
Abstract
Around 36,000 km3 of freshwater flows through rivers and estuarine ecosystems and enter the world’s coastal fishing regions every year. The flow of freshwater and sediments creates regional changes in coastal circulation, stimulates marine productivity and helps define the hydrologic properties of estuarine and oceanic waters. These processes can affect different life stages of marine species either directly, through variations in salinity and temperature, or indirectly, due to changes in the availability of food and habitat. This paper reviews the relationship between freshwater flowing through estuarine and coastal ecosystems, and the variable productivity associated with global marine capture fisheries. The results of a global synthesis revealed that 72% of species representing 77% of the total catch (43 million tonnes) were linked to river flows for at least some part of their life cycle. Insights into how the relationship between flow and fishery production varies globally indicates wild capture fisheries and freshwater resources would benefit from an integrated planning and management approach.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Many of the world’s wild marine fisheries are impacted by large and sometimes abrupt shifts in the abundance and distribution of fish species (Caddy and Gulland 1983; Vert-pre et al. 2013). These changes are often more profound in coastal and continental shelf areas that support around 80% of the world’s wild fish and invertebrate catch (Jackson et al. 2001; Watson et al. 2016). The cause of this variability has been linked to the effects of anthropogenic impacts such as overfishing, pollution and habitat destruction due to industrial-level fishing and coastal development (Baum and Worm 2009; Pikitch et al. 2012). These impacts can have immense effects on population dynamics that influence the growth and decline of fish stocks (Houde 1997; Vert-pre et al. 2013). It is also common for many fish stocks to experience fluctuations in productivity regimes that are influenced by environmental processes (Hare and Mantua 2000). In many coastal regions, river flows are strong drivers of seasonal environmental conditions that create regional changes in coastal circulation, stimulate marine productivity and help define the hydrologic properties of estuarine and coastal waters (Grimes and Kingsford 1996; Milliman and Farnsworth 2013). These processes can affect different life stages of marine species either directly, through variations in salinity and temperature, or indirectly, due to changes in food availability and habitat (Bianchi 2007; Saeck et al. 2013).
Determining how freshwater flowing through rivers and into coastal oceans drives fluctuations in the productivity of marine stocks is a key challenge in fisheries science (Myers 1998; Winemiller 2005). The extent of the dependence between the life cycle of many marine species and freshwater flows is poorly understood (Robins et al. 2005; Costa et al. 2007). To further complicate our understanding of marine species and freshwater flows, the construction of dams for agriculture, industry and urban use has led to the alteration of flows, sediment and nutrient loads in a vast majority of the world’s rivers (Benson 1981; Milliman and Farnsworth 2013). There are now more than 58,000 registered large dams in major rivers globally and many more barriers to river flow such as barrages and weirs are planned (International Commission on Large Dams [ICOLD] 2021). The regulation of freshwater flows coupled with changes to land use in and around coastal areas is believed to have contributed to the decline and collapse of many fish and invertebrate stocks globally (Vörösmarty and Sahagian 2000). In light of these stressors and gaps in our knowledge, it is important to determine how river flows are influencing the productivity regimes of fish stocks at a global scale to improve the management of marine fisheries.
It is widely accepted that coastal ecosystems support many commercially important marine fish and invertebrate species that inhabit these environments for growth and survival (Cowen et al. 2000; Beck et al. 2001; Whitfield and Pattrick 2015). For example, the coastal waters adjacent to large rivers, such as those around the Mississippi Delta (southern United States of America) and Changjiang River estuary (mid-eastern coast of China), host some the most productive marine fisheries in the world (Grimes and Kingsford 1996). While some large rivers have been associated with productive fisheries, much less is known about whether the aggregate influence of small and large rivers has a significant effect on the productivity of fisheries at a global scale. In regions of river influence freshwater flow is a ‘master variable’ that interacts with other environmental factors (e.g. temperature and salinity) to influence fisheries production (Gillanders and Kingsford 2002). However, it is difficult to measure the effects of freshwater flow on fisheries because of the confounding impacts of human development in and around rivers, tidal activity, upwelling and mixing of oceanic waters (Caddy and Bakun 1995; Caddy 2000). Much of the evidence used to study the effects of river flows on fisheries stems from correlative studies that used climate indices, salinity or surface water temperature as indicators of freshwater flow (Hallett et al. 2004; Bakun 2010). A synthesis of the mechanisms underlying the dynamics between river flows and marine fisheries would enable greater clarity on the drivers of fishery productivity and aid the management of coastal fisheries.
Accordingly, the purpose of this review is to examine the drivers and mechanisms underpinning the contribution of river flows to the productivity of wild capture marine fisheries. First, we synthesize literature to propose three drivers that directly and indirectly influence physical and biological mechanisms that cause variability in fisheries production. In the second part of this review, we present a global synthesis of the 19 Food and Agriculture Organisation (FAO) major fishing regions in the world by analysing the top ten fisheries species defined by catch weight and their known connection with freshwater flows. Finally, the implications of changes in river flows on fisheries management are discussed.
Linking river flows to fisheries production
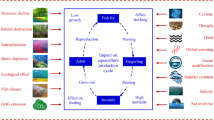
This review distinguishes between three main drivers that link river flows to the productivity of coastal marine fisheries: physical change, biological response and fishery dynamics (Fig. 1) (Bunn and Arthington 2002; Elliott and Whitfield 2011). Within a particular river, estuary or coastal area, the quantity, quality and timing of freshwater flow changes physical conditions in the receiving waters and can alter the availability of suitable habitat (MacCready and Geyer 2009; Poff et al. 2010). From the onset of these changed conditions the growth and survival of many marine species can be affected by short-term and long-term behavioural and physiological changes that might cause them to move to other parts of the river, estuary or coastal area and in extreme cases cause mortality (Drinkwater and Frank 1994). At the same time, the influx of nutrients and organic material from flowing rivers can influence food web dynamics through changes in primary and secondary productivity (Schlacher et al. 2008; Nunn et al. 2012). When the availability of prey changes with physical conditions, many species will be affected by population level processes such as predation and competition (Martino and Able 2003). Adding to these pressures, the level of fishing activity and size selectivity of target stocks can directly or indirectly alter the size and structure of both target and non-target populations (Baum and Worm 2009; Pikitch et al. 2012). Some or all of these drivers, as well as the interactions among drivers, affect the dynamics of fisheries, including: changes in the condition of the catch, recruitment, strength of a year class, catchability of target species or fishers’ harvesting behaviour (Drinkwater and Frank 1994; Gillson 2011).

The conceptual framework reviewing how freshwater flowing from rivers into coastal areas influences the productivity of marine fisheries. Individual images within the biological response box were sourced from Lowerre‐Barbieri et al. (2017) and Capuzzo et al. (2018)
Physical change
The inflow of freshwater has a critical role in driving physical processes in many rivers, estuaries and coastal areas (Poff et al. 1997; Elliott and Whitfield 2011). The key elements of freshwater flow include the quantity, quality and timing of flows (Poff et al. 2010). Quantity refers to the magnitude and frequency of freshwater that flows through rivers. The quality element represents the physical and chemical properties of freshwater flow. Timing is the regularity, duration and rate of change in magnitude of flowing freshwater (Poff et al. 1997). At a global level, there is significant variability in each of these elements due to differences in climatic conditions and drainage basin characteristics such as size, topography, geology, vegetation and human activity (Milliman and Farnsworth 2013).
Once freshwater flowing from rivers enters estuaries and coastal areas it creates a dynamic physical and chemical environment (MacCready and Geyer 2009). The interaction of freshwater flows with tides, wind and coastal currents changes fish habitat conditions such as salinity, water temperature, water circulation, and the level of nutrients and sediments within estuaries and coastal areas (MacCready and Geyer 2009; Milliman and Farnsworth 2013). At times of high river flow the buoyant freshwater that drains into coastal oceans can create low salinity plumes that extend a few kilometres to a thousand kilometres offshore depending on the size of the river (Belkin et al. 2009; Hetland and Hsu 2013).
Salinity
Seasonal changes in the quantity of freshwater flow combined with tidal currents largely determines the spatial extent of salinity within estuarine and coastal areas (Chícharo and Barbosa 2011). When freshwater flows are low or cease to flow, tidal currents enable saline waters to intrude further upstream (MacCready and Geyer 2001). Shallow water estuaries, such as coastal lagoons, in regions that have low flow and high evaporation will often experience sustained periods of high salinity (Telesh and Khlebovich 2010). At times of high flow, the salinity level of shallow water estuaries can be significantly reduced (MacCready and Geyer 2001). In contrast, deep water estuaries found in fjords and large rivers develop horizontal and vertically stratified layers of fresh and saline water (Farmer and Freeland 1983). In coastal areas adjacent to large rivers, such as the North Sea, or regions where multiple smaller rivers flow into the ocean, such as those found in the Gulf of Alaska, low salinity frontal systems may extend well into coastal shelf areas (Belkin et al. 2009; Strub et al. 2013).
