that modes of behavior were not related to size or age, and Losee et al. (2017b) showed no effect of size on movement patterns across a broad length range. Thus, size and age are not consistently associated with migratory patterns, and small brown trout and cutthroat trout have been encountered far from their natal stream in their respective ranges (Pearcy et al. 1990; Thorstad 2016). Movement patterns of anadromous trout at various stages of the life cycle are likely determined by environmental and genetic tradeoffs between growth potential (e.g. temperature and food availability) affecting physiology (Archer et al. 2020), and mortality risk in differing environments (Zydlewski et al. 2008). Studies of movement, indicate that the most important determinants of migratory distance and behavior at sea include season (Moore et al. 1998; Losee et al. 2017b), fish condition (Haque 2008; Nevoux et al. 2019; Shry et al. 2019), predation (Dieperink et al. 2001), and ectoparasite intensity (Halttunen et al. 2018) but it is worth noting that few quantitative estimates of the proportion of anadromous trout found offshore vs. inshore exist, especially for cutthroat.
Following the marine phase of the brown and coastal cutthroat trout life cycle, the age and timing of the return to freshwater varies and can include trips to freshwater outside the spawning period (Taal et al. 2018; Claiborne et al. 2020). Furthermore, the iteroparous nature of both species adds complexity including individuals that return to freshwater for the first time but do not spawn, and individuals remaining at sea for more than one year, skipping a spawning event (Trotter 1989; Thorstad 2016). Typically, the initial return to freshwater occurs between total ages three and five when maturity is reached (Trotter 1989; Jonsson and Jonsson 2006; Claiborne et al. 2020), and is annual thereafter. The diversity of behaviors expressed by adult anadromous brown and cutthroat trout in freshwater have not been fully described but range from short intermittent visits, extended freshwater residence to overwinter and complex fluvial migrations.
The spawning season represents one of the greatest dissimilarities between these two species; coastal cutthroat trout typically spawn in the late winter and early spring (Losee et al. 2016) whereas brown trout spawn in the fall to early winter (Jepsen et al. 1998; Birnie‐Gauvin et al. 2021). Importantly, both species show considerable variation in the period between entry into fresh water and the spawning period, including so-called “premature migration” that may occur many months or even years prior to spawning, or migration immediately prior to spawning (Quinn et al. 2016; García-Vega et al. 2022). While the spawning season differs significantly between brown and cutthroat trout, both may spawn up to five times or more (Jones 1975; Peoples et al. 1988; Lobón-Cerviá and Sanz 2017) during their life. The flexible nature of both species has been revealed through observations of anadromous cutthroat trout spawning in the fall in the Elwha River system, WA, USA (McMillan et al. 2014) and spring spawning of anadromous brown trout in a tributary of the Black Sea (Latiu et al. 2020). This protracted or variable spawning period suggests that environmental influences and other co-occurring species are important determinants of the spawning period along with differences between the biology of brown and cutthroat trout.
The timing of freshwater entry is determined in large part by local stream conditions (Trotter 1989; Svendsen et al. 2004; García-Vega et al. 2022) but also biological/genetic controls given the repeatability in spawn timing across years of variable environmental conditions (Eldøy et al. 2019; Jensen et al. 2020). Specifically, adult anadromous brown and cutthroat trout can overwinter in freshwater (Pearcy et al. 1990; Birnie‐Gauvin et al. 2021), marine (Rikardsen et al. 2006; Jensen and Rikardsen 2012; Losee et al. 2018), and estuary habitats (Loch and Miller 1988; Etheridge et al. 2008; Jensen and Rikardsen 2008), and both display two main periods of freshwater entry: in the fall and winter (Trotter 1989; Dêbowski 2018) with summer river entry also common in some systems (Birnie‐Gauvin et al. 2021). While the conditions that separate the freshwater entry period from the spawning period are not completely known, bigger river systems and those with lakes may encourage an earlier entry timing as a result of safer and less energetically demanding habitat, as was shown for brown trout in streams in southern (Jonsson et al. 2018) and coastal Ireland (Kennedy et al. 2022), and Atlantic salmon in Ireland (Reed et al. 2017). For cutthroat, nutrient subsidies produced from sympatric semelparous salmon populations (e.g., eggs and flesh) provide a growth advantage for this fish entering earlier in the fall (Trotter 1989). In addition to the importance of stream size within a given region, the duration of feeding in marine waters also varies with latitude. For example, brown trout in northern Norway averaged only 68 d at sea (Berg and Jonsson 1989) with compressed migrations downstream and upstream, whereas those in some Irish rivers have very protracted migrations lasting more than 10 months (Quinn et al. 2016). Thus some anadromous brown trout and cutthroat trout are essentially freshwater fish that briefly enter marine waters to feed, whereas others are, after the smolt transformation, essentially marine fish that briefly return to streams to spawn, and then quickly return to marine waters (Quinn and Myers 2004; Quinn et al. 2016; Losee et al. 2018).
Given their small average size relative to other anadromous salmonids and physically variable spawning streams across the range, anadromous brown and cutthroat trout are limited by stream flow, sediment size, and temperature to successfully fertilize eggs and deposit them in redds (Quinn 2018; Ferguson et al. 2019; Nevoux et al. 2019). These size-based limitations and associated preferences for higher order spawning tributaries represent a key similarity between these species and may provide the benefit of reduction in hybridization by naturally segregating brown trout and cutthroat trout from their counterparts—Atlantic salmon and steelhead (Buehrens et al. 2013). However, conclusions regarding the selective pressures driving habitat preferences are speculative and need further investigation. Characteristics of typical spawning areas are similar between anadromous brown and cutthroat trout (Table 1). However, like other aspects of anadromous trout life history, variation from typical spawning locations has been described, including spawning in brackish water by brown trout (Limburg et al. 2001; Gabrielsen et al. 2021). Together, this information highlights the diverse movements of anadromous brown and cutthroat trout but also reveals predictable migratory and spawning patterns driven by biotic and abiotic factors and highlights key data gaps, particularly for cutthroat trout.
Physical characteristics
Anadromous brown and cutthroat trout share important physical similarities that differ from many non-anadromous trout species. Both anadromous brown and anadromous cutthroat trout have numerous irregularly shaped spots, as opposed to the round spots that are common among fish in the Oncorhynchus family (Fig. 4). Additionally, both lack spots on the ventral surface in the marine environment. The maxillary of anadromous brown and cutthroat trout extends beyond the eye, creating a relatively wide gape that supports an ontogenetic shift to a piscivorous diet at a small body size relative to other salmonids (Brodeur et al. 1987; Knutsen et al. 2001). In the marine environment, brown trout and cutthroat trout have a silvery coloration similar to other marine fish of similar size (Denton and Nicol 1966) but with heavier spotting and countershading, likely highlighting the importance of camouflage in the nearshore marine environment. Upon their return to freshwater as adults, yellowish or brown coloration is common. These changes in color are used by biologists and anglers to identify recent transitions from one environment to the other.

Illustration of anadromous brown trout (above) and cutthroat trout (below). By Joseph Tomelleri
Both species display great variation in color patterns among populations and subspecies, but there are some consistent differences between them. Notably, cutthroat trout have red slashes under the jaw (especially prominent when they are in fresh water) that are the source of their common name, and spotting on both the dorsal and ventral lobes of the caudal fin. In contrast, brown trout often lack spots on the tail or carry light spotting on the upper lobe. When brown trout are in freshwater, red spotting is common with a white or blue “halo” around spots, but these are less common or absent in the anadromous form (Klemetsen et al. 2003). Sex specific traits observed in the field are common, and especially evident during the spawning phase. Male characteristics are affected by environmental conditions and population specific genetic controls and include longer upper and lower jaw, shorter dorsal and anal fins and a larger adipose fin relative to females (Gruchy and Vladykov 1968; Jonsson and Jonsson 2015). Females commonly have a round belly as a result of developing gonads and greater length, weight and reduced diversity in size relative to male brown trout (Jonsson and Jonsson 2006). This is consistent with other iteroparous salmonids (Beacham and Murray 1987; Young 2005; Tamate and Maekawa 2006; Englmaier et al. 2021). These patterns are likely consistent for cutthroat trout but sex-specific differences in size for anadromous cutthroat trout have not been documented. Variation in sex-specific traits across environments and life stages results in sex-specific chromosomes that allow for sex determination using genetic methods when field methods are not adequate (Li et al. 2011). A similar, non-lethal, approach to determine sex has not been developed for cutthroat trout but would be an important step to understanding behavioral differences and patterns of life-history expression between male and female cutthroat trout in the marine environment.
Anadromous brown trout and coastal cutthroat occupy a similar ecological niche and as a result their morphology and meristics are similar and were well described in Linnaeus (1758) for brown trout and in Behnke’s Native Trout of Western North America (1992) for cutthroat. Both have teeth developed on upper and lower jaws and two rows on the tongue. Coastal cutthroat trout have an additional set of teeth between the gill arches on the floor of the pharynx on the basibranchial (hyoid or basibranchial teeth) that are absent in brown trout, perhaps indicating slightly different diet or feeding behavior. Scale counts are similar, ranging from 118–130 for anadromous brown trout and 120–180 for anadromous cutthroat trout (Linnaeus 1758; Page and Burr 1991) (Table 1). Vertebrae counts range from 59 to 64 for coastal cutthroat and 56 to 61 for anadromous brown trout (Linnaeus 1758). Gill raker counts are low in both species, relative to other salmonids, ranging from 13–18 in anadromous brown trout and 15–21 in coastal cutthroat trout (Linnaeus 1758; Tåking 1950). Given that gill rakers restrict food from exiting the mouth, few gills rakers indicate a diet dominated by macroinvertebrates and fishes rather than plankton.
The broad distribution of both anadromous brown trout and cutthroat trout has resulted in diverse physical characteristics for adaptation to unique environmental variability. For instance, Vatandoust et al. (2014) reported significant differences between populations across a set of 31 morphological and 7 meristic characters for brown trout in a small geographic area in the southern Caspian Sea. Tåking (1950) demonstrated the importance of changes in temperature in regulating vertebrae count for brown trout suggesting phenotypic plasticity in some traits. Similarly, Zimmerman et al. (2015) documented four distinct meristic patterns among cutthroat trout in a tributary of the Nisqually River, Muck Creek, in Washington State, USA; differences in scale counts and dorsal and anal fin rays of cutthroat trout coincided with genetic differences between individuals from different study sites.
Adult anadromous brown trout and cutthroat exhibit similar body condition (weight at length) when sampled in marine waters (Fig. 5). At sea, both typically range in size from 20 to 56 cm (Jonsson and Jonsson 2007; Quinn 2018) but larger specimens are common in some areas. Moreover, the average size of trout sampled in marine water can vary considerably across seasons (L'Abée-Lund 1991), particularly for brown trout, which can exceed 80 cm, however sampling effort throughout the year in marine waters is limited (Fig. 6). Far fewer samples have been collected of anadromous cutthroat trout relative to anadromous brown trout, limiting a comparison of maximum size. Biologists measured cutthroat up to 60 cm in an upstream trap just above tide water in Puget Sound Washington (WDFW unpublished data) and one individual was reported measuring 75.6 cm in Victoria Harbor, B. C. (Roger Ptolemy pers. comm., Table 1). However, the average lengths of brown trout and cutthroat trout are less than those of their closest relatives occupying marine habitats (i.e., Atlantic and Pacific salmon and steelhead) (Brodeur et al. 1987; Jonsson and Jonsson 2011; Quinn 2018).

Fork length vs. weight of cutthroat (black) and brown trout (grey) captured in marine waters. Different shapes represent different data sets from Norway (J. Davidsen, Pers. Comm.), United Kingdom (B. Brown, Pers. comm.), Baltic Sea (SLU 2022), Denmark (Kristensen et al. 2019b), South Puget Sound (WDFW, Pers. comm.), North Puget Sound (T. Quinn, Pers. Comm.), Vancouver Island, B.C. (B. O’Connor Pers. Comm.) and Pacific Ocean from Central Oregon to Northern Washington (Pearcy et al. 2018)

Mean monthly fork length of anadromous brown trout Salmo trutta (black) and coastal cutthroat trout Oncorhynchus clarkii clarkii (grey) sampled from Baltic Sea (SLU 2022), Norway (J. Davidsen, Pers. Comm.), Wales (B. Brown, Pers. comm.), Ireland (W. Roche, pers. Comm.), Puget Sound (WDFW), Vancouver Island, B.C. (B. O’Connor, Pers. Comm.) and coastal Oregon to northern Washington (Pearcy et al. 2018) and coastal California (W. Duffy Pers. Comm.)
