
Abstract
Chemical analysis of calcified structures continues to flourish, as analytical and technological advances enable researchers to tap into trace elements and isotopes taken up in otoliths and other archival tissues at ever greater resolution. Increasingly, these tracers are applied to refine age estimation and interpretation, and to chronicle responses to environmental stressors, linking these to ecological, physiological, and life-history processes. Here, we review emerging approaches and innovative research directions in otolith chemistry, as well as in the chemistry of other archival tissues, outlining their value for fisheries and ecosystem-based management, turning the spotlight on areas where such biomarkers can support decision making. We summarise recent milestones and the challenges that lie ahead to using otoliths and archival tissues as biomarkers, grouped into seven, rapidly expanding and application-oriented research areas that apply chemical analysis in a variety of contexts, namely: (1) supporting fish age estimation; (2) evaluating environmental stress, ecophysiology and individual performance; (3) confirming seafood provenance; (4) resolving connectivity and movement pathways; (5) characterising food webs and trophic interactions; (6) reconstructing reproductive life histories; and (7) tracing stock enhancement efforts. Emerging research directions that apply hard part chemistry to combat seafood fraud, quantify past food webs, as well as to reconcile growth, movement, thermal, metabolic, stress and reproductive life-histories provide opportunities to examine how harvesting and global change impact fish health and fisheries productivity. Ultimately, improved appreciation of the many practical benefits of archival tissue chemistry to fisheries and ecosystem-based management will support their increased implementation into routine monitoring.
Graphical abstract

Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Information extracted from otoliths and other calcified structures is increasingly used to inform sustainable resource management in a rapidly changing world; and such information is often unattainable from sensors, data storage tags or other methods. This is particularly true for early life stages when fish are too small to tag, and otoliths and other archival tissues are used instead as the inert growth layers chronicle each individual’s entire life history. As a result, research using otoliths and other archival tissues (e.g., scales, vertebrae, fin rays, eye lenses, statoliths) is growing rapidly. Increasingly, more established techniques such as age, growth and shape analyses are paired with novel chemical approaches to provide new insights into fish life histories, health, population dynamics and demography (Avigliano 2021; Cadrin et al. 2014; Campana 2005; Nazir & Khan 2021; Tanner et al. 2016; Walther 2019).
Otoliths have been at the centre of fisheries science for many decades, but their use has evolved rapidly in recent years, triggered by landmark discoveries, and analytical and technological advances (Campana 2005; Walther 2019). Chemical analyses traditionally applied to otoliths are now being used on analogous archival tissues such as eye lenses, scales, vertebrae and operculae (Izzo et al. 2016a; Tzadik et al. 2017). While age and growth analyses dominated early otolith research, many recent studies have shown that shape and chemical analyses are powerful approaches to define stocks, reconstruct movements, and unravel individual life histories (Avigliano 2021; Gillanders et al. 2015; Smoliński et al. 2020; Tanner et al. 2016; Tzadik et al. 2017). Importantly, continued advances in analytical techniques have enabled researchers to measure a growing number of elemental and isotopic tracers at increasing fine temporal resolution, allowing them to link adverse environmental conditions to ecological, physiological and life-history processes (Chung et al. 2019b; Heimbrand et al. 2020; Johnson et al. 2020; Reis-Santos et al. 2021; Sturrock et al. 2020). This is a rapidly developing field, and the expansion in research was evident in the “Biomarkers in Calcified Structures” session at the 2021 World Fisheries Congress [Theme 4: Future of Fish and Fisheries (Innovations in Fisheries)], highlighting the many research directions that can be rapidly translated into fisheries management applications.
As an outcome of this session, we synthesize emerging approaches and major developments in the chemical analysis of otoliths and other archival tissues, outlining their value and pertinence to fisheries and ecosystem-based management, turning the spotlight on areas where such biomarkers can inform fisheries management and support decision making. The current paper does not focus on stock structure [as this has been addressed recently, e.g., Avigliano (2021); Cadrin et al. (2014); Nazir and Khan (2021); Tanner et al. (2016)] but rather reviews recent groundwork on other emerging and innovative research directions, noting areas that are particularly promising for rapid expansion and implementation. Overall, we summarise recent milestones and explore the challenges that lie ahead to using biogeochemical markers in otoliths and archival tissues, grouped into seven application-oriented and interdisciplinary research areas: (1) supporting fish age estimation; (2) evaluating environmental stress, ecophysiology and individual performance; (3) confirming seafood provenance; (4) resolving connectivity and individual movement pathways; (5) characterising food webs and trophic interactions; (6) reconstructing reproductive life histories; and (7) tracing stock enhancement efforts. We focus mainly on otolith research but also draw attention to state-of-the-art research into other archival tissues (e.g., scales, operculae, vertebrae, eye lenses) to highlight synergies and lessons learnt, focusing on the practical benefits of using biomarkers in such structures and next steps to support their application in routine fisheries monitoring.
Chemistry to support age estimation
Age estimation of fish, based on counts of growth increments in otoliths and other archival tissues, underpins calculations of population growth, mortality, longevity. These metrics are fundamental to understanding population dynamics for fisheries assessments and making accurate assignment of ages, which is critical for reliable stock assessments (Hüssy et al. 2016a). However, some species groups (e.g., Clupeiformes and Gadiformes) possess otoliths with diffuse banding patterns, impeding increment counts. This in turn increases reader subjectivity, leading to poor accuracy and precision for age estimates. Because the periodic formation of growth increments is not ubiquitous among fish, there is a need to validate increment formation, with chemical approaches increasingly explored to overcome issues in growth increment interpretation.
Classic approaches: validation of absolute age
Radiochemical and bomb radiocarbon dating provide indirect chemical methods to validate estimates of the absolute age and growth of fish (Campana 2001). The use of radiochemical methods is based on the uptake of environmental elements and isotopes to otoliths and is particularly useful in long lived species. Radiochemical dating is a geochemical approach that uses naturally occurring radioactivity (e.g., 228Th/228Ra and 210Pb/226Ra) as a clock within the inert crystalline otolith structure (Andrews et al. 2009, 2012; Andrews 2016; Campana et al. 1993). In contrast, bomb radiocarbon dating uses an anthropogenic radiocarbon signal created by atmospheric testing of thermonuclear devices in the 1950–60s, and their decline over time, as a reference in time relative to 14C concentration levels absorbed by otoliths, or other calcified structures (Andrews et al. 2012; Kalish 1993; Lackmann et al. 2019; Tracey et al. 2017), and can provide absolute ages of individuals depending on their birth year.
Emerging approaches: verification of visual patterns and age estimation
Seasonal patterns in element concentration
Although otoliths are routinely used for age estimation, the formation of ambiguous growth zones can prevent visual interpretation. Accordingly, interest in applying chemical methods for assessing seasonality has increased. One approach to reconstruct seasonal contrasts is temperature-dependent oxygen isotope fractionation (Høie et al. 2004a; Kalish 1991c) in temperate and arctic latitudes (Høie et al. 2004b). To date, oxygen isotope analysis has been used to estimate the timing of growth zone formation in Atlantic cod (Gadus morhua) (Høie & Folkvord 2006) and for validating age estimates in Pacific cod (G. macrocepahlus) (Kastelle et al. 2017). Recently, variations in chemical concentrations have been explored as tracers of physiological processes to underpin age estimation. Elements P, Mg and Zn are associated with the organic matrix of the otolith (Izzo et al. 2016b; Miller et al. 2006; Thomas et al. 2017) and reflect seasonal variations in growth (Heimbrand et al. 2020; Hüssy et al. 2021a; Limburg et al. 2018) (Fig. 1). Minima often correlate with visually identified translucent winter growth zones (Friedrich & Halden 2010; Halden et al. 2000; Halden & Friedrich 2008; Hüssy et al. 2016b; Limburg & Elfman 2010) or occur during the coldest months of the year (Hüssy et al. 2021a). Seasonal differences have also been described in Sr (Brophy et al. 2021; Siskey et al. 2016; Tzeng et al. 1999), though this element is often considered a proxy for environmental salinity.

Transverse sections of otoliths of Baltic cod Gadhus morhua, and superimposed variations in the chemical composition of P:Ca (red) and Mg:Ca (yellow) from the otolith core to the dorsal edge (measured with Laser ablation ICPMS) demonstrate the usefulness of chemical profiles to support age estimation. A is a transverse section of an otolith from an 8-year-old cod from the Baltic Sea, with good readability, associated with clear increments, and high contrast between translucent and opaque zones, that match variations in P:Ca and Mg:Ca. B is a transverse section of a 5-year-old cod from the Baltic Sea, lacking clear visual increments, and where oscillations between maxima and minima of the chemical profiles support age estimation where visual increments were unreliable. Dashed lines represent chemical age estimation points and grey lines the laser ablation path. Transverse sections of otoliths were viewed under reflected light (photos: Y. Heimbrand)
Biological challenges
The key to the successful application of chemical approaches to support age estimation is a detailed understanding of developmental processes and how elements pass various membranes and organs from water and food into the otolith matrix. Tracers of physiology and growth are essential co-factors in many metalloenzymes and are needed for a range of physiological processes, including otolith biomineralization. These interacting mechanisms are complex and hard to tease apart. Even in the well-studied otolith, more knowledge is needed on element uptake dynamics, transport, regulation, and incorporation into the growing otolith crystal lattice to distinguish controlling endogenous and exogenous factors (Hüssy et al. 2021b; Thomas et al. 2017; Thomas & Swearer 2019). In the plasma, elements may occur as free ions (Campana 1999; Payan et al. 2002), protein-bound (P. E. Fletcher & Fletcher 1980; Sturrock et al. 2014) or both. Transport of free ions from plasma into the endolymph is not fully understood and is presumed to occur primarily via passive diffusion along a concentration gradient, whereas elements bound to plasma proteins presumably require active transport across the endolymphatic epithelium. Physiological regulation of element uptake and incorporation is thought to be exerted by a wide range of factors, including ontogeny (Arai et al. 2007; Macdonald et al. 2020; Ranaldi & Gagnon 2010; Reis-Santos et al. 2018b), maturation and spawning (Sturrock et al. 2014, 2015a), diet (Ranaldi & Gagnon 2008), and growth (Hüssy et al. 2021b; Sturrock et al. 2015a) in addition to temperature (Reis-Santos et al. 2013a) which influences metabolic rate, but with considerable uncertainty regarding the magnitude—and even the direction—of the impact on otolith growth and chemistry. It also remains to be seen to what extent uptake mechanisms are universal or subject to phylogenetic differences in incorporation mechanisms (Limburg & Elfman 2010).
Methodological challenges
The ability to detect age-related patterns in chemical composition is determined by the spatial resolution of the analytical method relative to the size and annual growth rate of the archival tissue. The achievable temporal resolution declines towards the edge of the structure as age increases and growth slows (Høie et al. 2004b; Kastelle et al. 2017). Generally, probe-based techniques [e.g., laser ablation—inductively coupled plasma mass spectrometry (LA-ICPMS)], enable more detailed reconstruction of life histories than methods requiring sample dissolution or combustion [e.g., solution based ICPMS, isotope ratio mass spectrometry (IRMS)]. High-resolution micro-milling followed by IRMS has sufficient resolution to detect temperature related cycles in δ18O values in cod up to 6 years of age (Høie et al. 2004b; Kastelle et al. 2017), but long-lived species require higher resolution probe-based secondary ion mass spectrometry (SIMS) (Matta et al. 2010). With LA-ICPMS, continuous line scans can capture oscillations in trace elements at high spatial and temporal resolution allowing for the reconstruction of entire life history profiles (Heimbrand et al. 2020; Hüssy et al. 2016b, 2021a). Alternatively, high-resolution 2D elemental maps, produced using LA-ICPMS (Artetxe-Arrate et al. 2021; Heimbrand et al. 2020; McGowan et al. 2014), particle induced x-ray emission (PIXE) (Halden et al. 2000; Walther & Limburg 2012) or scanning X-ray fluorescence microscopy (Hermann et al. 2016; Tzeng et al. 1999) can reveal spatial heterogeneity in elemental composition, which is useful for observing banding patterns and identifying the optimal transect for analysis of seasonal trends (Heimbrand et al. 2020).
Investigations of age-related patterns in otolith chemistry employ a variety of analytical techniques, including visual inspection of chemical patterns in relation to growth increments (Seyama et al. 1991; Tzeng et al. 1999), statistical comparison of concentrations between opaque and translucent bands (Tomás et al. 2006) and the use of time-series methods to identify peaks or cycles in concentrations (Hüssy et al. 2016b, 2021a; Kastelle et al. 2017; Siskey et al. 2016; Sturrock et al. 2015a). Even with advanced quantitative techniques, the identification of seasonal patterns in chemical data can involve a degree of subjectivity. Standard peak detection algorithms require the user to define the threshold amplitude of a peak and the number of data points (window) over which to search for changes above that threshold (Jarman et al. 2003; Kay 1998). Settings can be optimised with reference to visual bands (Hüssy et al. 2021a) but this requires samples with clear unambiguous increments and somewhat reduces the independence of the chemistry-based age estimates. Alternatively, spectral analysis methods can detect periodic patterns along a chemical profile and statistically test for annual periodicity; the Lomb-Scargle periodogram is particularly appropriate as it can deal with data that are irregularly spaced (Lomb 1976; Scargle 1982). A potential limitation is the requirement for a temporal reference point, which can be supplied by the position of annual growth marks (Siskey et al. 2016). Nonetheless, even when peaks in elemental concentrations were identified using only visual inspection, inter-reader agreement was higher than with interpretation of visual growth marks for a difficult to age species (Eastern Baltic Sea cod, G. morhua) (Heimbrand et al. 2020), indicating that chemical tools can improve the objectivity of age estimation and be a key part in fisheries managers’ tool chest.
Two-dimensional mapping of stable isotopes in otoliths is also possible using nuclear reaction analysis (Limburg & Elfman 2017); with further development, this approach could provide a powerful tool for detecting seasonal oscillations and identify spatial patterns in isotope concentration to focus future one-dimensional analyses on. Additionally, further exploration of the periodic table may yield more chemical tracers with applications in novel environments such as deep-sea marine trenches or suboxic environments such as in oxygen minimum zones.
Overview and future directions
Overall, for chemical patterns to be broadly applicable in age estimation, future efforts should be directed towards understanding the mechanisms regulating elemental uptake and incorporation in space and time, validating seasonality of the observed patterns in the field, and identifying statistical methods that provide a more objective estimate of age than the ones used to date. Age estimation based on otolith chemistry is not yet cost-efficient enough to replace traditional visual growth increment reading directly. The approach, however, provides crucial calibration for assessing the accuracy of traditional age readings or size at age information used in length-based assessment models, in particular for species or populations with ambiguous growth increments. Depending on intended use, cost–benefit analyses will help identify to what extent the use of chemistry-based age estimation is sensible.
Environmental stress and ecophysiology
The use of archival tissues to identify exposure and responses to environmental stressors in aquatic ecosystems is one of the most promising applications of otolith chemistry. Otolith chemistry reflects external and internal processes expressed within a temporal framework. Therefore, variations in otolith tracer composition can indicate the extent, frequency, and duration of exposure to environmental stressors (external influences) as well as aspects of the individual’s response to those stressors (internal factors). Such information is valuable for resource managers to disentangle potential exogenous stressors on fish populations (i.e., beyond fishing effects). However, to extract the maximum information from otolith chemical markers reliably, we need a clear mechanistic understanding of the processes underpinning variations in otolith chemistry, whether internal, external or both. We also need to be mindful of the implications of sampling processes, particularly survivor bias. Significant advances have been made towards understanding the relationship between physiology, environmental conditions and otolith elemental proxies for environmental stressors such as hypoxia and selenium toxicity, and for internal tracers such as hormone residues and metabolic rate (e.g., Chung et al. 2021b; Johnson et al. 2020; Limburg & Casini 2019). More progress has been made where the biochemistry underpinning the proxy is well understood and where validation studies have been conducted to provide robust interpretations of observed patterns in nature for particular species and environmental conditions, although experimental evidence for some influences (e.g., temperature and salinity) have been contradictory (Hüssy et al. 2021b; Izzo et al. 2018). The need for specialized validation studies relating environmental conditions to elemental values in otoliths is one of the primary factors limiting broader application of otolith proxies to quantify environmental stressors.
Environmental stressors
Hypoxia
Reconstructing lifetime exposure to low oxygen conditions requires redox-sensitive geochemical proxies that respond to aqueous dissolved oxygen concentrations and are reliably incorporated into biogenic structures such as otoliths. Manganese appears to meet these criteria in many marine, estuarine and freshwater habitats (Limburg et al. 2015). Mn has varying solubility depending on its valency: in basic and oxic settings Mn is present as highly insoluble oxides, whereas in acidic and reducing (anoxic) waters Mn2+ ions are relatively soluble and available for biological uptake. In addition to fluxing predictably from organic-rich sediments to overlying waters during deoxygenation events (Lenz et al. 2015), and below aquaculture facilities (Adey et al. 2009), Mn has been categorized as an intermediate acid in otoliths that appears to be strongly driven by ambient water concentration at the time of otolith increment accretion (Mohan et al. 2012; Sturrock et al. 2015a). However, endogenous factors may also impact uptake dynamics given the impact of hypoxia on metabolism and growth rate. Some researchers have proposed accounting for these endogenous impacts by ratioing manganese to magnesium (Mn/Mg) to produce a hypoxia indicator normalized for metabolism (Limburg & Casini 2018), given the hypothesized influence of metabolism on magnesium uptake (Limburg et al. 2018). Other potential otolith-derived hypoxia indicators include iodine (Lu et al. 2010) and δ34S (Limburg et al. 2015), although further experimental validations are needed.
The use of hypoxia indicators in fish otoliths has expanded in recent years across species and systems to understand the direct and indirect impacts of this growing environmental stressor. This includes quantifying historical shifts in duration and magnitudes of hypoxia exposure in Baltic cod that are associated with worsening fish condition, size, and performance (Limburg et al. 2011; Limburg & Casini 2018, 2019). Conversely, condition and size-at-age do not appear to be strongly affected by hypoxia exposure in the first year of life for Atlantic croaker (Micropogonias undulatus) in the Gulf of Mexico (Altenritter et al. 2018; Altenritter & Walther 2019) nor flounder (Platichthys spp.) in the Baltic Sea (Samson 2021), perhaps due to their relatively higher physiological tolerance to hypoxia exposure. However, coupled measurements of otolith Mn and tissue stable isotopes revealed that hypoxia displaces some individual Atlantic croaker from benthic to pelagic food webs, while others remain feeding on benthic prey, leading to an overall expansion of trophic niches for hypoxia-exposed fish (Mohan & Walther 2016; Steube et al. 2021). The complexity of lethal and sublethal effects of hypoxia on fishes and their associated ecosystems are manifold, and the expanding use of geochemical proxies to assess their impacts hold great promise for a more comprehensive understanding of ecosystem level responses.
Contaminants
Aquatic ecosystems are increasingly threatened by human-mediated sources of contaminants (Guédron & Acha 2021; Schwarzenbach et al. 2006). Elevated levels of metal contaminants have been measured in archival tissues such as otoliths, fin rays, and eye lenses, to monitor elevated exposures and corroborate water quality concerns (e.g., Dove & Kingsford 1998; Mounicou et al. 2019; Selleslagh et al. 2016; Sun et al. 2019). Contaminants such as selenium have also been measured in otoliths from several species and systems to evaluate increased exposure. These include polluted lakes in New York, areas downstream of coal mines in British Columbia, and in freshwater and estuarine habitat impacted by oil refineries and legacy agricultural practices (Friedrich & Halden 2010; Johnson et al. 2020; Limburg et al. 2010). Detection of sublethal levels of contaminants in biominerals is a promising approach for diagnosing sources, pathways, and potential for cumulative exposures relevant to aquatic conservation (Barbee et al. 2014; Johnson et al. 2020). Traditional toxicological investigations resolve only recent exposures whereas contaminants deposited in biominerals can be used to track provenance, life stage, and history and duration of exposures over the lifetime of an animal. Ultimately, otoliths allow us to link past sublethal exposures to long term impacts (e.g., suboptimal early growth) and resulting poor quality phenotypes (Barbee et al. 2014). Combining contaminant chronologies with other assessments of fish conduction such as growth, deformities, modified behaviours, and stress responses will assist managers and regulatory agencies in assessing the magnitudes and impacts of contaminants on fishes (Barbee et al. 2014; Bengtsson et al. 1985; Johnson et al. 2020).
