
Abstract
Captive rearing is a conservation strategy where juveniles are collected from the natural environment, reared to maturity in a hatchery environment, and then released back into the natural environment at maturity for volitional spawning. This strategy has been used to produce adult outplants for stock enhancement where natural escapement is poor or capture of adults is difficult. In both Idaho (Chinook salmon, Oncorhynchus tshawytscha) and Maine (Atlantic salmon, Salmo salar), captive rearing programs have been initiated as an experimental strategy to prevent cohort collapse and conserve genetic integrity of select depressed populations. In this paper, we provide an overview of these programs and describe some of the methods used to evaluate the effectiveness of this approach. Behaviors such as habitat selection, courting, and spawn timing were monitored. Data collected for both programs indicate that the captive fish display similar behaviors as their wild conspecifics in terms of habitat selection and spawning, although there were some differences in spawn timing. Evaluations of egg and fry production also indicate that captive-reared adults are successfully spawning and producing offspring. Each program is still waiting on final evaluations of reproductive success through genetic analyses of returning adults, but results so far indicate that this could be an additional captive propagation strategy for depressed populations.
Similar content being viewed by others

Introduction
Stocks of wild salmon in both eastern and western North America sustained precipitous declines in abundance in the 20th century (Baum 1997; NRC 1996; Saunders et al. 2006). As a response to these declines, hatchery propagation has been implemented widely on both coasts (Baum 1997; Lichatowich 1999; Saunders et al. 2006). Hatchery programs have been developed both as a means to mitigate for lost abundance by providing hatchery fish for harvest (Mahnken et al. 1998) and as a tool to recover or conserve wild stocks (reviewed in Naish et al. 2007). Many conservation programs using hatchery propagation to boost wild salmon have employed either supplementation or captive broodstocking approaches (Berejikian et al. 2004; Hebdon et al. 2004; Hess et al. 2012). Supplementation approaches commonly collect adults from their habitat and spawn these fish in captivity. Juvenile progeny from hatchery spawned adults are released and returning anadromous adults are either spawned in captivity or allowed to spawn in the natural environment along with wild fish (Cuenco et al. 1993, Wang and Ryman 2001). Captive broodstock programs typically collect adults or juveniles from their habitat and rear them to sexual maturity in the hatchery. Then these adults are spawned in captivity, and their progeny are reared and released at one or more juvenile stages, while a subset of the progeny are kept in captive culture to serve as broodstock and a genetic bank or safety net (Flagg et al. 2004). While these strategies are becoming more widely applied as a conservation tool, there is still uncertainty how well each of these types of conservation hatchery strategies can conserve genetic diversity and fitness, or re-establish self-sustaining populations in the wild (Fraser 2008).
A much less common hatchery conservation program termed captive rearing has been used for a few select populations (Berejikian et al. 1997, 2001a, b). In the captive rearing approach, naturally produced juveniles (eggs, parr, or smolts) are collected, taken into captivity and reared to sexual maturity in the hatchery. However, mature adults are not spawned in captivity, but are released into their natal streams and allowed to spawn naturally (see Kuligowski et al. 2005). Fish are raised in captivity to adulthood to bypass major sources of mortality that may occur from the juvenile to adult stage (Berejikian et al. 2004). The primary objectives of this experimental strategy are to preserve severely depressed populations in the short term while maintaining the long-term genetic integrity of the population at large. If successful, this strategy could prevent cohort collapse in the target populations, maintain a continuum of generation–generation smolt production, and augment depressed natural escapement (or replace it in years when no natural escapement occurs).
Captive-reared fish differ substantially from captive broodstock fish in that natural and sexual selection may play a larger role (Berejikian et al. 2004). By allowing fish to spawn in the natural environment, selection is occurring as the adults are competing for mates and suitable spawning locations. Additionally, selection on eggs and juveniles is also occurring in the natural environment through redd substrates, water temperatures, and possible predation. Captive-reared offspring that have emerged from the gravel may also have better imprinting and homing capabilities than captive brood released smolts (Berejikian et al. 2004; Sweka et al. 2006). But, captive-reared fish undoubtedly experience a different environment and diet in culture and there may be some unintended domestication selection and alteration of spawning behavior and reproductive success as a result.