Water temperature
Within many estuaries, water temperature is a key physical property that influences water quality and ecosystem function (Van Vliet et al. 2011). The relationship between changes in water temperature caused by freshwater flows varies considerably between different regions of the world. In many temperate estuaries, surface water temperatures can have an inverse relationship with the quantity of freshwater flowing from rivers (Van Vliet et al. 2011). Increasing water velocity associated with seasonal freshwater flow can reduce estuarine residence times and transports heat towards coastal areas (Uncles and Stephens 2001). Flowing rivers may cool temperate estuaries but maximum water temperatures in arctic and subarctic estuaries generally occur during periods of peak flow from melting snow and ice (Hamman et al. 2017). In the tropics, freshwater flows during the wet season generally reflect rainfall temperatures (Strauch et al. 2017). There are some exceptions, with rising water temperatures occurring during periods of sustained low flows and hot ambient air temperatures. Under these conditions, freshwater flow is likely to have a cooling effect, reverting water temperature to ambient air temperature conditions (Van Vliet et al. 2011).
Circulation, nutrients and sediments
Water circulation and nutrient and sediment levels are important to water quality and the health of estuaries and coastal ecosystems (Levin et al. 2001; Ludwig et al. 2009). The movement of water within estuaries and coastal areas is influenced by tides, wind, topography and freshwater flows (MacCready and Geyer 2009). The combined effects of these physical factors can determine the extent of circulation of fresh and ocean water between estuarine and coastal areas (Alber 2002). When high flows occur, rainfall and run-off can carry land-based nutrients, sediments and contaminants into rivers and estuaries (Ludwig et al. 2009). At the same time, increasing water velocity can resuspend bottom sediments and increase bank erosion (Milliman and Farnsworth 2013). The bulk movement of freshwater can flush an estuary of nutrients, sediments and contaminants that become entrained in freshwater flowing to coastal areas (MacCready and Geyer 2009). The mixing of buoyant fresh and tidal oceanic waters can increase levels of turbidity in coastal areas and generate nutrient rich frontal systems (Belkin et al. 2009). During low flows, estuaries mainly rely on tidal currents to circulate water and those with poor flushing mechanisms tend to retain nutrients within the system (Alber 2002). In estuaries with little tidal water movement the slower currents allow suspended sediments to settle to the bottom resulting in reduced turbidity (MacCready and Geyer 2009).
Biological response
Freshwater flows can drive physical changes within estuarine and coastal environments that directly or indirectly affect marine species (Aleem 1972; Drinkwater and Frank 1994). Changes to an individual’s environment can directly affect their physiology and behaviour, and influence growth, mortality, reproduction and the distribution of marine species (Pauly 1980; Able 2005). Many marine species can also be indirectly affected by environmental change to habitat and food availability (Vance et al. 1985; Caddy 2000). As a result, flow induced environmental change can have a profound impact on populations of marine species that inhabit estuarine and coastal areas (Drinkwater and Frank 1994; Kimmerer 2002).
Food web effects
One of the dominant features of food webs in many estuarine and coastal marine ecosystems is the coupling between pelagic and benthic trophic pathways (Alongi 1998). The connections between existing trophic pathways are often decoupled by the onset of seasonal freshwater flows, which create an environment where new trophic pathways are formed (Schlacher et al. 2008; Black et al. 2016). During times of high flow, biota and primary producers that are intolerant to freshwater may disappear (Burford et al. 2012). After high flow conditions have subsided continued stratification of the water column may shift the distribution of phytoplankton into the photic zone (Bianchi 2007). The inflow of inorganic nutrients such as phosphate, nitrate or ammonium, combined with radiation from the sun facilitates ideal algal growth, and potentially bloom conditions (Saeck et al. 2013). In addition, nutrients and organic carbon runoff from catchments into rivers can provide a large food subsidy for the receiving estuarine and coastal ecosystems (Levin et al. 2001). However, excessively high nutrient loads resulting from inputs of fertilisers and large quantities of decomposing organic material can lead to very low dissolved oxygen levels in bottom marine waters that can cause mortality in marine organisms (Diaz and Rosenberg 2008; Reed and Harrison 2016).
In many coastal pelagic food webs, high levels of primary production typically occur for sustained periods of time after a freshwater flow event, although it might not occur immediately because nutrients may need to be transformed into more bioavailable forms (Winder and Cloern 2010). Many zooplankton and forage fish species take advantage of the abundant food supply that become available (Nunn et al. 2012). For example, Pacific herring (Clupea pallasii) in the Strait of Georgia, Canada, has developed a life history strategy where the timing of spawning events is matched with spring flows and phytoplankton blooms. It is thought this match in timing increases the successful transition to the first feeding of larval C. pallasii and supports its ongoing growth and survival. The match or mismatch between the spring bloom and spawning has been found to largely explain the variability in the number of C. pallasii (Schweigert et al. 2013).
Ontogenetic development and dietary changes may also be an important part of the response of marine species to changes in river flow. The abundance and diversity of available prey is particularly important for many species that use estuaries as nursery habitats since their preferred diet and size of prey often shifts between larval and juvenile life stages (Peck et al. 2012). A shift in the diet of larval fish is thought to occur in most species from the ontogenetic development of an enhanced swimming ability and improved vision (Teodosio et al. 2016). For example, in the Gulf of Mexico, the gulf menhaden (Brevoortia patronus) young larvae (< 5 mm) are thought to prey primarily on both phytoplankton and microzooplankton, e.g. dinoflagellates and tintinnids, while older larvae consume mainly microzooplankton, e.g. copepod nauplii, which are found in large numbers after the onset of spring freshwater flows (Stoecker and Govoni 1984; Stoecker and Capuzzo 1990). As the larvae develop into juveniles B. patronus prefer phytoplankton and detritus (Olsen et al. 2014). The timing of freshwater flows combined with a species behaviour, like those found in B. patronus, are particularly important for marine species that need to maximise lipid storage prior to emigrating offshore (Nunn et al. 2012).
Retention and transport
Many marine species spawn offshore and rely on ocean currents to transport eggs or larvae to coastal nursery habitats (Levin 2006). The interaction between freshwater flows and ocean currents can create hydrodynamic conditions that either positively or negatively impact early life stages (Pineda 2007). For example, the level of mixing between freshwater flow from rivers along the Norwegian coast and the Atlantic current has been positively linked to changes in the productivity regimes of the Barents Sea cod (Gadus morhua) population. For recently spawned G. morhua eggs to reach nursery grounds up to 1,200 km away requires optimal buoyancy conditions. Changes in the salinity and temperature of the Norwegian coastal current determines the vertical position of G. morhua eggs and thus affects the distance travelled toward their Barents Sea nursery grounds (Ottersen et al. 2014). An example of a negative impact occurs in the white shrimp (Penaeus setiferus) population in the area of the Terminos Lagoon, southwestern Gulf of Mexico (Gracia 1991). High freshwater flows from the Palizada River during the wet season restricted the access of P. setiferus to estuarine nursery habitats and had a negative effect on growth and survival.
The interaction between freshwater flows and coastal currents can also create hydrodynamic features that retain early life stages within the same region (Epifanio and Garvine 2001). For example, recently emerged capelin larvae (Mallotus villosus) within the Gulf of St. Lawrence, Canada, are concentrated in the upper estuary regions. Freshwater flow during spring and early summer creates a low salinity coastal jet that advects larvae toward the lower estuary and forces the separation of coastal currents creating a large gyre within the boundaries of the estuary. The change in estuarine circulation helps both transport capelin larvae to the north-west area of the estuary and retains them there while they grow. Once the freshwater flow subsides, the gyre loses strength and the direction of currents in the lower estuary allows M. villosus to emigrate into deeper waters (Ouellet et al. 2013).
Movement and migration
The changes in estuarine and coastal conditions forced by the influx of freshwater can cause variability in the movement and migration of marine species (Grimes and Kingsford 1996; Caddy 2000). For example, the effect of river flow from the Congo and Niger Rivers on the distribution of sardinella (Sardinella maderensis and Sardinella aurita) is an important driver of population movement in coastal waters near Gabon and the Republic of Congo, equatorial West Africa. At times when the Congo River plume extends northward, S. aurita are concentrated within the coastal boundaries of the warm and low saline water mass. When the freshwater outflow from Niger River shifts south so does S. maderensis, where the population is distributed between both countries. However, these same conditions force S. aurita further offshore (Binet et al. 2001).