Whilst a wide variety of metal contaminants have been measured in both otolith inorganic and organic components of fish from contaminated areas (Hüssy et al. 2021b; Limburg et al 2010; Selleslagh et al 2016), elements such as mercury and other complex chemical compounds have rarely been detected in otoliths, restricting a broader application for some classes of contaminants, in part due to limitations in measuring these compounds with ICPMS approaches. Overall, structures with purely organic matrices such as eye lenses (Dove & Kingsford 1998), or with high organic contents such as scales (Dua & Gupta 2005; Rahman et al. 2018; Sultana et al. 2017) or fin rays (Sun et al. 2019), may show greater promise for diet-based sources of contaminants (reviewed in Tzadik et al. 2017). Ultimately, to advance the application of otoliths as markers of contamination stress in fish, further research is needed relating values in the environment, fish tissues and otoliths. And also on the role of fish physiology in the transport, regulation, and incorporation of environmental contaminants into tissues and the existence of potential non-linear relationships in the incorporation of contaminants, in particular at high environmental concentrations or under stressful conditions (Bendell-Young & Harvey 1986). Nevertheless, otolith chemistry can augment understanding of contaminant histories by its complementary information (e.g., providing location at time of contamination).
Temperature
Marine heatwaves are extended periods of anomalously warm ocean temperatures that can have unprecedented impacts on marine species and major fisheries (Cavole et al. 2016; Sanford et al. 2019; Santora et al. 2020). They are increasing in frequency with one of the most common responses of organisms to warming ocean conditions being a shift in species distributions and/or spawning phenology. Migratory species that use multiple aquatic habitats to complete their life cycle are vulnerable to shifts that can have impacts at multiple life stages [e.g., California Chinook salmon (Oncorhynchus tshawytscha) exposed as juveniles to drought conditions and anomalously warm ocean conditions as adults (Johnson et al. 2017)]. Oxygen isotope compositions (δ18O values) in foraminifera, corals and other biomineralising invertebrates are used in paleo-temperature reconstructions because of the predictable, largely abiotically driven temperature-dependent fractionation (Eakin & Grottoli 2006). Similarly, δ18O values in otoliths can be used to reconstruct temperature if the δ18O of water is known (Weidman & Millner 2000; Willmes et al. 2019).
Advances in microdrilling and probe-based δ18O measurements have allowed temperature profiles to be revealed across the lifetime of fishes at high temporal (days to weeks) and thermal (< 1 ℃) resolution (Hanson et al. 2010; Sakamoto et al. 2019; Zazzo et al. 2006). As temperature regimes in the world’s oceans shift with climate change, the ability to track changes in realised thermal exposures of individual animals as well as their distributions will become increasingly important. For fish species that are constrained (e.g., in riverine networks or place holding taxa) and unable to move to optimally suitable temperatures, δ18O in otoliths can be used to quantify temperature-dependent mortality, thermal sensitivity of performance and temperatures experienced by survivors. Noting that, warming may increase performance where the ambient temperature is below the ecological thermal optimum for the species or population. Chemical thermometry of biomineralized tissue should also be compared to optical properties of otoliths for additional information, as daily increment formation is highly sensitive to temperature (Volk et al. 1999). Continued laboratory studies are necessary especially for species living close to their thermal limits since there is some evidence that growth rate, otolith precipitation rate, and metabolic effects could influence oxygen isotope fractionation (Darnaude et al. 2014; Storm-Suke et al. 2007).
Ecophysiology
Individual fish respond to changes in the external environmental conditions through behavioural modifications, physiological responses, and/or changes in performance. The nature and extent to which these responses are expressed reflects an interaction between the individual phenotype and the local environmental and ecological context. Consequently, laboratory-based studies of fish responses to external environmental drivers may have limited predictive power in complex field settings. There is therefore an urgent need for retrospective measures of fish responses to observed environmental stressors.
Otolith and other archival tissues have relatively untapped potential to reconstruct fish responses to external environmental drivers. For instance, stress responses are coordinated through the endocrine system: the expression of hormones (such as cortisol) in body tissues, including incrementally grown tissues, can indicate triggering of systemic responses to an external stressor. Similarly, an animal’s field metabolic rate reflects the sum of energy consuming processes and therefore acts as a holistic record of performance in response to the external environment. Variations in mass-specific field metabolic rates in response to changes in the external environment may indicate the relative sensitivity of whole animal performance to that perturbation. Below we briefly review the state of the art in approaches to recover markers of physiological responses to external environmental and ecological conditions.
Stress hormones
Several studies have demonstrated that exposure to stress causes an increase in the concentrations of cortisol in fish scales (Aerts et al. 2015; Laberge et al. 2019) at levels that are proportional to plasma concentrations (Carbajal et al. 2019). Exposure to temperatures outside of a species’ optimal range can activate the hypothalamic-pituitary-interrenal (HPI) axis, thereby inducing an endocrine stress response, which elevates systemic cortisol and is recorded in the scale (Hanke et al. 2019). Scale cortisol has therefore been proposed as an ideal biomarker for monitoring stress responses to climate change for fishes (Goikoetxea et al. 2021). See section Maturity and Reproduction, for more details on links between stress and reproductive hormones on archival tissues.
Metabolic rate
Responses of individuals to external conditions typically have an energetic expression, through changes in behaviour (e.g., energy partitioning) or changes in the availability or accessibility of resources. Estimating total energy use (field metabolic rate, FMR) is one way to understand how marine organisms respond to environmental pressure and changes. Field metabolic rate has been extremely challenging to determine, especially in aquatic organisms (Treberg et al. 2016). It has long been known that stable carbon isotope values (δ13C) recorded in biogenic carbonates are not in equilibrium with external waters, and that this disequilibrium reflects contributions from metabolic (respiratory) carbon. Metabolic carbon has much lower δ13C values than ambient dissolved inorganic carbon, so increased contributions of metabolic carbon to the biomineralising fluid result in lower δ13C values in the biomineral. This process was first identified in foraminifera (Berger et al. 1978) and then widely among marine organisms (e.g., Kalish 1991a; McConnaughey et al. 1997; Solomon et al. 2006). In most biogenic carbonates, the carbon forming the biomineral is dominated either by the ambient or the metabolic source. In fish otoliths (but also cephalopod shells and to a lesser extent bivalves), both carbon sources typically contribute more than 10% of the total carbon in the blood, so that the relative proportion of metabolic compared to ambient carbon provides an analytically tractable proxy for variations in metabolic rate (Chung et al. 2021a; McConnaughey et al. 1997). Otolith δ13C values have been applied to infer relative metabolic rates of teleosts in relation to their life stages, functional behaviours, and experienced temperatures (Dufour et al. 2007; Sherwood & Rose 2003; Sinnatamby et al. 2015).
In a further development, experimental work has validated a direct relationship between the proportion of metabolic carbon in the otolith δ13C and fish respiratory rates manipulated through ambient temperature (Chung et al. 2019a, 2019b). Establishing this statistical calibration between the otolith isotope proxy for FMR and oxygen consumption rate extends the use of the otolith metabolic proxy in broader ecophysiological research. A key advantage of using otoliths as proxies of field metabolic rate is that the analysis required to obtain δ13C values also returns δ18O values. Each otolith sample therefore provides an estimate of the time-averaged temperature experienced by the individual fish and the time-averaged field metabolic rate expressed at that temperature. Combined with body size data, this allows insight into thermal sensitivity and allometric scaling of FMR as well as identifying conditions (i.e., stressors) where field metabolic performance is reduced, providing added value to routine monitoring (Alewijnse et al. 2021; Chung et al. 2021b; Smoliński et al. 2021).
Mortality/selection
Environmental stressors in aquatic systems such as warming water, hypoxia, and pollutants often cause direct or latent mortality to individuals with often unquantified impacts to populations. Selective mortality during early life stages can have significant population-level consequences, yet critical periods when selective mortality occurs, the strength of selection, and under what environmental conditions, can be difficult to identify (Garrido et al. 2015; Sinclair et al. 2002). Otoliths and archival tissues are ideal structures to assess the magnitude of survival bias (mortality) through cohort-based sampling. Age reconstructions in fisheries have revealed poor recruitment of particular year-classes and growth variability due to environmental fluctuations or extreme events that may result in fish kills (Martino et al. 2019; Pinto et al. 2021; Reis-Santos et al. 2021; Wang et al. 2011). Similarly, Woodson et al. (2013) reconstructed daily growth rates in Chinook salmon from cohorts through time and demonstrated that individuals that were larger and growing faster during their early lives had greater survival than smaller and slower growing individuals during poor oceanographic conditions. Additional monitoring data can be particularly powerful when paired with otolith reconstructions to estimate the strength of selection for particular traits such as fish size, growth, or migration timing as a function of environmental conditions (Martino et al. 2019; Sturrock et al. 2015b, 2020). For example, Sturrock et al. (2015b, 2019) used routine monitoring data of fish sizes collected from juvenile Chinook salmon leaving rivers and compared them to reconstructed fish sizes at emigration using Sr isotopes in the otoliths from surviving adults from the same year-classes. This work quantified size-selective mortality as a function of hydrologic condition and identified water release strategies from reservoirs to support a portfolio of outmigration behaviours in salmon (See section: Connectivity and Movement Reconstructions Across Life-stages). A similar approach can be used linking water temperatures and fish distributions at early life stages and temperature-dependent mortality reconstructed from survivors in that cohort using δ18O in otoliths. Conversely, consistent observed chemical patterns over time may indicate a lack of stress-selective mortality. For instance, Altenritter and Walther (2019) used multivariate analyses of otolith chemical proxies of hypoxia exposure patterns during the first year of life (e.g., duration and magnitude) and found that exposure patterns of surviving Atlantic croaker did not change significantly over time. Thus, this species appeared to be relatively tolerant to hypoxia exposure, at least for the life history stages that were sampled. Future efforts should aim to explore shifts in demographic metrics recorded in archival tissues linked to landscape scale environmental stressors.
Overview and future directions
Overall, chemical tracers in biomineralised tissues of fishes have been used to infer aspects of the external environment such as water temperature, pollutant exposure, or hypoxia. However, alteration of biomineral chemical composition based on internal processes, or retention of metabolites or hormones within the biomineral provides a concomitant signal to infer physiological responses to the external environment. Biomineralised tissues can therefore offer a simultaneous record of the change in the external setting, together with a record of the physiological and/or behavioural response to that stressor, all within a temporal context offered from the incremental nature of the tissue. Furthermore, by working retrospectively using tissues collected from free-swimming wild fish, we record the realised effects of a stressor on individuals after they have taken whatever mitigation actions are available to them within the specific ecological/environmental context. Identifying responses that cannot be behaviourally mitigated or that are overcome at physiological cost is arguably more ecologically relevant and more pertinent to management than determining lethal limits in laboratory contexts.
All biomarker approaches described here can provide more information when used in combination with other biomarkers, including tagging and gene markers. In combination such data will make significant progress in validating and evaluating individual responses to environmental pressures, like warming, hypoxia, harvesting on fish and fisheries. However, it is important to note that studies drawing on retrospective analysis of biomineralised tissues recovered from live caught individuals commonly suffer from survivor bias. One remedy will be to track early life cohorts occurring in stressed environments, taking advantage of daily growth increments to track mortality in combination with chemical information. In the context of environmental stressors, sampling natural mortalities and/or animals in poor condition is important, but may be complicated where a lack of growth reduces accretion of the biomineralised tissue.
Seafood provenance
A significant challenge for fisheries management and the seafood industry is to accurately pinpoint the harvest location of seafood products (used here in its broadest sense to encompass both freshwater, estuarine and marine products). Unlike most food commodities, many seafood products are sourced from wild populations, where high seas transshipping and processing operations allied to complex and obscure supply chains make it increasingly challenging to trace product origin, and provide a backdrop for illegal and fraudulent activities (Fox et al. 2018; Kroetz et al. 2020; A. J. Silva et al. 2021). In this context, seafood provenance and forensics is an emerging area of research driven by the growing recognition that malpractices undermine sustainable fisheries, erode management and conservation efforts, and pose potential risks to consumer health (Fonseca et al. 2022; Leal et al. 2015; A. J. Silva et al. 2021).
Provenance (i.e., the geographic area of collection or production of seafood), together with product authenticity (i.e., a product is genuine regarding species, wild vs farmed, or location of origin) and traceability (i.e., means to follow a product along the supply chain) are straightforward concepts and key components in seafood, fishery and aquaculture regulations (e.g. EU, No 1379/2013), but their implementation requires effective tools for testing and verifying information (Carter & Chesson 2017; Cusa et al. 2021; Gopi et al. 2019a). Therefore, capitalizing on the intrinsic properties of calcified structures and their chemical composition provides an opportunity to develop systems to validate seafood provenance. In doing so, we are empowering managers to safeguard legal and sustainably managed fisheries, as well as guarantee compliance and the legitimacy of certification programmes (Armenta & de la Guardia 2016; Leal et al. 2015).
Chemical and isotopic composition in calcified structures such as otoliths or bivalve shells have long been used by fishery scientists and ecologists to inform stock or management units (Albuquerque et al. 2016; Avigliano 2021; Tanner et al. 2016), and discriminate habitat use at small and large scales (see Section on Connectivity and references therein). These intrinsic markers have the advantage that all individuals are naturally marked, reflect geographical conditions mediated by variations in the surrounding environment (e.g., water chemistry, salinity, temperature) (Campana 1999; Izzo et al. 2018; Reis-Santos et al. 2013a), and therefore can also underpin provenance assessments. Critically, calcified structures allow retrospective analyses through the lifetime of the individual that can be used for more detailed assignments among locations, or a potential tracer for interventions during aquaculture [e.g., changes in rearing conditions (salinity, hypoxia, feed, other treatments), mass-markings for origin identification]. Despite these benefits, the application of natural chemical markers in calcified structures specifically for seafood provenance is surprisingly minimal (see Table 1). Furthermore, of the known examples, few studies have used LA-ICPMS (Daryanani et al. 2021; Pereira et al. 2019), even though this is by far the leading analytical method used by ecologists to analyse the trace element chemistry of biominerals. Compared to solution based ICPMS or IRMS of soft and hard tissues that dominate seafood provenance research and where samples are digested in acid before analysis (Cusa et al. 2021; Gopi et al. 2019a, 2019b), LA-ICPMS has minimal sample preparation, low contamination risk, and allows precise analytical spatial resolution of a large number of samples. Whilst instrument availability still varies across the globe, these traits have made LA-ICPMS routine outside the seafood provenance field, and we should capitalize on its high throughput to shift the current model and advance the implementation of LA-ICPMS to speed up response times for regulators aiming to safeguard compliance but also claims on geographic origin.
Provenance assessments based on otolith chemistry are to date centred in distinguishing wild versus farmed seafood products (Arechavala-Lopez et al. 2016; Pereira et al. 2019) (Table 1). In this case, both studies successfully trace back wild and farmed fish from different origins, with Pereira et al. (2019) also combining the use of radiogenic markers (87Sr/86Sr) and δ13C from otoliths to discriminate provenance of freshwater fish along a large expanse of the Amazon. While we acknowledge there are other otolith studies that could be usefully applied to the question of seafood provenance (e.g., Gillanders & Joyce 2005; Johnson et al. 2012) we limited Table 1 to studies that directly address seafood provenance. Considering other calcified structures, recently, the chemical composition of octopus statoliths demonstrated these are a powerful tool to track the provenance of octopus at regional/domestic and international scales (Daryanani et al. 2021; Martino et al. 2021). Scale chemistry has also identified wild versus farmed salmon (Adey et al. 2009). But, thus far, bivalve shells are the most commonly used calcified structure to assess provenance, where chemical composition can reveal harvest period (Morrison et al. 2019) and has shown incredible spatial resolutions (down to 1 km apart) (Ricardo et al. 2015), with the influence of harvest location strong enough to overcome species-specific variability and allow the use of species as proxies of others (Mamede et al. 2021).
Building reference libraries of geographical locations of interest is key to the successful application of otoliths and other calcified structures to test if particular geographic claims are true. Alternatively, spatial models of chemical variation can be developed (e.g., isoscapes or maps of elemental variation), ideally encompassing seasonal or other temporal variations (Cusa et al. 2021; Gopi et al. 2019a) (like in connectivity and movement reconstructions, see respective section). Such baselines boost the certainty of the retrospective assignment of seafood provenance and legitimise managers and regulatory agents to support sustainable and well-managed fisheries, but also underpin denomination of origin programmes, identify wild versus farmed, or discriminate sources of aquaculture products. While in need of further development, portable X-Ray Fluorescence (XRF) could also be a game changer enabling real-time, on site, forensic applications for seafood provenance.
Provenance testing can be framed as a set of questions with increasing requirements in terms of reference data and assumptions (Cusa et al. 2021). The simplest type of question requiring the least additional data can be framed as:
- (1):
-
Is the test sample consistent with coming from a specific, spatially restricted, and known place? In this case, the sample design requires testing against reference data or estimates of the chemical matching that of the claimed origin, with the spatio-temporal scale of the reference data ideally matching that of the claimed origin. Note that matching consistent patterns may not always indicate origin, as there is potential for unsampled regions to also match – but identifying mis-matching tracers can demonstrate that a claimed origin is unlikely.
- (2):
-
Which of a set of discrete sites is the most likely origin? This question is an extension of the above, simply requiring additional reference datasets capturing the likely tracer composition among sites. Addressing the question adds extra sample complexity as each potential origin site should be characterised with equal efficiency in the reference. Noting that the effective likelihood of assignments to any site is likely to reduce as the number of candidate sites increases – therefore choice of candidate sites is important.
- (3):
-
Where did the test sample likely come from? This is by far the most challenging application, as it requires complete characterisation of all possible origin areas. Successful applications of biomarker approaches to unconstrained provenance problems are likely to be limited to broader questions (e.g., comparing temperate to tropical origins, distinguishing wild from farmed).
In all three of these cases, demonstrating the accuracy of the approach can be challenging. Assignment accuracy is often estimated by sub-sampling the population used to create a reference dataset and assessing the likely origin of that subsample using decision rules built from the reference population. Clearly in this case the reference and test populations are not independent, and estimates of assignment accuracy are estimates of maximum possible assignment accuracy. There are very few examples in the seafood provenance (but also stock structure or connectivity) literature where geographic assignment accuracy has been estimated using genuinely independently-collected datasets (Cusa et al. 2021).
Although the use of natural biogeochemical tracers in seafood provenance is increasing, most of these applications draw on the chemical composition of organic tissues. This is understandable where seafood products are traded as processed, and whereby calcified structures are typically removed. However, when calcified tissues are retained (e.g., whole seafood or partially processed seafood) they are a formidable tool for establishing provenance. A major advantage of otoliths and other calcified structures for provenance assessments is that the natural chemical markers are tamper resistant, as they are permanently locked in the calcified matrix, and collections to develop reference libraries can be easily compiled, as these samples can be stored dry for decades, with minimum maintenance. Importantly, otolith collections are widely available, as otoliths are routinely used in global fisheries assessments, and can be collected at the point of landing (often without decreasing product value) or throughout the supply chain in specimens that are traded whole. Yet, existing monitoring programmes and otolith repositories remain underutilised for provenance assessments and to build reference libraries or maps of chemical variation. Ultimately, otoliths and calcified structures are an untapped resource that can enable the development of a clear and easy to implement framework to underpin compliance and food labelling laws by identifying the geographical origin of wild and farmed seafood.
Land-sea connectivity and lifetime movement reconstructions
Maintaining connectivity and life history diversity in freshwater and diadromous fishes
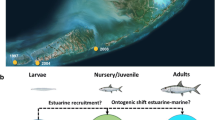
To cope with climate change and increasing population demands, water infrastructure and hydropower developments are rapidly increasing world-wide (Winemiller et al. 2016). These developments have the potential to further fragment and simplify freshwater and estuarine habitats and threaten the migratory pathways of fishes (Silva et al. 2018). Understanding the potential impacts of planned and future infrastructure is critical to address the practical challenges of river development and to protect feeding, spawning and dispersal habitats (Hall et al. 2011). Traditionally, it was thought that migration pathways were mostly important for adult fish (Gillanders et al. 2015). But recent research has identified that movement among habitats is important from eggs-to-adults (Grüss et al. 2011). For instance, eggs and/or larvae may have extended drifting periods across many kilometres (Lechner et al. 2016). Juveniles are known to migrate significant distances both upstream and downstream to access critical habitats. Adults might perform spawning or feeding migrations. It is important to understand the movement requirements of each life stage if fish are to be protected across their entire life across river watersheds (Northcote 1984) but also land- to seascape for diadromous species. Furthermore, maintaining inter and intra-specific diversity in life history pathways and timings (e.g., contingents or portfolios of migratory strategies, Fig. 2) allows fish to respond to changing environmental conditions and is critical for resilient fisheries and ecosystems (Des Roches et al. 2021; Moore et al. 2014; Schindler et al. 2010). This requires quantifying individual fish movements among different habitats and across all life-stages.