Captive rearing is considered an experimental approach and it is uncertain as to what behavioral, physiological, or morphological changes the fish may experience in culture. Furthermore, the reproductive success of the outplanted adults and fitness of their progeny in the natural environment is also unknown (Fleming and Gross 1992, 1993; Joyce et al. 1993; Flagg and Mahnken 1995; Carr et al. 2004). There have been some studies that have investigated the spawning behavior of captive-reared coho salmon (Oncorhynchus kisutch) in artificial channels (Berejikian et al. 1997, 1999), Chinook salmon (O. tshawytscha) in artificial channels (Berejikian et al. 2001b), Chinook salmon in natural environments (Chebanov and Riddell 1998; Venditti et al. 2013) and Atlantic salmon (Salmo salar) (Fleming et al. 1996; Carr et al. 2004). Berejikian et al. (2001a) also compared the competitive behavior of male captive-reared and wild coho salmon during spawning, and the differences between newly emerged fry produced by captive-reared and wild coho salmon (Berejikian et al. 1999). These studies all demonstrated that captive-reared fish, despite differences in size and spawn timing, were able to successfully spawn and produce progeny.
In this paper, we review the monitoring and evaluation efforts for two captive rearing programs for salmon on the West and East coasts of North America: Chinook salmon in Idaho and Atlantic salmon in Maine. As captive rearing is an experimental approach, it was unknown as to whether captive Chinook salmon could be reared their entire lives in captivity and then be released as adults and spawn successfully or whether Atlantic Salmon could be spawned in captivity and then released to successfully spawn again. We describe the culture evaluations for Chinook salmon and provide background on the decision-making process for releasing post-spawn captive brood Atlantic salmon adults for spawning. We also describe the critical evaluations used to address spawning behavior and reproductive success in terms of post-release behavior and movement of adults, spawn timing of adults and the number of redds per female. Comparisons are also made to wild con-specifics, where available. Lastly, we highlight some of the main similarities and differences in these captive rearing approaches and thoughts on where this tool is best employed as well as future evaluations needed for both programs.
Chinook salmon in Idaho
Background
Chinook salmon return to spawning areas from May through September. These wild fish must travel 1,200–1,400 km inland, pass eight large hydroelectric dams and 522 km of slack water reservoirs in the Columbia and Snake River corridors (Petrosky and Schaller 2010), to reach natal streams 1,600–2,000 m in elevation (above sea level) in the upper Salmon River drainage. These populations spawn from late July through early October, and like all Chinook salmon are semelparous (i.e.—die after spawning only once); (Healey 1991). Fry emerge from gravels from early April through May, then generally exhibit a stream-type life history, rearing for 1 year in freshwater before migrating to the ocean as age-1 smolts (Gilbert 1913); although juvenile life history can vary (Connor et al. 2001, Copeland and Venditti 2009). Age-1 smolts typically emigrate from early March to mid-May (Copeland and Venditti 2009), then spend one to 4 years in the ocean, before returning to freshwater to spawn as adults. Thus, returning adults are generally described as being 1-ocean (age-3 or Jack), 2-ocean (age-4), or 3-ocean (age-5).
In 1992, Snake River Spring/Summer Chinook salmon were listed as threatened under the Endangered Species Act (ESA; NMFS 1992). Idaho and Oregon state, tribal, and federal fish managers met during 1993 and 1994 to discuss captive culture research and the implementation of strategies to boost abundance of wild populations in the Snake River basin. Flagg and Mahnken (1995) provided an initial literature review of captive rearing technology, which provided the foundation for the program design. Using this work, the Idaho Department of Fish and Game (IDFG) captive rearing program for Salmon River Chinook salmon was initiated to further develop this technology. It was designed to be small in scale and a less invasive rearing technique for populations at risk, since it involved collecting a small number of juveniles or eggs from the natural environment and theoretically it would avoid the impacts of multigenerational hatchery culture described in Reisenbichler and Rubin (1999). This research program was developed cooperatively with state, tribal, and federal fish managers, and used an adaptive management approach through all phases of development of the program.
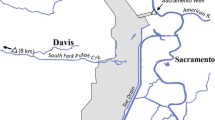
The IDFG captive rearing program was developed to evaluate a new strategy to offset major sources of mortality in the natural environment and increase the number of naturally spawning adults in three select populations in the upper Salmon River drainage: the West Fork Yankee Fork (WFYF), Lemhi River (LEM), and East Fork Salmon River (EFSR) (Fig. 1). These streams were chosen because in 1995 their total wild adult spawner abundances had dropped to record lows of ten, zero, and eleven Chinook salmon in the LEM, WFYF, and EFSR; respectively (NOAA Fisheries Interior Columbia Technical Recovery Team (ICTRT) unpublished data, IDFG unpublished data). These streams were also selected because spawning habitat quality was considered to be relatively pristine with minimal anthropogenic land use practices and disturbances. The IDFG captive rearing program started in 1995 in all three study streams, but the focus of releases and monitoring efforts in each stream changed through time.