Many diadromous fish species use olfactory senses to detect changes in water chemistry that are carried by freshwater flows (Dittman and Quinn 1996; Harrison et al. 2014). These olfactory cues are thought to be one of the main mechanisms used by salmonids to return to their natal rivers to spawn (Milner et al. 2012). For example, Atlantic salmon (Salmo salar) can spend up to 5 years at sea to feed and grow to attain sexual maturity before undertaking their return migration to natal rivers (Webb et al. 2007). The return migration coincides with increased river flow that helps S. salar locate their home river system and spawning area within the catchment (Milner et al. 2012).
Freshwater flows were also found to be an important factor in the migration of eastern king prawns (Penaeus plebejus), in Moreton Bay, Australia (Loneragan 1999). High summer flows increase the delivery of nutrients, stimulating phytoplankton production, and providing a trigger for juvenile prawns to commence emigration from rivers into the bay area where P. plebejus continue to grow and mature.
Competition
It has long been recognised that the population dynamics of many fish and invertebrate populations are driven by changes in competition for food and habitat (Myers 2001; Le Pape and Bonhommeau 2015). Marine species that inhabit estuarine and coastal areas are particularly susceptible to changes in competition caused by variable freshwater flows (Martino and Able 2003). Periods of high flow can shift the available habitat for many marine species downstream toward the mouth of the estuary, which concentrates competition for food within this region (Rulifson and Manooch 1990). For instance, in the Chikuro River estuary, Japan, the abundance of juvenile Japanese seaperch (Lateolabrax japonicas) was lower at times of high freshwater flow. A shift in prey due to increases in water temperature led to the density-dependent regulation of juvenile L. japonicas from greater intra-specific competition for food that negatively impacted their growth and survival (Shoji and Tanaka 2008).
At times of high flow and flood, marine species will often aggregate in coastal plumes and frontal systems where they converge and compete for food and seek refuge from a wide range of predators, including mammals and sea birds (Grimes and Kingsford 1996). During peak flows from the Columbia River (in north-western United States of America), a large river plume extends along the coast for hundreds of kilometres to the north and south of the river mouth. In the vicinity of the plume many forage fish such as northern anchovy (Engraulis mordax), Pacific herring and Pacific sardine (Sardinops sagax) are found to aggregate in large numbers. The abundance of forage fish increased inter-specific competition between those fish seeking shelter from predators. At the same time, outward migrating juvenile Pacific salmonids (Oncorhynchus spp.) remained under the protective cover of the turbid river plume, which aided predator avoidance and improved salmonid survival (Phillips et al. 2017).
Fishery dynamics
The previous sections have identified many of the physical and biological mechanisms resulting from changes in freshwater flows that influence fisheries productivity. The effects of these mechanisms are interrelated and operate over different spatial and temporal scales, which causes variability in marine fish populations and the dynamics of fisheries (Drinkwater and Frank 1994). Fishery dynamics can be defined as the way in which exploited populations respond to changes in physical and biological mechanisms, and fishers' harvesting behaviour (e.g. fishing effort or the type of gear) (Szuwalski and Thorson 2017). The level of fishing pressure exerted upon fish stocks can affect the geographic distribution, size and structure of target and non-target populations (Baum and Worm 2009; Pikitch et al. 2012). Separating the interactive effects of fishing pressure and freshwater flows on exploited populations is very difficult (Planque et al. 2010). Therefore, the objective here is to propose four fishery characteristics that demonstrate how freshwater flows can either positively or negatively impact fisheries productivity, including: recruitment, condition of the catch, strength of a year class and catchability (Table 1). Also, recognising that fishers' harvesting behaviour and the level of fishing pressure applied to a stock has a critical role in influencing fisheries production.
Global synthesis of major FAO fishing areas and freshwater flow
The aim of this section is to synthesise the relationships between rivers that flow into the ocean and the world’s commercial fisheries. The objective here is to analyse literature and FAO data to gain an insight into how freshwater flows might influence commercially important fish and invertebrate species in different regions of the world.
Method
The top ten species of marine fish and invertebrates caught, by weight, for each of the 19 FAO major fishing areas from around the world were identified using the FAO FishStatJ 3.03.0 application (FAO 2015). The data were selected by grouping each species and FAO major areas, once grouped they were aggregated by weight in tonnes and ranked by their average catch weight from 2011 to 2015. These years contained most data at the species-level for each of the FAO major areas. The non-specific class Osteichthyes (bony fish) was removed from the dataset because there were too many possible species (i.e. around 28,000) that could belong this group. Those FAO areas with data at a higher taxonomic resolution, such as Sciaenidae in the eastern Indian Ocean (area 57), were cross checked with supplementary FAO records from FAO Fisheries and Aquaculture online to identify which commercially fished species belonged to this higher taxon for a particular area. This additional species-level information was included in the analysis and remained linked to the higher taxonomic group within the database created for this study. The existence of these higher taxa in the FAO data is the reason why there are more than ten species for some FAO areas. Further, no attempt to calculate catch per unit effort was made in this study because of the known difficulties in estimating fishing effort for each fishery within the FAO areas (Bell et al. 2017). It should also be acknowledged that the FAO catch data used in this study does not include discards or catches originating from illegal, unreported or unregulated (IUU) fishing, which may cause the catch in some areas to be underestimated.
A literature search was conducted using Google Scholar and Web of Science bibliographic databases to identify the habitat used by each of the top ten FAO listed species throughout their lifecycle. The search terms were confined to the name of each species within the geographic boundaries of the FAO area being searched. In cases where the literature implied a species did not inhabit riverine or estuarine environments, then Fishbase was used to confirm their life history status (Froese and Pauly 2010). Fishbase was also used to confirm the life history of species where the literature provided contrasting findings for the same species. The information for these species were then entered into our database. Literature that identified species inhabiting riverine and estuarine environments for at least some part of their life cycle were further analysed to determine the extent of their association with freshwater flow. Studies that provided a statistical analysis which linked freshwater flow to particular estuaries and fish, or invertebrate species were preferred, but not always available. In the cases where a statistical analysis was not available the relationship was either observed or inferred, by changes in environmental conditions such as salinity, or the association between river flow and the presence or absence of species. Once a publication was found containing all of the necessary information, such as the latitude and longitude of the estuary, and the species association with freshwater, it was then recorded in a database.
After reviewing the literature, we decided to focus our study on the type of association (i.e. trophic, ontogenetic, migration or no association) and its potential link to commercial catch instead of evaluating the positive and negative effects of changes in flow on catch. Determining a positive or negative effect is complicated by the variability in river flows throughout a species lifecycle. For example, (a) high flows might negatively affect larval fish by physically forcing them out of areas of suitable estuarine habitat given their small size and limited swimming ability but positively affect adult fish by increasing suitable habitat and food availability in coastal areas, and (b) low to moderate flows might be positively associated with fish abundance, but once flow exceeds an optimum threshold (i.e. at extreme low or very high flows) the association could become negative (Aleem 1972; Drinkwater and Frank 1994; Grimes and Kingsford 1996).
We define estuaries generally following the Coastal and Marine Ecological Classification Standard (CMECS) as waters that are tidally influenced, have an open connection to the ocean, receive freshwater input and are partly enclosed by land. Estuaries may be located on continents or islands in waters of any depth, such as fjords, large embayments, the mouth of coastal creeks, lagoonal and riverine estuaries (FGDC 2012).
Species guild associations were used to identify patterns in the relationships between river flow and catches that might not be apparent at the species-level. The top ten species from all the major FAO areas were classified into marine (Straggler, Estuarine-opportunist and Estuarine-dependent) and diadromous (Anadromous and Catadromous) guilds defined in Potter et al. (2015) (Table 2). This subset of guilds was chosen because of the focus on marine wild capture fisheries in this study. In cases where the literature for a particular species did not provide enough information to classify its guild then a secondary source, such as Fishbase or FAO Online, was used to confirm their classification.
In order to analyse the effects of river flow on catches in each of the FAO major fishing areas the river flow, total suspended solids (TSS) and total dissolved solids (TDS) data from Milliman and Farnsworth (2013) were used. Milliman and Farnsworth (2013) was chosen because it provides a comprehensive, global data source for river flow, suspended solids and dissolved solids. These authors defined TSS as fluvially suspended material that was between 0.45 and 0.62 µm. TDS was classified as material finer than 0.45 µm, which included the combined total of different inorganic compounds, such as salts and metals. The data contained within the supplementary information from Milliman and Farnsworth (2013) were analysed using R version 3.5.2 (R Core Team 2018). Each of the rivers found within the geographical boundaries of a particular FAO region were assigned to that area. This enabled the calculation of total flows, TSS and TDS for each of the major FAO fishing areas.