Life history diversity of juvenile Chinook salmon (Oncorhynchus tshawytscha) in the California Central Valley revealed by otolith strontium isotope profiles measured in the otoliths of post-spawned (i.e. ‘successful’) adults, emphasizing A the temporal diversity in emigration timing of spring run salmon (both from their natal river and into the ocean), and B the spatial diversity exhibited by nomadic winter run, which exhibit frequent stop-overs in non-natal tributaries on their way to the ocean (habitat colours indicated on inset map). The mean isotopic value of each population’s natal river and the ocean are indicated by short and long dashed horizontal lines, respectively. In B the spawning area is indicated by a star and black profile lines indicate fish that reared exclusively in the natal river. Data used with permission from A Cordoleani et al. (2021) sourced from https://doi.org/10.5061/dryad.bk3j9kdc9 and B Phillis et al. (2018)
Otolith analyses have provided unique insights into the spatial and temporal scales of fish migrations, revealing diverse migratory pathways and allowing the categorisation of associated life-history traits (Campana 2005; Cordoleani et al. 2021; Starrs et al. 2016). In some cases, the spatial scale of migration can be vast, and otolith chemistry has been particularly useful, for instance, in identifying diadromy. This is due to substantial chemical differences between marine and freshwater environments, defined by salinity gradients, and their incorporation into the otolith structure which is typically expressed as the ratio of strontium or barium to calcium (i.e. Sr:Ca or Ba:Ca) (e.g., Elsdon et al. 2008; Gillanders et al. 2015; Walther & Limburg 2012). Migrations that occur solely within freshwater require a different approach, and changes in otolith 87Sr/86Sr ratios can represent movements of individuals between isotopically distinct regions (Barnett-Johnson et al. 2008; Hegg et al. 2013), including within and among rivers (Brennan et al. 2019a; Hermann et al. 2021; Humston et al. 2017; Tran et al. 2021; Walther & Limburg 2012). This latter approach has recently revealed exceptionally long-distance migrations (> 8000 km) by goliath catfish (Brachyplatystoma rousseauxii) in the Amazon Basin, including natal homing (Duponchelle et al. 2016).
In many cases, the combination of multiple geochemical tracers can be used to enhance the life history information gained from otoliths (Avigliano et al. 2021; Crook et al. 2016; Macdonald & Crook 2010). For example, using 87Sr/86Sr ratios alone Duponchelle et al. (2016) were unable to demonstrate natal homing to the upper Amazon River due to less distinct environmental gradients. A follow up study undertaken by Hauser et al. (2020) used Sr:Ca and Se:Ca ratios to confirm natal homing to both the Madeira and Amazon rivers. The large spatial scale of these movements highlights the complexities of fisheries management, particularly where movements cross international borders (Cooke et al. 2012). While otolith chemistry has been instrumental in quantifying the spatial scale of movements, including highlighting long-distance migrations, it has also identified fine scale movements and substantial variability in life-history traits within species. For example, Bell-Tilcock et al. (2021) showed that otolith and eye lens δ34S values could be used to reconstruct movements between main channel and floodplain habitats. Phillis et al. (2018) used otolith 87Sr/86Sr ratios to show the importance of short excursions into small tributaries along the main migratory corridor, demonstrating the importance of maintaining an interconnected riverscape (Fig. 2). Many species exhibit phenotypic variability in migratory tendencies and a single stock can be comprised of multiple contingents, often including both residents and migrants (Chapman et al. 2012a; Secor 1999). Termed partial migration, the prevalence of this phenomena is increasingly being identified through the application of otolith chemistry in freshwater, estuarine and marine environments including across salinity gradients (Chapman et al. 2012b; Gillanders et al. 2015; Hobbs et al. 2019). Finally, otolith chemical analyses have uncovered complex life history portfolios across many different fish species and ecosystems which have implications for management strategies (Brennan et al. 2019a; Hegg et al. 2015; Hodge et al. 2016; Schindler et al. 2010; Vu et al. 2022). Therefore, only by embracing approaches that quantify individual fish movements within populations can we protect ecological heterogeneity and the critical habitats supporting our fisheries. Fisheries and habitat management should consider a holistic approach that recognises that diverse migratory strategies are an integral component of functional land-seascapes and key to safeguarding population resilience and fisheries stability. The future challenge is to link plasticity in individual fish movements to population-level outcomes, requiring innovative data analysis approaches that maximise the information stored in otoliths to reveal both population connectivity but also the factors driving individual variation in migration pathways, timing and survival (see also Food Webs section).
Linking individual movements with growth and size information
Otoliths’ time-keeping properties allow linking chemical measurements with daily and annual growth rings to reconstruct the age and timing of individual movements, as well as habitat or region-specific growth rates (see also Environmental Stress and Ecophysiology). Several recent studies have combined otolith age, growth and elemental or isotopic measurements to demonstrate the importance of divergent movement strategies under varying climatic conditions and differences in growth performance among phenotypes and habitats (Barrow et al. 2021; Cordoleani et al. 2021; Gillanders et al. 2015). In freshwater systems characterized by sharp changes in geology, otolith 87Sr/86Sr ratios have the potential to reconstruct individual migration pathways at spatiotemporal resolutions comparable to acoustic and data storage tags (Phillis et al. 2018). Emerging time series analyses such as space‐time isotope models (Brennan et al. 2019b) and Dynamic Time Warping (Hegg & Kennedy 2021) are increasing the sophistication of such analyses. Such detailed life history reconstructions provide unique opportunities to reveal the impact of specific management actions, such as whether habitat restoration or reservoir releases resulted in appreciable changes in growth, or whether pulse flows cued fish movement (Sturrock et al. 2015b, 2020). Even without time-consuming microstructural analyses, the positive relationship between otolith size and fish size (usually particularly reliable for early life stages) means that the relative growth obtained in different habitats can be estimated (Brennan et al. 2019a, 2019b). Thus, by reconstructing the life of a fish from egg to adult, we can better-understand migratory pathways and provide these data to inform policy decisions and reduce the impacts of future developments across the land-sea interface.
While fine scale movement and growth reconstructions have typically been applied in freshwater systems or between fresh and marine environments due to larger chemical gradients and thus higher spatiotemporal resolution, there are also applications for fully marine species. For example, growth, movement, thermal and metabolic reconstructions in marine fishes using otolith microstructure in tandem with δ18O and δ13C values is gaining traction (Chung et al. 2019b; Sakamoto et al. 2019; Trueman & St John Glew 2019) with finer sampling and detection capabilities through SIMS (Hanson et al. 2010) and new micromilling techniques (Sakamoto et al. 2019) increasing the spatiotemporal resolution possible (See also Environmental Stress and Ecophysiology).
Natal origin, and estuarine and coastal nursery contributions
Many marine fish species use spatially segregated habitats over their life cycle, connecting spawning and nursery areas via pelagic larval dispersal (Cowen & Sponaugle 2009), and juvenile nursery areas and adult populations via recruitment (Beck et al. 2001). Understanding and quantifying these connectivity links is key to assess population structure and dynamics, and to identify important areas and pathways for dispersal and movement, thus providing essential information to ensure management at biologically relevant spatial scales (Gillanders 2002b; Kerr et al. 2016; Phillis et al. 2018; Reis-Santos et al. 2018a). Once more, otolith chemistry provides a unique and versatile approach to identify and quantify connectivity patterns between different life stages of fish, enabling managers to adopt and support fishery and habitat protection decisions.
The pelagic larval stage that connects spatially segregated spawning grounds and nursery areas is heavily influenced by physical processes allowing for large-scale dispersal and connectivity between populations. Yet, evaluating connectivity during the larval stage is initially dependent on the ability to discriminate between individuals from different spawning grounds. This can be achieved by analysing the elemental chemistry of otoliths collected from developing embryos and recently hatched larvae in both marine (Brophy et al. 2003; Rogers et al. 2019; Schaffler et al. 2009; Tripp et al. 2020) and freshwater environments (Barbee & Swearer 2007; Lazartigues et al. 2018). Subsequently, the elemental signatures for each spawning ground can be compared to the natal signatures of juveniles collected from nursery areas to establish larval connectivity and quantify the relationships between specific spawning grounds and nursery areas (Shima & Swearer 2016; Walther & Thorrold 2010). Because elemental signatures and dispersal patterns can change over multiple temporal scales (Avigliano et al. 2018; Gillanders 2002a; Reis-Santos et al. 2012), it may be necessary to repeat studies, or build libraries of chemical signatures over multiple years to understand variability in dispersal and subsequent recruitment to nursery areas (akin to building reference libraries to test Seafood Provenance). An alternate approach is to simulate dispersal using biophysical models (Swearer et al. 2019). The biological component of these models describes the ontogenetic development of the larvae, which can be retrospectively determined from the otolith microstructure of settled juveniles (i.e., spawn date, growth rate, settlement date) (Rogers et al. 2021; Vaz et al. 2019). Furthermore, the simulated patterns of larval dispersal can be compared to the relationships identified from otolith biomarkers to assess model predictions and best interpret connectivity.
Estuarine and coastal areas are often cited as important nursery habitats (Beck et al. 2001; Gillanders et al. 2003). Juvenile fish recruit into the sheltered estuarine habitats settling in structured habitat such as seagrass, mangroves or saltmarsh where there can be abundant food and reduced numbers of predators. At larger sizes and ages, they then move out to coastal habitats including reefs. In some cases, fish can recruit to both estuarine and reef habitat as juveniles but adults are only found on the reef. Overall, otolith chemistry provides a means of determining the origins and movement of fish, allowing the estimation of contribution rates of the different nursery sites (e.g., Gillanders 2002b; Reis-Santos et al. 2013b). Many studies have demonstrated differences in otolith chemistry among estuarine and coastal habitats (Brown et al. 2019; Russell et al. 2021; Tanner et al. 2012), which is a key precursor for determining connectivity between habitats. To demonstrate connectivity additional steps are required including being able to match adult fish back to estuaries or juvenile habitats within estuaries and coastal areas using classification approaches (Chittaro et al. 2009; Gillanders 2002b; Reis-Santos et al. 2013b; Tanner et al. 2013). Issues may occur with increasing the number of baseline sampling sites such that sampling sites are often aggregated (Gillanders 2002b). Alternatively, spatially explicit approaches involving continuous assignment where boundaries between sampling sites are not delineated have recently been used on otolith elemental data (Burns et al. 2020). Such an approach building on models of chemical and isotopic variation (i.e., isoscapes) is likely to be particularly beneficial for fisheries and ecosystem-based management, including to identify critical areas for stock replenishment or target areas for conservation (McMahon et al. 2013; Trueman & St John Glew 2019).
In fully marine settings and for highly migratory fish species with ocean-wide or cosmopolitan distribution such as tunas and billfishes, otolith chemistry has also been used to investigate nursery area use and contribution rates as well as adult movements (Artetxe-Arrate et al. 2021; Fraile et al. 2016; Rooker et al. 2008, 2021; Wells et al. 2021). These species regularly cross management boundaries, often migrating large distances to complete their life cycles and some exhibiting homing behaviour that further complicates stock assessment and management efforts (Collette et al. 2011). Since the physicochemical characteristics of the marine environment are rather homogenous compared to inshore areas (e.g., estuaries) the investigated nursery areas in these species are generally farther apart and the analysed otolith chemical composition includes both trace elements and stable isotopes (Artetxe-Arrate et al. 2021; Fraile et al. 2016). Otolith stable isotope δ18O values are particularly reliable markers at large spatial scales (cross ocean basin) due to lower temporal variability compared to other tracers (Rooker et al. 2008; Trueman & St John Glew 2019). As above, baseline libraries or models of chemical and isotopic variation of known nursery areas can be used to then assign adults retrospectively to different nursery/natal areas. Moreover, adult movements in these highly migratory species can also be assessed by analysing the chemical composition along the otolith growth axis to provide information over the lifetime of the individuals (Fraile et al. 2016) or by comparing capture location with natal or nursery origin based on otolith core signatures (Artetxe-Arrate et al. 2021; Rooker et al. 2008). Like individual migration pathway reconstruction in freshwater environments (see above), time series analysis approaches, such as characteristic-based clustering or behavioural change point analysis, are being increasingly applied to otolith chemical transect data allowing movement reconstruction over individual lifetime and providing valuable continuous information on habitat use patterns and larval dispersal trajectories (Sabetian et al. 2021; Shima & Swearer 2016). Ultimately, technological advances will contribute to increased spatio-temporal resolutions in movement reconstructions (Hanson et al. 2010; Sakamoto et al. 2019) but a key next step to consolidate ecosystem-based management is identifying the mechanisms that determine variations in movement behaviour as well as its consequences. Tying movement to other biomarkers in otoliths will empower managers to have a broader perspective on the significance of differential movement for growth performance to evaluate fish responses to environmental variations, as well as its implications on reproductive success, population biomass or trophodynamics (Chung et al. 2019b; Grønkjær et al. 2013; Oxman et al. 2021; Reis-Santos et al. 2021; Sturrock et al. 2015a) (see sections on Environmental stress and Ecophysiology, Food Webs, and Reproduction).
Food webs and species interactions
Changes in fish diet occur as fish grow, and these ontogenetic shifts often present critical bottlenecks for survival and recruitment. Increasingly, fisheries management aims to include dietary information when modelling sustainability and fish population projections. While still not implemented at a large scale, ecosystem-based fisheries management relies on multi-species models that account for species interactions, of which predator–prey relationships are paramount (Eddy et al. 2021; Heymans et al. 2016). Ecosystem models often include mechanisms that account for prey availability, body size of consumers and prey, natural mortality from predation, and the different trophic positions of fishes across their life history stages (Heymans et al. 2016; Pethybridge et al. 2018). Therefore, effective ecosystem-based fisheries management requires comprehensive assessments of trophic interactions and dietary requirements for fish survival and reproduction.
In modern fishes, stable isotope compositions of nitrogen (δ15N values) and carbon (δ13C values) are used in conjunction with stomach contents to estimate trophic positions and basal sources of primary production, respectively (Boecklen et al. 2011). δ15N values reflect the weighted-average trophic position of the fish’s prey, plus a trophic discrimination factor that results from preferential excretion of the light isotope. Fish δ13C values can vary significantly within and across ecosystems, such as across estuarine salinity gradients, due to divergent sources of primary production creating isotopic shifts in baseline organic material (Fry 2002; Davias et al. 2014). Isotope analysis of soft tissues, including muscle, eye lenses, and fin clips, form the basis of trophic level analyses in the modern ocean (Boecklen et al. 2011). However, soft tissues quickly degrade and decompose, with few opportunities for measuring δ15N values in historical samples. The basis for understanding fish diets is therefore biased toward modern fishes and their recent feeding history. In addition, most soft tissue isotope signatures are time integrated and continuously change to reflect the most recent dietary isotope signatures and lifetime ontogenetic diet shifts are not preserved. Therefore, as methods advance, isotopic research on the organic matrix of otoliths, vertebrae, eye lenses or fish teeth to reconstruct dietary information is gaining momentum, allied to the chronological properties of these archival tissues, that allow ontogenetic shifts in diet histories to be resolved across time (e.g., Bell‐Tilcock et al. 2021; Matsubayashi et al. 2020). These qualities make biogenic hard parts of fishes extremely valuable for the broader goals of effective ecosystem-based fisheries management.
Methodological developments
The tools outlined below serve as ‘tracers’ of dietary signals, and our ability to interpret these signals rests strongly on controlled, laboratory experiments that validate the behaviour of the tracers. Otolith δ15N measurements have been limited by the small amount of N in fish otoliths. Nitrogen in otoliths is in the form of proteins, comprising only 0.1–3% by mass (Campana 1999; Dauphin & Dufour 2003; Thomas & Swearer 2019). Until recently, the majority of δ15N studies combined multiple otoliths to obtain enough N for elemental-analysis coupled to isotope ratio mass spectrometry (EA-IRMS) (Godiksen et al. 2019; Grønkjær et al. 2013; Sirot et al. 2017; Vandermyde & Whitledge 2008). However, advances using a persulfate-denitrifier method, allow N isotope measurements from small, individual otoliths (Cheng et al. 2018; Lueders-Dumont et al. 2018, 2020, 2022). For δ13C measurements of organics, otoliths are demineralized prior to isotopic analysis, facilitating measurements of organic C without dilution from the inorganic carbonate C that comprises most of the otolith mass (Godiksen et al. 2019; Grønkjær et al. 2013; McMahon et al. 2011a, 2011b; Sirot et al. 2017; Vane et al. 2018a). Otoliths must be demineralized as the mineral fraction is a tracer of other, non-diet related variables (see Ecophysiology section). While the advantage of the persulfate-denitrifier method is the lower mass requirements for N, one of the primary advantages of EA-IRMS methods is that both δ15N and δ13C can be measured from the same otoliths. Micromill sampling, allowing for discrete analyses from juvenile and adult life history, is emerging for δ15N (Lueders-Dumont et al. 2022; Vane et al. 2018b) and for δ13C (Vane et al. 2018a) analyses of the organic matrix, with higher resolution sampling tied to continued improvements in instrument sensitivity. δ34S values in otoliths have also emerged as a useful marker in freshwater systems to identify individuals who spent their early lives feeding in different aquatic habitats such as floodplains, rivers, and/or aquaculture facilities (Bell-Tilcock et al. 2021; Johnson et al. 2012; Weber et al. 2002).
Compound-specific isotopic analysis of amino acids (CSIA-AA) in proteins separates isotopic ‘baseline’ from ‘trophic’ patterns more clearly than bulk (non-CSIA) δ15N or δ13C analysis (Chikaraishi et al. 2009; McClelland & Montoya 2002). CSIA-AA of muscle tissues is becoming a widely used technique in ecology, providing additional insight into processes at low trophic levels and physiological processing of nutrients (Skinner et al. 2021; Wyatt et al. 2019). Several studies use CSIA-AA analyses of the otolith organic matrix (McMahon et al. 2011a, 2011b; Vane et al. 2018a, 2018b) but to date, CSIA-AA analysis of otolith proteins has only been conducted for large otoliths, as it requires greater amounts of organic N and C than bulk measurements.
For historical reconstruction of food webs using the aforementioned techniques, two additional methodological factors must be considered. First, due to potential diagenetic alteration of organic matter, cleaning methods may be required when analysing δ15N of proteins in otolith fossils as trophic archives (Das et al. 2021; Lueders-Dumont et al. 2018). Second, identifying contemporaneous baseline δ15N and δ13C changes is required for interpreting dietary or food web structural changes, as for modern studies using δ15N and δ13C in fish tissues (Boecklen et al. 2011; Post 2002). Increased sensitivity of CSIA-AA methods allowing for analysis of smaller otoliths will be a powerful tool for reconstructing past food webs as baseline and trophic isotopic signals can be measured in the same specimen. Until then, bulk δ15N and δ13C analyses of shell-bound organics of contemporaneous filter-feeding animals known to record isotopic baselines (e.g., coral skeletal material, bivalve shells; Wang et al. 2015; Das et al. 2021), can be compared with isotopic data from otoliths analysed using the methods mentioned above. Matching the spatial and temporal scale of otolith isotopic records with the isotopic baseline is required for interpreting isotopic changes in historical otoliths.
Groundtruthing stable isotope composition of otolith protein to tissues
Muscle tissues have high concentrations of organic N and C, and are the most frequently measured tissue to extract dietary information and food web structure (e.g., Boecklen et al. 2011; Post 2002). In contrast, otoliths have high concentrations of mineral (calcium carbonate), and low concentrations of organics. Studies have only recently begun to measure the isotopic composition of otolith organics, including studies comparing muscle to otolith organic matrix isotope composition. In the few studies to compare signatures, otolith matrix and muscle protein δ13C values are similar and highly correlated with one another, including using CSIA-AA (McMahon et al. 2011a, 2011b) and bulk analyses (Godiksen et al. 2019; Grønkjær et al. 2013; Sirot et al. 2017). The same is true for CSIA-AA δ15N values (Vane et al. 2018b) and bulk analyses (Cheng et al. 2018; Lueders-Dumont et al. 2020; Vandermyde & Whitledge 2008). Cross-species comparisons revealed the potential for species-level differences in the N isotope fractionation in otoliths (e.g., Atlantic cod (G. morhua, and others in the Gadidae family). Yet, otolith matrix δ15N values were always highly correlated to muscle and dietary values (Grønkjær et al. 2013; Lueders-Dumont et al. 2018, 2020), and inter-specific differences were largely explained by otolith size and biomineralization (Lueders-Dumont et al. 2020). A smaller number of studies have assessed paired muscle and otolith matrix δ2H values (Whitledge et al. 2006) and δ34S values (Bell-Tilcock et al. 2021; Godbout et al. 2010; Weber et al. 2002). Here too, otolith matrix values were similar to muscle values for both δ2H and δ34S, therefore reflecting the provenance (for δ2H) or diet (for δ34S) of the fish. The findings of the small number of studies explicitly comparing the isotope composition of soft tissue and the otolith organic matrix are encouraging, indicating the otolith macromolecular matrix preserves isotopic histories comparable to tissues such as muscle. Further field and laboratory experimental validations are needed to understand the universality of these trends and whether non-dietary physiological signals are an important factor in otolith organic isotope records.