It is not clear why anadromous brown and cutthroat trout are so much smaller than other anadromous salmonids. Pearcy et al. (1990) hypothesized that the small size of adult anadromous cutthroat trout, relative to sympatric salmon and steelhead, allows them to spawn, and offspring to rear, in small streams with abundant marginal habitat that young of the year favor (Moore and Gregory 1988). Larger-bodied steelhead overlap in distribution but breed in larger streams and farther downstream than cutthroat trout (Jones 1975; Trotter 1989; Buehrens et al. 2013). This size-biased habitat segregation may be reinforced by the apparent survival deficit experienced by hybrids. That is, naturally occurring hybrids occur regularly, as inferred from sampling of juveniles from throughout their overlapping range in northern California (Baumsteiger et al. 2005), coastal Washington: (Buehrens et al. 2013), Puget Sound, Washington, (Campton and Utter 1985; Moore et al. 2010), and south-central Alaska: (Williams et al. 2007). In these and other studies, authors report that most hybrids are the product of F1 backcrosses whereby adult hybrids spawned with pure cutthroat or steelhead, indicating that some hybrids survive to adulthood. However, reports of adult hybrids are uncommon relative to hybrids at the juvenile stage, suggesting hybrids experience reduced fitness through factors that select against adults favoring similar spawning habitats and interbreeding (Ostberg et al. (2004).
Small size as adults relative to many sympatric salmonids also makes brown and cutthroat trout less conspicuous to predatory birds and mammals in the shallow nearshore environment that they inhabit (Pemberton 1976; Lyse et al. 1998). In this way, relatively small anadromous size of brown trout and cutthroat is likely a trait selected for that enables them to occupy a unique ecological niche that supports successful feeding and predator avoidance in the nearshore marine environment and as mentioned earlier, may reduce competition and hybridization with their larger congeners in fresh water. Recent work focused on trout physiology has strengthened understanding of anadromous brown trout and cutthroat (Boel et al. 2014; Desforges et al. 2021; Anlauf-Dunn et al. 2022), highlighting the role of temperature in determining rates of growth and life-history expression, particularly in freshwater. As this area of research grows, experimental studies on growth in controlled conditions (e.g., comparing brown and cutthroat trout) would be especially fruitful to understanding the eco-physiology of anadromous trout and the evolutionary significance of documented growth and size traits.
Feeding and growth
As visual, opportunistic predators, brown and cutthroat trout exhibit diverse feeding habits and diet studies reveal fine-scale differences in habitat utilization and prey availability (Keeley and Grant 2001). Diet analysis from anadromous trout suggests the prey consumed vary with the trout’s size, age, and season, but are typically dominated by prey from the marine littoral zone, reflecting movements largely restricted to the nearshore environment (discussed later). During the day, both species are observed feeding and can be caught by anglers, but brown trout also show some specialization for crepuscular or nocturnal feeding, based on diet differences and stomach fullness during daylight hours vs. night (Pemberton 1976), and reduced feeding intensity during daylight (Heggenes et al. 1993). Studies of this kind are limited for anadromous brown trout and absent in the literature for cutthroat, and this would be a good topic for future work to better understand behavioral adaptations, food web dynamics and habitat prioritization to aid in management of coastal trout.
Feeding studies are much more common for anadromous brown trout than cutthroat but both rely heavily on stomach content analysis. For anadromous brown trout, broad scale patterns of feeding have been described with a diet dominated by nearshore invertebrates and fish. Piscivory is consistently reported as important for anadromous brown and cutthroat trout, with increasing contribution of fish in the diet with increasing trout size, especially in the summer and fall (May–October) (Lyse et al. 1998; Keeley and Grant 2001; Knutsen et al. 2001; Sánchez-Hernández 2020). For instance, in coastal waters of western Scotland, Pemberton (1976) observed increased feeding on small fish among brown trout > 21 cm, especially in summer when clupeids such as Atlantic herring (Clupea harengus) and sand eels (Ammodytidae) were abundant. As mentioned previously, information on the marine diet and feeding behavior of anadromous cutthroat is limited relative to brown trout, however their diet is dominated by nearshore biota including small, schooling, planktivorous fishes, juvenile salmonids, and invertebrates (Pearcy et al. 2018). Stomach content analysis has revealed that food items with the greatest contribution by weight and number in the coastal ocean and Columbia River plume included herring, Pacific sand lance, anchovy and juvenile cabezon as well as crab megalope, euphausiids and mysiid shrimp (Brodeur et al. 1987; Loch and Miller 1988). Similarly, the diet of cutthroat captured in central Puget Sound was dominated by nearshore fishes throughout the year (Duffy and Beauchamp 2008). In the southern Puget Sound, where chum salmon are the most abundant anadromous fish species (Losee et al. 2019), Jauquet (2005) demonstrated the importance of their fry, flesh and eggs to the diet of cutthroat trout in marine waters. Similarly, Ellings et al. (2005) found that 90% of cutthroat trout sampled in tributaries of south Puget Sound during the chum salmon spawning period had diets dominated by chum salmon eggs. Similarly variable movement patterns to maximize seasonal prey resources was also described for brown trout, linking patterns of feeding and migration (Rikardsen et al. 2006; Boel et al. 2014; Bordeleau et al. 2018). Differences in diet by season and geographical area of both brown and cutthroat trout highlights the opportunistic nature of anadromous trout and are an important determinant of migration patterns. In this way, the presence and absence of anadromous trout may provide some insight into habitat quality and guide habitat improvement projects in the nearshore marine environment.
Maximum size may differ significantly between anadromous trout and other Pacific and Atlantic salmonids but maximum growth rates are often comparable between trout and salmon, and can exceed 1 mm per day for immature fish (Ricker 1981; Hartt and Dell 1986; Gonçalves et al. 2013). Specifically, Pearcy et al. (1990) documented growth rates as high as 2.6 mm a day for cutthroat trout, estimated by back-calculating ocean growth from scales. Similarly, Degerman et al. (2012) recorded growth rates up to 1.8 mm/day for brown trout marked and recaptured in Northern parts of the Baltic Sea. These high growth rates during the first summer at sea suggest that trout species, despite their small maximum size, enjoy the benefit of rapid growth during the early marine period documented for Pacific and Atlantic salmon (Hansen et al. 1998; Claiborne et al. 2011; Anderson et al. 2021). The rapid growth rate at a young age is likely an adaptation to avoid gape limited predators and improve body condition prior to over-wintering (Pearcy 1992; Beamish and Mahnken 2001; Thorstad 2016).
While growth rates of Pacific and Atlantic salmon and trout are rapid during the spring early marine period, a second phase of high growth rate and feeding occurs in the late fall for some trout populations (Knutsen et al. 2001; Rikardsen et al. 2006; Jonsson and Jonsson 2009a; Losee et al. 2018). For instance, Rikardsen et al. (2006) documented peak feeding rates, body condition and lipid content among brown trout captured in two fjords in Northern Norway (Ranafjord and Balsfjord) in August and September opposed to the typical spring period when anadromous salmon and trout first enter salt water. A similar pattern was documented among cutthroat sampled in southern Puget Sound over a 12 month period (Losee et al. 2018). This period of accelerated growth was associated with nutrient subsidies from spawning chum salmon for cutthroat trout, and increased abundance and access to clupeids, sand eels and crustaceans (e.g. shrimp and benthic amphipods) in brown trout (Haluch and Skora 1997; Rikardsen et al. 2006), again highlighting the highly opportunistic nature of anadromous trout and their ability to overwinter in marine water successfully.
Abundance and threats
Despite the scarcity of comprehensive population size assessments for anadromous trout, declines in abundance have been documented across the respective ranges of cutthroat (Giger 1972; Jones 1975; Hooton 1997; Johnson et al. 1999; Pearcy et al. 2018) and brown trout (ICES 2013; Pedersen et al. 2017). Despite these declines in abundance of for both species in the 1980’s and 1990’s, trends may have stabilized in recent decades. For instance, catches of anadromous brown trout on the west coast of Ireland (Connemara district) declined from approximately 10,000 fish annually prior to the 1990’s to less than 1,000 per year in the past 3 decades (Fig. 7). This unique data set reveals trends similar to those observed from of catch from other areas of Ireland and shows a consistent pattern across the country (Gargan et al. 2006). Additionally, in northern Wales, Davidson et al. (2007) documented a relatively stable anadromous brown trout run over more than 10 years based on counts of upstream migrating adults in the River Dee, beginning in the early 1990’s, but there were no estimates prior to the 1990’s. In the Baltic Sea, trends in anadromous brown trout abundance have declined relative to historic levels consistent with other areas (Pedersen et al. 2017; Dêbowski 2018), and many populations are vulnerable or have uncertain status (ICES 2021). However, HELCOM (2018) reported increasing abundance in four Swedish rivers (Kalixälven, Piteälven, Byskeälven and Vindelälven) following changes to fisheries practices in response to historic low levels in the 1990’s as was evident in River Umeälven where fish primarily from the tributary Vindelälven are counted (Fig. 7). In places where Baltic Sea populations remain at low levels such as the Gulf of Bothnia, Southern Baltic Sea and those originating in Germany, changes in fishing regulations that would result in reduced exploitation have been recommended (ICES 2021).

Trends in abundance for anadromous brown trout (upper tiles in grey) and anadromous cutthroat trout (lower tiles in black) from trapping locations across the range of distribution. Note that Y axis are not consistent between tiles
Similarly, several populations of anadromous cutthroat trout declined in the 1980’s and 1990s but information on abundance is limited overall. Catch of anadromous cutthroat trout from traps in tributaries of Puget Sound (Minter Creek), the Columbia River (Kalama River) and the Oregon Coast (Umpqua River) demonstrated a significant decline followed by a recent period of stability at lower abundances (Fig. 7). Additionally, fish managers in Puget Sound, Washington have used redd surveys to assess adult abundance in small streams (Losee et al. 2017a). Redd counts ranged from 50 to 150 over the last 15 years but estimates of redd abundance prior to 2006 are not available. Together, the trends observed for anadromous brown and cutthroat trout highlight the importance of long term time series to avoid the masking of past declines (i.e., shifting baseline (Pauly 1995)) and the importance of the late 1980’s and 1990’s in altering baseline abundance levels for many populations of anadromous fish. Increased use of redd surveys may be an economical approach to estimating abundance of brown trout across a broader range as has been done for cutthroat trout.
Leading causes cited for the declines observed for both brown and cutthroat trout are consistent with other anadromous salmonids and include habitat degradation, anthropogenic barriers (e.g. dams, hydropower operation etc.), hatchery production and harvest (Trotter et al. 1993; Lichatowich et al. 2006; Skaala et al. 2014; HELCOM 2018). The mechanisms that lead to association between anadromous trout abundance and these constraints are described well in (Connolly 1996; Johnson et al. 1999; Lichatowich 1999; Costello 2008; ICES 2013; Williams et al. 2015). Other less understood factors include predation, parasite infections, hybridization, and climate related factors, which we discuss here. While predation can affect population size of anadromous trout to some extent, rates are typically low (Lyse et al. 1998; Lance et al. 2012). Specifically, aquatic mammals and birds feed on anadromous trout, especially at certain life stages and habitats (Collis et al. 2002; Hansson et al. 2018; Penaluna et al. 2021; Källo et al. 2023). For example, Hansson et al. (2018) indicates seals and cormorants have been the target of research and public scrutiny in the Baltic Sea especially given the increased abundance by both. However, recent evidence that cormorants eat few anadromous trout suggests additional information is needed to understand the role of predators in regulating trout abundance (Boström et al. 2009, 2012). Additionally, parasite induced mortality likely represents an important factor for anadromous trout, particularly for brown trout in Norway (Thorstad et al. 2015). The best documented source of parasite associated mortality is the parasitic copepod (Lepeophtheirus salmonis) commonly known as “sea louse,” with the highest infection levels typically associated with areas supporting commercial net pen operations. In a review of the effects of sea lice on anadromous brown trout, Thorstad et al. (2015) provided evidence for a direct relationship between net pen operation, associated infections of sea louse, and declining abundance of anadromous brown trout in European marine waters. Negative effects of copepod infestations include secondary infections (Johnson et al. 2004), changes in behavior (Gjelland et al. 2014; Halttunen et al. 2018), and reduced survival (Gargan et al. 2003). For coastal cutthroat trout, the effect of sea lice is much less understood but presence, abundance and intensity have been documented (Hoffman 1999; Losee et al. 2020). Of particular interest is the parasitic argulid, Argulis pugettensis that appears to specialize on coastal cutthroat trout. Argulids feed on the flesh of their host and have caused premature mortality of their host, therefore infections of A. pugettensis likely represents a threat to coastal cutthroat trout. While investigations of ectoparasites in cutthroat have increased in recent years including an online angler reporting tool produced by WDFW and the Coastal Cutthroat Coalition (www.coastalcutthroatcoalition.com) (Losee et al. 2022), to our knowledge, no study comparing movement patterns or survival of individuals with differing parasite loads has been conducted. This represents an important data gap, given the negative effects of these infections described on anadromous brown trout, the current/observed prevalence of ectoparasites on cutthroat hosts, and the increased interest in commercial aquaculture in habitats occupied by cutthroat trout that have been linked to increased ectoparasite loads (https://www.fisheries.noaa.gov/west-coast/aquaculture/aquaculture-west-coast).