Results and discussion
A review of the literature revealed that 87% of the studies (n = 258) identified had linked commercially fished FAO species to freshwater flowing from rivers into the global oceans (Table 3). Among these, 95% were peer reviewed journal articles and the remaining 5% were technical reports and theses. The majority of the literature came from studies in United States of America (26%), South Africa (7%), India (6%) followed by Brazil, Canada and Russia (around 5% each) with the remaining countries comprising less than 4% of the identified studies. Although the distribution of literature was geographically biased, we managed to find studies for each of the top ten species from all of the FAO major fishing areas. Those research articles that provided a statistical analysis between the commercial species caught and freshwater flows comprised 60% of the studies, whereas 22% observed this relationship, e.g. through changes in salinity, and for 18% of the articles it was inferred, e.g. from the presence or absence of species and freshwater flows. Most of the studies (89%) used fishery-independent surveys to collect their data and only 11% relied solely on commercial fisheries landings or catch data. The largest proportion of studies (10%) were found in the east coast of South America (area 41), and the coastlines bordering the northwest Pacific Ocean (area 61). Very few of the studies (4%) were found in the regions around Antarctica (areas 48, 58 and 88). A single estuary was found in the Kerguelen Islands, southern Atlantic Ocean (area 48), and no estuaries were found in Antarctic and Southern Indian Ocean (area 58) and Pacific Antarctic (area 88). The deep embayments and fjords along the coast in the Gulf of Alaska and Bering Sea (area 67) accounted for 13% of the estuaries found in this study. The coastal areas in the eastern Indian Ocean (area 57) represented the largest proportion of species (10%). This was mainly due to this area having the largest number of FAO species at a higher taxon, thus more species level information representing these higher taxa were found.
The total average annual catch of all marine wild capture fish and invertebrate species from 2011 to 2015 was 73.23 million tonnes (Fig. 2a). The top ten species from all of the major FAO fishing areas represents close to 60% of the global catch (Fig. 2c). Moreover, the top ten species that were linked to freshwater flows accounted for around 45% of the total global catch (Fig. 2e). Those species having no association with freshwater flow represented 13% of the global catch. The largest proportion of the global catch for those species linked to freshwater flows was found in the Atlantic Northeast and Pacific Northwest, each sharing around 9% of the total harvest, with the Pacific Southeast having just over 8% of the catch. The main species caught in these areas were Atlantic herring (Clupea harengus), walleye pollock (Theragra chalcogramma), and Peruvian anchoveta (Engraulis ringins). It is also worth noting that the Pacific Western Central had the second largest total catch, 11 million tonnes, but one of the lower catches of fish that were related to river flows, 1 million tonnes. The high catch and low association with river flows in this region was due to a large share of the catch belonging to highly migratory pelagic species such as skipjack tuna (Katsuwonus pelamis) and yellowfin tuna (Thunnus albacares). Fishers target the tuna as they follow forage fish prey near the nutrient rich warm pool-cold tongue convergence zone, a unique feature of the equatorial Pacific Western Central region (Lehodey 2001). The lowest catch was recorded in the Arctic Sea, which was less than 1% of the total global catch. A common problem in this area is the absence of monitoring, control and surveillance of fishing activity reported to the FAO, thus this result is probably not a reliable indicator of the total catch (Pauly et al. 2008).

A series of charts showing each of the FAO major fishing areas with (a) total average catch of fish and invertebrate species from 2011 to 2015, (b) total catch represented by the top ten fish and invertebrate species, (c) catch of those top ten species that were found to have an association with freshwater flows, (d) total river flow into the ocean, (e) total suspended solids (TSS), and (f) total dissolved solids (TDS). The FAO catch data were sourced from FAO (2015) and the TSS and TDS data were sourced from Milliman and Farnsworth (2013)
There is approximately 36,000 km3 of freshwater, 18 billion tonnes of sediments and 4 billion tonnes of dissolved material flowing into the world’s coastal fishing areas every year (Fig. 2b, d, f) (Milliman and Farnsworth 2013). The river outflow from the Atlantic Southwest (9,700 km3 per year) was the most significant feature shown in Fig. 2b. Nearly 65% of the flow from this area stems from the large drainage basin of the Amazon River. The debris and sedimentation deposited in the shelf area near the Amazon River has created a vast region of muddy and coralline benthic habitat that has little biodiversity (Moura et al. 2016). This meant that much of the northern section of the Atlantic Southwest is unsuitable for commercial fishing (Food and Agriculture Organisation [FAO] 2011). The Pacific Western Central had the second largest inflow of freshwater (4,830 km3 per year) combined with the greatest load of suspended and dissolved sediments (7,220 and 840 million tonnes per year, respectively) (Fig. 2b, d, f). Episodic and intense rainfall occurring in areas throughout this region can generate significant river flows with large sediment loads originating from the rapid erosion of volcanic bedrock (Milliman and Farnsworth 2013). Overall, there was considerable variability in river flows, sediments and dissolved material flowing into each FAO area.
Globally, those fish and invertebrate species that were linked to freshwater flows accounted for 77% of the total catch (43 million tonnes) from the FAO’s top ten fished species for each area. However, the relationship between each species, freshwater flows and fishing area varied considerably among FAO areas (Fig. 3). Some of this variability can be explained by the way individual species have adapted to abiotic conditions within different regions of the world. More than half of the species (57%) associated with freshwater flows were found in temperate regions. Patterns in the life history of many marine species in temperate areas are strongly cued to seasonal changes in river flows (Winemiller 2005). Unlike the temperate regions, in tropical marine environments only 17% of species were associated with freshwater flows. It is likely that many marine species in the tropics have developed life history strategies that avoid the potentially lethal effects of abrupt and unpredictable river flows (Connell 1978; Winemiller and Rose 1992). The subarctic and arctic regions accounted for 15% of species. Species in these areas have either biologically adapted to cold water environments or show strong migratory behaviour (Clarke and Johnston 1999). Lastly, dry and arid areas had the least number species (11%) associated with river flows. Most species in these regions were found to be migratory and were likely to take advantage of the benefits from flowing rivers when the opportunity arises.

World map of the FAO major fishing areas with pie charts inset within each region showing the percentage of catch for the top ten species that have an association with freshwater flow (blue) and those where no freshwater association was found in the literature (red). The small cyan coloured dots represent the location of estuaries where commercial fish and invertebrate species were identified at some point throughout their lifecycle
Overall, 199 out of the 276 (72%) commercially fished species in this study were linked to river flows. Further analyses of these species revealed that 83% had a trophic association with flowing freshwaters (Fig. 4). This relationship was mainly driven by an increase in the availability of both pelagic and benthic prey, which for many species included phytoplankton, zooplankton, detritus or forage fish. Around 14% of species, such as salmonids and clupeids, required changes in their physiochemical habitat (e.g. salinity or temperature) brought about by freshwater flows to complete a life stage. Fewer species, around 3%, were found to rely on freshwater flows in order to successfully migrate to or from estuarine areas.

The percentage of fisheries species having a migratory, trophic or ontogenetic relationship with flowing freshwaters
Most of the top ten species (59%) that were linked to freshwater flows were classified as estuarine-opportunists (Fig. 5). In these cases, species were identified as inhabiting both coastal and estuarine areas, predominately as juveniles. The presence of estuarine-opportunists ranged from 20 to 100% in all FAO areas, excluding Antarctic waters. Just over 19% of marine species were found to be estuarine-dependent, which means throughout their juvenile life stage they were only found within estuarine environments. Around 40% of these species were crustaceans from the penaeid family. Those classified as marine stragglers accounted for 18% of species, since they were only found within estuaries on an irregular basis. A majority of stragglers (87%) were found in estuaries and coastal areas boarding both the Atlantic Ocean and Mediterranean Sea. Anadromous species made up 4% of the guild associations classified in this study and occurred in large numbers in the Arctic Sea and Pacific Northeast. Lastly, none of the species in this study were associated with the catadromous guild.

World map of the FAO major fishing areas with pie charts inset within each region showing percentage of species classified as either anadromous (blue), estuarine-dependent (green), opportunist (orange) or straggler (red). The guild definitions from Potter et al (2015) were used to categorise each species. The small cyan coloured dots represent the location of estuaries where commercial fish and invertebrate species were identified at some point throughout their lifecycle
The data suggest that the majority of species (77%) have a facultative association with freshwater flows because they enter estuaries irregularly, or in low numbers (i.e. estuarine-opportunists and marine stragglers). Fewer species (23%) were found to have an obligate relationship where they require estuarine and riverine habitats to complete a life stage (i.e. estuarine-dependent and anadromous). Moreover, the results from both the analyses of flow and guild associations indicates many of the commercial fish and invertebrate species have a preference for the food resources available in productive estuarine and coastal waters, which are driven by changes in freshwater flows. However, the facultative nature of this relationship does make it challenging to quantify how the variability in river flows affects these species throughout their lifecycle.