Controlled laboratory experiments demonstrate stable isotopes of otolith organics are effective tracers of diet (e.g., Grønkjær et al. 2013; Lueders-Dumont et al. 2020, 2022), with dietary tracing ability of otoliths immune to temperature effects (Godiksen et al. 2019). Overall, otolith proteins rapidly incorporate N from diet (Shiao et al. 2018) and in timescales similar to liver, the tissue with the shortest turnover time (Lueders-Dumont et al. 2022). However, life history stage must be considered, especially for young fish, which may have smaller trophic discrimination factors, including for otoliths (Lueders-Dumont et al. 2022).
Historical ecological baselines
δ15N and δ13C measurements of historical otoliths demonstrate the potential for food web data to inform fisheries management. Gulf of Maine Atlantic cod (G. morhua) remain commercially important despite the 1990s fishery collapse, with δ15N measurements of midden otoliths from fishing outposts showing that seventeenth century cod were nearly one trophic level higher than the largest modern cod from the same region (Lueders-Dumont et al. 2018). Another study, on commercially harvested American silver perch (Bardiella chrysoura) in the Gulf of Mexico, also revealed higher trophic levels of historical fish, a shift that was attributed to anthropogenic impacts on prey and habitat availability (Sirot et al. 2017). These studies show that fisheries managers, by measuring only modern conditions, may have insufficient data for successfully managing and restoring fisheries. Some of the largest ecological changes in structure likely occurred prior to the modern management efforts. Therefore, ecosystem-based fisheries management should strive to include historical food web data to improve their predictions. Together with growth and age information from otoliths and other calcified structures, reconstructing dietary changes in populations and individuals over time may reveal links between complex life history portfolios, differential movement and migration behaviour seen across many different fish species, and changes in food webs.
We are just beginning to apply geochemical tools and other biomarkers to quantify past food webs. Reconstructing ecological baselines in food webs is an especially critical goal now, as we are likely to miss our climate goals at the same time that fish are a vital source of nutrition globally. Overall, examining how past climate changes affected diets of fishes, as well as quantifying the relative impacts of human- and climate-caused changes on food webs is crucial. Trophic level is the basis of food web structure and moreover, is known to be a plastic trait that is sensitive to shifting prey availability, fish behaviour, and fish habitat among other variables. Identifying past ecological conditions will also allow for concrete ecological benchmarks for informing ecosystem-based fisheries management.
Maturity and reproduction
The proportion of mature individuals present in a population and the frequency of reproductive events during a species’ lifetime are key parameters used to estimate stock production and manage fishery populations, and are essential for estimating spawning stock biomass, reproductive output, and yearly recruitment (Murawski et al. 2001; Quinn & Deriso 1999). For fish, collecting such data can be challenging. Size-at-age, age of sexual maturity, and reproductive life histories are commonly estimated through seasonal fishery operations, costly survey observations, and/or histological examinations. Such observations, however, provide only a snapshot in time of a population’s reproductive capacity and provide no historical perspective for individuals or populations. This limitation is concerning because reproductive stages change seasonally and the relative proportion of individuals estimated within maturity stages can vary depending on the timing of sampling such that estimation of a population’s reproductive potential can be biased (Rideout et al. 2005). Also, spawning frequency and senescence, which age-based population models often assume as constant, can vary with age and environment (Rideout & Tomkiewicz 2011). The ability to monitor reproductive reference points over time would improve stock assessments and our understanding of factors influencing reproductive success (Hutchings & Baum 2005).
Fish growth varies seasonally, and these changes are reflected in the structure of incrementally grown archival tissues (e.g., bones and otoliths). Trumble et al. (2013) demonstrated for the first time that in addition to isotopes and elements, reproductive and stress hormones could also be extracted from incremental growth zones in a baleen whale’s earplug using a modified Soxhlet method. The resulting age-specific chemical profile was used to estimate the whale’s age of sexual maturity, reconstruct its reproductive life history, and assess physiological responses to intrinsic and extrinsic factors throughout its life span. Analysis of cetacean baleen and blubber have yielded similar results (Hunt et al. 2014). Recently, these hormone extraction techniques have been successfully applied to the annual growth increments in bones of several fish species. Charapata et al. (2022) used material removed from the operculae of long lived benthic yelloweye rockfish (Sebastes ruberrimus) to reconstruct their reproductive life history (Fig. 3). The opercula of these rockfish have large, well defined annual growth increments from which material is easily sampled. Hormones associated with maturation and reproduction (progesterone, estradiol) as well as stress (cortisol) were targeted for recovery. Progesterone’s role in gamete maturation and reproductive behaviour makes it an ideal indicator of reproductive activity (Pankhurst 2016). Similarly, estradiol is associated with the formation of secondary sexual characteristics, egg development and spermatogenesis. Cortisol is a corticosteroid released in response to stress that can have both inhibitory and stimulatory effects on the endocrine system and behaviour. Although it is not directly involved in reproduction, cortisol is secreted in response to stress and can either enhance or suppress reproduction, depending on the strength of the stress response (Pankhurst 2016). Consequently, cortisol concentrations are often used to assess animal health and environmental disruptions (Chrousos & Gold 1992; Romero et al. 2009). Overall, hormone concentrations varied among annual increments, thereby allowing inferences to be made regarding age of maturity, reproductive frequency, senescence, and stress levels (Charapata et al. 2022) (Fig. 3).

Cortisol (grey), estradiol (orange) and progesterone (blue) normalized concentrations recovered from annual growth increments within the operculum via immunoassay extraction of a 20-year-old female yelloweye rockfish (Sebastes ruberrimus) caught in 2018 (Charapata, Oxman and Trumble, unpublished). The sampled region of the operculum is highlighted yellow (light shade). The entirety of each growth increment was sampled for analysis with the black line showing the trajectory of growth. The currently accepted age of maturity for yelloweye rockfish is 20 years, but peaks in reproductive hormones (progesterone and estradiol) indicated this individual began to reproduce at 9 – 10 years of age. Cortisol concentrations exceeded those of reproductive hormones between the ages of 9 and 10 but did not appear to have a negative effect. After the onset of maturity, cortisol levels declined significantly until age 18 and there was little evidence of senescence
Lifetime hormone reconstructions have also been conducted with opercula from black rockfish (Sebastes melanops) and Chinook salmon (Oncorhynchus tshawytscha) as well as vertebrae from bluefin tuna (Thunnus thynnus) (Oxman et al. 2021). Opercular bones and vertebra are preferred for this analysis because, relative to otoliths and scales which are typically used to construct biochronologies, their annual growth zones are large and provide the sample mass necessary for hormone extraction.
For management purposes, the ability to construct biochronologies using hormones stored in hard parts provides an effective alternative for assessing a population’s reproductive potential relative to more traditional approaches involving costly field surveys or laboratory studies. Moreover, for the first time, it is possible to reconstruct past reproductive activities that can provide insights into drivers of productivity, trends in abundance, and changes in size and age of maturity. This information could be used to inform forecast and assessment models within the context of environmental change thereby improving the efficacy of management.
While still in its infancy, there is growing interest in the potential for using trace element concentrations in archival tissues such as fish otoliths to reconstruct age at maturity and the frequency of reproductive cycles (Sturrock et al. 2014, 2015a) (Fig. 4, see also Clark et al. (2020), for successful application in mammalian teeth). Given the relative ease and low cost of measuring elemental concentrations at high spatiotemporal resolution across otolith growth rings using LA-ICPMS, and the widespread collection and archiving of otoliths from harvested species, this could provide an economical approach to reconstructing long term shifts in onset at maturity and skipped spawning behaviour to improve the accuracy of metrics such as spawning stock biomass. Because Zn is an important co-factor of collagen proteins, including otolin-1, and is essential for yolk formation (vitellogenesis), Zn and Ca are the elements most likely to inform these types of analyses due to the elevated concentrations needed for egg production (G. L. Fletcher & King 1978; Hüssy et al. 2021b, Thomas et al. 2019). Pb and Sr are potentially also of interest due to similar behaviours and binding affinities, respectively (Clark et al. 2020; Kalish 1991b; Sturrock et al. 2015a) (Fig. 4). While not easy to measure in otoliths, Se may also be a useful ‘spawning marker’ in the future, being important in vertebrate reproduction (Bedwal & Bahuguna 1994) and showing clear divergence in blood concentrations between mature and immature females and mature male and female fish across the reproductive cycle (Sturrock et al. 2014). The applicability of using element concentrations to reconstruct spawning events is likely to be sex, system and species-specific, and may require multivariate and/or time-series approaches. For example, it was an increase in both concentration and variability in teeth Zn and Pb concentrations that marked the onset of maturity in female walruses (Clark et al. 2020).

Potential for reconstructing past spawning events using otolith element concentrations. In tank-reared female plaice (Pleuronectes platessa), increased gonadosomatic index (GSI) was associated with increased strontium concentrations in the blood A and otoliths of mature females (B and E), and with reduced zinc concentrations in the blood (C) and otoliths of successful spawners (D and E). A to D show repeat measurements and relationships within individuals (red dashed lines; fish in E highlighted by red circles and solid lines), and the predicted line for the entire population (black line; grey shading showing 95% CI). E shows otolith Sr and Zn concentrations, GSI and temperature for one female across the year-long experiment. Inset images show a gravid plaice and a plaice otolith, with the approximate laser track shown as a black line. Data originally published in Sturrock et al. 2014, Sturrock et al.2015
As with many previous applications described above, the accuracy of chemical reconstructions will likely be augmented when used in combination with otolith and/or scale microstructural analyses, which often exhibit distinct ‘spawning zones’ relating to ontogenetic shifts in energy allocation (Denechaud et al. 2021; Engelhard & Heino 2005; Irgens et al. 2019). This is a clear area for development in the use of otoliths and calcified tissues and opens a window to go back in time and examine how harvesting and global change have impacted population productivity, maturity and reproductive cycles and to use this information to boost future ecosystem-based fisheries management strategies.
Stock enhancement and tracing
Stock enhancement, the practice of rearing fish in a hatchery and releasing them into the wild, is undertaken globally in freshwater, estuarine and marine systems to supplement natural recruitment. This management technique is often implemented where wild populations are heavily depleted or to provide recreational fishing opportunities, and has the potential to enhance other ecosystem functions. For depleted populations, stock enhancement is used to supplement natural recruitment where the potential for spawning and natural recruitment has been significantly reduced. Similarly, hatchery-reared fish may be stocked into areas where they do not naturally occur to enhance recreational fishing opportunities (e.g., impoundments, reservoirs and dams). Furthermore, it is increasingly becoming a legal requirement for hatcheries in many countries and jurisdictions to enable the identification of escaped fish from aquaculture facilities.
The ability to differentiate between wild and hatchery-reared individuals is fundamental to evaluate the performance of stock enhancement programmes and to quantify the relative contribution of natural and artificial recruitment in wild populations (Curtis et al. 2014; Warren-Myers et al. 2018). Chemical markers provide one option to differentiate between individuals. They can be artificially induced or naturally occurring and are often retained in all calcified structures within a fish (otoliths, scales, bone). Fluorescent compounds such as oxytetracycline, calcein, alizarin complexone, and alizarin red S have been widely used to mark fish for age validation experiments and have also been used for batch marking large numbers of fish for stock enhancement programmes (Fig. 5). Marking of small fish, particularly fingerlings, is best achieved via immersion in a concentrated solution, although the fluorescent compound can also be absorbed from the diet or injection in larger individuals (Crook et al. 2005, 2007; Warren-Myers et al. 2018). Alizarine complexone (ALC) is a fluorescent compound that has been used successfully to mass mark millions of juvenile red sea bream (Pagrus major) in Japan since the 1980s (Kitada 2020; Masuda & Tsukamoto 1998). Immersion in ALC has been shown to produce high-quality marks in otoliths and other archival tissues with low mortality rates (i.e., < 5%) and successfully used to quantify the efficacy of restocking (Cottingham et al. 2020; Kitada 2020).

Sagittae (top) and lapilli (bottom) otoliths from a reared juvenile snapper (Chrysophrys auratus, Sparidae) viewed under a dissecting microscope showing a purple band as a result of marking with alizarin complexone (ALC). The fish (38 mm FL, 60 days old) was immersed in a bath of 10 ppm ALC for 6 h and was sampled six days after staining. Several cohorts of juvenile snapper were marked using this method to differentiate between wild and reared fish as part of a stock enhancement programme in northern Spencer Gulf, South Australia, in 2021 and 2022. The scale bar is 500 µm (photo – T. Rogers)
Fish for stock enhancements have also been successfully marked using trace elements (e.g., Sr, rare earth metals) and enriched stable isotopes (e.g., 24 Mg, 86Sr, 88Sr, 137Ba, 138Ba,). For elements or isotopes to be suitable markers, they must have no effect on the fish, and be simple to apply, long-lasting and non-toxic (Crook et al. 2005; Woodcock et al. 2011). In addition, the cost of the mark, the application process, and its recovery from marked fish need to be considered for the long-term success of the restocking and monitoring efforts. Although Sr is widely used as an artificial tag, it is naturally occurring across a broad range of concentrations, and so differentiating between artificial and natural signatures can be challenging without a comprehensive understanding of natural variation in water Sr:Ca (Pollard et al. 1999). Subsequently, elements that naturally occur at low concentrations in the environment may provide a more reliable and easier to interpret artificial tag than Sr. Rare earth elements or lanthanides have shown variable success in marking fish especially over short time periods (e.g., Ennevor 1994; Warren-Myers et al. 2018). If either the water or food source differs between wild fish and hatcheries (as well as among hatcheries) then naturally occurring elements may also provide a suitable signature (Johnson et al. 2012; Weber et al. 2002). Similar to Sr:Ca, a comprehensive understanding of natural variation in both the hatchery and the wild is required across a range of spatial and temporal scales. To overcome potential limitations, enriched stable isotopes have been used to unequivocally mark larvae and fingerlings through immersion, dietary spiking, or via the injection of an enriched isotope into the mother (i.e., transgenerational marking). Various enriched isotopes of Ba, Mg and Sr either singly or in combination have been successfully used for marking and identifying hatchery reared fish (e.g., Munro et al. 2008, 2009; Woodcock et al. 2011).
A further method widely used on salmonids, while not a chemical one, involves taking advantage of natural variations in microstructure (Barnett-Johnson et al. 2007) or inducing marks on otoliths through changes in temperature or photoperiod (e.g. thermal marking) (Brothers 1981; Volk et al. 1999; Warren-Myers et al. 2018). Abrupt changes in the physical environment disrupt the daily periodicity of increment formation, which is manifested in the otolith microstructure as a discontinuity in the width and contrast of the daily increment. This approach is more suited towards species that can tolerate abrupt changes in physical conditions.
Numerous otolith marking techniques have demonstrated the capacity to produce distinctive marks in large batches of individuals which are very simple to apply, easily detected, and are suitable for long-term monitoring. However, the most appropriate technique will depend on the aim of the restocking effort, the number of individuals to be marked, and the availability of funds. For example, fluorescent compounds that retain a distinctive mark for a short period of time are well suited to experimental studies for age validation that consider a small number of individuals. Similarly, batch-marking of large numbers of fish via immersion is a cost-effective technique for long term stock enhancement and monitoring programmes. Combining fluorescent tags, which can be detected non-lethally in the field, with other approaches may also provide a cost-effective approach (Crook et al. 2011). In the end, managers and researchers must consider the characteristics of different compounds and select the most appropriate approach for the objective being undertaken (Crook et al. 2005; Warren-Myers et al. 2018).
Final remarks
Advances in chemical techniques have enabled researchers to tap into information derived from elements, isotopes and biomolecules recorded within the growth layers of otoliths and other incrementally grown tissues, such as eye lenses, vertebrae, operculae and scales. Increasingly, these chemical tracers are being applied for age estimation and to link the impact of differing environmental conditions on the ecology, physiology, and life history expression of fish across habitats and life stages. These emerging technologies combine otolith science with a host of other disciplines, most prominently geochemistry, ecophysiology, metabolism, food provenance and forensics, or trophodynamics, that can be leveraged to support contemporary applications of ecosystem-based fisheries management. While we have summarised several key advances in detecting environmental stressors, other stressors may yet be detectable in the future. We also note some approaches may not yet be necessarily cost-effective for widespread application, however, they already reveal management and conservation-relevant issues, and often already contribute to ongoing monitoring programmes [e.g., Sr isotopes in juvenile and adult otoliths to identify salmon province, nursery habitats, and diversity in outmigration size and timing in California (Cordoleani et al. 2021; Phillis et al. 2018)].
Given the unrivalled time-keeping properties of otoliths, the widespread collection and archiving of otoliths from harvested species provides an economical way to create large reference libraries for connectivity and movement reconstructions (Izzo et al. 2016a). These archives provide links to past ecological conditions from which we can evaluate long-term changes in fish behaviour, physiology and growth performance. Such benchmarks will inform ecosystem-based fisheries management by allowing managers to assess the impacts of stressors such as warming, food web shifts and overfishing on species and population performance. These archives will also allow managers to assess long term changes in age at maturity and skipped spawning behaviour to estimate spawning stock biomass more accurately. Ultimately, coupling multiple lines of evidence extracted from otolith biomarkers (e.g., that trace movement, growth, thermal, metabolic, reproductive, or food web structure) can greatly enhance our ability to manage fisheries more sustainably. Using otolith-derived data to highlight the significance of differential movement for growth performance, evaluate fish responses to environmental variations, as well as implications of those changes for reproductive success, population biomass or trophodynamics will deliver tangible benefits for fisheries managers and the development of ecosystem-based management efforts.
Calcified tissues also provide unequivocal natural tags to test and validate claims on geographical origin, enabling us to confirm whether a product is from where it claims to be, or to assign potential locations of origin for unlabelled products. Overall, there is an underlying need by managers and regulators to understand where aquatic food products originate from, both regarding fishery and aquaculture industries, and for this information to be verifiable. Benefiting from advances in otolith chemistry, reference libraries, and high-throughput methodologies, seafood provenance is a major area for otolith research to advance, catapulting the effectiveness of fisheries and regulatory agencies to combat food fraud (Cusa et al. 2021; Kroetz et al. 2020).
To extract the maximum information from otolith chemical markers, we need a robust and mechanistic understanding of the environmental drivers, physiological regulation and calcification processes that influence otolith development and composition (Walther 2019). Key research areas include gaining clearer knowledge of the mechanisms governing incorporation of different biomarkers into the proteinaceous and calciferous matrices of the otolith to improve the accuracy of environmental reconstructions (Hüssy et al. 2021b; Thomas et al. 2017; Thomas & Swearer 2019). Improved understanding of the environmental, ecological and phenotypic factors influencing fish responses to stressors, together with continued progress in instrumentation (including, to reduce analytical costs, increase accessibility and sample sizes) will be invaluable to drive the field forward and to increase uptake into routine monitoring and management. Better integrating tissue chemistry approaches with other individual-based measures such as gene expression, genetic markers (Brophy et al. 2020; Reis-Santos et al. 2018a; Tanner et al. 2016), and biologging (Darnaude et al. 2014; Sturrock et al. 2015a; von Leesen et al. 2021) will also help us forecast species responses to pressures such as warming, hypoxia or harvesting. Methodological advances that allow us to take advantage of genetic data archived in otoliths (Momigliano et al. 2019) will also open doorways into long-term changes in fish population structure, gene flow, and effective population size. The analyses of genetic data from otolith collections is an area poised for continued advancement, and offers unique opportunities to identify shifting baselines prior to modern management efforts (Izzo et al. 2016a), and to evaluate long term impacts of environmental stressors and harvesting on genetic diversity. Overall, the goal of using otoliths as biomarkers is to allow us to reconcile management with biological parameters (Kerr et al. 2016), and promote bespoke management actions relevant to specific species, environments, and harvest pressures.