Hybridization with closely related salmonids that overlap spatially and temporally may be a threat for both brown and cutthroat trout (Matthews et al. 2000; Al-Chokhachy et al. 2014) as briefly discussed earlier. The fossil record suggests strong geographic separation (ca. 8 mya) of cutthroat trout from their closest relative, rainbow trout (Smith and Stearley 2018). Despite this separation, hybridization between these species occurs across the range (Campton and Utter 1985; Ostberg et al. 2004; Baumsteiger et al. 2005; Williams et al. 2007), though documentation of hybrids surviving to maturity is limited. However, over the last two decades numerous studies have described patterns of hybridization and hybrid swarms where rainbow trout and cutthroat overlap geographically, particularly for the subspecies of cutthroat that did not co-evolve with rainbow trout (Rubidge and Taylor 2004; Metcalf et al. 2008). In Washington State, U.SA., Moore et al. (2010) showed that cutthroat/steelhead hybrids had an intermediate migration pattern in marine water relative to parent species and suggested that this conferred a survival disadvantage. Like cutthroat, brown trout hybridization is relatively common when sympatric with Atlantic salmon (Makhrov 2008; Adams et al. 2014) but has been selected against because hybrids rarely reach adulthood (Solomon and Child 1978; Verspoor and Hammart 1991) or are sterile (Palm et al. 2013). This reduced fitness may exert selective pressure on size and behavioral patterns associated with habitat use and spatial and temporal patterns of reproduction for both brown and cutthroat trout. The role and effect of hybrids for anadromous trout is an area ripe for future research. By assessing rates of hybridization and associated survival across all life stages, the population level effect of hybridizing with closely related species would be clearer. Until more is known, managers should preserve diverse habitat types and limit interaction with artificially propagated trout species, to limit hybridization and support natural mechanisms that optimize fitness for brown and cutthroat trout in their native range.
Climate related factors play a major role in regulating abundance (e.g., flows during incubation and rearing stages, temperatures in summer and winter, etc.) and natural fluctuations in the environment have been linked to variability in salmon returns for decades (Lichatowich 1999; Quinn 2018). Anthropogenic changes in climate are a major threat for fish across the globe and is expected to exacerbate many of the threats and challenges highlighted above, especially for mid-high latitude species that need multiple habitat types to complete their life cycle, like anadromous trout (Bernstein et al. 2008; Williams et al. 2015; Jonsson et al. 2016; Mottola et al. 2020). Specifically, in northern Europe and North America, increased frequency of extreme weather events is expected, coupled with wetter winters and dryer, hotter summers, and less productive marine conditions. Together, these forecasted changes should be expected to intensify current problems and reduce productivity of fish populations that are adapted to historic conditions (Bernstein et al. 2008; Jonsson and Jonsson 2009b, ICES 2013; Freeman In review.). In contrast, the flexibility in life history for anadromous trout (stream, lake, marine waters, iteroparity etc.) and resilience associated with a diverse life history may convey some benefits in a changing climate.
Management
Status and threats are similar for anadromous trout across much of their native range and recognition of their economic value associated with sport fisheries is shared among management agencies. However, approaches to their conservation and recovery vary and management plans specific to anadromous trout are rare, resulting in diverse regulations that are not linked to population performance. One example of a place where management plans do exist is for anadromous rainbow trout (steelhead). Across the range, many steelhead are monitored and regulations may be changed within the fishing season based on estimated abundance. In contrast, in the case of cutthroat trout, there are typically neither abundance estimates nor management plans contingent on them. In European countries, some level of commercial harvest of anadromous brown trout is permitted—primarily in marine waters using commercial traps and nets but also in some lake systems on a smaller scale. In the Scandinavian countries, commercial and recreational gill net fishing is extensive in some coastal areas, and brown trout are either directly targeted by the fishery or caught as bycatch (e.g. (Kuhn et al. 2022)). For anadromous cutthroat trout, low abundance and small size, relative to Pacific salmon species, precludes commercial fishing but some limited ceremonial and subsistence food fisheries occur among Indigenous peoples. Altering harvest regulations for commercial fisheries has shown promise in recovering wild trout populations in some areas of the United Kingdom and Baltic Sea as was observed in the early 2000’s (Fig. 7) (Jutila et al. 2006; ICES 2012). However, given the limited scale of commercial fisheries targeting wild anadromous trout today, recent focus from management agencies has been directed towards sport fisheries. For both anadromous brown and cutthroat trout, sport fishing regulations rely on a diversity of harvest control rules and gear restrictions as well as limitations on time and place to harvest. Broadly, regulations include minimum size limits and maximum bag limits for harvest, depth restrictions to avoid encounters of larger individuals and restriction of the use of bait to limit encounters and critical hooking locations such as the gills and throat (Pauley and Thomas 1993) however both commercial and sport regulations across the range of brown and cutthroat have been and are currently extremely diverse and often inconsistent within a management unit (Johnson and Bjornn 1975; Johnson et al. 1999; Guðbergsson 2014; ICES 2020; Government 2023). For instance, in marine waters of Washington State, USA, fishing regulations have changed considerably from the 1970’s when the daily harvest limit was 16 fish, to the current prohibition of harvest of cutthroat in marine waters as a response to declining abundance (Fig. 8). This change to “catch and release only” regulations in marine waters contrasts with diverse regulations in Washington fresh waters that range from conservative catch and release regulations to more liberal harvest regulations including a two fish daily harvest limit of fish over 8 inches (20 cm). Similarly, Along the northwestern coast of the Baltic Sea, a ban on gillnets shallower than 3 m has been enforced to protect declining stocks of anadromous brown trout (Degerman et al. 2015), however harvest is permitted in some nearby waters. Furthermore, formal evaluations of management strategies are uncommon, in part due to the lack of management plans that provide well-defined management objectives (ICES 2020), policy guidance and decision frameworks (i.e., regulations contingent on abundance estimates) for managers. This is concerning because the mixed stocks (Degerman et al. 2012; Losee et al. 2017b) and nearshore marine distribution make them vulnerable to overexploitation (Blyth and Rönnbäck 2022). Importantly, the regulations and management regimes have not adapted to the increasing scientific knowledge of life-history complexity and movement patterns and remain inconsistent and lacking formal management strategy evaluation or policy guidance.

Fishing regulations for cutthroat trout in marine waters of Washington State, USA between 1955 and 2022
Both anadromous brown and cutthroat trout are targeted by fishers across their respective ranges. To support long term-sustainable fisheries, managers rely heavily on fishing regulations. To our knowledge a comprehensive understanding of fishing regulations and their effect on abundance of anadromous trout does not exist and represents an area for future research. However, some small scale studies with limited geographic scope have been conducted (Pauley and Thomas 1993; Solomon et al. 2007; Losee et al. 2017b; Skov et al. 2022) and suggest that conservative regulations (e.g. catch and release) in all environments occupied by resident and anadromous life histories can increase catch rates in marine waters, given the close association of resident and anadromous forms as described previously. Additionally, involving anglers in conservation and recovery work has been proven effective to achieve shared goals associated with fishing opportunity and protection of the resource and should be explored more broadly (Shephard et al. 2023). Overall, fishing regulations that consider life history diversity, such as those that protect juveniles, kelts and individuals migrating at the beginning and end of the typical run, combined with improved passage and habitat, are the most important tools to support the long-term recovery and sustainability of anadromous trout as has been adopted in the Shetland Islands and elsewhere (King et al. 2021). In the short term, catch and release regulations maximize fishing opportunity while minimizing impact to wild trout, given high rates of recapture, low rates of release mortality, high economic revenue associated with fishing, and anglers’ willingness to accept stricter regulations to protect the opportunity to fish (Elliot 1989; Degerman et al. 2012; Lothrop and Losee 2016; Liu et al. 2019).
Hatchery production has been applied broadly to management of anadromous brown trout to support fisheries and conservation goals. In the Shetland Islands of the United Kingdom, King et al. (2021) used genetic analysis to evaluate the contribution of hatchery produced brown trout to the total trout catch in the marine environment. They found that release of hatchery brown trout had little to no positive effect on the fishery, and highlighted environmental conditions and habitat quality, including fish passage, as the most important factors affecting wild trout population restoration and improved catch rates. Regardless, releases of hatchery produced brown trout continue across the range to improve catch rates in lakes, rivers and marine waters and in some cases to bolster declining wild populations. In contrast, the production of anadromous cutthroat has received limited attention from managers since the 1970’s when cutthroat trout programs were extensively evaluated (Jones 1977; Mercer and Johnston 1980; Mercer 1982; Rempel 1984; Tipping 1986). Those studies showed low survival rates among smolts released, and limited return to the fishery relative to wild cutthroat trout. Today, hatchery programs producing anadromous cutthroat trout are uncommon with stocking limited to British Columbia (Slaney et al. 1996; Costello 2008), and in Washington on the Columbia River (Tipping and Blankenship 1993; Tipping 2001). These programs are very small relative to salmon and steelhead hatchery programs. This contrast may be important when considering factors affecting wild cutthroat trout versus other species. Consequently, the scientific focus on genetic, ecological, and fishery conflicts between wild and hatchery-produced or farmed salmonids that is so prominent for Atlantic and Pacific salmon (Naish et al. 2007) is less relevant for anadromous brown and especially cutthroat trout.
Habitat protection and restoration represents an area of increasing importance for fisheries managers responsible for anadromous trout. Their use of small streams makes anadromous trout vulnerable to passage barriers such as poorly designed or maintained road culverts, and this is increasingly recognized as a priority area for salmonid conservation (Price et al. 2010; Januchowski‐Hartley et al. 2014), especially considering the effect freshwater conditions can have on marine survival and adult abundance of salmonids. For instance Flávio et al. (2020) demonstrated a high degree of mortality of Atlantic salmon in Northern Ireland, occurring in lower river areas that could have been mitigated through simple habitat-based management actions (e.g. improvement to riparian and in-river habitat complexity), potentially decreasing marine mortality, assuming compensatory mortality is low. Similarly, Losee et al. (2019) analyzed trends in survival rates across six species of hatchery and natural origin Pacific salmon in Washington State and reported the greatest declines occurring for those species that depend on extended rearing in freshwater (e.g. coho salmon and steelhead). There is a growing body of evidence from these and other studies that highlight the potential significant benefit from improved freshwater habitat conditions as a step toward improving marine survival rates and adult returns for anadromous salmon and trout. Analytical approaches that allow for forecasted recovery gains associated with freshwater habitat improvements such as Chasco et al. (2021) may provide managers and elected officials the incentive necessary to increase efforts to improve freshwater habitats.
In contrast to Atlantic salmon, Pacific salmon and steelhead, genetic baseline libraries for stock identification of anadromous brown trout and cutthroat trout have not yet been established and implemented in management and conservation work, however pioneering efforts have been conducted in the northern Baltic Sea and British North Sea coast (Bekkevold et al. 2021; Östergren et al. 2016) and have just begun in North America. Comprehensive genetic baselines for anadromous trout are challenging to establish given the often-numerous small populations occurring along the coasts. However, such work would provide a valuable tool for advancing current management work, especially in situations where mixed stock fisheries occur (as is generally the case for anadromous trout in the sea). For salmon, in some areas of North American and Europe, precise estimates of stock specific mortality are estimated for various stocks, aiming to protect small populations from overexploitation when intermingling with abundant populations in the presence of fisheries. Across the range of anadromous trout, population-specific management targets are seldom applied, although genetic conservation and an awareness of the genetic integrity of populations is being considered (Johnson et al. 1999; ICES 2020). As more sophisticated tools become available (genetics stock identification, stock specific forecasting of runs, etc.) and are applied to management of specific species of anadromous trout, information sharing by managers across the globe should lead to an acceleration of improved management techniques.
Finally, it should be mentioned that brown trout are among the most invasive species in the world (Budy et al. 2013). For example, the Rio Grande in Argentina supports one of the most abundant and diverse populations of anadromous brown trout in the world (O'Neal 2008). Where brown trout are not native, some eradication programs have been implemented to protect native fishes but have been largely unsuccessful. However, in other areas non-native populations of brown trout have been protected by management agencies and are highly prized for their economic value associated with fisheries, particularly for fluvial populations (Jones and Closs 2018). Interestingly, anadromous brown trout have been reported from watersheds also containing native coastal cutthroat (Idyll 1942; Bisson et al. 1986). Given the overlap in ecological niche and trophic position of the two species, sympatric existence would likely be to the detriment of native cutthroat, as has been seen in landlocked sympatric populations in the Yellowstone Lake system (Al‐Chokhachy and Sepulveda 2019). In their native range, introduced species have been identified as a risk to cutthroat and brown trout through predation, competition and genetic introgression and is expected to be exacerbated with an increasingly changing climate (Rahel and Olden 2008; Bae et al. 2018). Research on this topic has primarily been focused on brook trout, rainbow trout, round goby, and most recently invasive pink salmon for brown trout (Landergren 1999; Lehtonen 2002; Larranaga et al. 2019; Lutz et al. 2020; Staveley and Ahlbeck Bergendahl 2022). However, invasive pink salmon in western Europe provide some benefit through nutrient subsidies so more work is needed to understand the level of threat and appropriate response by managers (Dunlop et al. 2021).