For the fish and invertebrates considered in this study, 85% of species had an association with freshwater flow at the juvenile stage of development (Fig. 6). This result is consistent with many other studies that have identified estuarine and coastal areas as critical nursery habitats and areas of refuge from predators for marine species (Beck et al. 2001; Sheaves et al. 2015; Whitfield and Pattrick 2015). Most adult species (58%) were also found to be inhabiting estuarine and coastal areas throughout their life cycle. Though, the presence of adult species in estuaries and coastal areas is more than likely facultative, it goes some way to supporting the idea that many species are taking advantage of the food subsidies associated with flowing rivers. Fewer species (46%) were found associated with freshwater flows during their larval stage (from 16 of the FAO areas), and only 19% as eggs (from seven areas). Unlike the juvenile and adult life stages the association between freshwater flow and species found spawning was low (11%). The lower proportion of species found with eggs or spawning in estuaries and coastal areas may be an observation bias because it is more difficult to survey spawning events and eggs compared to other life stages. The presence of large numbers of adult marine species found associated with freshwater flows in this study does warrant further investigation into the possibility of spawning activity in these areas.

FAO major areas and species that have an association with freshwater flow for the life stages shown as a circle, consecutive stages are linked by a line
Assumptions and caveats
A key assumption of this review is that factors affecting individual species at different life stages could influence fish stocks within each FAO area. However, species within some FAO areas have been genetically identified as belonging to separate fish stocks. For example, Pacific whiting in FAO Area 67 have been identified as two separate stocks, one found in the Strait of Georgia and the other migrating along the coast (Phillips et al. 2007). In this case, we found literature that linked flows from the Fraser River to the survival of larval and juvenile Pacific whiting in the Strait of Georgia (Guan et al. 2018). A separate study was found for the coastal stock, where the abundance of juvenile Pacific whiting was positively influenced by an increase in food availability associated with the Columbia River plume (Emmet et al. 2006).
A limitation of this review is that only two bibliographic databases (Google Scholar and Web of Science) were used for literature searches. As a result, the literature search may not have been as exhaustive as possible. To reduce the risk of omitting important information the database search terms were intentionally broad and included the name of each species within the geographic boundaries of the FAO area being searched. Fishbase was also used to confirm the life history status of species, which was particularly useful where the literature provided contrasting findings for the same species (Froese and Pauly 2010).
Finding multiple studies for each species, fish stock and FAO area was desirable but not always possible. The disadvantage of this approach was that some FAO areas were better represented than others, which led to geographical bias in the number of studies found in some regions of the world. In particular, the number of studies in tropical regions was low in comparison with temperate areas. The lack of studies in tropical regions provides an opportunity for future research to bridge this gap in knowledge.
Another potential limitation is the use of FAO catch data instead of a fishery catch rate such as catch per unit effort (CPUE). Estimating fishing effort for each fishery within the FAO areas is difficult because of the lack of reporting of vessel information and the type of gear used by many countries (Bell et al. 2017). Consequently, effort statistics are not always available from the FAO or need to be verified to account for fishing power, efficiency and the type of gear (FAO 2011). The FAO catch data are based on landings from each country that tend to be more complete and provide important insights into the status of global marine wild capture fisheries (FAO 2011; Bell et al. 2017).
Implications for fisheries management
The strong association between river flows and most of the world’s commercially important fishery species found in this study raises concerns about how changes in flow may affect fish stocks on a global scale. The effects of river flow regulation from the construction large dams have already been implicated in the decline and collapse of many fish stocks globally (Drinkwater and Frank 1994; Grimes and Kingsford 1996). However, the ability to determine the effect of changes in river flows on the productivity of marine stocks remains a key challenge in fisheries science (Myers 1998; Winemiller 2005). It is challenging because the interaction between a species’ population dynamics and river flows are not well-understood in many regions of the world (Robins et al. 2005; Costa et al. 2007). Additionally, changes in river flows are often managed within the jurisdiction of catchment managers and hydrologists (Richter 2010). Allocations of water for use by industries downstream, such as coastal fisheries, may not be prioritised in the planning process (Poff et al. 2003). Therefore, identifying how changes in river flows influence marine populations is imperative to proactively manage the potential impact on fish stocks.
One way to incorporate changes in river flow into fisheries management planning is through stock assessment models (Myers 1998). This has been achieved by considering river flow in stock-recruitment relationships. For example, accounting for the impact of freshwater flowing from the Fraser River, Canada, helped explain 58% of the recruitment variability in Pacific herring stocks. In this case, river flow was thought to enhance primary productivity, which positively benefited larval and juvenile herring growth and survival (Stocker et al. 1985). Similarly, the interannual changes in river flow were used to explain the variability in the recruitment strength of multiple cohorts of white shrimp (Gracia 1991). The author developed multiple stock-recruitment models that reflected periods in time where river flows had a positive or negative effect on recruitment. These models were then used to explore how the different scenarios of river flows combined with fishing pressure would impact fishery yield and allow fishery managers to consider environmental changes in their management strategies.
Conclusions
The regulation of freshwater flows coupled with changes to land use in and around river basins has far-reaching consequences not only for the ecology of rivers but also for fisheries in the receiving estuarine and coastal ecosystems (Bunn and Arthington 2002). Even in regions where there are low numbers of species associated with flow, managers should consider how the impact of changes in quality, quantity and timing of freshwater flows affects the productivity of fishing areas (Benson 1981). Likewise, water resource managers should take into account the downstream effects of future freshwater allocations.
Further exploring the effects of climate change on the association between river flow and marine fisheries production may be an important area for future research. The effects of climate change can impact river flow regimes by altering spatial and temporal rainfall patterns and the frequency of very cold or hot seasonal temperatures that can affect the timing of snow melt (Adam et al. 2009; Graham and Harrod 2009). An increase in temperature of 1.5 to 3.0° C from global warming is predicted to increase periods of extreme low and high flow events (Thompson et al. 2021). Modifications to river flow regimes could affect the physiology, behaviour and population dynamics of marine species (Rijnsdorp et al. 2009). Many marine species that inhabit coastal regions have complex life histories where growth and survival in the early life stages is largely dependent on conditions in coastal, estuarine and riverine nursery areas (Vörösmarty and Sahagian 2000; Drinkwater and Frank 1994). Changes in coastal habitat caused by global warming could affect the growth and survival of marine species and lead to reduced recruitment that negatively impacts the productivity of fish stocks (Rijnsdorp et al. 2009). To protect global fishery resources in the face these challenges future research could provide valuable insights on the links between climate change and river flows and their effect on marine species.
Data availability
The FAO data that support the findings of this study are publicly available from the FAO Fisheries Division (http://www.fao.org/fishery/statistics/software/fishstatj/en). The remaining data analysed during the current study are available from the corresponding author on reasonable request.
Code availability
Not applicable.