The advances described in this article were made possible by technological breakthroughs, the development of new conceptual frameworks, and through interdisciplinary collaborations. Application of these tools also requires interdisciplinary partnerships and cross-sector knowledge transfer to support sustainable fisheries management. Over the next few years, the true value of these emerging approaches using otoliths and other archival tissues to support fisheries and boost ecosystem-based management will be measured by their successful implementation into routine monitoring. To ensure widespread uptake and application, the challenge for the research community is to continue to develop, test and demonstrate their benefits, build partnerships with managers and industry, and communicate and translate these innovations widely across academic and applied spheres.
Change history
12 September 2022
A Correction to this paper has been published: https://doi.org/10.1007/s11160-022-09729-4
References
Adey E, Black K, Sawyer T, Shimmield T, Trueman C (2009) Scale microchemistry as a tool to investigate the origin of wild and farmed Salmo salar. Mar Ecol Prog Ser 390:225–235
Aerts J, Metz JR, Ampe B, Decostere A, Flik G, De Saeger S (2015) Scales tell a story on the stress history of fish. PLoS ONE 10:e0123411. https://doi.org/10.1371/journal.pone.0123411
Albuquerque R, Queiroga H, Swearer SE, Calado R, Leandro SM (2016) Harvest locations of goose barnacles can be successfully discriminated using trace elemental signatures. Sci Rep 6:1–9. https://doi.org/10.1038/srep27787
Alewijnse SR, Stowasser G, Saunders RA, Belcher A, Crimmen OA, Cooper N, Trueman CN (2021) Otolith-derived field metabolic rates of myctophids (Family Myctophidae) from the Scotia Sea (Southern Ocean). Mar Ecol Prog Ser 675:113–131
Altenritter ME, Walther BD (2019) The legacy of hypoxia: tracking carryover effects of low oxygen exposure in a demersal fish using geochemical tracers. Trans Am Fish Soc 148:569–583. https://doi.org/10.1002/tafs.10159
Altenritter ME, Cohuo A, Walther BD (2018) Proportions of demersal fish exposed to sublethal hypoxia revealed by otolith chemistry. Mar Ecol Prog Ser 589:193–208
Andrews AH (2016) Lead–radium dating of Pacific cod (Gadus macrocephalus) – validation of the young-fish scenario. Mar Freshw Res 67:1982–1982. https://doi.org/10.1071/MF15228
Andrews AH, Tracey DM, Dunn MR (2009) Lead-radium dating of orange roughy (Hoplostethus atlanticus): validation of a centenarian life span. Can J Fish Aquat Sci 66:1130–1140. https://doi.org/10.1139/F09-059
Andrews AH, Demartini EE, Brodziak J, Nichols RS, Humphreys RL (2012) A long-lived life history for a tropical, deepwater snapper (Pristipomoides filamentosus): bomb radiocarbon and lead-radium dating as extensions of daily increment analyses in otoliths. Can J Fish Aquat Sci 69:1850–1869. https://doi.org/10.1139/f2012-109
Arai T, Ohji M, Hirata T (2007) Trace metal deposition in teleost fish otolith as an environmental indicator. Water Air Soil Pollut 179:255–263. https://doi.org/10.1007/s11270-006-9229-4
Arechavala-Lopez P, Milošević-González M, Sanchez-Jerez P (2016) Using trace elements in otoliths to discriminate between wild and farmed European sea bass (Dicentrarchus labrax L.) and Gilthead sea bream (Sparus aurata L.). Int Aquatic Res 8:263–273. https://doi.org/10.1007/s40071-016-0142-1
Armenta S, de la Guardia M (2016) Analytical approaches for the evaluation of food protected designation of origin. In: Espiñeira M, Santaclara FJ (eds) Advances in food traceability techniques and technologies. Woodhead Publishing, pp 275–301
Artetxe-Arrate I, Fraile I, Clear N, Darnaude AM, Dettman DL, Pécheyran C, Farley J, Murua H (2021) Discrimination of yellowfin tuna Thunnus albacares between nursery areas in the Indian Ocean using otolith chemistry. Mar Ecol Prog Ser 673:165–181. https://doi.org/10.3354/meps13769
Avigliano E (2021) Optimizing the methodological design in fish stock delineation from otolith chemistry: review of spatio-temporal analysis scales. Rev Fish Sci Aquac. https://doi.org/10.1080/23308249.2021.1961679
Avigliano E, Carvalho B, Velasco G, Tripodi P, Volpedo AV (2018) Inter-annual variability in otolith chemistry of catfish Genidens barbus from South-western Atlantic estuaries. J Mar Biol Assoc UK 98:855–865. https://doi.org/10.1017/S0025315417000212
Avigliano E, Pisonero J, Bouchez J, Pouilly M, Domanico A, Sánchez S, Clavijo C, Scarabotti P, Facetti JF, Caffetti JD, del Rosso FR, Llamazares Vegh S, Volpedo AV, Rosso FR, Vegh SL, Alejandra V (2021) Otolith Sr/Ca ratio complements Sr isotopes to reveal fish migration in large basins with heterogeneous geochemical landscapes. Environ Biol Fishes 104:277–292. https://doi.org/10.1007/s10641-021-01074-y
Barbee NC, Swearer SE (2007) Characterizing natal source population signatures in the diadromous fish Galaxias maculatus, using embryonic otolith chemistry. Mar Ecol Prog Ser 343:273–282. https://doi.org/10.3354/meps06886
Barbee NC, Ganio K, Swearer SE (2014) Integrating multiple bioassays to detect and assess impacts of sublethal exposure to metal mixtures in an estuarine fish. Aquat Toxicol 152:244–255. https://doi.org/10.1016/j.aquatox.2014.04.012
Barnett-Johnson R, Grimes CB, Royer CF, Donohoe CJ (2007) Identifying the contribution of wild and hatchery Chinook salmon (Oncorhynchus tshawytscha) to the ocean fishery using otolith microstructure as natural tags. Can J Fish Aquat Sci 64:1683–1692. https://doi.org/10.1139/f07-129
Barnett-Johnson R, Pearson TE, Ramos FC, Grimes CB, MacFarlane RB (2008) Tracking natal origins of salmon using isotopes, otoliths, and landscape geology. Limnol Oceanogr 53:1633–1642. https://doi.org/10.4319/lo.2008.53.4.1633
Barrow JS, Yen JDL, Koehn JD, Zampatti BP, Thiem JD, Tonkin Z, Strawbridge A, Morrongiello JR (2021) Lifetime movement history is associated with variable growth of a potamodromous freshwater fish. J Anim Ecol 90:2560–2572. https://doi.org/10.1111/1365-2656.13561
Beck MW, Heck KL, Able KW, Childers DL, Eggleston DB, Gillanders BM, Halpern B, Hays CG, Hoshino K, Minello TJ, Orth RJ, Sheridan PF, Weinstein MR (2001) The identification, conservation, and management of estuarine and marine nurseries for fish and invertebrates. Bioscience 51:633–641
Bedwal RS, Bahuguna A (1994) Zinc, copper and selenium in reproduction. Cell Mol Life Sci 50:626–640. https://doi.org/10.1007/bf01952862
Bell-Tilcock M, Jeffres CA, Rypel AL, Willmes M, Armstrong RA, Holden P, Moyle PB, Fangue NA, Katz JVE, Sommer TR, Conrad JL, Johnson RC (2021) Biogeochemical processes create distinct isotopic fingerprints to track floodplain rearing of juvenile salmon. PLoS ONE 16:e0257444. https://doi.org/10.1371/journal.pone.0257444
Bendell-Young LI, Harvey HH (1986) Uptake and tissue distribution of manganese in the white sucker (Catostomus commersoni) under conditions of low Ph. Hydrobiologia 133:117–125. https://doi.org/10.1007/BF00031861
Bengtsson B-E, Bengtsson Å, Himberg M (1985) Fish deformities and pollution in some swedish waters. Ambio 14:32–35
Bennion M, Morrison L, Brophy D, Carlsson J, Abrahantes JC, Graham CT (2019) Trace element fingerprinting of blue mussel (Mytilus edulis) shells and soft tissues successfully reveals harvesting locations. Sci Total Environ 685:50–58. https://doi.org/10.1016/j.scitotenv.2019.05.233
Berger W, Killingley J, Vincent E (1978) Sable isotopes in deep-sea carbonates - box core erdc-92, west equatorial pacific. Oceanol Acta 1:203–216
Boecklen WJ, Yarnes CT, Cook BA, James AC (2011) On the use of stable isotopes in trophic ecology. Annu Rev Ecol Evol Syst 42:411–440. https://doi.org/10.1146/annurev-ecolsys-102209-144726
Brennan SR, Schindler DE, Cline TJ, Walsworth TE, Buck G, Fernandez DP (2019a) Shifting habitat mosaics and fish production across river basins. Science 364:783–786. https://doi.org/10.1126/science.aav4313
Brennan SR, Cline TJ, Schindler DE (2019b) Quantifying habitat use of migratory fish across riverscapes using space-time isotope models. Methods Ecol Evol 10:1036–1047. https://doi.org/10.1111/2041-210X.13191
Brophy D, Danilowicz BS, Jeffries TE (2003) The detection of elements in larval otoliths from Atlantic herring using laser ablation ICP-MS. J Fish Biol 63:990–1007. https://doi.org/10.1046/j.1095-8649.2003.00223.x
Brophy D, Rodríguez-Ezpeleta N, Fraile I, Arrizabalaga H (2020) Combining genetic markers with stable isotopes in otoliths reveals complexity in the stock structure of Atlantic bluefin tuna (Thunnus thynnus). Sci Rep 10:14675. https://doi.org/10.1038/s41598-020-71355-6
Brophy D, Pérez-Mayol S, Duncan R, Hüssy K, Geffen AJ, Gerritsen HD, Villanueva MC, Morales-Nin B (2021) Elemental composition of illicia and otoliths and their potential application to age validation in white anglerfish (Lophius piscatorius linnaeus, 1758). Estuar Coast Shelf Sci 261:107557–107557. https://doi.org/10.1016/j.ecss.2021.107557
Brothers EB (1981) What can otolith microstructure tell us about daily and sub-daily events in the early life history of fish? Rapports Et Proces-Verbaux De Reunions Counseil Int Pour L’exploration De La Mer 178:393–394
Brown EJ, Reis-Santos P, Gillanders BM, Støttrup JG (2019) Juvenile fish habitat across the inner Danish waters: using otolith chemistry to discriminate between hybridising con-familials and contiguous, coastal habitat. Estuar Coast Shelf Sci 220:111–119. https://doi.org/10.1016/j.ecss.2019.02.025
Burns NM, Hopkins CR, Bailey DM, Wright PJ (2020) Otolith chemoscape analysis in whiting links fishing grounds to nursery areas. Commun Biol. https://doi.org/10.1038/s42003-020-01433-y
Cadrin SX, Kerr LA, Mariani S (2014) Stock identification methods, 2nd edn. Academic Press, San Diego
Campana SE (1999) Chemistry and composition of fish otoliths: pathways, mechanisms and applications. Mar Ecol Prog Ser 188:263–297. https://doi.org/10.3354/meps188263
Campana SE (2001) Accuracy, precision and quality control in age determination, including a review of the use and abuse of age validation methods. J Fish Biol 59:197–242. https://doi.org/10.1111/j.1095-8649.2001.tb00127.x
Campana SE (2005) Otolith science entering the 21st century. Mar Freshw Res 56:485–495. https://doi.org/10.1071/MF04147
Campana SE, Oxenford HA, Smith JN (1993) Radiochemical determination of longevity in flyingfish Hirundichthys affinis using Th-228/Ra-228. Mar Ecol Prog Ser 100:211–219
Carbajal A, Reyes-López FE, Tallo-Parra O, Lopez-Bejar M, Tort L (2019) Comparative assessment of cortisol in plasma, skin mucus and scales as a measure of the hypothalamic-pituitary-interrenal axis activity in fish. Aquaculture 506:410–416. https://doi.org/10.1016/j.aquaculture.2019.04.005
Carter JF, Chesson LA (2017) Food forensics: stable isotopes as a guide to authenticity and origin. CRC Press, USA
Cavole LM, Demko AM, Diner RE, Giddings A, Koester I, Pagniello CMLS, Paulsen M-L, Ramirez-Valdez A, Schwenck SM, Yen NK, Zill ME, Franks PJS (2016) Biological impacts of the 2013–2015 warm-water anomaly in the Northeast Pacific winners, losers, and the future. Oceanography 29:273–285
Chapman BB, Skov C, Hulthén K, Brodersen J, Nilsson PA, Hansson LA, Brönmark C (2012a) Partial migration in fishes: definitions, methodologies and taxonomic distribution. J Fish Biol 81:479–499. https://doi.org/10.1111/j.1095-8649.2012.03349.x
Chapman BB, Hulthén K, Brodersen J, Nilsson PA, Skov C, Hansson LA, Brönmark C (2012b) Partial migration in fishes: causes and consequences. J Fish Biol 81:456–478. https://doi.org/10.1111/j.1095-8649.2012.03342.x
Charapata P, Oxman D, McNeel K, Keith A, Mansouri F, Trumble S (2022) Lifetime hormone profiles for a long-lived teleost: Opercula reveal novel estimates of age-specific reproductive parameters and stress trends in yelloweye rockfish (Sebastes ruberrimus). Canadian J Fish Aquatic Sci. https://doi.org/10.1139/cjfas-2022-0048
Cheng L-C, Shiao J-C, Hsiao S-Y, Wang P-L (2018) Fractionation of otolith nitrogen stable isotopes measured by peroxodisulphate oxidation-bacterial conversion and isotopic ratio mass spectrometry. Rapid Commun Mass Spectrom. https://doi.org/10.1002/rcm.8259
Chikaraishi Y, Ogawa NO, Kashiyama Y, Takano Y, Suga H, Tomitani A, Miyashita H, Kitazato H, Ohkouchi N (2009) Determination of aquatic food-web structure based on compound-specific nitrogen isotopic composition of amino acids. Limnol Oceanogr Methods 7:740–750
Chittaro PM, Finley RJ, Levin PS (2009) Spatial and temporal patterns in the contribution of fish from their nursery habitats. Oecologia 160:49–61. https://doi.org/10.1007/s00442-009-1282-4
Chrousos GP, Gold PW (1992) The concepts of stress and stress system disorders. Overview of physical and behavioral homeostasis. JAMA 267:1244–1252. https://doi.org/10.1001/jama.1992.03480090092034
Chung M-T, Trueman CN, Godiksen JA, Grønkjær P (2019a) Otolith δ13C values as a metabolic proxy: approaches and mechanical underpinnings. Mar Freshw Res 70:1747–1756. https://doi.org/10.1071/MF18317
Chung M-T, Trueman CN, Godiksen JA, Holmstrup ME, Grønkjær P (2019b) Field metabolic rates of teleost fishes are recorded in otolith carbonate. Commun Biol 2:24. https://doi.org/10.1038/s42003-018-0266-5
Chung M-T, Chen C-Y, Shiao J-C, Shirai K, Wang C-H (2021a) Metabolic proxy for cephalopods: stable carbon isotope values recorded in different biogenic carbonates. Methods Ecol Evol 12:1648–1657. https://doi.org/10.1111/2041-210X.13630
Chung M-T, Jørgensen K-EM, Trueman CN, Knutsen H, Jorde PE, Grønkjær P (2021b) First measurements of field metabolic rate in wild juvenile fishes show strong thermal sensitivity but variations between sympatric ecotypes. Oikos 130:287–299. https://doi.org/10.1111/oik.07647
Clark CT, Horstmann L, Misarti N (2020) Zinc concentrations in teeth of female walruses reflect the onset of reproductive maturity. Conserv Physiol. https://doi.org/10.1093/conphys/coaa029
Collette BB, Carpenter KE, Polidoro BA, Juan-Jorda MJ, Boustany A, Die DJ, Elfes C, Fox W, Graves J, Harrison LR, McManus R, Minte-Vera CV, Nelson R, Restrepo V, Schratwieser J, Sun CL, Amorim A, Brick Peres M, Canales C, Cardenas G, Chang SK, Chiang WC, de Oliveira LN, Jr., Harwell H, Lessa R, Fredou FL, Oxenford HA, Serra R, Shao KT, Sumaila R, Wang SP, Watson R, Yanez E, (2011) Conservation. high value and long life–double jeopardy for tunas and billfishes. Science 333:291–292. https://doi.org/10.1126/science.1208730
Cooke SJ, Paukert C, Hogan Z (2012) Endangered river fish: factors hindering conservation and restoration. Endangered Spec Res 17:179–191. https://doi.org/10.3354/esr00426
Cordoleani F, Phillis CC, Sturrock AM, FitzGerald AM, Malkassian A, Whitman GE, Weber PK, Johnson RC (2021) Threatened salmon rely on a rare life history strategy in a warming landscape. Nat Clim Chang 11:982–988. https://doi.org/10.1038/s41558-021-01186-4
Cottingham A, Hall NG, Loneragan NR, Jenkins GI, Potter IC (2020) Efficacy of restocking an estuarine-resident species demonstrated by long-term monitoring of cultured fish with alizarin complexone-stained otoliths. A Case Study Fish Res 227:105556. https://doi.org/10.1016/j.fishres.2020.105556
Cowen RK, Sponaugle S (2009) Larval dispersal and marine population connectivity. Ann Rev Mar Sci 1:443–466. https://doi.org/10.1146/annurev.marine.010908.163757
Crook DA, O’Mahony D, Gillanders BM, Munro AR, Sanger AC (2007) Production of external fluorescent marks on golden perch fingerlings through osmotic induction marking with alizarin red S. N Am J Fish Manage 27:670–675
Crook DA, Lacksen K, King AJ, Buckle DJ, Tickell S, Woodhead JD, Maas R, Townsend SA, Douglas MM (2016) Temporal and spatial variation in strontium in a tropical river: implications for otolith chemistry analyses of fish migration. Canadian J Fish Aquatic Sci. https://doi.org/10.1139/cjfas-2016-0153
Crook DA, Munro AR, Gillanders BM, Sanger AC, Thurstan S, Macdonald J (2005) Review of existing and proposed methodologies for discriminating hatchery and wild-bred fish (Native fish strategy project R5003). Murray-Darling Basin Commission (Canberra, Australia)
Crook DA, O'Mahony DJ, Gillanders BM, Munro AR, Sanger AC. (2011). Quantitative measurement of calcein fluorescence for non-lethal, field based discrimination of hatchery and wild fish. In: J R McKenzie, B Parsons, A C Seitz, R K Kopf, M G Mesa & Q Phelps (Eds.), Advances in fish tagging and marking technology (Vol. 76, pp. 389–396).