Through improved stock assessment work, including genetic stock analysis and biologically relevant management objectives, a more refined approach to management of sport fisheries could be implemented across the range. However, given the complicated life history and numerous, small populations of brown and cutthroat trout, stock recruitment relationships may be applicable only on the stream where they were derived or very similar ones and so not broadly generalizable. Höjesjö et al. (2017) suggested that an approach consisting of many index areas across a broad range of habitat types and geographic distribution may provide managers with an understanding of changes in abundance across the range. This, combined with broad conservative regulations that protect both small, understudied populations and those that are large and receive monitoring efforts could provide a reliable management approach compared to stream-specific regulations that are not based on high-quality data. Together, this review highlights the need for fishery managers and recovery biologists to focus on improving conditions in freshwater for anadromous trout, combined with protective fishing regulations. Habitat improvement projects that promote diversity to buffer extreme weather events, maximize available spawning and rearing habitat, prioritize production of juvenile trout and encourage segregation between natural and hatchery origin fish, are likely to provide the greatest chance of restoring anadromous trout abundance to historic levels and support sustainable fisheries.
Data gaps
By applying a comparative approach to the review of anadromous brown and cutthroat trout, this work has revealed numerous data gaps for two similar species of anadromous trout from opposite sides of the globe. Areas for future research include increased sampling of trout in the marine water for diet and condition throughout the year, identification of genetic sex markers, improved understanding of the role of freshwater conditions in determining rates of marine survival, and investigation into site fidelity of brown trout in the marine environment. Additionally, this review has identified a number of data gaps that should be considered high priority to support long term sustainability of anadromous brown and cutthroat trout. Most conspicuous is the lack of good estimates of abundance patterns and trends across a range of rivers. With a greater understanding of stock abundance and status, more refined management approaches could be utilized to support the recovery of anadromous trout in some places and improved fishing opportunity in others. In some areas, traditional tools like redd counts, riverscapes, tagging (Floy, elastomer and PIT tag technology) and genetics, have been applied and tailored specifically for anadromous trout given their diverse life histories providing great promise for these two species and the management around them. Additionally, new tools being developed for other focal species will be available to apply to anadromous trout in the near future including SONAR (Gaudet 1990; Connolly et al. 2022), seismic monitoring (Dietze et al. 2020) and drones (Groves et al. 2016) to improve estimates of abundance.
Some of the most important knowledge gaps that exist, in addition to lack of abundance estimates, is an understanding of the underlying mechanisms driving life history variation and migration patterns of anadromous trout. This information would support more focused conservation efforts aimed at identifying limiting factors to abundance and resilience. If researchers look to the study designs and results that have emerged from the brown trout research across Europe, the state of knowledge for coastal cutthroat trout could accelerate, particularly with the utilization of otolith microchemistry and acoustic telemetry. In the same way, the propensity for brown trout to exhibit high site fidelity in the marine environment is unknown but likely occurring and would have important implications for fisheries managers in identifying important habitat. This is one recent advancement in the understanding of fine scale movements for anadromous cutthroat that could lead to a novel study design for anadromous brown trout, and to test hypotheses related to migration patterns, variation in life-history types, population abundance and vulnerability to fisheries in the marine environment among others. Together, an improved understanding of environmental and genetic factors explaining growth and performance would clarify the tradeoffs between various migratory patterns exhibited by anadromous trout.
Along with anadromous brown and cutthroat trout, numerous other closely related species exhibit similar behavior but suffer from the same lack of information needed to support sustainable fisheries management. Nearshore distributions of anadromous grayling Thymallus thymallus (Müller and Karlsson 1983; Hellström et al. 2023) and different species of char have been documented, including brook trout, Salvelinus fontinalis referred to as “coasters” and “salters”(Morinville and Rasmussen 2006), bull trout, S. confluentus (Hayes et al. 2011; Goetz et al. 2021), Dolly Varden, S. malma (Bond et al. 2013) and Arctic charr, S. alpinus (Mulder et al. 2020), though some of these species may also occupy offshore waters. Each of these facultatively anadromous species shows some similarities with brown and cutthroat trout as described here, hence future research may highlight important aspects of the behavior and biology of any one species that can help understand and refine management approaches of others.
Conclusion
This review aims to inform sustainable fisheries management of anadromous cutthroat and brown trout and aid in the progression of science relating to anadromous trout and char across the globe. Research is limited on anadromous trout, but these unique salmonids, with their use of many small, short coastal streams for spawning, restricted migrations at sea, and occupancy of primarily nearshore marine habitats, require a unique approach to management and conservation. Recent research in protected waters of the North Atlantic Ocean has increased understanding of abundance, maximum body size, maximum migration distance, feeding, ectoparasite infections, sexual dimorphism and potential for individuals to spawn in brackish marine waters. For anadromous cutthroat trout, these aspects remain poorly understood across their range and represent important data gaps, as are the major sources of mortality and especially the key predators. Recent developments in fish tracking technology, stable isotope analysis, and otolith chemistry have provided researchers with an improved toolbox to address these challenges, and researchers in Europe adopted these techniques and have been leading their application to anadromous trout. Recent studies have revealed spatially and temporally diverse migration patterns, thus the classic pattern “enter the marine environment in the spring and return to freshwater in the fall” is only one of many patterns and not always the most common one. As researchers use the current review as a roadmap for future work, managers should seek ways to incorporate less understood aspects of salmonid life history such as iteroparity, diverse river and marine entry timings and production of anadromous offspring by resident parents into management frameworks tailored specifically to anadromous trout and remain aware of new findings that separate anadromous trout from their semelparous relatives.
Brown trout and cutthroat trout that spawn near marine waters rely on diverse life history strategies spanning the freshwater, estuary and marine environments adding to their resilience in the face of a changing climate and increasing human population growth. Due to this diversity and broad geographic range the challenges anadromous trout face are multifaceted as highlighted in this review. It is evident that anadromous trout, with their opportunistic nature, have taken advantage of a unique niche resource to maximize growth, minimize predation and competition and adapt to variable environments in both the marine and freshwater ecosystems. With declining abundance, and the absence of anadromous trout specific management plans across most of their native range, managers should, in the short term, focus on improving connection between diverse habitat types given the benefit of removing anthropogenic barriers and estuary habitat improvement projects have on marine survival and ultimate adult abundance. Longer term goals should include changing to conservative fishing regulations in some areas that protect resident and anadromous forms and development of comprehensive anadromous trout management plans to provide a pathway to rebuilding anadromous trout populations to historic abundance and diversity while protecting those that are performing well.
Availability of data and materials
The data presented are available upon request.
References
Adams C, Burrows A, Thompson C, Verspoor E (2014) An unusually high frequency of Atlantic salmon x brown trout hybrids in the Loch Lomond catchment, west-central Scotland. The Glasglow Natl 26:75–81
Ahn S, De Steiguer JE, Palmquist RB, Holmes TP (2000) Economic analysis of the potential impact of climate change on recreational trout fishing in the southern Appalachian Mountains: an application of a nested multinomial logit model. Clim Change 45:493–509
Al-Chokhachy R, Sepulveda AJ (2019) Impacts of nonnative brown trout on Yellowstone cutthroat trout in a tributary stream. N Am J Fish Manag 39:17–28
Al-Chokhachy R, Muhlfeld CC, Boyer MC, Jones LA, Steed A, Kershner JL (2014) Quantifying the effectiveness of conservation measures to control the spread of anthropogenic hybridization in stream salmonids: a climate adaptation case study. N Am J Fish Manag 34:642–652
Alt DD, Hyndman DW (1995) Northwest exposures: a geologic story of the Northwest. Mountain Press
Anderson AJ, Claiborne AM, Agha M, Litz MN (2021) Puget sound chum salmon growth linked to competitor abundance, climate indices, and copepod species richness. Trans Am Fish Soc 150:707–729
Anlauf-Dunn K, Kraskura K, Eliason EJ (2022) Intraspecific variability in thermal tolerance: a case study with coastal cutthroat trout. Conserv Physiol 10:coac029
Archer LC, Hutton SA, Harman L, McCormick SD, O’Grady MN, Kerry JP, Poole WR, Gargan P, McGinnity P, Reed TE (2020) Food and temperature stressors have opposing effects in determining flexible migration decisions in brown trout (Salmo trutta). Glob Change Biol 26:2878–2896
Arostegui MC, Quinn TP (2019) Reliance on lakes by salmon, trout and charr (Oncorhynchus, Salmo and Salvelinus): An evaluation of spawning habitats, rearing strategies and trophic polymorphisms. Fish Fish 20:775–794
Bachman RA (1984) Foraging behavior of free-ranging wild and hatchery brown trout in a stream. Trans Am Fish Soc 113:1–32
Bae M-J, Murphy CA, García-Berthou E (2018) Temperature and hydrologic alteration predict the spread of invasive Largemouth Bass (Micropterus salmoides). Sci Total Environ 639:58–66
Baumsteiger J, Hankin D, Loudenslager EJ (2005) Genetic analyses of juvenile steelhead, coastal cutthroat trout, and their hybrids differ substantially from field identifications. Trans Am Fish Soc 134:829–840
Beacham TD, Murray CB (1987) Adaptive variation in body size, age, morphology, egg size, and developmental biology of chum salmon (Oncorhynchus keta) in British Columbia. Can J Fish Aquat Sci 44:244–261
Beamish RJ, Mahnken C (2001) A critical size and period hypothesis to explain natural regulation of salmon abundance and the linkage to climate and climate change. Prog Oceanogr 49:423–437
Behnke RJ (2002) Trout and Salmon of North America. The Free Press, New York
Behnke RJ (1992) Native trout of western North America. Monograph 6, American Fisheries Society
Bekkevold D, Piper A, Campbell R, Rippon P, Wright RM, Crundwell C, Wysujack K, Stevens JR, King RA, Aarestrup K, Maltby A (2021) Genetic stock identification of sea trout (Salmo trutta L.) along the British North Sea Coast shows prevalent long-distance migration. ICES J Mar Sci 78:952–966
Berg OK, Jonsson B (1989) Migratory pattern of anadromous Atlantic salmon, brown trout and Arctic charr from the Vardnes river in northern Norway. In: Proceedings of the Salmonid Migration and Distribution Symposium, 1989. University of Washington
Berg O, Berg M (1987) Migrations of sea trout, Salmo trutta L., from the Vardnes river in northern Norway. J Fish Biol 31:113–121
Berg OK, Jonsson B (1990) Growth and survival rates of the anadromous trout, Salmo trutta, from the Vardnes River, northern Norway. Environ Biol Fishes 29:145–154
Bernatchez L (2001) The evolutionary history of brown trout (Salmo trutta L.) inferred from phylogeographic, nested clade, and mismatch analyses of mitochondrial DNA variation. Evolution 55:351–379