References
Able KW (2005) A re-examination of fish estuarine dependence: evidence for connectivity between estuarine and ocean habitats. Estuar Coast Shelf Sci 64:5–17. https://doi.org/10.1016/j.ecss.2005.02.002
Adam JC, Hamlet AF, Lettenmaier DP (2009) Implications of global climate change for snowmelt hydrology in the twenty-first century. Hydrol Process 23:962–972. https://doi.org/10.1002/hyp.7201
Alber M (2002) A conceptual model of estuarine freshwater inflow management. Estuaries 25:1246–1261. https://doi.org/10.1007/BF02692222
Aleem AA (1973) Effect of river outflow management on marine life. Mar Biol 15:200–208. https://doi.org/10.1007/BF00383550
Alongi DM (1998) Coastal Ecosystem Processes. CRC Press, Boca Raton. https://doi.org/10.1201/9781003057864
Bakun A (2010) Linking climate to population variability in marine ecosystems characterized by non-simple dynamics: conceptual templates and schematic constructs. J Mar Syst 79:361–373. https://doi.org/10.1016/j.jmarsys.2008.12.008
Baum JK, Worm B (2009) Cascading top-down effects of changing oceanic predator abundances. J Anim Ecol 78:699–714. https://doi.org/10.1111/j.1365-2656.2009.01531.x
Beck MW, Heck KL Jr, Able KW, Childers DL, Eggleston DB, Gillanders BM, Halpern B, Hays CG, Hoshino K, Minello TJ (2001) The identification, conservation, and management of estuarine and marine nurseries for fish and invertebrates: a better understanding of the habitats that serve as nurseries for marine species and the factors that create site-specific variability in nursery quality will improve conservation and management of these areas. Bioscience 51:633–641. https://doi.org/10.1641/0006-3568(2001)051[0633:TICAMO]2.0.CO;2
Belkin IM, Cornillon PC, Sherman K (2009) Fronts in large marine ecosystems. Prog Oceanogr 81:223–236. https://doi.org/10.1016/j.pocean.2009.04.015
Bell JD, Watson RA, Ye Y (2017) Global fishing capacity and fishing effort from 1950 to 2012. Fish Fish 18:489–505. https://doi.org/10.1111/faf.12187
Benson NG (1981) The freshwater-inflow-to-estuaries issue. Fisheries 6:8–10. https://doi.org/10.1577/1548-8446(1981)006%3c0008:TFI%3e2.0.CO;2
Bianchi TS (2007) Biogeochemistry of Estuaries. Oxford University Press, UK
Binet D, Gobert B, Maloueki L (2001) El Niño-like warm events in the Eastern Atlantic (6 N, 20 S) and fish availability from Congo to Angola (1964–1999). Aquat Living Resour 14:99–113. https://doi.org/10.1016/S0990-7440(01)01105-6
Black KP, Longmore AR, Hamer PA, Lee R, Swearer SE, Jenkins GP (2016) Linking nutrient inputs, phytoplankton composition, zooplankton dynamics and the recruitment of pink snapper, Chrysophrys auratus, in a temperate bay. Estuar Coast Shelf Sci 183:150–162. https://doi.org/10.1016/j.ecss.2016.10.032
Bunn SE, Arthington AH (2002) Basic principles and ecological consequences of altered flow regimes for aquatic biodiversity. Environ Manage 30:492–507. https://doi.org/10.1007/s00267-002-2737-0
Burford M, Webster I, Revill A, Kenyon R, Whittle M, Curwen G (2012) Controls on phytoplankton productivity in a wet–dry tropical estuary. Estuar Coast Shelf Sci 113:141–151. https://doi.org/10.1016/j.ecss.2012.07.017
Caddy J (2000) Marine catchment basin effects versus impacts of fisheries on semi-enclosed seas. ICES J Mar Sci 57:628–640. https://doi.org/10.1006/jmsc.2000.0739
Caddy JF, Bakun A (1995) Marine catchment basins and anthropogenic effects on coastal fishery ecosystems. In: Effects of Riverine Inputs on Coastal Ecosystems and Fisheries Resources. FAO Fisheries Technical Papers, Volume 349. FAO, Rome, pp 119–133. https://www.fao.org/3/V4890E/V4890E00.htm#TOC
Caddy J, Gulland J (1983) Historical patterns of fish stocks. Mar Policy 7:267–278. https://doi.org/10.1016/0308-597X(83)90040-4
Capuzzo E, Lynam CP, Barry J, Stephens D, Forster RM, Greenwood N, McQuatters‐Gollop A, Silva T, Leeuwen SM, Engelhard GH (2018) A decline in primary production in the North Sea over 25 years, associated with reductions in zooplankton abundance and fish stock recruitment. Global Change Biol 24 https://doi.org/10.1111/gcb.13916
Chícharo A, Barbosa AB (2011) Hydrology and biota interactions as driving forces for ecosystem functioning. In: Wolanski E, McLusky D (ed) Treatise on Estuarine and Coastal Science, Vol 10. Academic Press, Waltham, 7–47. https://doi.org/10.1016/B978-0-12-374711-2.01002-0
Clarke A, Johnston NM (1999) Scaling of metabolic rate with body mass and temperature in teleost fish. J Anim Ecol 68:893–905. https://doi.org/10.1046/j.1365-2656.1999.00337.x
Connell JH (1978) Diversity in tropical rain forests and coral reefs. Science 199:1302–1310. https://doi.org/10.1126/science.199.4335.1302
Costa MJ, Vasconcelos R, Costa J, Cabral H (2007) River flow influence on the fish community of the Tagus estuary (Portugal). Hydrobiologia 587:113–123. https://doi.org/10.1007/s10750-007-0690-x
Cowen RK, Lwiza KM, Sponaugle S, Paris CB, Olson DB (2000) Connectivity of marine populations: open or closed? Science 287:857–859. https://doi.org/10.1126/science.287.5454.857
Diaz RJ, Rosenberg R (2008) Spreading dead zones and consequences for marine ecosystems. Science 321:926–929. https://doi.org/10.1126/science.1156401
Dittman AH, Quinn TP (1996) Homing in pacific salmon: Mechanisms and ecological basis. J Exp Biol 199:83–91. https://doi.org/10.1242/jeb.199.1.83
Drinkwater KF, Frank KT (1994) Effects of river regulation and diversion on marine fish and invertebrates. Aquat Conserv: Mar Freshwat Ecosyst 4:135–151. https://doi.org/10.1002/aqc.3270040205
Elliott M, Whitfield AK (2011) Challenging paradigms in estuarine ecology and management. Estuar Coast Shelf Sci 94:306–314. https://doi.org/10.1016/j.ecss.2011.06.016
Emmett RL, Krutzikowsky GK, Bentley P (2006) Abundance and distribution of pelagic piscivorous fishes in the Columbia River plume during spring/early summer 1998–2003: relationship to oceanographic conditions, forage fishes, and juvenile salmonids. Prog Oceanogr 68:1–26. https://doi.org/10.1016/j.pocean.2005.08.001
Epifanio C, Garvine R (2001) Larval transport on the Atlantic continental shelf of North America: a review. Estuar Coast Shelf Sci 52:51–77. https://doi.org/10.1006/ecss.2000.0727
FAO (2011) Review of the State of World Marine Fishery Resources. FAO Fisheries and Aquaculture Technical Paper 569. FAO, Rome. http://www.fao.org/3/i2389e/i2389e.pdf
FAO (2015) FishStatJ: Universal software for fishery statistical time series. FAO Fisheries Division, Statistics and Information Branch. https://www.fao.org/fishery/en/statistics/software/fishstatj/en. Accessed 15 July 2019.
Farmer DM, Freeland HJ (1983) The physical oceanography of fjords. Prog Oceanogr 12:147–219. https://doi.org/10.1016/0079-6611(83)90004-6
FGDC (2012) Coastal and Marine Ecological Classification Standard. Marine and Coastal Spatial Data Subcommittee. FGDC- STD-018–2012. Federal Geographic Data Committee. https://www.fgdc.gov/standards/projects/cmecs-folder/CMECS_Version_06-2012_FINAL.pdf Accessed 21 October 2019.