Curtis JM, Stunz GW, Overath RD, Vega RR (2014) Otolith chemistry can discriminate signatures of hatchery-reared and wild spotted seatrout. Fish Res 153:31–40. https://doi.org/10.1016/j.fishres.2013.12.013
Cusa M, St John Glew K, Trueman C, Mariani S, Buckley L, Neat F, Longo C (2021) A future for seafood point-of-origin testing using DNA and stable isotope signatures. Rev Fish Biol Fish. https://doi.org/10.1007/s11160-021-09680-w
Darnaude AM, Sturrock A, Trueman CN, Mouillot D, Eimf CSE, Hunter E (2014) Listening In on the past: what can otolith δ18O values really tell us about the environmental history of fishes? PLoS ONE 9:e108539. https://doi.org/10.1371/journal.pone.0108539
Daryanani DS, Martino JC, Doubleday ZA (2021) Statolith chemistry: a new tool to understand the ecology and provenance of octopus. Rev Fish Biol Fisheries. https://doi.org/10.1007/s11160-021-09671-x
Das S, Judd EJ, Uveges BT, Ivany LC, Junium CK (2021) Variation in δ15N from shell-associated organic matter in bivalves: Implications for studies of modern and fossil ecosystems. Palaeogeogr Palaeoclimatol Palaeoecol 562:110076. https://doi.org/10.1016/j.palaeo.2020.110076
Dauphin Y, Dufour E (2003) Composition and properties of the soluble organic matrix of the otolith of a marine fish: Gadus morhua Linne, 1758 (Teleostei, Gadidae). Comp Biochem Physiol a: Mol Integr Physiol 134:551–561. https://doi.org/10.1016/S1095-6433(02)00358-6
Davias LA, Kornis MS, Breitburg DL (2014) Environmental factors influencing δ 13C and δ 15N in three Chesapeake Bay fishes. ICES J Mar Sci 71:689–702
Denechaud C, Geffen AJ, Smoliński S, Godiksen JA (2021) Otolith “spawning zones” across multiple Atlantic cod populations: do they accurately record maturity and spawning? PLoS ONE 16:e0257218. https://doi.org/10.1371/journal.pone.0257218
Des Roches S, Pendleton LH, Shapiro B, Palkovacs EP (2021) Conserving intraspecific variation for nature’s contributions to people. Nat Ecol Evolution. https://doi.org/10.1038/s41559-021-01403-5
Dove SG, Kingsford MJ (1998) Use of otoliths and eye lenses for measuring trace-metal incorporation in fishes: a biogeographic study. Mar Biol 130:377–387. https://doi.org/10.1007/s002270050258
Dua A, Gupta N (2005) Mercury toxicology as assessed through fish scales. Bull Environ Contam Toxicol 74:1105–1110. https://doi.org/10.1007/s00128-005-0695-y
Dufour E, Gerdeaux D, Wurster CM (2007) Whitefish (Coregonus lavaretus) respiration rate governs intra-otolith variation of δ13C values in Lake Annecy. Can J Fish Aquat Sci 64:1736–1746. https://doi.org/10.1139/f07-132
Duponchelle F, Pouilly M, Pécheyran C, Hauser M, Renno J-F, Panfili J, Darnaude AM, García-Vasquez A, Carvajal-Vallejos F, García-Dávila C, Doria C, Bérail S, Donard A, Sondag F, Santos RV, Nuñez J, Point D, Labonne M, Baras E (2016) Trans-amazonian natal homing in giant catfish. J Appl Ecol 53:1511–1520. https://doi.org/10.1111/1365-2664.12665
Eakin MC, Grottoli AG. (2006). Coral reef records of past climatic change. In: coral reefs and climate change: science and management (pp. 33–54).
Eddy TD, Bernhardt JR, Blanchard JL, Cheung WWL, Colléter M, du Pontavice H, Fulton EA, Gascuel D, Kearney KA, Petrik CM, Roy T, Rykaczewski RR, Selden R, Stock CA, Wabnitz CCC, Watson RA (2021) Energy flow through marine ecosystems: confronting transfer efficiency. Trends Ecol Evol 36:76–86. https://doi.org/10.1016/j.tree.2020.09.006
Elsdon TS, Wells BK, Campana SE, Gillanders BM, Jones CM, Limburg KE, Secor DH, Thorrold SR, Walther BD (2008) Otolith chemistry to describe movements and life-history parameters of fishes: Hypotheses, assumptions, limitations and inferences. Oceanogr Mar Biol Annu Rev 46:297–330
Engelhard GH, Heino M (2005) Scale analysis suggests frequent skipping of the second reproductive season in Atlantic herring. Biol Let 1:172–175
Ennevor BC (1994) Mass marking coho salmon, Oncorhynchus kisutch, fry with lanthanum and cerium. Fish B-Noaa 92:471–473
Fletcher PE, Fletcher GL (1980) Zinc- and copper-binding proteins in the plasma of winter flounder (Pseudopleuronectes americanus). Can J Zool 58:609–613. https://doi.org/10.1139/z80-086
Fletcher GL, King MJ (1978) Seasonal dynamics of Cu2+, Zn2+, Ca2+, and Mg2+ in gonads and liver of winter flounder (Pseudopleuronectes americanus): evidence for summer storage of Zn2+ for winter gonad development in females. Can J Zool 56:284–290. https://doi.org/10.1139/z78-038
Fonseca VF, Duarte IA, Matos AR, Reis-Santos P, Duarte B (2022) Fatty acid profiles as natural tracers of provenance and lipid quality indicators in illegally sourced fish and bivalves. Food Control 134:108735. https://doi.org/10.1016/j.foodcont.2021.108735
Fox M, Mitchell M, Dean M, Elliott C, Campbell K (2018) The seafood supply chain from a fraudulent perspective. Food Sec 10:939–963. https://doi.org/10.1007/s12571-018-0826-z
Fraile I, Arrizabalaga H, Santiago J, Goñi N, Arregi I, Madinabeitia S, Wells RJD, Rooker JR (2016) Otolith chemistry as an indicator of movements of albacore (Thunnus alalunga) in the North Atlantic Ocean. Mar Freshw Res 67:1002. https://doi.org/10.1071/mf15097
Friedrich LA, Halden NM (2010) Determining exposure history of northern pike and walleye to tailings effluence using trace metal uptake in otoliths. Environ Sci Technol 44:1551–1558. https://doi.org/10.1021/es903261q
Fry B (2002) Conservative mixing of stable isotopes across estuarine salinity gradients: a conceptual framework for monitoring watershed influences on downstream fisheries production. Estuaries 25:264–271
Garrido S, Ben-Hamadou R, Santos AMP, Ferreira S, Teodósio MA, Cotano U, Irigoien X, Peck MA, Saiz E, Ré P (2015) Born small, die young: Intrinsic, size-selective mortality in marine larval fish. Sci Rep 5:17065. https://doi.org/10.1038/srep17065
Gillanders BM (2002a) Temporal and spatial variability in elemental composition of otoliths: implications for determining stock identity and connectivity of populations. Can J Fish Aquat Sci 59:669–679. https://doi.org/10.1139/f02-040
Gillanders BM (2002b) Connectivity between juvenile and adult fish populations: do adults remain near their recruitment estuaries? Mar Ecol Prog Ser 240:215–223
Gillanders BM, Joyce TC (2005) Distinguishing aquaculture and wild yellowtail kingfish via natural elemental signatures in otoliths. Mar Freshw Res 56:693–704. https://doi.org/10.1071/MF04133
Gillanders BM, Able KW, Brown JA, Eggleston DB, Sheridan PF (2003) Evidence of connectivity between juvenile and adult habitats for mobile marine fauna: an important component of nurseries. Mar Ecol Prog Ser 247:281–295
Gillanders BM, Izzo C, Doubleday ZA, Ye QF (2015) Partial migration: growth varies between resident and migratory fish. Biol Let 11:20140850. https://doi.org/10.1098/rsbl.2014.0850
Godbout L, Trudel M, Irvine JR, Wood CC, Grove MJ, Schmitt AK, McKeegan KD (2010) Sulfur isotopes in otoliths allow discrimination of anadromous and non-anadromous ecotypes of sockeye salmon (Oncorhynchus nerka). Environ Biol Fishes 89:521–532. https://doi.org/10.1007/s10641-010-9689-7
Godiksen JA, Chung M-T, Grønkjær P (2019) Effects of temperature on tissue – diet isotopic spacing of nitrogen and carbon in otolith organic matter. Marine Freshwater Res 70(12):1757–1767
Goikoetxea A, Sadoul B, Blondeau-Bidet E, Aerts J, Blanc M-O, Parrinello H, Barrachina C, Pratlong M, Geffroy B (2021) Genetic pathways underpinning hormonal stress responses in fish exposed to short- and long-term warm ocean temperatures. Ecol Ind 120:106937. https://doi.org/10.1016/j.ecolind.2020.106937
Gopi K, Mazumder D, Sammut J, Saintilan N (2019a) Determining the provenance and authenticity of seafood: a review of current methodologies. Trends Food Sci Technol 91:294–304. https://doi.org/10.1016/j.tifs.2019.07.010
Gopi K, Mazumder D, Sammut J, Saintilan N, Crawford J, Gadd P (2019b) Combined use of stable isotope analysis and elemental profiling to determine provenance of black tiger prawns (Penaeus monodon). Food Control 95:242–248. https://doi.org/10.1016/j.foodcont.2018.08.012
Grønkjær P, Pedersen JB, Ankjærø TT, Kjeldsen H, Heinemeier J, Steingrund P, Nielsen JM, Christensen JT (2013) Stable N and C isotopes in the organic matrix of fish otoliths: validation of a new approach for studying spatial and temporal changes in the trophic structure of aquatic ecosystems. Can J Fish Aquat Sci 70:143–146. https://doi.org/10.1139/cjfas-2012-0386
Grüss A, Kaplan DM, Hart DR (2011) Relative impacts of adult movement, larval dispersal and harvester movement on the effectiveness of reserve networks. PLoS ONE 6:e19960–e19960. https://doi.org/10.1371/journal.pone.0019960
Guédron S, Acha D (2021) Mercury and methylmercury contamination of terrestrial and aquatic ecosystems. Appl Sci 11:4807
Halden NM, Friedrich LA (2008) Trace-element distributions in fish otoliths: natural markers of life histories, environmental conditions and exposure to tailings effluence. Mineral Mag 72:593–605. https://doi.org/10.1180/minmag.2008.072.2.593
Halden NM, Mejia SR, Babaluk JA, Reist JD, Kristofferson AH, Campbell JL, Teesdale WJ (2000) Oscillatory zinc distribution in Arctic char (Salvelinus alpinus) otoliths: the result of biology or environment? Fish Res 46:289–298. https://doi.org/10.1016/S0165-7836(00)00154-5
Hall CJ, Jordaan A, Frisk MG (2011) The historic influence of dams on diadromous fish habitat with a focus on river herring and hydrologic longitudinal connectivity. Landscape Ecol 26:95–107. https://doi.org/10.1007/s10980-010-9539-1
Hanke I, Ampe B, Kunzmann A, Gärdes A, Aerts J (2019) Thermal stress response of juvenile milkfish (Chanos chanos) quantified by ontogenetic and regenerated scale cortisol. Aquaculture 500:24–30. https://doi.org/10.1016/j.aquaculture.2018.09.016
Hanson NN, Wurster CM, EIMF, Todd CD, (2010) Comparison of secondary ion mass spectrometry and micromilling/continuous flow isotope ratio mass spectrometry techniques used to acquire intra-otolith δ18O values of wild Atlantic salmon (Salmo salar). Rapid Commun Mass Spectrom 24:2491–2498. https://doi.org/10.1002/rcm.4646
Hauser M, Duponchelle F, Hermann TW, Limburg KE, Castello L, Stewart DJ, Torrente-Vilara G, García-Vásquez A, García-Davila C, Pouilly M, Pecheyran C, Ponzevera E, Renno J-f, Moret AS, Doria CRC (2020) Unmasking continental natal homing in goliath catfish from the upper Amazon. Freshw Biol 65:325–336. https://doi.org/10.1111/fwb.13427
Hegg JC, Kennedy BP (2021) Let’s do the time warp again: non-linear time series matching as a tool for sequentially structured data in ecology. Ecosphere 12:e03742. https://doi.org/10.1002/ecs2.3742
Hegg JC, Kennedy BP, Fremier AK (2013) Predicting strontium isotope variation and fish location with bedrock geology: Understanding the effects of geologic heterogeneity. Chem Geol 360–361:89–98. https://doi.org/10.1016/j.chemgeo.2013.10.010
Hegg JC, Giarrizzo T, Kennedy BP (2015) Diverse early life-history strategies in migratory amazonian catfish: implications for conservation and management. PLoS ONE 10:e0129697–e0129697. https://doi.org/10.1371/journal.pone.0129697
Heimbrand Y, Limburg KE, Hüssy K, Casini M, Sjöberg R, Palmén Bratt AM, Levinsky SE, Karpushevskaia A, Radtke K, Öhlund J (2020) Seeking the true time: exploring otolith chemistry as an age-determination tool. J Fish Biol. https://doi.org/10.1111/jfb.14422
Hermann TW, Stewart DJ, Limburg KE, Castello L (2016) Unravelling the life history of Amazonian fishes through otolith microchemistry. Royal Soc Open Sci. https://doi.org/10.1098/rsos.160206
Hermann TW, Duponchelle F, Castello L, Limburg KE, Pereira LA, Hauser M (2021) Harnessing the potential for otolith microchemistry to foster the conservation of Amazonian fishes. Aquat Conserv Mar Freshwat Ecosyst 31:1206–1220. https://doi.org/10.1002/aqc.3567
Heymans JJ, Coll M, Link JS, Mackinson S, Steenbeek J, Walters C, Christensen V (2016) Best practice in ecopath with ecosim food-web models for ecosystem-based management. Ecol Model 331:173–184. https://doi.org/10.1016/j.ecolmodel.2015.12.007
Hobbs JA, Lewis LS, Willmes M, Denney C, Bush E (2019) Complex life histories discovered in a critically endangered fish. Sci Rep 9:16772–16772. https://doi.org/10.1038/s41598-019-52273-8
Hodge BW, Wilzbach MA, Duffy WG, Quiñones RM, Hobbs JA (2016) Life history diversity in Klamath river steelhead. Trans Am Fish Soc 145:227–238. https://doi.org/10.1080/00028487.2015.1111257
Høie H, Folkvord A (2006) Estimating the timing of growth rings in Atlantic cod otoliths using stable oxygen isotopes. J Fish Biol 68:826–837. https://doi.org/10.1111/j.0022-1112.2006.00957.x
Høie H, Otterlei E, Folkvord A (2004a) Temperature-dependent fractionation of stable oxygen isotopes in otoliths of juvenile cod (Gadus morhua L.). ICES J Mar Sci 61:243–251. https://doi.org/10.1016/j.icesjms.2003.11.006
Høie H, Andersson C, Folkvord A, Karlsen Ø (2004b) Precision and accuracy of stable isotope signals in otoliths of pen-reared cod (Gadus morhua) when sampled with a high-resolution micromill. Mar Biol 144:1039–1049
Humston R, Doss SS, Wass C, Hollenbeck C, Thorrold SR, Smith S, Bataille CP (2017) Isotope geochemistry reveals ontogeny of dispersal and exchange between main-river and tributary habitats in smallmouth bass Micropterus dolomieu. J Fish Biol 90:528–548. https://doi.org/10.1111/jfb.13073
Hunt KE, Stimmelmayr R, George C, Hanns C, Suydam R, Brower H Jr, Rolland RM (2014) Baleen hormones: a novel tool for retrospective assessment of stress and reproduction in bowhead whales (Balaena mysticetus). Conserv Physiol 2:cou030. https://doi.org/10.1093/conphys/cou030
Hüssy K, Mosegaard H, Jessen F (2004) Effect of age and temperature on amino acid composition and the content of different protein types of juvenile Atlantic cod (Gadus morhua) otoliths. Can J Fish Aquat Sci 61:1012–1020. https://doi.org/10.1139/f04-037
Hüssy K, Radtke K, Plikshs M, Oeberst R, Baranova T, Krumme U, Sjöberg R, Walther Y, Mosegaard H (2016a) Challenging ICES age estimation protocols: lessons learned from the eastern Baltic cod stock. ICES J Mar Sci 73:2138–2149. https://doi.org/10.1093/icesjms/fsw107
Hüssy K, Gröger J, Heidemann F, Hinrichsen HH, Marohn L (2016b) Slave to the rhythm: seasonal signals in otolith microchemistry reveal age of eastern Baltic cod (Gadus morhua). ICES J Mar Sci 73:1019–1032. https://doi.org/10.1093/icesjms/fsv247
Hüssy K, Krüger-Johnsen M, Thomsen TB, Heredia BD, Næraa T, Limburg KE, Heimbrand Y, McQueen K, Haase S, Krumme U, Casini M, Mion M, Radtke K (2021a) It’s elemental, my dear watson: Validating seasonal patterns in otolith chemical chronologies. Can J Fish Aquat Sci 78:551–566. https://doi.org/10.1139/cjfas-2020-0388
Hüssy K, Limburg KE, de Pontual H, Thomas ORB, Cook PK, Heimbrand Y, Blass M, Sturrock AM (2021b) Trace element patterns in otoliths: the role of biomineralization. Rev Fish Sci Aquac 29:445–477. https://doi.org/10.1080/23308249.2020.1760204
Hutchings JA, Baum JK (2005) Measuring marine fish biodiversity: temporal changes in abundance, life history and demography. Philosoph Trans Royal Soc B: Biol Sci 360:315–338. https://doi.org/10.1098/rstb.2004.1586
Irgens C, Folkvord A, Otterå H, Kjesbu OS (2019) Otolith growth and zone formation during first maturity and spawning of Atlantic cod (Gadus morhua). Can J Fish Aquat Sci 77:113–123. https://doi.org/10.1139/cjfas-2019-0033
Izzo C, Doubleday ZA, Grammer GL, Gilmore KL, Alleway HK, Barnes TC, Disspain MCF, Giraldo AJ, Mazloumi N, Gillanders BM (2016a) Fish as proxies of ecological and environmental change. Rev Fish Biol Fish 26:265–286. https://doi.org/10.1007/s11160-016-9424-3
Izzo C, Doubleday ZA, Gillanders BM (2016b) Where do elements bind within the otoliths of fish? Mar Freshw Res 67:1072–1076. https://doi.org/10.1071/MF15064
Izzo C, Reis-Santos P, Gillanders BM (2018) Otolith chemistry does not just reflect environmental conditions: a meta-analytic evaluation. Fish Fish 19:441–454. https://doi.org/10.1111/faf.12264
Jarman KH, Daly DS, Anderson KK, Wahl KL (2003) A new approach to automated peak detection. Chemom Intell Lab Syst 69:61–76. https://doi.org/10.1016/S0169-7439(03)00113-8
Johnson RC, Weber PK, Wikert JD, Workman ML, MacFarlane RB, Grove MJ, Schmitt AK (2012) Managed metapopulations: do salmon hatchery ‘sources’ lead to in-river ‘sinks’ in conservation? PLoS ONE 7:e28880. https://doi.org/10.1371/journal.pone.0028880
Johnson RC, Windell S, Brandes PL, Conrad JL, Ferguson J, Goertler PAL, Harvey BN, Heublein J, Isreal JA, Kratville DW, Kirsch JE, Perry RW, Pisciotto J, Poytress WR, Reece K, Swart BG (2017) Science advancements key to increasing management value of life stage monitoring networks for endangered Sacramento River winter-run Chinook salmon in California. San Francisco Estuary Watershed Sci 15:1–41
Johnson RC, Stewart AR, Limburg KE, Huang R, Cocherell D, Feyrer F (2020) Lifetime chronicles of selenium exposure linked to deformities in an imperiled migratory fish. Environ Sci Technol 54:2892–2901. https://doi.org/10.1021/acs.est.9b06419
Kalish JM (1991a) 13C and 18O isotopic disequilibria in fish otoliths: metabolic and kinetic effects. Mar Ecol Prog Ser 75:191–203
Kalish JM (1991b) Determinants of otolith chemistry: seasonal variation in the composition of blood plasma, endolymph and otoliths of bearded rock cod Pseudophysis barbatus. Mar Ecol Prog Ser 74:137–159
Kalish JM (1991c) Oxygen and carbon stable isotopes in the otoliths of wild and laboratory-reared Australian salmon (Arripis trutta). Mar Biol 110:37–47
Kalish JM (1993) Pre- and post-bomb radiocarbon in fish otoliths. Earth Planet Sci Lett 114:549–554
Kastelle CR, Helser TE, McKay JL, Johnston CG, Anderl DM, Matta ME, Nichol DG (2017) Age validation of Pacific cod (Gadus macrocephalus) using high-resolution stable oxygen isotope (δ18O) chronologies in otoliths. Fish Res 185:43–53. https://doi.org/10.1016/j.fishres.2016.09.024
Kay SM (1998) Fundamentals of statistical signal processing | Guide books. Upper Saddle River, NJ: Prentice-Hall PTR.