Bernatchez L, Hendry A, Stearns S (2004) Evolution illuminated: salmon and their relatives
Bernstein L, Bosch P, Canziani O, Chen Z, Christ R, Davidson O, Hare W, Huq S, Karoly DJ, Kattsov V (2008) Climate change 2007 synthesis report. Intergovernmental Panel on Climate Change
Birnie-Gauvin K, Aarestrup K (2019) A call for a paradigm shift: assumed-to-be premature migrants actually yield good returns. Ecol Freshw Fish 28:62–68
Birnie-Gauvin K, Koed A, Aarestrup K (2021) Repeatability of migratory behaviour suggests trade-off between size and survival in a wild iteroparous salmonid. Funct Ecol 35:2717–2727
Bisson PA, Nielsen JL, Chilcote MW, Crawford B, Leider SA (1986) Occurrence of anadromous brown trout in two lower Columbia river tributaries. N Am J Fish Manag 6:290–292
Blicharska M, Rönnbäck P (2018) Recreational fishing for sea trout—Resource for whom and to what value? Fish Res 204:380–389
Blyth S, Rönnbäck P (2022) To eat or not to eat, coastal sea trout anglers’ motivations and perceptions of best practices for catch and release. Fish Res 254:106412
Boel M, Aarestrup K, Baktoft H, Larsen T, Søndergaard Madsen S, Malte H, Skov C, Svendsen JC, Koed A (2014) The physiological basis of the migration continuum in brown trout (Salmo trutta). Physiol Biochem Zool 87:334–345
Bohlin T, Dellefors C, Faremo U (1996) Date of smolt migration depends on body-size but not age in wild sea-run brown trout. J Fish Biol 49:157–164
Bond MH, Quinn TP, Jonsson B (2013) Patterns and influences on Dolly Varden migratory timing in the Chignik Lakes, Alaska, and comparison of populations throughout the northeastern Pacific and Arctic oceans. Can J Aquat Sci 70:655–665
Bordeleau X, Davidsen JG, Eldøy SH, Sjursen AD, Whoriskey FG, Crossin GT (2018) Nutritional correlates of spatiotemporal variations in the marine habitat use of brown trout (Salmo trutta) veteran migrants. Can J Fish Aquat Sci 75:1744–1754
Boström MK, Lunneryd S-G, Karlsson L, Ragnarsson B (2009) Cormorant impact on trout (Salmo trutta) and salmon (Salmo salar) migrating from the river Dalälven emerging in the Baltic Sea. Fish Res 98:16–21
Boström MK, Östman Ö, Bergenius MAJ, Lunneryd S-G (2012) Cormorant diet in relation to temporal changes in fish communities. ICES J Mar Sci 69:175–183
Brodeur RD, Lorz HV, Pearcy WG (1987) Food habits and dietary variability of pelagic nekton off Oregon and Washington, 1979–1984
Budy P et al (2013) Limitation and facilitation of one of the world’s most invasive fish: an intercontinental comparison. Ecology 94(2):356–367
Buehrens TW, Glasgow J, Ostberg CO, Quinn TP (2013) Spatial segregation of spawning habitat limits hybridization between sympatric native steelhead and coastal cutthroat trout. Trans Am Fish Soc 142:221–233
Burns R (1985) The shape and form of Puget Sound. University of Washington Press, Sea Grant Program, Seattle
Campton DE, Utter FM (1985) Natural hybridization between steelhead trout (Salmo gairdneri) and coastal cutthroat trout (Salmo clarki clarki) in two Puget Sound streams. Can J Fish Aquat Sci 42:110–119
Campton DE, Utter FM (1987) Genetic structure of anadromous cutthroat trout (salmo clarki clarki) populations in the Puget Sound area: evidence for restricted gene flow. Can J Fish Aquat Sci 44:573–582
Chapman BB, Brönmark C, Nilsson J-Å, Hansson L-A (2011) The ecology and evolution of partial migration. Oikos 120:1764–1775
Chapman BB, Hulthén K, Brodersen J, Nilsson PA, Skov C, Hansson L-A, Brönmark C (2012) Partial migration in fishes: causes and consequences. J Fish Biol 81:456–478
Chasco B, Burke B, Crozier L, Zabel R (2021) Differential impacts of freshwater and marine covariates on wild and hatchery Chinook salmon marine survival. PLoS ONE 16:e0246659
Claiborne AM, Fisher JP, Hayes SA, Emmett RL (2011) Size at release, size-selective mortality, and age of maturity of Willamette River hatchery yearling Chinook salmon. Trans Am Fish Soc 140:1135–1144
Claiborne AM, Losee JP, Miller JA (2020) Estimating migratory behavior and age for anadromous coastal cutthroat trout in south puget sound: evaluation of approaches based on fish scales versus otoliths. N Am J Fish Manag 40:1313–1323
Collis K, Roby DD, Craig DP, Adamany S, Adkins JY, Lyons DE (2002) Colony size and diet composition of piscivorous waterbirds on the lower Columbia River: implications for losses of juvenile salmonids to avian predation. Trans Am Fish Soc 131:537–550
Connolly PJ (1996) Resident cutthroat trout in the central Coast Range of Oregon: logging effects, habitat associations, and sampling protocols. Oregon State University, Corvallis, OR
Connolly RM, Jinks KI, Shand A, Taylor MD, Gaston TF, Becker A, Jinks El (2022) Out of the shadows: automatic fish detection from acoustic cameras. Aquat Ecol
Costello AB (2008) The status of coastal cutthroat trout in British Columbia. In: The 2005 Coastal cutthroat trout symposium, p 24
Crawford SS, Muir AM (2008) Global introductions of salmon and trout in the genus Oncorhynchus: 1870–2007. Rev Fish Biol Fisheries 18:313–344
Crete-Lafreniere A, Weir LK, Bernatchez L (2012) Framing the Salmonidae family phylogenetic portrait: a more complete picture from increased taxon sampling. PLoS ONE
Davidson I, Cove R, Hazlewood M (2007) Annual variation in age composition, growth and abundance of sea trout returning to the River Dee at Chester. Sea Trout: Biology, Conservation and Management, p 76
Dêbowski P (2018) The largest Baltic population of sea trout (Salmo trutta L.): its decline, restoration attempts, and current status. Fish Aquat Life 26:81–100
Degerman E, Leonardsson K, Lundqvist H (2012) Coastal migrations, temporary use of neighbouring rivers, and growth of sea trout (Salmo trutta) from nine northern Baltic Sea rivers. ICES J Mar Sci 69:971–980
Degerman E, Näslund I, Kagervall A, Östergren J (2015) Anadromous brown trout—a challenge for management. Swedish University of Agricultural University (in Swedish)
Denton EJ, Nicol JAC (1966) A survey of reflectivity in silvery teleosts. J Mar Biol Assoc UK 46:685–722
Desforges J, Birnie-Gauvin K, Aarestrup K, Cooke SJP (2021) Upper thermal tolerance indicated by CTmax fails to predict migration strategy and timing, growth, and predation vulnerability in juvenile brown trout (Salmo trutta). Physiol Biochem Zool 94:215–227
Dieperink C, Pedersen S, Pedersen MI (2001) Estuarine predation on radiotagged wild and domesticated sea trout (Salmo trutta L. ) smolts. Ecol Freshw Fish 10:177–183
Dietze M, Losee J, Polvi L, Palm D (2020) A seismic monitoring approach to detect and quantify river sediment mobilization by steelhead redd-building activity. Earth Surf Processes 45:2840–2849
Duffy EJ, Beauchamp DA (2008) Seasonal patterns of predation on juvenile Pacific salmon by anadromous cutthroat trout in Puget Sound. Trans Am Fish Soc 137:165–181
Dunlop K, Eloranta AP, Schoen E, Wipfli M, Jensen JL, Muladal R, Christensen GN (2021) Evidence of energy and nutrient transfer from invasive pink salmon (Oncorhynchus gorbuscha) spawners to juvenile Atlantic salmon (Salmo salar) and brown trout (Salmo trutta) in northern Norway. Ecol Freshw Fish 30:270–283
Duval E, Skaala Ø, Quintela M, Dahle G, Delaval A, Wennevik V, Glover KA, Hansen MM (2021) Long-term monitoring of a brown trout (Salmo trutta) population reveals kin-associated migration patterns and contributions by resident trout to the anadromous run. BMC Ecol Evol 21:143
Eldøy SH, Davidsen JG, Thorstad EB, Whoriskey FG, Aarestrup K, Næsje T, Rønning L, Sjursen AD, Rikardsen AH, Arnekleiv JV (2017) Marine depth use of sea trout Salmo trutta in fjord areas of central Norway. J Fish Biol 91:1268–1283
Eldøy SH, Bordeleau X, Crossin GT, Davidsen JG (2019) Individual repeatability in marine migratory behavior: a multi-population assessment of anadromous brown trout tracked through consecutive feeding migrations 7
Ellings CS, Cederholm CJ, Chin-Leo G (2005) The influence of spawning pacific salmon on the stable isotope composition, feeding behavior, and caloric intake of coastal cutthroat trout. In: The 2005 coastal cutthroat trout symposium, p 47
Elliot JM (1989) Wild brown trout Salmo trutta: an important national and international resource. Freshw Biol 21:1–5
Emeis K-C, Struck U, Blanz T, Kohly A, Voβ M (2003) Salinity changes in the central Baltic Sea (NW Europe) over the last 10000 years. The Holocene 13:411–421
Englmaier GK, Antonov A, Weiss SJ, Fisheries (2021) General patterns of sexual dimorphism in graylings (Thymallus), with a comparison to other salmonid species. Rev Fish Biol Fishe 1–23
Etheridge E, Harrod C, Bean C, Adams C (2008) Continuous variation in the pattern of marine v. freshwater foraging in brown trout Salmo trutta L. from Loch Lomond, Scotland. J Fish Biol 73:44–53
Ferguson A, Reed TE, Cross TF, McGinnity P, Prodöhl PA (2019) Anadromy, potamodromy and residency in brown trout Salmo trutta: the role of genes and the environment. J Fish Biol 95:692–718
Ferguson A (2006) Genetics of sea trout, with particular reference to Britain and Ireland. J Sea Trout Biol Conserv Manag 157–182
Flávio H, Kennedy R, Ensing D, Jepsen N, Aarestrup K (2020) Marine mortality in the river? Atlantic salmon smolts under high predation pressure in the last kilometres of a river monitored for stock assessment. Fish Manag Ecol 27:92–101
Freeman R, Losee JP, Madel GM In review. Environmental variables and their relationship with coastal cutthroat trout spawn timing
Gabrielsen S-E, Lennox RJ, Wiers T, Barlaup BT (2021) Saltwater spawning grounds of sea-run brown trout (Salmo trutta) in tidal waters of a major Norwegian river. Environ Biol Fishes 104:1207–1213
García-Vega A, Fuentes-Pérez JF, Leunda Urretabizkaia PM, Ardaiz Ganuza J, Sanz-Ronda FJ (2022) Upstream migration of anadromous and potamodromous brown trout: patterns and triggers in a 25-year overview. Hydrobiologia 849:197–213
Gargan P, Tully O, Poole W (2003) Relationship between sea lice infestation, sea lice production and sea trout survival in Ireland, 1992–2001
Gargan P, Poole W, GJSTB (2006) Forde conservation, and management. A review of the status of Irish sea trout stocks 25–44
Gaudet DM (1990) Enumeration of migrating salmon populations using fixed-location sonar counters. Rapports Et Proces-Verbaux Des Reunions, Conseil International Pour L’exploration De La Mer 189:197–209
Giger RD (1972) Ecology and management of coastal cutthroat trout in Oregon
Gjelland KØ, Serra-Llinares RM, Hedger RD, Arechavala-Lopez P, Nilsen R, Finstad B, Uglem I, Skilbrei OT, Bjørn PA (2014) Effects of salmon lice infection on the behaviour of sea trout in the marine phase. Aquac Environ Interact 5:221–233
Goetz FA, Baker B, Buehrens T, Quinn TP (2013) Diversity of movements by individual anadromous coastal cutthroat trout Oncorhynchus clarkii clarkii. J Fish Biol 83:1161–1182
Goetz FA, Beamer E, Connor EJ, Jeanes E, Kinsel C, Chamberlin JW, Morello C, Quinn TP (2021) The timing of anadromous bull trout migrations in estuarine and marine waters of Puget Sound, Washington. Environ Biol Fishes 104:1073–1088
Gonçalves JFM, Carraça S, Damasceno-Oliveira A, Vicente C, P. M. d. Costa, M. Lopes-Lima, and R. O. d. A. Ozório. (2013) Growth and osmoregulation in Salmo salar L. juveniles 1+, 1½+ and 2+ reared under restrained salinity. Scientia Agricola 70:12–20
Government BC (2023) 2023–2025 Freshwater fishing regulations synopsis. In: Fisheries, editor
Griswold KE, Currens KP, Reeves GH (1997) Genetic and meristic divergence of coastal cutthroat trout residing above and below barriers in two coastal basins. In: Hall JD, Bisson PA, Gresswell RE (eds) Sea-run cutthroat trout: biology, management, and future conservation. Oregon Chapter of the American Fisheries Society, pp 167–169
Groves PA, Alcorn B, Wiest MM, Maselko JM, Connor WP (2016) Testing unmanned aircraft systems for salmon spawning surveys. Facets 1:187–204
Gruchy C, Vladykov V (1968) Sexual dimorphism in anal fin of brown trout, Salmo trutta, and close relatives. J Fish Board Can 25:813–815
Guðbergsson G (2014) Catch statistics for Atlantic salmon, Arctic char and brown trout in Icelandic rivers and lakes 2013
Guy TJ, Gresswell RE, Banks MA (2008) Landscape-scale evaluation of genetic structure among barrier-isolated populations of coastal cutthroat trout, Oncorhynchus clarkii clarkii. Can J Fish Aquat Sci 65:1749–1762
Halttunen E, Gjelland K-Ø, Hamel S, Serra-Llinares R-M, Nilsen R, Arechavala-Lopez P, Skarðhamar J, Johnsen IA, Asplin L, Karlsen Ø, Bjørn P-A, Finstad B (2018) Sea trout adapt their migratory behaviour in response to high salmon lice concentrations. J Fish Dis 41:953–967
Haluch M, Skora K (1997) Food of sea trout [Salmo trutta m. trutta L.] caught in the Gulf of Gdansk in 1994–1995 [Baltic Sea]. Bull Sea Fish Inst
Hansen L, Quinn T, Sciences A (1998) The marine phase of the Atlantic salmon (Salmo salar) life cycle, with comparisons to Pacific salmon. Can J Fish 55:104–118