Froese R, Pauly D (2018) Editors. Fishbase. http://www.fishbase.org/search.php. Accessed 6 Aug 2018
Gillanders BM, Kingsford MJ (2002) Impact of changes in flow of freshwater on estuarine and open coastal habitats and the associated organisms. In: Gibson RN, Barnes M, Atkinson RJ (ed) Oceanography and Marine Biology: An annual review Volume 40, Taylor and Francis, London, pp 233–309. http://books.google.com.au/books?id=VBVGouVOOAIC&pg=PA233
Gillson J (2011) Freshwater flow and fisheries production in estuarine and coastal systems: where a drop of rain is not lost. Rev Fish Sci 19:168–186. https://doi.org/10.1080/10641262.2011.560690
Gracia A (1991) Spawning stock–recruitment relationships of white shrimp in the southwestern Gulf of Mexico. Trans Am Fish Soc 120:519–527. https://doi.org/10.1577/1548-8659(1991)120%3c0519:SSROWS%3e2.3.CO;2
Graham CT, Harrod C (2009) Implications of climate change for the fishes of the British Isles. J Fish Biol 74:1143–1205. https://doi.org/10.1111/j.1095-8649.2009.02180.x
Grimes CB, Kingsford MJ (1996) How do riverine plumes of different sizes influence fish larvae: do they enhance recruitment? Mar Freshwater Res 47:191–208. https://doi.org/10.1071/MF9960191
Guan L, Dower JF, Pepin P (2018) Characterizing spatial structures of larval fish assemblages at multiple scales in relation to environmental heterogeneity in the Strait of Georgia (British Columbia, Canada). Can J Fish Aquat Sci 75:1902–1914. https://doi.org/10.1139/cjfas-2017-0194
Hallett T, Coulson T, Pilkington J, Clutton-Brock T, Pemberton J, Grenfell B (2004) Why large-scale climate indices seem to predict ecological processes better than local weather. Nature 430:71. https://doi.org/10.1038/nature02708
Hamman J, Nijssen B, Roberts A, Craig A, Maslowski W, Osinski R (2017) The coastal streamflow flux in the Regional Arctic System Model. J Geophys Res Oceans 122:1683–1701. https://doi.org/10.1002/2016JC012323
Hare SR, Mantua NJ (2000) Empirical evidence for North Pacific regime shifts in 1977 and 1989. Prog Oceanogr 47:103–145. https://doi.org/10.1016/S0079-6611(00)00033-1
Harrison AJ, Walker AM, Pinder AC, Briand C, Aprahamian MW (2014) A review of glass eel migratory behaviour, sampling techniques and abundance estimates in estuaries: implications for assessing recruitment, local production and exploitation. Rev Fish Biol Fish 24:967–983. https://doi.org/10.1007/s11160-014-9356-8
Hetland, RD, Hsu, T (2013) Freshwater and sediment dispersal in large river plumes. In: Bianchi T, Allison M, Cai W (ed), Biogeochemical Dynamics at Major River-Coastal Interfaces: Linkages with Global Change. Cambridge University Press, Cambridge, 55–8. doi:https://doi.org/10.1017/CBO9781139136853.006
Houde E (1997) Patterns and trends in larval-stage growth and mortality of teleost fish. J Fish Biol 51:52–83. https://doi.org/10.1111/j.1095-8649.1997.tb06093.x
ICOLD (2021) General Synthesis. International Commission on Large Dams. http://www.icold-cigb.net/article/GB/world_register/general_synthesis/general-synthesis. Accessed 15 July 2021
Jackson JB, Kirby MX, Berger WH, Bjorndal KA, Botsford LW, Bourque BJ, Bradbury RH, Cooke R, Erlandson J, Estes JA (2001) Historical overfishing and the recent collapse of coastal ecosystems. Science 293:629–637. https://doi.org/10.1126/science.1059199
Kimmerer WJ (2002) Effects of river flow on abundance of estuarine organisms: physical effects or trophic linkages? Mar Ecol Prog Ser 243:39–55. https://doi.org/10.3354/meps243039
Lehodey P (2001) The pelagic ecosystem of the tropical Pacific Ocean: dynamic spatial modelling and biological consequences of ENSO. Prog Oceanogr 49:439–468. https://doi.org/10.1016/S0079-6611(01)00035-0
Le Pape O, Bonhommeau S (2015) The food limitation hypothesis for juvenile marine fish. Fish Fish 16:373–398. https://doi.org/10.1111/faf.12063
Levin LA, Boesch DF, Covich A, Dahm C, Erséus C, Ewel KC, Kneib RT, Moldenke A, Palmer MA, Snelgrove P (2001) The function of marine critical transition zones and the importance of sediment biodiversity. Ecosystems 4:430–451. https://doi.org/10.1007/s10021-001-0021-4
Loneragan NR (1999) River flows and estuarine ecosystems: implications for coastal fisheries from a review and a case study of the Logan River, southeast Queensland. Austral Ecol 24:431–440. https://doi.org/10.1046/j.1442-9993.1999.00975.x
Lowerre-Barbieri S, DeCelles G, Pepin P, Catalán IA, Muhling B, Erisman B, Cadrin SX, Alós J, Ospina-Alvarez A, Stachura MM (2017) Reproductive resilience: a paradigm shift in understanding spawner-recruit systems in exploited marine fish. Fish Fish 18:285–312. https://doi.org/10.1111/faf.12180
Ludwig W, Dumont E, Meybeck M, Heussner S (2009) River discharges of water and nutrients to the Mediterranean and Black Sea: major drivers for ecosystem changes during past and future decades? Prog Oceanogr 80:199–217. https://doi.org/10.1016/j.pocean.2009.02.001
MacCready P, Geyer WR (2001) Estuarine salt flux through an isohaline surface. J Geophys Res Oceans 106:11629–11637. https://doi.org/10.1029/2001JC900006
MacCready P, Geyer WR (2009) Advances in estuarine physics. Ann Rev Mar Sci 2:35–58. https://doi.org/10.1146/annurev-marine-120308-081015
Martino EJ, Able KW (2003) Fish assemblages across the marine to low salinity transition zone of a temperate estuary. Estuar Coast Shelf Sci 56:969–987. https://doi.org/10.1016/S0272-7714(02)00305-0
Milliman JD, Farnsworth KL (2013) River discharge to the coastal ocean: a global synthesis. Cambridge University Press, UK. https://doi.org/10.1017/CBO9780511781247
Milner NJ, Solomon DJ, Smith GW (2012) The role of river flow in the migration of adult Atlantic salmon, Salmo salar, through estuaries and rivers. Fish Manag Ecol 19:537–547. https://doi.org/10.1111/fme.12011
Moura RL, Amado-Filho GM, Moraes FC, Brasileiro PS, Salomon PS, Mahiques MM, Bastos AC, Almeida MG, Silva JM, Araujo BF (2016) An extensive reef system at the Amazon River mouth. Sci Adv 2:e1501252. https://doi.org/10.1126/sciadv.1501252
Myers RA (1998) When do environment–recruitment correlations work? Rev Fish Biol Fish 8:285–305. https://doi.org/10.1023/A:1008828730759
Myers RA (2001) Stock and recruitment: generalizations about maximum reproductive rate, density dependence, and variability using meta-analytic approaches. ICES J Mar Sci 58:937–951. https://doi.org/10.1006/jmsc.2001.1109
Nunn AD, Tewson LH, Cowx IG (2012) The foraging ecology of larval and juvenile fishes. Rev Fish Biol Fish 22:377–408. https://doi.org/10.1007/s11160-011-9240-8
Olsen Z, Fulford R, Dillon K, Graham W (2014) Trophic role of gulf menhaden Brevoortia patronus examined with carbon and nitrogen stable isotope analysis. Mar Ecol Prog Ser 497:215–227. https://doi.org/10.3354/meps10519
Ottersen G, Bogstad B, Yaragina NA, Stige LC, Vikebø FB, Dalpadado P (2014) A review of early life history dynamics of Barents Sea cod (Gadus morhua). ICES J Mar Sci 71:2064–2087. https://doi.org/10.1093/icesjms/fsu037
Ouellet P, Bui AOV, Lavoie D, Chassé J, Lambert N, Ménard N, Sirois P (2013) Seasonal distribution, abundance, and growth of larval capelin (Mallotus villosus) and the role of the lower estuary (Gulf of St. Lawrence, Canada) as a nursery area. Can J Fish Aquat Sci 70:1508–1530. https://doi.org/10.1139/cjfas-2013-0227
Pauly D (1980) On the interrelationships between natural mortality, growth parameters, and mean environmental temperature in 175 fish stocks. ICES J Mar Sci 39:175–192. https://doi.org/10.1093/icesjms/39.2.175
Pauly D, Alder J, Booth S, Cheung W, Christensen V, Close C, Sumaila U, Swartz W, Tavakolie A, Watson R (2008) Fisheries in large marine ecosystems: descriptions and diagnoses. In: Sherman K, Hempel G (ed) The UNEP large marine ecosystem report: a perspective on changing conditions in LMEs of the World’s Regional Seas. UNEP Regional Seas Reports and Studies No. 182. United Nations Environment Programme, pp 23–40. https://iwlearn.net/resolveuid/848c0f306710edd035e8cfe6e7cd1def
Peck MA, Huebert KB, Llopiz JK (2012) Intrinsic and extrinsic factors driving match–mismatch dynamics during the early life history of marine fishes. Adv Ecol Res 47:177–302. https://doi.org/10.1016/B978-0-12-398315-2.00003-X
Phillips EM, Horne JK, Zamon JE (2017) Predator–prey interactions influenced by a dynamic river plume. Can J Fish Aquat Sci 74:1375–1390. https://doi.org/10.1139/cjfas-2016-0302
Pikitch E, Boersma PD, Boyd I, Conover D, Cury P, Essington T, Heppell S, Houde E, Mangel M, Pauly D (2012) Little fish, big impact: managing a crucial link in ocean food webs. Washington, DC
Pineda J, Hare JA, Sponaugle S (2007) Larval transport and dispersal in the coastal ocean and consequences for population connectivity. Oceanography 20:22–39. https://www.jstor.org/stable/24860094
Planque B, Fromentin J-M, Cury P, Drinkwater KF, Jennings S, Perry RI, Kifani S (2010) How does fishing alter marine populations and ecosystems sensitivity to climate? J Mar Syst 79:403–417. https://doi.org/10.1016/j.jmarsys.2008.12.018
Poff NL, Allan JD, Bain MB, Karr JR, Prestegaard KL, Richter BD, Sparks RE, Stromberg JC (1997) The natural flow regime. Bioscience 47:769–784. https://doi.org/10.2307/1313099
Poff NL, Allan JD, Palmer MA, Hart DD, Richter BD, Arthington AH, Rogers KH, Meyer JL, Stanford JA (2003) River flows and water wars: emerging science for environmental decision making. Front Ecol Env 1:298–306. https://doi.org/10.1890/1540-9295(2003)001[0298:RFAWWE]2.0.CO;2
Poff NL, Richter BD, Arthington AH, Bunn SE, Naiman RJ, Kendy E, Acreman M, Apse C, Bledsoe BP, Freeman MC (2010) The ecological limits of hydrologic alteration (ELOHA): a new framework for developing regional environmental flow standards. Freshwat Biol 55:147–170. https://doi.org/10.1111/j.1365-2427.2009.02204.x
Potter IC, Tweedley JR, Elliott M, Whitfield AK (2015) The ways in which fish use estuaries: a refinement and expansion of the guild approach. Fish Fish 16:230–239. https://doi.org/10.1111/faf.12050
R Core Team (2018) R: A language and environment for statistical computing. R Foundation for Statistical Computing. https://www.R-project.org/.