Kerr LA, Hintzen NT, Cadrin SX, Clausen LW, Dickey-Collas M, Goethel DR, Hatfield EMC, Kritzer JP, Nash RDM (2016) Lessons learned from practical approaches to reconcile mismatches between biological population structure and stock units of marine fish. ICES J Marine Sci. https://doi.org/10.1093/icesjms/fsw188
Kitada S (2020) Lessons from Japan marine stock enhancement and sea ranching programmes over 100 years. Rev Aquac 12:1944–1969. https://doi.org/10.1111/raq.12418
Kroetz K, Luque GM, Gephart JA, Jardine SL, Lee P, Chicojay Moore K, Cole C, Steinkruger A, Donlan CJ (2020) Consequences of seafood mislabeling for marine populations and fisheries management. Proc Natl Acad Sci 117:30318–30323. https://doi.org/10.1073/pnas.2003741117
Laberge F, Yin-Liao I, Bernier NJ (2019) Temporal profiles of cortisol accumulation and clearance support scale cortisol content as an indicator of chronic stress in fish. Conserv Physiol. https://doi.org/10.1093/conphys/coz052
Lackmann AR, Andrews AH, Butler MG, Bielak-Lackmann ES, Clark ME (2019) Bigmouth Buffalo Ictiobus cyprinellus sets freshwater teleost record as improved age analysis reveals centenarian longevity. Commun Biol 2(1):1–14
Lazartigues A, Girard C, Brodeur P, Lecomte F, Mingelbier M, Sirois P (2018) Otolith microchemistry to identify sources of larval yellow perch in a fluvial lake: an approach towards freshwater fish management. Can J Fish Aquat Sci 75:474–487. https://doi.org/10.1139/cjfas-2016-0289
Leal MC, Pimentel T, Ricardo F, Rosa R, Calado R (2015) Seafood traceability: current needs, available tools, and biotechnological challenges for origin certification. Trends Biotechnol 33:331–336. https://doi.org/10.1016/j.tibtech.2015.03.003
Lechner A, Keckeis H, Humphries P (2016) Patterns and processes in the drift of early developmental stages of fish in rivers: a review. Rev Fish Biol Fish 26:471–489. https://doi.org/10.1007/s11160-016-9437-y
Lenz C, Jilbert T, Conley DJ, Slomp CP (2015) Hypoxia-driven variations in iron and manganese shuttling in the Baltic sea over the past 8 kyr. Geochem Geophys Geosyst 16:3754–3766. https://doi.org/10.1002/2015GC005960
Limburg KE, Casini M (2018) Effect of marine hypoxia on Baltic Sea Cod Gadus morhua: evidence from otolith chemical proxies. Front Mar Sci. https://doi.org/10.3389/fmars.2018.00482
Limburg KE, Casini M (2019) Otolith chemistry indicates recent worsened Baltic cod condition is linked to hypoxia exposure. Biol Let 15:20190352. https://doi.org/10.1098/rsbl.2019.0352
Limburg KE, Elfman M (2010) Patterns and magnitude of Zn: Ca in otoliths support the recent phylogenetic typology of Salmoniformes and their sister groups. Can J Fish Aquat Sci 67:597–604
Limburg KE, Elfman M (2017) Insights from two-dimensional mapping of otolith chemistry. J Fish Biol 90:480–491. https://doi.org/10.1111/jfb.13048
Limburg KE, Lochet A, Driscoll D, Dale DS, Huang R (2010) Selenium detected in fish otoliths: a novel tracer for a polluted lake? Environ Biol Fishes 89:433–440. https://doi.org/10.1007/s10641-010-9671-4
Limburg KE, Olson C, Walther Y, Dale D, Slomp CP, Høie H (2011) Tracking Baltic hypoxia and cod migration over millennia with natural tags. Proc Natl Acad Sci 108:E177–E182. https://doi.org/10.1073/pnas.1100684108
Limburg KE, Walther BD, Lu Z, Jackman G, Mohan J, Walther Y, Nissling A, Weber PK, Schmitt AK (2015) In search of the dead zone: Use of otoliths for tracking fish exposure to hypoxia. J Mar Syst 141:167–178. https://doi.org/10.1016/j.jmarsys.2014.02.014
Limburg KE, Wuenschel MJ, Hüssy K, Heimbrand Y, Samson M (2018) Making the otolith magnesium chemical calendar-clock tick: plausible mechanism and empirical evidence. Rev Fish Sci Aquac 26:479–493. https://doi.org/10.1080/23308249.2018.1458817
Lomb NR (1976) Least-squares frequency analysis of unequally spaced data. Astrophys Space Sci 39:447–462. https://doi.org/10.1007/BF00648343
Lu Z, Jenkyns HC, Rickaby REM (2010) Iodine to calcium ratios in marine carbonate as a paleo-redox proxy during oceanic anoxic events. Geology 38:1107–1110. https://doi.org/10.1130/g31145.1
Lueders-Dumont JA, Wang XT, Jensen OP, Sigman DM, Ward BB (2018) Nitrogen isotopic analysis of carbonate-bound organic matter in modern and fossil fish otoliths. Geochim Cosmochim Acta 224:200–222. https://doi.org/10.1016/j.gca.2018.01.001
Lueders-Dumont JA, Sigman DM, Johnson BJ, Jensen OP, Oleynik S, Ward BB (2020) Comparison of the isotopic composition of fish otolith-bound organic N with host tissue. Can J Fish Aquat Sci 77:264–275
Lueders-Dumont JA, Forden AG, Kast ER, Mohan JA, Walther BD, Sigman DM, Ward BB (2022) Controls on the nitrogen isotopic composition of fish otolith organic matter: lessons from a controlled diet switch experiment. Geochim Cosmochim Acta 316:69–86. https://doi.org/10.1016/j.gca.2021.09.030
Macdonald JI, Crook DA (2010) Variability in Sr: Ca and Ba: Ca ratios in water and fish otoliths across an estuarine salinity gradient. Mar Ecol Prog Ser 413:147–161. https://doi.org/10.3354/meps08703
Macdonald JI, Drysdale RN, Witt R, Cságoly Z, Marteinsdóttir G (2020) Isolating the influence of ontogeny helps predict island-wide variability in fish otolith chemistry. Rev Fish Biol Fisheries 30:173–202. https://doi.org/10.1007/s11160-019-09591-x
Mamede R, Ricardo F, Gonçalves D, Ferreira da Silva E, Patinha C, Calado R (2021) Assessing the use of surrogate species for a more cost-effective traceability of geographic origin using elemental fingerprints of bivalve shells. Ecol Ind 130:108065. https://doi.org/10.1016/j.ecolind.2021.108065
Martino JC, Fowler AJ, Doubleday ZA, Grammer GL, Gillanders BM (2019) Using otolith chronologies to understand long-term trends and extrinsic drivers of growth in fisheries. Ecosphere 10:e02553. https://doi.org/10.1002/ecs2.2553
Martino JC, Mazumder D, Gadd P, Doubleday ZA (2021) Tracking the provenance of octopus using isotopic and multi-elemental analysis. Food Chem. https://doi.org/10.1016/j.foodchem.2021.131133
Masuda R, Tsukamoto K (1998) Stock Enhancement in Japan: review and perspective. Bull Mar Sci 62:337–358
Matsubayashi J, Osada Y, Tadokoro K, Abe Y, Yamaguchi A, Shirai K, Honda K, Yoshikawa C, Ogawa NO, Ohkouchi N, Ishikawa NF, Nagata T, Miyamoto H, Nishino S, Tayasu I (2020) Tracking long-distance migration of marine fishes using compound-specific stable isotope analysis of amino acids. Ecol Lett 23:881–890. https://doi.org/10.1111/ele.13496
Matta EM, Black BA, Wilderbuer TK (2010) Climate-driven synchrony in otolith growth-increment chronologies for three Bering Sea flatfish species. Mar Ecol Prog Ser 413:137–145
McClelland JW, Montoya JP (2002) Trophic relationships and the nitrogen isotopic composition of amino acids in plankton. Ecology 83:2173–2180
McConnaughey TA, Burdett J, Whelan JF, Paull CK (1997) Carbon isotopes in biological carbonates: respiration and photosynthesis. Geochim Cosmochim Acta 61:611–622. https://doi.org/10.1016/S0016-7037(96)00361-4
McGowan N, Fowler AM, Parkinson K, Bishop DP, Ganio K, Doble PA, Booth DJ, Hare DJ (2014) Beyond the transect: an alternative microchemical imaging method for fine scale analysis of trace elements in fish otoliths during early life. Sci Total Environ 494–495:177–186. https://doi.org/10.1016/j.scitotenv.2014.05.115
McMahon KW, Berumen ML, Mateo I, Elsdon TS, Thorrold SR (2011a) Carbon isotopes in otolith amino acids identify residency of juvenile snapper (Family: Lutjanidae) in coastal nurseries. Coral Reefs 30:1135–1145. https://doi.org/10.1007/s00338-011-0816-5
McMahon KW, Fogel ML, Johnson BJ, Houghton LA, Thorrold SR, Gillanders B (2011b) A new method to reconstruct fish diet and movement patterns from δ13C values in otolith amino acids. Can J Fish Aquat Sci 68:1330–1340. https://doi.org/10.1139/f2011-070
McMahon KW, Hamady LL, Thorrold SR (2013) A review of ecogeochemistry approaches to estimating movements of marine animals. Limnol Oceanogr 58:697–714. https://doi.org/10.4319/lo.2013.58.2.0697
Miller MB, Clough AM, Batson JN, Vachet RW (2006) Transition metal binding to cod otolith proteins. J Exp Mar Biol Ecol 329:135–143. https://doi.org/10.1016/J.JEMBE.2005.08.016
Mohan JA, Walther B (2016) Out of breath and hungry: natural tags reveal trophic resilience of Atlantic croaker to hypoxia exposure. Mar Ecol Prog Ser 560:207–221
Mohan JA, Rulifson RA, Corbett DR, Halden NM (2012) Validation of oligohaline elemental otolith signatures of striped bass by use of in situ caging experiments and water chemistry. Marine and Coastal Fisheries 4:57–70. https://doi.org/10.1080/19425120.2012.656533
Momigliano P, Jokinen H, Calboli F, Aro E, Merilä J (2019) Cryptic temporal changes in stock composition explain the decline of a flounder (Platichthys spp.) assemblage. Evol Appl 12:549–559. https://doi.org/10.1111/eva.12738
Moore JW, Yeakel JD, Peard D, Lough J, Beere M (2014) Life-history diversity and its importance to population stability and persistence of a migratory fish: Steelhead in two large North American watersheds. J Anim Ecol 83:1035–1046. https://doi.org/10.1111/1365-2656.12212
Morrison L, Bennion M, Gill S, Graham CT (2019) Spatio-temporal trace element fingerprinting of king scallops (Pecten maximus) reveals harvesting period and location. Sci Total Environ 697:134121. https://doi.org/10.1016/j.scitotenv.2019.134121
Mounicou S, Frelon S, Le Guernic A, Eb-Levadoux Y, Camilleri V, Février L, Pierrisnard S, Carasco L, Gilbin R, Mahé K, Tabouret H, Bareille G, Simon O (2019) Use of fish otoliths as a temporal biomarker of field uranium exposure. Sci Total Environ 690:511–521. https://doi.org/10.1016/j.scitotenv.2019.06.534
Munro AR, Gillanders BM, Elsdon TS, Crook DA, Sanger AC (2008) Enriched stable isotope marking of juvenile golden perch (Macquaria ambigua) otoliths. Can J Fish Aquat Sci 65:276–285. https://doi.org/10.1139/F08-010
Munro AR, Gillanders BM, Thurstan S, Crook DA, Sanger AC (2009) Transgenerational marking of freshwater fishes with enriched stable isotopes: a tool for fisheries management and research. J Fish Biol 75:668–684. https://doi.org/10.1111/j.1095-8649.2009.02352.x
Murawski SA, Rago PJ, Trippel EA (2001) Impacts of demographic variation in spawning characteristics on reference points for fishery management. ICES J Mar Sci 58:1002–1014. https://doi.org/10.1006/jmsc.2001.1097
Nazir A, Khan MA (2021) Using otoliths for fish stock discrimination: status and challenges. Acta Ichthyol Piscat 51:199–218
Northcote TG (1984) Mechanisms of fish migration in rivers. In: W H Neil (Ed.), (pp. 317–355). New York, London: Plenum Press.
Oxman DS, Charapata P, McNeel K, Trumble SJ (2021) Reconstructing reproductive histories using hormones stored in incrementally grown structures in fish. Paper presented at the biomarkers in calcified structures (theme 4). world fish congress, Adelaide, Australia. September 20–24, 2021.
Pankhurst NW (2016) Reproduction and Development. In: C B Schreck, L Tort, A P Farrell, C J Brauner (Eds.), Fish Physiology (Vol. 35, pp. 295–331): Academic Press.
Payan P, Borelli G, Priouzeau F, de Pontual H, Boef G, Mayer-Gostan N (2002) Otolith growth in trout Oncorhynchys mykiss: supply of Ca2+ and Sr2+ to the saccular endolymph. J Exp Biol 205:2687–2695
Pereira LA, Santos RV, Hauser M, Duponchelle F, Carvajal F, Pecheyran C, Bérail S, Pouilly M (2019) Commercial traceability of Arapaima spp. fisheries in the Amazon basin: can biogeochemical tags be useful? Biogeosciences 16:1781–1797. https://doi.org/10.5194/bg-16-1781-2019
Pethybridge HR, Choy CA, Polovina JJ, Fulton EA (2018) Improving marine ecosystem models with biochemical tracers. Ann Rev Mar Sci 10:199–230
Phillis CC, Sturrock AM, Johnson RC, Weber PK (2018) Endangered winter-run Chinook salmon rely on diverse rearing habitats in a highly altered landscape. Biol Cons 217:358–362. https://doi.org/10.1016/j.biocon.2017.10.023
Pinto M, Monteiro JN, Crespo D, Costa F, Rosa J, Primo AL, Pardal MA, Martinho F (2021) Influence of oceanic and climate conditions on the early life history of European seabass dicentrarchus labrax. Mar Environ Res 169:105362. https://doi.org/10.1016/j.marenvres.2021.105362
Pollard MJ, Kingsford MJ, Battaglene SC (1999) Chemical marking of juvenile snapper, Pagrus auratus (Sparidae), by incorporation of strontium into dorsal spines. Fish B-Noaa 97:118–131
Post DM (2002) Using stable isotopes to estimate trophic position: models, methods, and assumptions. Ecology 83:703–718. https://doi.org/10.2307/3071875
Quinn TJ, Deriso RB (1999) Quantitative fish dynamics. Oxford University Press, USA
Rahman SA, Abdullah NA, Chowdhury AJK, Yunus K (2018) Fish scales as a bioindicator of potential marine pollutants and carcinogens in Asian sea bass and red tilapia within the coastal waters of pahang. Malaysia J Coast Res 82(120–125):126
Ranaldi MM, Gagnon MM (2008) Zinc incorporation in the otoliths of juvenile pink snapper (Pagrus auratus Forster): the influence of dietary versus waterborne sources. J Exp Mar Biol Ecol 360:56–62. https://doi.org/10.1016/J.JEMBE.2008.03.013
Ranaldi MM, Gagnon MM (2010) Trace metal incorporation in otoliths of pink snapper (Pagrus auratus) as an environmental monitor. Comp Biochem Physiol C: Toxicol Pharmacol 152:248–255. https://doi.org/10.1016/J.CBPC.2010.04.012
Reis-Santos P, Gillanders BM, Tanner SE, Vasconcelos RP, Elsdon TS, Cabral HN (2012) Temporal variability in estuarine fish otolith elemental fingerprints: implications for connectivity assessments. Estuar Coast Shelf Sci 112:216–224. https://doi.org/10.1016/j.ecss.2012.07.027
Reis-Santos P, Tanner SE, Elsdon TS, Cabral HN, Gillanders BM (2013a) Effects of temperature, salinity and water composition on otolith elemental incorporation of Dicentrarchus labrax. J Exp Mar Biol Ecol 446:245–252. https://doi.org/10.1016/j.jembe.2013.05.027
Reis-Santos P, Tanner SE, Vasconcelos RP, Elsdon TS, Cabral HN, Gillanders BM (2013b) Connectivity between estuarine and coastal fish populations: contributions of estuaries are not consistent over time. Mar Ecol Prog Ser 491:177–186. https://doi.org/10.3354/meps10458
Reis-Santos P, Tanner SE, Aboim MA, Vasconcelos RP, Laroche J, Charrier G, Pérez M, Presa P, Gillanders BM, Cabral HN (2018a) Reconciling differences in natural tags to infer demographic and genetic connectivity in marine fish populations. Sci Rep 8:10343. https://doi.org/10.1038/s41598-018-28701-6
Reis-Santos P, Vasconcelos RP, Tanner SE, Fonseca VF, Cabral HN, Gillanders BM (2018b) Extrinsic and intrinsic factors shape the ability of using otolith chemistry to characterize estuarine environmental histories. Mar Environ Res 140:332–341. https://doi.org/10.1016/j.marenvres.2018.06.002
Reis-Santos P, Condini MV, Albuquerque CQ, Saint’Pierre TD, Garcia AM, Gillanders BM, Tanner SE (2021) El Niño – Southern Oscillation drives variations in growth and otolith chemistry in a top predatory fish. Ecol Ind 121:106989. https://doi.org/10.1016/j.ecolind.2020.106989
Ricardo F, Génio L, Costa Leal M, Albuquerque R, Queiroga H, Rosa R, Calado R (2015) Trace element fingerprinting of cockle (Cerastoderma edule) shells can reveal harvesting location in adjacent areas. Sci Rep 5:11932. https://doi.org/10.1038/srep11932
Rideout RM, Tomkiewicz J (2011) Skipped spawning in fishes: more common than you might think. Marine Coast Fish 3:176–189. https://doi.org/10.1080/19425120.2011.556943
Rideout RM, Rose GA, Burton MPM (2005) Skipped spawning in female iteroparous fishes. Fish Fish 6:50–72. https://doi.org/10.1111/j.1467-2679.2005.00174.x
Rogers TA, Fowler AJ, Steer MA, Gillanders BM (2019) Discriminating natal source populations of a temperate marine fish using larval otolith chemistry. Front Mar Sci. https://doi.org/10.3389/fmars.2019.00711
Rogers TA, Redondo Rodriguez A, Fowler AJ, Doubell MJ, Drew MJ, Steer MA, Matthews D, James C, Gillanders BM (2021) Using a biophysical model to investigate connectivity between spawning grounds and nursery areas of King George whiting (Sillaginodes punctatus: Perciformes) in South Australia’s gulfs. Fish Oceanogr 30:51–68. https://doi.org/10.1111/fog.12502
Romero LM, Dickens MJ, Cyr NE (2009) The reactive scope model — a new model integrating homeostasis, allostasis, and stress. Horm Behav 55:375–389. https://doi.org/10.1016/j.yhbeh.2008.12.009
Rooker JR, Secor DH, De Metrio G, Schloesser R, Block BA, Neilson JD (2008) Natal homing and connectivity in Atlantic bluefin tuna populations. Science 322:742–744. https://doi.org/10.1126/science.1161473
Rooker JR, Wells RJD, Block BA, Liu H, Baumann H, Chiang WC, Sluis MZ, Miller NR, Mohan JA, Ohshimo S, Tanaka Y, Dance MA, Dewar H, Snodgrass OE, Shiao JC (2021) Natal origin and age-specific egress of Pacific bluefin tuna from coastal nurseries revealed with geochemical markers. Sci Rep 11:14216. https://doi.org/10.1038/s41598-021-93298-2
Russell AL, Gillanders BM, Barnes TC, Johnson DD, Taylor MD (2021) Inter-estuarine variation in otolith chemistry in a large coastal predator: a viable tool for identifying coastal nurseries? Estuaries Coasts 44:1132–1146. https://doi.org/10.1007/s12237-020-00825-x
Sabetian A, Zhang J, Campbell M, Walter R, Allen H, Reid M, Wijenayake K, Lilkendey J (2021) Fish nearshore habitat-use patterns as ecological indicators of nursery quality. Ecol Ind 131:108225. https://doi.org/10.1016/j.ecolind.2021.108225
Sakamoto T, Komatsu K, Shirai K, Higuchi T, Ishimura T, Setou T, Kamimura Y, Watanabe C, Kawabata A (2019) Combining microvolume isotope analysis and numerical simulation to reproduce fish migration history. Methods Ecol Evol 10:59–69. https://doi.org/10.1111/2041-210X.13098
Samson M. (2021). Otolith microchemistry in stressful environments: otoliths as tools for species identification and as recorders of hypoxia exposure. (Doctoral dissertation). State University of New York, College of environmental science and forestry, Syracuse, New York, USA.