Hansson S, Bergström U, Bonsdorff E, Härkönen T, Jepsen N, Kautsky L, Lundström K, Lunneryd S-G, Ovegård M, Salmi J (2018) Competition for the fish–fish extraction from the Baltic Sea by humans, aquatic mammals, and birds. ICES J Mar Sci 75:999–1008
Haque SR (2008) Movement patterns of coastal cutthroat trout (Oncorhynchus clarki clarki) in South Puget Sound, Washington 2006–2007. Evergreen State College
Harff J, Hoth P (2011) The baltic sea basin: introduction. In: H J, B S, H P, (eds) The Baltic Sea Basin. Central and Eastern European Development Studies (CEEDES). Springer, Berlin, Heidelberg
Harris G, Milner N (2008) Sea trout: biology, conservation and management. John Wiley & Sons
Hartt AC, Dell MB (1986) Early oceanic migrations and growth of juvenile Pacific salmon and steelhead trout. Int North Pac Fish Comm Bull 46
Hayes MC, Rubin SP, Reisenbichler RR, Goetz FA, Jeanes E, McBride A (2011) Marine habitat use by anadromous bull trout from the Skagit River, Washington. Mar Coast Fish 3:394–410
Hayes SA, Hanson CV, Pearse DE, Bond MH, Garza JC, MacFarlane RB (2012) Should I stay or should I go? The Influence of Genetic Origin on Emigration Behavior and Physiology of Resident and Anadromous Juvenile Oncorhynchus Mykiss 32:772–780
Heggenes J, Salveit SJ (1990) Seasonal and spatial microhabitat selection and segregation in young atlantic salmon, (Salmo salar L.), and brown trout, (Salmo trutta), in a Norwegian river. J Fish Biol 36:707–720
Heggenes J, Northcote TG, Peter A (1991) Spatial stability of cutthroat trout (Oncorhynchus clarki) in a small, coastal stream. Can J Fish Aquat Sci 48:757–762
Heggenes J, Krog OMW, Lindås OR, Dokk JG (1993) Homeostatic behavioural responses in a changing environment: brown trout (Salmo trutta) become Nocturnal during Winter. J Anim Ecol 62:295–308
HELCOM (2018) Abundance of sea trout spawners and parr:HELCOM core indicator report. Online. 891
Hellström G, Palm D, Losee J, Sandström A, Persson L, Berglund J, Brodin T (2023) Behaviour and migration of anadromous grayling in the Baltic sea. in review
Hindar K, b. Jonsson, N. Ryman and G. Stahl. (1991) Genetic relationships among landlocked, resident, and anadromous brown trout, Salmo trutta L. Heredity 66:83–91
Hoffman GL (1999) Parasites of North American freshwater fishes. Cornell University Press
Höjesjö J, Aldvén D, Davidsen JG, Pedersen S, Degerman E (2017) Perspectives on sea trout stocks in Sweden, Denmark & Norway: monitoring, threats and management. In: 2nd international sea trout symposium. Troubador Publishing, pp 443–459
Honda K, Arai T, Kobayashi S, Tsuda Y, Miyashita K (2012) Migratory patterns of exotic brown trout Salmo trutta in south-western Hokkaido, Japan, on the basis of otolith Sr: Ca ratios and acoustic telemetry. J Fish Biol 80:408–426
Hooton RM (1997) Status of coastal cutthroat trout in Oregon
Huitfeldt-Kaas H (1918) Ferskvandsfiskenes utbredelse og indvandring i Norge. Centraltrykkeriet, Kristiania
Hutchings JA (2004) Norms of reaction and phenotypic plasticity in salmonid life histories. In: Hendry AP, Stearns SC (eds) Evolution illuminated: salmon and their relatives. Oxford University Press, Oxford, pp 154–174
ICES (2021) Baltic Salmon and Trout Assessment Working Group (WGBAST). ICES Sci Rep 3(26):331
ICES (2012) Report of the Baltic Salmon and Trout Working Group (WGBAST). Uppsala Sweden
ICES (2013) ICES WK Trutta Report 2013
ICES (2020) Working Group with the Aim to Develop Assessment Models and Establish Biological Reference Points for Sea Trout (Anadromous Salmo trutta) Populations (WGTRUTTA; outputs from 2019 meeting)
Idyll C (1942) Food of rainbow, cutthroat and brown trout in the cowichan river system, B.C. J Fish Board Can 5:448–458
Januchowski-Hartley SR, Diebel M, Doran PJ, McIntyre PB (2014) Predicting road culvert passability for migratory fishes. Divers Distrib 20:1414–1424
Jauquet JM (2005) Diet of coastal cutthroat trout in south puget sound. In: The 2005 coastal cutthroat trout symposium, p 152
Jensen JLA, Rikardsen AH (2008) Do northern riverine anadromous Arctic charr Salvelinus alpinus and sea trout Salmo trutta overwinter in estuarine and marine waters? J Fish Biol 73:1810–1818
Jensen J, Rikardsen A (2012) Archival tags reveal that Arctic charr Salvelinus alpinus and brown trout Salmo trutta can use estuarine and marine waters during winter. J Fish Biol 81:735–749
Jensen AJ, Finstad B, Fiske P, Diserud OH, Thorstad EB (2020) Repeatable individual variation in migration timing in two anadromous salmonids and ecological consequences. Ecol Evol 10:11727–11738
Jensen AJ, Diserud OH, Finstad B, Fiske P, Thorstad EB (2022) Early-season brown trout (Salmo trutta) migrants grow and survive better at sea. J Fish Biol 100:1419–1431
Jepsen S, Aarestrup K, Økland F, Rasmussen G (1998) Survival of radio-tagged Atlantic salmon (Salmo salar L.) and trout (Salmo trutta L.) smolts passing a reservoir during seaward migration. Hydrobiologia 372:347–353
Johnson SC, Bravo S, Nagasawa K, Kabata Z, Hwang J, Ho J, Shih C (2004) A review of the impact of parasitic copepods on marine aquaculture. Zool Stud 43:229–243
Johnson JR, Baumsteiger J, Zydlewski J, Hudson JM, Ardren W (2010) Evidence of panmixia between sympatric life history forms of coastal cutthroat trout in two lower Columbia River tributaries. N Am J Fish Manag 30:691–701
Johnson T, Bjornn TC (1975) Evaluation of angling regulations in management of cutthroat trout. Idaho Fish & Game Department
Johnson OW, Ruckelshaus MH, Grant WS, Waknitz FW, Garrett AM, Bryant GJ, Neely KG, Hard JJ (1999) Status review of Coastal Cutthroat Trout from Washington, Oregon, and California
Johnston JM, Mercer SP (1976) Sea-run cutthroat in saltwater pens: broodstock development and extended juvenile rearing (with life history compendium). Washington State Game Department
Jones P, Closs G (2018) The introduction of brown trout to New Zealand and their impact on native fish communities. John Wiley and Sons, New Jersey
Jones DE (1975) Steelhead and sea-run cutthroat trout life history study in Southeast Alaska. Alaska Department of Fish Game
Jones DE (1977) Development of a technique for enhancement and management of anadromous cutthroat trout in southeast Alaska. Alaska Department of Fish Game, Annual Performance Report
Jonsson B, Jonsson N (1993) Partial migration: niche shift versus sexual maturation in fishes. Rev Fish Biol Fish 3:348–365
Jonsson B, Jonsson N (2006) Life-history effects of migratory costs in anadromous brown trout. J Fish Biol 69:860–869
Jonsson B, Jonsson N (2009a) Migratory timing, marine survival and growth of anadromous brown trout Salmo trutta in the River Imsa, Norway. J Fish Biol 74:621–638
Jonsson B, Jonsson N (2009b) A review of the likely effects of climate change on anadromous Atlantic salmon Salmo salar and brown trout Salmo trutta, with particular reference to water temperature and flow. J Fish Biol 75:2381–2447
Jonsson B, Jonsson N (2014) Naturally and hatchery produced European trout Salmo trutta: Do their marine survival and dispersal differ? J Coast Conserv 18:79–87
Jonsson B, Jonsson N (2015) Sexual size dimorphism in anadromous brown trout Salmo trutta. J Fish Biol 87:187–193
Jonsson B, L’Abee-Lund JH (1993) Latitudinal clines in life-history variables of anadromous brown trout in Europe. J Fish Biol 43:1–16
Jonsson B, Jonsson N, Jonsson M (2018) Water level influences migratory patterns of anadromous brown trout in small stream. Ecol Freshw Fish 27:1066–1075
Jonsson B, Jonsson N (2007) Life history of the anadromous trout Salmo trutta. Sea Trout: Biology, Conservation and Management, pp 196–223
Jonsson B, Jonsson N (2011) Ecology of Atlantic Salmon and Brown Trout: Habitat as a template for life histories. Springer
Jonsson B, Jonsson N, Albretsen J (2016) Environmental change influences the life history of salmon Salmo salar in the North Atlantic Ocean. J Fish Biol
Jonsson B, Jonsson N, Gresswell RE (2019) Life history diversity. Trout and Char of the World
Jutila E, Saura A, Kallio-Nyberg I, Huhmarniemi A (2006) Romakkaniemi, conservation, and management. The status and exploitation of sea trout on the Finnish coast of the Gulf of Bothnia in the Baltic Sea 128–138
Kallio-Nyberg I, Saura A, Ahlfors P (2002) Sea migration pattern of two sea trout (Salmo trutta) stocks released into the Gulf of Finland. in Annales Zoologici Fennici. JSTOR, pp 221–235
Källo K, Baktoft H, Kristensen ML, Birnie-Gauvin K, Aarestrup K (2022) High prevalence of straying in a wild brown trout (Salmo trutta) population in a fjord system. ICES J Mar Sci 79:1539–1547
Källo K, Birnie‐Gauvin K, Jepsen N, Aarestrup K (2023) Great cormorant (Phalacrocorax carbo sinensis) predation on adult anadromous brown trout (Salmo trutta). Ecol Freshw Fish
Keeley ER, Grant JWA (2001) Prey size of salmonid fishes in streams, lakes, and oceans. Can J Aquat Sci 58:1122–1132
Kendall NW, McMillan JR, Sloat M, R., T. W. Buehrens, T. P. Quinn, G. R. Pess, K. V. Kuzishchin, M. M. McClure, and R. W. Zabel. (2015) Anadromy and residency in steelhead and rainbow trout (Oncorhynchus mykiss): a review of the processes and patterns. Can J Fish Aquat Sci 72:319–342
Kennedy RJ, Barry J, Roche W, Rosell R, Allen M (2022) In-river behaviour and freshwater return rates of sea trout, Salmo trutta L., from two coastal river populations. J Fish Biol 101:1008–1020
King AR, Miller AL, Stevens JR (2021) Has stocking contributed to an increase in the rod catch of anadromous trout (Salmo trutta L.) in the Shetland Islands, UK? J Fish Biol 99:980–989
Klemetsen A, Amundsen PA, Dempson JB, Jonsson B, Jonsson N, O’Connell MF, Mortensen E (2003) Atlantic salmon Salmo salar L., brown trout Salmo trutta L. and Arctic charr Salvelinus alpinus (L.): a review of aspects of their life histories. Ecol Freshw Fish 12:1–59
Knutsen JA, Knutsen H, Gjøsæter J, Jonsson B (2001) Food of anadromous brown trout at sea. J Fish Biol 59:533–543
Kristensen ML, Birnie-Gauvin K, Aarestrup K (2019a) Behaviour of veteran sea trout Salmo trutta in a dangerous fjord system. Mar Ecol Prog Ser 616:141–153
Kristensen ML, Pedersen MW, Thygesen UH, del Villar-Guerra D, Baktoft H, Aarestrup K (2019b) Migration routes and habitat use of a highly adaptable salmonid (sea trout, Salmo trutta) in a complex marine area. Anim Biotelem 7:23
Kristensen ML, Righton D, D. d. Villar-Guerra, H. Baktoft, and K. Aarestrup. (2019c) Behaviour of adult sea trout Salmo trutta that survive or die at sea. Estuar Coast Shelf Sci 227:106310
Kuhn JA, Berg S, Støttrup JG (2022) Bycatch of brown trout Salmo trutta in a commercial gillnet fishery targeting whitefish Coregonus lavaretus. Fish Manag Ecol 29(2):143–155
Kuliskova P, Horky P, Slavik O, Jones JI (2009) Factors influencing movement behaviour and home range size in ide Leuciscus idus. J Fish Biol 74:1269–1279
L’abée-Lund JH (1989) Significance of mature male parr in a small population of Atlantic salmon (Salmo salar). Can J Fish Aquat Sci 46:928–931
L’Abée-Lund JH (1991) Variation within and between rivers in adult size and sea age at maturity of anadromous brown trout, Salmo trutta. Can J Fish Aquat Sci 48:1015–1021
Lance MM, Chang W-Y, Jeffries SJ, Pearson SF, Acevedo-Gutiérrez A (2012) Harbor seal diet in northern Puget Sound: implications for the recovery of depressed fish stocks. Mar Ecol Prog Ser 464:257–271
Landergren P (1999) Spawning of anadromous rainbow trout, Oncorhynchus mykiss (Walbaum): A threat to sea trout, Salmo trutta L., populations? Fish Res 40:55–63
Larranaga N, Wallerius ML, Guo H, Cucherousset J, Johnsson JI (2019) Invasive brook trout disrupt the diel activity and aggregation patterns of native brown trout. Can J Fish Aquat Sci 76:1052–1059
Latiu C, Cocan D, Uiuiu P, Ihut A, Nicula S-A, Constantinescu R, Miresan V, Biotechnologies. (2020) The Black Sea Trout, Salmo labrax Pallas, 1814 (Pisces: Salmonidae) in Romanian Waters. Bull UASVM Anim Sci 77:2
Lecomte F, Beall E, Chat J, Davaine P, Gaudin P (2013) The complete history of salmonid introductions in the Kerguelen Islands, Southern Ocean. Polar Biol 36:457–475
Lehtonen H (2002) Alien freshwater fishes of Europe. Invasive aquatic species of Europe. Distribution, impacts management Decision 153–161
Lemopoulos A, Uusi-Heikkilä S, Huusko A, Vasemägi A, Vainikka A (2018) Comparison of migratory and resident populations of brown trout reveals candidate genes for migration tendency. Genome Biol Evol 10:1493–1503
Leppäranta M, Myrberg K (2009) Physical oceanography of the Baltic Sea. Springer Science & Business Media
Li J, Phillips R, Harwood A, Koop B, Davidson W (2011) Identification of the sex chromosomes of brown trout (Salmo trutta) and their comparison with the corresponding chromosomes in Atlantic salmon (Salmo salar) and rainbow trout (Oncorhynchus mykiss). Cytogenet Genome Res 133:25–33
Lichatowich J (1999) Salmon without rivers. Island Press, Washington, D. C.