Reed DC, Harrison JA (2016) Linking nutrient loading and oxygen in the coastal ocean: A new global scale model. Global Biogeochem Cy 30:447–459. https://doi.org/10.1002/2015GB005303
Richter BD (2010) Re-thinking environmental flows: from allocations and reserves to sustainability boundaries. River Res Appl 26:1052–1063. https://doi.org/10.1002/rra.1320
Rijnsdorp AD, Peck MA, Engelhard GH, Möllmann C, Pinnegar JK (2009) Resolving the effect of climate change on fish populations. ICES J Mar Sci 66:1570–1583. https://doi.org/10.1093/icesjms/fsp056
Robins JB, Halliday IA, Staunton-Smith J, Mayer DG, Sellin MJ (2005) Freshwater-flow requirements of estuarine fisheries in tropical Australia: a review of the state of knowledge and application of a suggested approach. Mar Freshwater Res 56:343–360. https://doi.org/10.1071/MF04087
Rulifson RA, Manooch CS III (1990) Recruitment of juvenile striped bass in the Roanoke River, North Carolina, as related to reservoir discharge. N Am J Fish Manage 10:397–407. https://doi.org/10.1577/1548-8675(1990)010%3c0397:ROJSBI%3e2.3.CO;2
Saeck EA, Hadwen WL, Rissik D, O’Brien KR, Burford MA (2013) Flow events drive patterns of phytoplankton distribution along a river–estuary–bay continuum. Mar Freshwater Res 64:655–670. https://doi.org/10.1071/MF12227
Schlacher TA, Skillington AJ, Connolly RM, Robinson W, Gaston TF (2008) Coupling between marine plankton and freshwater flow in the plumes off a small estuary. Int Rev Hydrobiol 93:641–658. https://doi.org/10.1002/iroh.200711050
Schweigert J, Thompson M, Fort C, Hay D, Therriault T, Brown L (2013) Factors linking Pacific herring (Clupea pallasi) productivity and the spring plankton bloom in the Strait of Georgia, British Columbia, Canada. Prog Oceanogr 115:103–110. https://doi.org/10.1016/j.pocean.2013.05.017
Sheaves M, Baker R, Nagelkerken I, Connolly RM (2015) True value of estuarine and coastal nurseries for fish: incorporating complexity and dynamics. Estuar Coast 38:401–414. https://doi.org/10.1007/s12237-014-9846-x
Shoji J, Tanaka M (2008) Recruitment processes of Japanese sea bass in the Chikugo estuary Japan: shift from density-independence to density-dependence during the early life stages. J Northwest Atl Fish Sci 41:85–91. https://doi.org/10.2960/J.v41.m612
Stocker M, Haist V, Fournier D (1985) Environmental variation and recruitment of Pacific herring (Clupea harengus pallasi) in the Strait of Georgia. Can J Fish Aquat Sci 42:174–180. https://doi.org/10.1139/f85-272
Stoecker D, Govoni J (1984) Food selection by young larval gulf menhaden (Brevoortia patronus). Mar Biol 80:299–306. https://doi.org/10.1007/BF00392825
Stoecker DK, Capuzzo JM (1990) Predation on protozoa: its importance to zooplankton. J Plankton Res 12:891–908. https://doi.org/10.1093/plankt/12.5.891
Strauch AM, MacKenzie RA, Tingley RW III (2017) Base flow-driven shifts in tropical stream temperature regimes across a mean annual rainfall gradient. Hydrol Process 31:1678–1689. https://doi.org/10.1002/hyp.11084
Strub PT, Combes V, Shillington FA, Pizarro O (2013) Currents and processes along the eastern boundaries. In: Siedler G, Griffies S, Gould J, Church J (ed) Ocean Circulation and Climate: A 21st Century Perspective. Academic Press, UK, 339–384. https://doi.org/10.1016/B978-0-12-391851-2.00014-3
Szuwalski CS, Thorson JT (2017) Global fishery dynamics are poorly predicted by classical models. Fish Fish 18:1085–1095. https://doi.org/10.1111/faf.12226
Telesh IV, Khlebovich VV (2010) Principal processes within the estuarine salinity gradient: a review. Mar Pollut Bull 61:149–155. https://doi.org/10.1016/j.marpolbul.2010.02.008
Teodosio MA, Paris CB, Wolanski E, Morais P (2016) Biophysical processes leading to the ingress of temperate fish larvae into estuarine nursery areas: A review. Estuar Coast Shelf Sci 183:187–202. https://doi.org/10.1016/j.ecss.2016.10.022
Thompson JR, Gosling SN, Zaherpour J, Laizé CL (2021) Increasing risk of ecological change to major rivers of the world with global warming. Earth’s Future 9:1–20. https://doi.org/10.1029/2021EF002048
Uncles R, Stephens J (2001) The annual cycle of temperature in a temperate estuary and associated heat fluxes to the coastal zone. J Sea Res 46:143–159. https://doi.org/10.1016/S1385-1101(01)00078-8
Van Vliet M, Ludwig F, Zwolsman J, Weedon G, Kabat P (2011) Global river temperatures and sensitivity to atmospheric warming and changes in river flow. Water Resour Res 47. https://doi.org/10.1029/2010WR009198
Vance D, Staples D, Kerr J (1985) Factors affecting year-to-year variation in the catch of banana prawns (Penaeus merguiensis) in the Gulf of Carpentaria, Australia. ICES J Mar Sci 42:83–97. https://doi.org/10.1093/icesjms/42.1.83
Vert-pre KA, Amoroso RO, Jensen OP, Hilborn R (2013) Frequency and intensity of productivity regime shifts in marine fish stocks. Proc Natl Acad Sci 110:1779–1784. https://doi.org/10.1073/pnas.1214879110
Vörösmarty CJ, Sahagian D (2000) Anthropogenic disturbance of the terrestrial water cycle. Bioscience 50:753–765. https://doi.org/10.1641/0006-3568(2000)050[0753:ADOTTW]2.0.CO;2
Watson RA, Green BS, Tracey SR, Farmery A, Pitcher TJ (2016) Provenance of global seafood. Fish Fish 17:585–595. https://doi.org/10.1111/faf.12129
Webb JE, Verspoor E, Aubin-Horth N, Romakkaniemi A, Amiro P (2007) The Atlantic salmon. In: Verspoor E, Stradmeyer L, Nielsen JL (eds) The Atlantic salmon: genetics, conservation and management. Blackwell Publishing, UK, pp 17–56
Whitfield AK, Pattrick P (2015) Habitat type and nursery function for coastal marine fish species, with emphasis on the Eastern Cape region, South Africa. Estuar Coast Shelf Sci 160:49–59. https://doi.org/10.1023/A:1008994405375
Winder M, Cloern JE (2010) The annual cycles of phytoplankton biomass. Philos T R Soc B 365:3215–3226. https://doi.org/10.1098/rstb.2010.0125
Winemiller KO (2005) Life history strategies, population regulation, and implications for fisheries management. Can J Fish Aquat Sci 62:872–885. https://doi.org/10.1139/f05-040
Winemiller KO, Rose KA (1992) Patterns of life-history diversification in North American fishes: implications for population regulation. Can J Fish Aquat Sci 49:2196–2218. https://doi.org/10.1139/f92-242
Acknowledgements
AB was supported by an Australian Government Research Training Program Scholarship and a Northern Australia Environmental Resources/Northern Environmental Sciences Programme scholarship. CJB was supported by a Discovery Early Career Researcher Award (DE160101207) from the Australian Research Council. Finally, we thank the two anonymous reviewers whose comments helped improve the manuscript.
Funding
Open Access funding enabled and organized by CAUL and its Member Institutions. Andrew Broadley was supported by an Australian Government Research Training Program Scholarship and a Northern Australia Environmental Resources/Northern Environmental Sciences Programme scholarship. Christopher Brown was supported by a Discovery Early Career Researcher Award (DE160101207) from the Australian Research Council.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors have no conflicts of interest to declare that are relevant to the content of this article.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Broadley, A., Stewart-Koster, B., Burford, M.A. et al. A global review of the critical link between river flows and productivity in marine fisheries. Rev Fish Biol Fisheries 32, 805–825 (2022). https://doi.org/10.1007/s11160-022-09711-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11160-022-09711-0