Sanford E, Sones JL, García-Reyes M, Goddard JHR, Largier JL (2019) Widespread shifts in the coastal biota of northern California during the 2014–2016 marine heatwaves. Sci Rep 9:4216. https://doi.org/10.1038/s41598-019-40784-3
Santora JA, Mantua NJ, Schroeder ID, Field JC, Hazen EL, Bograd SJ, Sydeman WJ, Wells BK, Calambokidis J, Saez L, Lawson D, Forney KA (2020) Habitat compression and ecosystem shifts as potential links between marine heatwave and record whale entanglements. Nat Commun 11:536. https://doi.org/10.1038/s41467-019-14215-w
Scargle JD (1982) Studies in astronomical time series analysis. II - statistical aspects of spectral analysis of unevenly spaced data. Astrophys J 263:835–835. https://doi.org/10.1086/160554
Schaffler JJ, Reiss CS, Jones CM (2009) Spatial variation in otolith chemistry of Atlantic croaker larvae in the Mid-Atlantic Bight. Mar Ecol Prog Ser 382:185–195. https://doi.org/10.3354/meps07993
Schindler DE, Hilborn R, Chasco B, Boatright CP, Quinn TP, Rogers LA, Webster MS (2010) Population diversity and the portfolio effect in an exploited species. Nature 465:609–612
Schwarzenbach RP, Escher BI, Fenner K, Hofstetter TB, Johnson CA, Uv G, Wehrli B (2006) The challenge of micropollutants in aquatic systems. Science 313:1072–1077. https://doi.org/10.1126/science.1127291
Secor D (1999) Specifying divergent migrations in the concept of stock: the contingent hypothesis. Fish Res 43:13–34. https://doi.org/10.1016/S0165-7836(99)00064-8
Selleslagh J, Echard A, Pécheyran C, Baudrimont M, Lobry J, Daverat F (2016) Can analysis of Platichthys flesus otoliths provide relevant data on historical metal pollution in estuaries? Experimental and in situ approaches. Sci Total Environ 557–558:20–30. https://doi.org/10.1016/j.scitotenv.2016.03.014
Seyama H, Edmonds JS, Moran MJ, Shibata Y, Soma M, Morita M (1991) Periodicity in fish otolith Sr, Na, and K corresponds with visual banding. Experientia 47:1193–1196. https://doi.org/10.1007/BF01918383
Sherwood GD, Rose GA (2003) Influence of swimming form on otolith 13C in marine fish. Mar Ecol Prog Ser 258:283–289
Shiao JC, Shirai K, Tanaka K, Takahata N, Sano Y, Sung-Yun Hsiao S, Lee DC, Tseng YC (2018) Assimilation of nitrogen and carbon isotopes from fish diets to otoliths as measured by nanoscale secondary ion mass spectrometry. Rapid Commun Mass Spectrom 32:1250–1256. https://doi.org/10.1002/rcm.8171
Shima JS, Swearer SE (2016) Evidence and population consequences of shared larval dispersal histories in a marine fish. Ecology 97:25–31. https://doi.org/10.1890/14-2298.1
Silva AT, Lucas MC, Castro-Santos T, Katopodis C, Baumgartner LJ, Thiem JD, Aarestrup K, Pompeu PS, O’Brien GC, Braun DC, Burnett NJ, Zhu DZ, Fjeldstad H-P, Forseth T, Rajaratnam N, Williams JG, Cooke SJ (2018) The future of fish passage science, engineering, and practice. Fish Fish 19:340–362. https://doi.org/10.1111/faf.12258
Silva AJ, Hellberg RS, Hanner RH (2021) Seafood fraud. In: Hellberg RS, Everstine K, Sklare SA (eds) Food fraud. Academic Press, pp 109–137
Sinclair AF, Swain DP, Hanson JM (2002) Measuring changes in the direction and magnitude of size-selective mortality in a commercial fish population. Can J Fish Aquat Sci 59:361–371. https://doi.org/10.1139/f02-015
Sinnatamby NR, Dempson BJ, James RD, Power M (2015) Latitudinal variation in growth and otolith-inferred field metabolic rates of Canadian young-of-the-year Arctic charr. Ecol Freshw Fish 24:478–488. https://doi.org/10.1111/eff.12166
Sirot C, Grønkjær P, Pedersen JB, Panfili J (2017) Using otolith organic matter to detect diet shifts in Bardiella chrysoura during a period of environmental changes. Mar Ecol Prog Ser 575:137–152
Siskey MR, Lyubchich V, Liang D, Piccoli PM, Secor DH (2016) Periodicity of strontium: calcium across annuli further validates otolith-ageing for Atlantic bluefin tuna (Thunnus thynnus). Fish Res 177:13–17. https://doi.org/10.1016/j.fishres.2016.01.004
Skinner C, Mill AC, Fox MD, Newman SP, Zhu Y, Kuhl A, Polunin NVC (2021) Offshore pelagic subsidies dominate carbon inputs to coral reef predators. Sci Adv. https://doi.org/10.1126/sciadv.abf3792
Smoliński S, Schade FM, Berg F (2020) Assessing the performance of statistical classifiers to discriminate fish stocks using fourier analysis of otolith shape. Can J Fish Aquat Sci 77:674–683. https://doi.org/10.1139/cjfas-2019-0251
Smoliński S, Denechaud C, von Leesen G, Geffen AJ, Grønkjær P, Godiksen JA, Campana SE (2021) Differences in metabolic rate between two Atlantic cod (Gadus morhua) populations estimated with carbon isotopic composition in otoliths. PLoS ONE 16:e0248711. https://doi.org/10.1371/journal.pone.0248711
Solomon CT, Weber PK, Cech JJ, J, Ingram BL, Conrad ME, Machavaram MV, Pogodina AR, Franklin RL, (2006) Experimental determination of the sources of otolith carbon and associated isotopic fractionation. Can J Fish Aquat Sci 63:79–89. https://doi.org/10.1139/f05-200
Starrs D, Ebner BC, Fulton CJ (2016) All in the ears: unlocking the early life history biology and spatial ecology of fishes. Biol Rev 91:86–105. https://doi.org/10.1111/brv.12162
Steube TR, Altenritter ME, Walther BD (2021) Distributive stress: individually variable responses to hypoxia expand trophic niches in fish. Ecology 102:e03356. https://doi.org/10.1002/ecy.3356
Storm-Suke A, Dempson JB, Reist JD, Power M (2007) A field-derived oxygen isotope fractionation equation for Salvelinus species. Rapid Commun Mass Spectrom 21:4109–4116. https://doi.org/10.1002/rcm.3320
Sturrock AM, Trueman CN, Milton JA, Waring CP, Cooper MJ, Hunter E (2014) Physiological influences can outweigh environmental signals in otolith microchemistry research. Mar Ecol Prog Ser 500:245–264. https://doi.org/10.3354/meps10699
Sturrock AM, Hunter E, Milton JA, EIMF, Johnson RC, Waring CP, Trueman CN, (2015a) Quantifying physiological influences on otolith microchemistry. Methods Ecol Evol 6:806–816. https://doi.org/10.1111/2041-210X.12381
Sturrock AM, Wikert JD, Heyne T, Mesick C, Hubbard AE, Hinkelman TM, Weber PK, Whitman GE, Glessner JJ, Johnson RC (2015b) Reconstructing the migratory behavior and long-term survivorship of juvenile chinook salmon under contrasting hydrologic regimes. PLoS ONE 10:e0122380. https://doi.org/10.1371/journal.pone.0122380
Sturrock AM, Carlson SM, Wikert JD, Heyne T, Nusslé S, Merz JE, Sturrock HJW, Johnson RC (2020) Unnatural selection of salmon life histories in a modified riverscape. Glob Chang Biol 26:1235–1247. https://doi.org/10.1111/gcb.14896
Sultana T, Siddique A, Sultana S, Mahboob S, Al-Ghanim K, Ahmed Z (2017) Fish scales as a non-lethal tool of the toxicity of wastewater from the River Chenab. Environ Sci Pollut Res 24:2464–2475. https://doi.org/10.1007/s11356-016-7962-9
Sun J, Davis JA, Stewart R, Palace V (2019) Selenium in White Sturgeon from North San Francisco Bay: The 2015–2017 Sturgeon derby study. SFEI Contribution No. 897. San Francisco Estuary Institute: Richmond, CA.
Swearer SE, Treml EA, Shima JS (2019). A Review of Biophysical Models of Marine Larval Dispersal. https://doi.org/10.1201/9780429026379-7
Tanner SE, Reis-Santos P, Vasconcelos RP, França S, Thorrold SR, Cabral HN (2012) Otolith geochemistry discriminates among estuarine nursery areas of Solea solea and S. senegalensis over time. Mar Ecol Prog Ser 452:193–203
Tanner SE, Reis-Santos P, Vasconcelos RP, Thorrold SR, Cabral HN (2013) Population connectivity of Solea solea and Solea senegalensis over time. J Sea Res 76:82–88. https://doi.org/10.1016/j.seares.2012.11.005
Tanner SE, Reis-Santos P, Cabral HN (2016) Otolith chemistry in stock delineation: a brief overview, current challenges and future prospects. Fish Res 173:206–213. https://doi.org/10.1016/j.fishres.2015.07.019
Thomas ORB, Swearer SE (2019) Otolith biochemistry — a review. Rev Fish Sci Aquacu 27:458–489. https://doi.org/10.1080/23308249.2019.1627285
Thomas ORB, Ganio K, Roberts BR, Swearer SE (2017) Trace element–protein interactions in endolymph from the inner ear of fish: implications for environmental reconstructions using fish otolith chemistry. Metallomics 9:239–249. https://doi.org/10.1039/C6MT00189K
Tomás J, Geffen AJ, Millner RS, Piñeiro CG, Tserpes G (2006) Elemental composition of otolith growth marks in three geographically separated populations of European hake (Merluccius merluccius). Mar Biol 148:1399–1413. https://doi.org/10.1007/s00227-005-0171-6
Tracey DM, Andrews AH, Horn PL, Neil HL (2017) Another New Zealand centenarian: age validation of black cardinalfish (Epigonus telescopus) using lead–radium and bomb radiocarbon dating. Mar Freshw Res 68:352–352. https://doi.org/10.1071/MF15267
Tran NT, Labonne M, Chung M-T, Wang C-H, Huang K-F, Durand J-D, Grudpan C, Chan B, Hoang HD, Panfili J (2021) Natal origin and migration pathways of Mekong catfish (Pangasius krempfi) using strontium isotopes and trace element concentrations in environmental water and otoliths. PLoS ONE 16:e0252769. https://doi.org/10.1371/journal.pone.0252769
Treberg JR, Killen SS, MacCormack TJ, Lamarre SG, Enders EC (2016) Estimates of metabolic rate and major constituents of metabolic demand in fishes under field conditions: methods, proxies, and new perspectives. Comp Biochem Physiol Mol Integr Physiol 202:10–22. https://doi.org/10.1016/j.cbpa.2016.04.022
Tripp A, Murphy HM, Davoren GK (2020) Otolith chemistry reveals natal region of larval capelin in coastal newfoundland. Canada Front Marine Sci. https://doi.org/10.3389/fmars.2020.00258
Trueman CN, St John Glew K (2019) Tracking animal migration with stable isotopes (Second ed.): Elsevier.
Trumble SJ, Robinson EM, Berman-Kowalewski M, Potter CW, Usenko S (2013) Blue whale earplug reveals lifetime contaminant exposure and hormone profiles. Proc Natl Acad Sci 110:16922–16926. https://doi.org/10.1073/pnas.1311418110
Tzadik OE, Curtis JS, Granneman JE, Kurth BN, Pusack TJ, Wallace AA, Hollander DJ, Peebles EB, Stallings CD (2017) Chemical archives in fishes beyond otoliths: a review on the use of other body parts as chronological recorders of microchemical constituents for expanding interpretations of environmental, ecological, and life-history changes. Limnol Oceanogr Method 15:238–263. https://doi.org/10.1002/lom3.10153
Tzeng W-N, Severin KP, Wickström H, Wang C-H (1999) Strontium bands in relation to age marks in otoliths of european eel Anguilla anguilla. https://scholars.lib.ntu.edu.tw/bitstream/123456789/137233/1/18.pdf.
Vandermyde JM, Whitledge GW (2008) Otolith δ15N distinguishes fish from forested and agricultural streams in southern Illinois. J Freshw Ecol 23:333–336. https://doi.org/10.1080/02705060.2008.9664206
Vane K, Larsen T, Kopke BMS-bB, Ekau W, (2018a) Ontogenetic resource utilization and migration reconstruction with δ13C values of essential amino acids in the Cynoscion acoupa otolith. Ecol Evol 8:9859–9869. https://doi.org/10.1002/ece3.4471
Vane K, Wallsgrove NJ, Ekau W, Popp BN (2018b) Reconstructing lifetime nitrogen baselines and trophic position of Cynoscion acoupa from δ15N values of amino acids in otoliths. Mar Ecol Prog Ser 597:1–11. https://doi.org/10.3354/meps12625
Vaz AC, Scarcella G, Pardal MA, Martinho F (2019) Water temperature gradients drive early life-history patterns of the common sole (Solea solea L.) in the Northeast Atlantic and Mediterranean. Aquat Ecol 53:281–294. https://doi.org/10.1007/s10452-019-09688-2
Volk EC, Schroder SL, Grimm JJ (1999) Otolith thermal marking. Fish Res 43:205–219. https://doi.org/10.1016/s0165-7836(99)00073-9
von Leesen G, Bardarson H, Halldórsson SA, Whitehouse MJ, Campana SE (2021) Accuracy of otolith oxygen isotope records analyzed by SIMS as an index of temperature exposure of wild icelandic cod (Gadus morhua). Front Mar Sci. https://doi.org/10.3389/fmars.2021.698908
Vu AV, BaumgartnerLJ M-C, Doran GS, Limburg KE, Gillanders BM, Thiem JD, Howitt JA, Kewish CM, Reinhardt J, Cowx IG (2022) Diverse migration tactics of fishes within the large tropical Mekong river system. Fish Manage Ecol 00:1–16. https://doi.org/10.1111/fme.12566
Walther BD (2019) The art of otolith chemistry: interpreting patterns by integrating perspectives. Mar Freshw Res 70:1643–1658. https://doi.org/10.1071/MF18270
Walther BD, Limburg KE (2012) The use of otolith chemistry to characterize diadromous migrations. J Fish Biol 81:796–825. https://doi.org/10.1111/j.1095-8649.2012.03371.x
Walther BD, Thorrold SR (2010) Limited diversity in natal origins of immature anadromous fish during ocean residency. Can J Fish Aquat Sci 67:1699–1707. https://doi.org/10.1139/f10-086
Wang C-H, Hsu C-C, Tzeng W-N, You C-F, Chang C-W (2011) Origin of the mass mortality of the flathead grey mullet (Mugil cephalus) in the Tanshui River, northern Taiwan, as indicated by otolith elemental signatures. Mar Pollut Bull 62:1809–1813. https://doi.org/10.1016/j.marpolbul.2011.05.011
Wang XT, Sigman DM, Cohen AL, Sinclair DJ, Sherrel RM, Weigand MA, Erler DV, Ren H (2015) Isotopic composition of skeleton-bound organic nitrogen in reef-building symbiotic corals: A new method and proxy evaluation at Bermuda. Geochimica et Cosmochimica Acta 148(2015):179–190. https://doi.org/10.1016/j.gca.2014.09.017
Warren-Myers F, Dempster T, Swearer SE (2018) Otolith mass marking techniques for aquaculture and restocking: benefits and limitations. Rev Fish Biol Fisheries 28:485–501. https://doi.org/10.1007/s11160-018-9515-4
Weber PK, Hutcheon ID, McKeegan KD, Ingram BL (2002) Otolith sulfur isotope method to reconstruct salmon (Oncorhynchus tshawytscha) life history. Can J Fish Aquat Sci 59:587–591. https://doi.org/10.1139/f02-038
Weidman CR, Millner R (2000) High-resolution stable isotope records from North Atlantic cod. Fish Res 46:327–342. https://doi.org/10.1016/S0165-7836(00)00157-0
Wells RJD, Quesnell VA, Humphreys RL Jr, Dewar H, Rooker JR, Alvarado Bremer J, Snodgrass OE (2021) Nursery origin and population connectivity of swordfish Xiphias gladius in the North Pacific Ocean. J Fish Biol 99:354–363. https://doi.org/10.1111/jfb.14723
Whitledge GW, Johnson BM, Martinez PJ (2006) Stable hydrogen isotopic composition of fishes reflects that of their environment. Can J Fish Aquat Sci 63:1746–1751. https://doi.org/10.1139/F06-076
Willmes M, Lewis LS, Davis BE, Loiselle L, James HF, Denny C, Baxter R, Conrad JL, Fangue NA, Hung T-C, Armstrong RA, Williams IS, Holden P, Hobbs JA (2019) Calibrating temperature reconstructions from fish otolith oxygen isotope analysis for California’s critically endangered delta smelt. Rapid Commun Mass Spectrom 33:1207–1220. https://doi.org/10.1002/rcm.8464
Winemiller KO, McIntyre PB, Castello L, Fluet-Chouinard E, Giarrizzo T, Nam S, Baird IG, Darwall W, Lujan NK, Harrison I, Stiassny MLJ, Silvano RAM, Fitzgerald DB, Pelicice FM, Agostinho AA, Gomes LC, Albert JS, Baran E, Petrere M, Zarfl C, Mulligan M, Sullivan JP, Arantes CC, Sousa LM, Koning AA, Hoeinghaus DJ, Sabaj M, Lundberg JG, Armbruster J, Thieme ML, Petry P, Zuanon J, Vilara GT, Snoeks J, Ou C, Rainboth W, Pavanelli CS, Akama A, Av S, Saenz L (2016) Balancing hydropower and biodiversity in the amazon, congo, and mekong. Science 351:128–129. https://doi.org/10.1126/science.aac7082
Woodcock SH, Gillanders BM, Munro AR, Crook DA, Sanger AC (2011) Determining mark success of 15 combinations of enriched stable isotopes for the batch marking of larval otoliths. N Am J Fish Manage 31:843–851. https://doi.org/10.1080/02755947.2011.623760
Woodson LE, Wells BK, Weber PK, MacFarlane RB, Whitman GE, Johnson RC (2013) Size, growth, and origin-dependent mortality of juvenile Chinook salmon Oncorhynchus tshawytscha during early ocean residence. Mar Ecol Prog Ser 487:163–175. https://doi.org/10.3354/meps10353
Wyatt ASJ, Matsumoto R, Chikaraishi Y, Miyairi Y, Yokoyama Y, Sato K, Ohkouchi N, Nagata T (2019) Enhancing insights into foraging specialization in the world’s largest fish using a multi-tissue, multi-isotope approach. Ecol Monogr 89:1–28. https://doi.org/10.1002/ecm.1339
Zazzo A, Smith GR, Patterson WP, Dufour E (2006) Life history reconstruction of modern and fossil sockeye salmon (Oncorhynchus nerka) by oxygen isotopic analysis of otoliths, vertebrae, and teeth: Implication for paleoenvironmental reconstructions. Earth Planet Sci Lett 249:200–215. https://doi.org/10.1016/j.epsl.2006.07.003
Acknowledgements
We thank all participants in the otoliths as biomarkers session at the 2021 World Fisheries Conference. Thank you also to Flora Cordoleani and Corey Phillis for letting us use their otolith data to generate Fig. 2. AMS is supported by a UKRI Future Leaders Fellowship (MR/V023578/1), BDW by an award from the National Science Foundation (OCE-1924011), JL-D by a TMON MarineGEO postdoctoral fellowship and a STRI postdoctoral fellowship, SET by Fundação para a Ciência e a Tecnologia (FCT) (UIDB/04292/2020, CEECIND/02710/2021), and ZAD by an Australian Research Council Future Fellowship (FT190100244). We also thank Nina Wootton for her assistance with the graphical abstract.
Funding
Open Access funding enabled and organized by CAUL and its Member Institutions.
Author information
Authors and Affiliations
Contributions
PRS coordinated manuscript preparation, drafted, edited and combined sections. AMS, BMG, BDW, CI, JL-D, KEL, PRS, SET, ZAD, conceptualized the manuscript building on ideas from the WFC 2021 session on biomarkers led by BDW, CI, KEL, PRS, SET. For individual sections, KH, YH, CI, KEL, AHA, DB drafted the section on chemistry to support age estimation; KEL, DB, M-TC, RJ, DSO, CT, BDW drafted the section on environmental stress and ecophysiology; PRS, ZAD, CI, CT drafted the section on seafood provenance; AMS, MW, LJB, JDT drafted the sections on life history diversity in freshwater and diadromous fishes, and linking individual movements with growth; SET, TR, PRS, BMG drafted the section on natal origins and coastal marine nurseries; JL-D and BDW drafted the section on food webs, DSO and AMS drafted the section on maturity and reproduction, with contributions from PC and ST, and TR and BMG drafted the stock enhancement section. All authors contributed and commented on previous versions of the manuscript, read, and approved the final manuscript.
Corresponding author
Ethics declarations
Conflicts of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
The original online version of this article was revised: the authorship is revised.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Reis-Santos, P., Gillanders, B.M., Sturrock, A.M. et al. Reading the biomineralized book of life: expanding otolith biogeochemical research and applications for fisheries and ecosystem-based management. Rev Fish Biol Fisheries 33, 411–449 (2023). https://doi.org/10.1007/s11160-022-09720-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11160-022-09720-z