Lichatowich J, Powell M, Williams R (2006) Artificial production and the effects of fish culture on native Salmonids. Return to the river: restoring salmon to the Columbia River 417
Limburg K, Landergren P, Westin L, Elfman M, Kristiansson P (2001) Flexible modes of anadromy in Baltic sea trout: making the most of marginal spawning streams. J Fish Biol 59:682–695
Linnaeus CJE (1758) Systema Naturae per regna tria naturae. Secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis 1:823
Liu Y, Bailey JL, Davidsen JG (2019) Social-cultural ecosystem services of sea trout recreational fishing in Norway. Front Mar Sci 6
Lobón-Cerviá J, Sanz N (2017) Brown trout: biology, ecology and management
Lobón-Cerviá J, Esteve M, Berrebi, Duchi A, Lorenzoni M, Young KA (2019) Trout and char of central and southern Europe and northern Africa
Loch J, Miller DR (1988) Distribution and Diet of sea-run cutthroat trout captured in and adjacent to the Columbia River Plume, May-July 1980. Northwest Science 62
Losee JP, Phillips L, Young W (2016) Spawn timing and redd morphology of anadromous coastal Cutthroat Trout Oncorhynchus clarkii clarkii in a tributary of south Puget Sound Washington. N Am J Fish Manag 36:375–384
Losee JP, Seamons TR, Jauquet J (2017b) Migration patterns of anadromous cutthroat trout in south puget sound: a fisheries management perspective. Fish Res 187:218–225
Losee JP, Claiborne AM, Dionne PD, Faulkner HS, Seamons TR (2018) Size, age, growth and site fidelity of anadromous cutthroat trout Oncorhynchus clarkii clarkii in the Salish Sea. J Fish Biol 93:978–987
Losee JP, Dufault A, Kendall NW (2019) Changing salmon: an analysis of body mass, abundance and survival trends across 45 years in Puget Sound. Fish Fish 20:934–951
Losee JP, Jones SRM, McKinstry CAE, Batts WN, Hershberger PK (2020) Anadromous Coastal Cutthroat Trout (Oncorhynchus clarkii clarkii) as a Host for Argulus pugettensis (Crustacea, Branchiura): Parasite Prevalence, Intensity and Distribution. Northwest Sci 94:111–117
Losee JP, Madel G, Faulkner H, Claiborne A, Seamons TR, Young W( 2017a) Cutthroat trout in saltwater: spawn timing, migration patterns and abundance of anadromous coastal cutthroat trout
Losee JP, Dapp DR, Madel G, Shimek G (2022) Volunteer angling and technology‐based solutions provide the first estimate of sea lice infections for wild searun cutthroat trout (Oncorhynchus clarkii clarkii). Fisheries
Lothrop R, Losee J (2016) Value of catch-and-release fishing for puget sound coastal cutthroat trout. Washington Department of Fish and Wildlife
Lutz E, Hirsch PE, Bussmann K, Wiegleb J, Jermann HP, Muller R, Burkhardt-Holm P, Adrian-Kalchhauser I (2020) Predation on native fish eggs by invasive round goby revealed by species-specific gut content DNA analyses. Aquat Conserv Mar Freshw Ecosyst 30:1566–1577
Lyse A, Stefansson S, Fernö A (1998) Behaviour and diet of sea trout post-smolts in a Norwegian fjord system. J Fish Biol 52:923–936
MacCrimmon HR, Marshall TL (1968) World distribution of brown trout. Salmo Trutta 25:2527–2548
MacCrimmon HR, Marshall TL, Gots BL (1970) World distribution of brown trout, Salmo trutta: further observations. J Fish Board Can 27:811–818
Makhrov AA (2008) Hybridization of the Atlantic salmon (Salmo salar L.) and brown trout (S. trutta L.). In: Zoosystematica Rossica, pp 129–143
Matthews MA, Poole WR, Thompson CE, McKillen J, Ferguson A, Hindar K, Wheelan KF (2000) Incidence of hybridization between Atlantic salmon, Salmo salar L., and brown trout, Salmo trutta L., in Ireland. Fish Manag Ecol 7:337–347
McKay SJ, Devlin RH, S. M. J. (1996) Phylogeny of Pacific salmon and trout based on growth hormone type-2 and mitochondrial NADH dehydrogenase subunit 3 DNA sequences. Can J Fish Aquat Sci 53:1165–1176
McMillan JR, Pess GR, McHenry ML, Moses R, Quinn TP (2014) Documentation of unusual, fall spawning by coastal cutthroat trout in the Elwha River System, Washington. Trans Am Fish Soc 143:1605–1611
McPhail J, Lindsey C (1986) Zoogeography of the freshwater fishes of Cascadia (the Columbia system and rivers north to the Stikine). Zoogeogr N Am Freshw Fishes 615:638
Mercer SP, Johnston JM (1980) Sea-run cutthroat: development and evaluation of a new enhancement technique. Washington State Game Department
Mercer SP (1982) Sea-run cutthroat: Broodstock development and evaluation of enhancement techniques. F-68-R
Metcalf JL, Love Stowell S, Kennedy CM, Rogers KB, McDonald D, Epp J, Keepers K, Cooper A, Austin JJ, Martin AP (2012) Historical stocking data and 19th century DNA reveal human-induced changes to native diversity and distribution of cutthroat trout. Mol Ecol 21:5194–5207
Metcalf JL, Siegle MR,Martin Ap (2008) Hybridization dynamics between Colorado's native cutthroat trout and introduced rainbow trout 99:149–156
Michael J (1989) Life history of anadromous coastal cutthroat trout in Snow and Salmon Creeks, Jefferson County, Washington, with implications for management. Washington Department of Fish and Wildlife 75
Minett JF, Garcia de Leaniz C, Sobolewska H, Brickle P, Crossin GT, Consuegra S (2021) SNP analyses and acoustic tagging reveal multiple origins and widespread dispersal of invasive brown trout in the Falkland Islands. Evol Appl 14:2134–2144
Moore KM, Gregory SV (1988) Response of young-of-the-year cutthroat trout to manipulation of habitat structure in a small stream. Trans Am Fish Soc 117:162–170
Moore A, Ives M, Scott M, Bamber S (1998) The migratory behaviour of wild sea trout (Salmo trutta L.) smolts in the estuary of the River Conwy. North Wales Aquaculture 168:57–68
Moore ME, Goetz FA, D. M. Van doornik, E. P. Tezak, T. P. Quinn, J. J. Reyes-Tomassini, and B. S. Berejikian. (2010) Early marine migration patterns of wild coastal cutthroat trout (Oncorhynchus clarkii clarkii), steelhead trout (Oncorhynchus mykiss), and their hybrids. PLoS ONE 5:e12881
Moore JW, Yeakel JD, Peard D, Lough J, Beere M (2014) Life-history diversity and its importance to population stability and persistence of a migratory fish: steelhead in two large North American watersheds. J Anim Ecol 83:1035–1046
Morinville GR, Rasmussen JB (2006) Marine feeding patterns of anadromous brook trout (Salvelinus fontinalis) inhabiting an estuarine river fjord. Can J Fish Aquat Sci 63:2011–2027
Mottola G, Kristensen T, Anttila K (2020) Compromised thermal tolerance of cardiovascular capacity in upstream migrating Arctic char and brown trout—Are hot summers threatening migrating salmonids? Conserv Physiol 8:coaa101
Muhlfeld CC et al (2019) Global status of trout and char: conservation challenges in the twenty-first century. Trout char of the world, pp 717–760
Mulder IM, Morris CJ, Dempson JB, Fleming IA, Power M (2020) Marine temperature and depth use by anadromous Arctic char correlates to body size and diel period. Can J Fish Aquat Sci 77:882–893
Müller K, Karlsson L (1983) The biology of the grayling, Thymallus thymallus L. in coastal areas of the Bothnian sea. Aquilo Ser Zool 68
Naish KA et al (2007) An evaluation of the effects of conservation and fishery enhancement hatcheries on wild populations of salmon. Adv Mar Biol 53:61–194
Nevoux M, Finstad B, Davidsen JG, Finlay R, Josset Q, Poole R, Höjesjö J, Aarestrup K, Persson L, Tolvanen O (2019) Environmental influences on life history strategies in partially anadromous brown trout (Salmo trutta, Salmonidae). Fish Fish 20:1051–1082
Nika N, Virbickas T, Kontautas A (2011) Spawning site selection and redd gravel characteristics of sea trout Salmo trutta in the lowland streams of Lithuania. Oceanol Hydrobiol Stud 40:46–56
O’Reilly P, Mawle G, and Management (2006) An appreciation of the social and economic values of sea trout in England and Wales. Sea Trout Bio Conser Manag 457–465
O’Reilly P (2015) The economic contribution of salmon and sea trout angling in Ireland. Report to the National Strategy for Angling Development
O’Neal SL (2008) Lessons to learn from all out invasion: life history of brown trout (Salmo trutta) in a Patagonian River. University of Montana, Polson, MT
O’Neal SL, Stanford JA (2011) Partial migration in a robust brown trout population of a Patagonian river. Trans Am Fish Soc 140:623–635
Ostberg CO, Slatton SL, Rodriguez R (2004) Spatial partitioning and asymmetric hybridization among sympatric coastal steelhead trout (Oncorhynchus mykiss irideus), coastal cutthroat trout (O. clarki clarki) and interspecific hybrids. Mol Ecol 13:2773–2788
Östergren J, Nilsson J, Lundqvist H, Dannewitz J, Palm S (2016) Genetic baseline for conservation and management of sea trout in the northern Baltic Sea. Conserv Genet 17:177–191
Ottaway EM, Carling PA, Clarke A, Reader NA (1981) Observations on the structure of brown trout, Salmo trutta Linnaeus, redds. J Fish Biol 19:593–607
Page LM (2008) Handbook of European freshwater fishes. Copeia 725–727:723
Page LM, Burr BM (1991) A field guide to freshwater fishes: North America north of Mexico. Houghton Mifflin Harcourt
Palm S, Dannewit J, Prestegaard T, Östergren JJ (2013) Aqua Report. 2013. Laxing och felvandrad lax i Mörrumsån—en genetisk analys (Salmon-trout hybrids and strayers in Mörrumsån River—a genetic analysis, In Swedish with an English abstract). 20
Pauley GB, Thomas GL (1993) Mortality of anadromous coastal cutthroat trout caught with artificial lures and natural bait. North Am J Fish Manag 13:337–345
Pauley GB, et al (1989) Species Profiles: life histories and enviromental requirements of coastal fishes and invertebrates (Pacific Northwest). Biological Report 82 (11, 86), U.S. Dept. of the Interior, and the U. S. Army Corps of Engineers, Lafayette, LA
Pauly D (1995) Anecdotes and shifting baseline syndrome of fisheries. Trends Ecol Evol 10:430
Pearcy WG (1992) Ocean ecology of North Pacific salmonids. University of Washington Press, Seattle, WA
Pearcy W, Brodeur R, Fisher J (1990) Distribution and biology of juvenile cutthroat trout Oncorhynchus clarki clarki and steelhead O. mykiss in coastal waters off Oregon and Washington. Fish Bull 88:697–711
Pearcy WG, Brodeur RD, McKinnell SM, Losee JP (2018) Ocean ecology of anadromous coastal cutthroat trout. In: Beamish RJ (ed) Ocean Ecology of Pacific Salmon and Trout. American Fisheries Society, Bethesda, MD, pp 905–930
Pedersen S, Degerman E, Debowski P, Petereit C (2017) Assessment and recruitment status of Baltic Sea trout populations. In: Sea trout: science and management: proceedings of the second international sea trout symposium. Troubador Publishing, Leicester, UK, pp 423–441
Pemberton R (1976) Sea trout in North Argyll sea lochs, population, distribution and movements. J Fish Biol 9:157–179
Penaluna BE, Dunham JB, Andersen HV (2021) Nowhere to hide: The importance of instream cover for stream-living Coastal Cutthroat Trout during seasonal low flow. Ecol Freshw Fish 30:256–269
Peoples M, Cenci M, Hunter J (1988) Sea-run cutthroat studies in south Puget Sound tributaries, July 1986 to June 1987. Washington State Department of Wildlife, Fisheries Management Division
Pielou EC (2008) After the ice age. University of Chicago Press, After the Ice Age
Price DM, Quinn T, Barnard RJ (2010) Fish passage effectiveness of recently constructed road crossing culverts in the Puget Sound region of Washington State. N Am J Fish Manag 30:1110–1125
Quinn TP (2018) The behavior and ecology of Pacific salmon and trout. University of Washington Press, Seattle, WA
Quinn TP, Myers KW (2004) Anadromy and the marine migrations of Pacific salmon and trout: Rounsefell revisited. Rev Fish Biol Fisheries 14:421–442
Quinn TP, McGinnity P, Reed TE (2016) The paradox of “premature migration” by adult anadromous salmonid fishes: patterns and hypotheses. Can J Fish Aquat Sci 73:1015–1030
Rahel FJ, Olden JD (2008) Assessing the effects of climate change on aquatic invasive species. Conserv Biol 22:521–533
Reed TE, de Eyto E, O’Higgins K, Gargan P, Roche W, White J, O’Maoileidigh N, Quinn TP, McGinnity P (2017) Availability of holding habitat in lakes and rivers affects the incidence of spring (premature) upriver migration by Atlantic salmon. Can J Fish Aquat Sci 74:668–679
Rempel M (1984) Wild and hatchery steelhead and cutthroat trout populations of the Little Campbell River, BC, 1983. Ministry of Environment, Fisheries Branch
Ricker WE (1981) Changes in the average size and average age of Pacific salmon. Can J Fish Aquat Sci 38:1636–1656
Rikardsen A, Amundsen P, Knudsen R, Sandring S (2006) Seasonal marine feeding and body condition of sea trout (Salmo trutta) at its northern distribution. J Mar Sci 63:466–475
Rohtla M, Matetski L, Svirgsden R, Kesler M, Taal I, Saura A, Vaittinen M, Vetemaa M (2017) Do sea trout Salmo trutta parr surveys monitor the densities of anadromous or resident maternal origin parr, or both? Fish Manag Ecol 24:156–162
Rohtla M, Matetski L, Taal I, Svirgsden R, Kesler M, Paiste P, Vetemaa M (2020) Quantifying an overlooked aspect of partial migration using otolith microchemistry. J Fish Biol 97:1582–1585
Rubidge EM, Taylor EB (2004) Hybrid zone structure and the potential role of selection in hybridizing populations of native westslope cutthroat trout (Oncorhynchus clarki lewisi) and introduced rainbow trout (O. mykiss). Mol Ecol 13:3735–3749
Saiget D, Sloat M, Reeves G (2007) Spawning and movement behavior of migratory coastal cutthroat trout on the western Copper River Delta, Alaska. N Am J Fish Manag 27:1029–1040
Sánchez-Hernández J (2020) Drivers of piscivory in a globally distributed aquatic predator (brown trout): a meta-analysis. Sci Rep 10:11258
Schindler DE, Armstrong JB, Reed TE (2015) The portfolio concept in ecology and evolution. Front Ecol Environ 13:257–263
Schöffmann J (2021) Trout and Salmon of the Genus Salmo. American Fisheries Society
Sepkoski JJ, Jr. (1992) A compendium of fossil marine animal families, 2nd edition. Contrib Biol Geol 83:1–156
Shephard S, List CJ, Arlinghaus R (2023) Reviving the unique potential of recreational fishers as environmental stewards of aquatic ecosystems. Fish Fish 24:339–351
Shry SJ, McCallum ES, Alanärä A, Persson L, Hellström G (2019) Energetic status modulates facultative migration in brown trout (Salmo trutta) differentially by age and spatial scale. Front Ecol Evol 7
Skaala Ø, Johnsen GH, Lo H, Borgstrøm R, Wennevik V, Hansen MM, Merz JE, Glover KA, Barlaup BT (2014) A conservation plan for Atlantic salmon (Salmo salar) and anadromous brown trout (Salmo trutta) in a region with intensive industrial use of aquatic habitats, the Hardangerfjord, western Norway. Mar Biol Res 10:308–322
Skov C, Gundelund C, Weltersbach MS, Ferter K, Bertelsen SK, Jepsen N (2022) Catch and release angling for sea trout explored by citizen science: Angler behavior, hooking location and bleeding patterns. Fish Res 255:106451
Slaney TL, Hyatt KD, Northcote TG, Fielden RJ (1996) Status of Anadromous Salmon and Trout in British Columbia and Yukon. Fisheries 21:20–35
Slavík O, Horký P (2019) Home range size decreases with increasing site fidelity in high-density subpopulations of brown trout. Ethol Ecol Evol 31:421–434
Sloat M, R. (2013) Born to run? Integrating individual behavior, physiology, and life histories in poartially migratory steelhead and rainbow trout (Oncorhynchus mykiss). Oregon State University, Corvallis, OR
Sloat MR, Fraser DJ, Dunham JB, Falke JA, Jordan CE, McMillan JR, Ohms HA (2014) Ecological and evolutionary patterns of freshwater maturation in Pacific and Atlantic salmonines. Rev Fish Biol Fish 24:689–707
SLU (2022) KUL-database. Institue of Coastal Research, Dpeartment of Aquatic Resources
Smith GR, Stearley RF (2018) The fossil record of Cutthroat Trout: implications for evolution and conservation. In: Trotter P, Bisson P, Schultz L, Roper B (eds) Cutthroat trout: evolutionary biology and taxonomy. American Fisheries Society, Symposium 36, pp 77–101
Snoj A, Bravničar J, Marić S, Sušnik Bajec S, Benaissa H, Schöffmann J (2021) Nuclear DNA reveals multiple waves of colonisation, reticulate evolution and a large impact of stocking on trout in north-west Africa. Hydrobiologia 848:3389–3405
Solomon D, Child A (1978) Identification of juvenile natural hybrids between Atlantic salmon (Salmo solar L.) and trout (Salmo trutta L.). J Fish Biol 12:499–501
Solomon D, Czerwinski M, and management (2007) Catch and release, net fishing and sea trout fisheries management
Solomon DJ (1997) Review of sea trout fecundity. Environment Agency Bristol, UK
Staveley TA, Ahlbeck Bergendahl I (2022) Pink salmon distribution in Sweden: The calm before the storm? Ecol Evol 12:e9194
Sturlaugsson J (2017) The Marine Migration & Swimming Depth of Sea Trout (Salmo trutta L.) in Icelandic Waters. In: Proceedings of 2nd international sea trout symposium sea trout: science and management, Dundalk, Ireland, Harris, G., Ed., Leicester: Troubador
Sumner FH (1962) Migration and growth of the coastal cutthroat trout in Tillamook County, Oregon. Trans Am Fish Soc 91:77–83
Svendsen JC, Koed A, Aarestrup K (2004) Factors influencing the spawning migration of female anadromous brown trout. J Fish Biol 64:528–540
Taal I, Rohtla M, Saks L, Kesler M, Jürgens K, Svirgsden R, Matetski L, Verliin A, Paiste P, Vetemaa M (2018) Parr dispersal between streams via a marine environment: a novel mechanism behind straying for anadromous brown trout? Ecol Freshw Fish 27:209–215
Tåking ÅV (1950) Influence of the environment on number of vertebrae in teleostean fishes. Nature 165:28–28
Tamate T, Maekawa K (2006) Latitudinal variation in sexual size dimorphism of sea-run Masu salmon, Oncorhynchus masou. Evolution 60:196–201
Thorstad EB (2016) Marine life of the sea trout. Mar Biol 163:47
Thorstad EB, Todd CD, Uglem I, Bjørn PA, Gargan PG, Vollset KW, Halttunen E, Kålås S, Berg M, Finstad B (2015) Effects of salmon lice Lepeophtheirus salmonis on wild sea trout Salmo trutta a literature review. Aquac Environ Interact 7:91–113
Tipping J (1986) Effect of release size on return rates of hatchery sea-run cutthroat trout. Progress Fish-Culturist 48:195–197
Tipping JM (2001) Communications: adult returns of hatchery sea-run cutthroat reared in a seminatural pond for differing periods prior to release. N Am J Aquac 63:131–133
Tipping JM, Blankenship H (1993) Effect of condition factor at release on smolt-to-adult survival of hatchery sea-run cutthroat trout. Progress Fish-Culturist 55:184–186
Tougard C (2022) Will the genomics revolution finally solve the Salmo systematics? Hydrobiologia 1–16
Trotter PC (1989) Coastal cutthroat trout: a life history compendium. Trans Am Fish Soc 118:463–473
Trotter PC, Bisson PA, Fransen BR (1993) Status and plight of searun cutthroat trout. Plenum Press
Trotter P, Bisson P, Shultz L, Roper B (2018) Cutthroat trout: evolutionary biology and raxonomy. American Fisheries Society
Trotter P (2008) Cutthroat: native trout of the west. University of California Press
Vatandoust S, Abdoli A, Anvarifar H, Mousavi-Sabet H (2014) Morphometric and meristic characteristics and morphological fario (Pisces: Salmonidae) along the southern Caspian Sea basin. Eur J Zool Res 3:56–65
Vélez-Espino LA, McLaughlin RL, Robillard M (2013) Ecological advantages of partial migration as a conditional strategy. Theor Popul Biol 85:1–11
Verspoor E, Hammart J (1991) Introgressive hybridization in fishes: the biochemical evidence. J Fish Biol 39:309–334
Viktorsson L (2018) Hydrography and oxygen in the deep basins. Helsinki: HELCOM Baltic Sea environment fact sheets
Waldman JR, Quinn TP (2022) North American diadromous fishes: Drivers of decline and potential for recovery in the Anthropocene. Sci Adv 8:eabl5486
Walker AM, Pawson MG, Potter ECE (2004) Sea trout fisheries management: should we follow the salmon Sea Trout. Biol Conserv Manag 466–479
Westley PA, Fleming IA (2011) Landscape factors that shape a slow and persistent aquatic invasion: brown trout in Newfoundland 1883–2010. Diversity 17:566–579
Whiteley AR, Penaluna BE, Taylor EB, Weiss E, Abadía-Cardoso A, Gomez-Uchida D, Koizumi I, Trotter P (2019) Trout and char: taxonomy, systematics, and phylogeography. In: K JL, W JE, G RE, L.-C. J (eds) Trout and Char of the World. American Fisheries Society
Williams I, Reeves GH, Graziano SL, Nielsen JL (2007) Genetic investigation of natural hybridization between rainbow and coastal cutthroat trout in the Copper River Delta, Alaska. Trans Am Fish Soc 136:926–942
Williams J, Isaak D, Imhof J, Hendrickson D, McMillan J (2015) Cold-water fishes and climate change in North America. Ref Module Earth Syst Environ Sci. https://doi.org/10.1016/B978-0-12-409548-9.09505-1
Young KA (2005) Life-history variation and allometry for sexual size dimorphism in Pacific salmon and trout. Proc R Soc B Biol Sci 272:167–172
Zimmerman MS, Irvine JR, O’Neill M, Anderson JH, Greene CM, Weinheimer J, Trudel M, Rawson K (2015) Spatial and temporal patterns in smolt survival of wild and hatchery coho salmon (Oncorhynchus kisutch) in the Salish Sea. Mar Coast Fish Dyn Manag Ecosys Sci 7:116–134
Zydlewski J, Johnson J, Brunzell J, Hogle J, Clements S, Karnowski M, Schreck C (2008) Seaward migration of coastal cutthroat trout Oncorhynchus clarkii clarkii from four tributaries of the Columbia River. Status, Management, Biology, and Conservation, Oregon Chapter, American Fisheries Society, Coastal Cutthroat Trout Symposium
Zydlewski J, Zydlewski G, Kennedy B, Gale W (2014) Smolting in coastal cutthroat trout Onchorhynchus clarkii clarkii. J Fish Biol 85:1111–1130
Acknowledgements
We appreciate the help of WDFW Librarian Connie Sterner, illustrations by Joseph Tomelleri, GIS expert Dale Gombert and data providers, Russell Poole (Marine Institute, Newport, Co. Mayo, Ireland), and Jan Davidson (NTNU Museum), Bryce O’Connor (Chu Cho Environmental), Bregan Brown (Natural Resources Wales) and thoughtful conversations with Greg Shimek (Coastal Cutthroat Coalition) and Rob Allan (WDFW). We also appreciate the comments on an earlier version of this document by Charlene Hurst (WDFW) and three anonymous reviewers.
Funding
Open access funding provided by Swedish University of Agricultural Sciences. Funding was provided by Washington Department of Fish and Wildlife and Swedish University of Agricultural Sciences.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The author has no relevant financial or non-financial interests to disclose.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Losee, J.P., Palm, D., Claiborne, A. et al. Anadromous trout from opposite sides of the globe: biology, ocean ecology, and management of anadromous brown and cutthroat trout. Rev Fish Biol Fisheries 34, 461–490 (2024). https://doi.org/10.1007/s11160-023-09824-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11160-023-09824-0