Location of Chinook salmon captive rearing program study streams in the Upper Salmon River, Idaho: (BVC), Big Springs Creek (BSC), and the mainstem Lemhi in the Lemhi River (LEM); West Fork Yankee Fork Salmon River (WFYF) in the Yankee Fork Salmon River; and the East Fork Salmon River (EFSR). Captive-reared adults were released upstream of weirs when used, and wild adult returns were trapped at the EFSR trap facility
Methods
Collections and culture
Progeny were collected exclusively for this captive rearing strategy. At the onset of the program, Chinook salmon had never been reared from juvenile to the adult stage so it was uncertain as to how well fish would survive in captivity. The objective for collection of wild Chinook salmon for the captive rearing experiment was to collect enough juveniles or eggs from the target populations to provide approximately 20 adults to release to spawn each year in each study stream (Hassemer et al. 1999). The survival of fish to maturity in culture was unknown and this was merely a minimum goal. Collections were cautiously employed since the loss of wild production with unknown benefits from culture could lead to further losses. The program initially collected wild Chinook salmon parr using rotary screw traps and beach seines to comprise annual rearing groups. A maximum of 25 % of the parr produced in each stream or a maximum of 200 parr was permitted for collection (section 10, permit 43230) in order to not over-mine the natural population. From 1994 through 1999, parr were collected from all three study streams (LEM, WFYF, and EFSR). Collected parr were temporarily held in streamside incubation boxes for a couple of hours until transport to the hatchery for rearing.
In the later years of the program, progeny were collected as eyed-eggs instead of parr. From 2000–2005, eyed-eggs were collected from natural spawning wild Chinook salmon but only in the WFYF and EFSR (Berejikian et al. 2011). Eyed-egg collections were made by monitoring embryo development by tracking cumulative Celsius thermal units (CTUs) from the observed or estimated date of spawning of wild adults. When eggs reached the eyed-stage of development (estimated), redds were sampled using hydraulic methods as described by Berejikian et al. 2011). Caution was exercised to not damage the collected eggs or eggs remaining in the redd. In both the EFSR and WFYF, approximately 300 eyed-eggs were targeted for collection from approximately six wild redds per stream. The number of targeted redds and eggs was also chosen to be conservative to not over-mine the population or impact the remaining eggs within each redd. In order to evaluate the success of each collection method, the number of fish surviving to the smolt stage and to maturity were summarized for each collection method and statistically analyzed with a z-test for differences in proportions (α = 0.05).
One of the main objectives of the program was to develop and implement culture practices needed to rear Chinook salmon to maturity in captivity. Two hatchery facilities, the IDFG Eagle Fish Hatchery (EFH) in Idaho and NOAA Manchester Research Station (MRS) in Washington, were used for rearing of Chinook salmon. Fish were reared using standardized fish culture practices in freshwater at EFH; and in seawater at the MRS facility (Baker et al. 2006; Maynard et al. 2012). During early rearing (first 15 months at EFH), juveniles were maintained in family groups by redd, then tagged with Passive Integrated Transponder (PIT) tags, then combined in larger rearing groups while maintaining segregation between stocks from each stream. When fish reached the smolt stage (age 1+) they were vaccinated and allowed to recover. During the time wild-origin smolts were migrating downstream through the Salmon, Snake, and Columbia Rivers to the Pacific Ocean, fish were transferred to MRS for rearing in saltwater. However, about 20 % of each cohort from brood years 1994–1999 were left in freshwater at EFH for the remainder of their life cycle and spawned as broodstock to (1) conduct gamete evaluations, (2) provide a measure of protection if no wild adults were forecasted to return to the study areas, and (3) provide a safety-net against possible catastrophic loss at MRS (Venditti et al. 2013). Also, in some years a portion of mature age-2 males (precocials) were culled, and their sperm was cryopreserved for future spawning events. Eyed-eggs from these spawning events would be placed into egg-boxes and out-planted into the study streams but are not a focus of the evaluations described here. All fish from brood years 2000–2005 were reared in seawater at MRS from the smolt stage through maturation (Maynard et al. 2012).
While in culture, the maturation status was determined for each fish and tracked for each brood year as well as length and weight (growth rates) and survival (as mentioned above). For years 1997–2001, the maturation status of each fish was determined at MRS in April using visual inspection. From 2002 to 2010, ultrasound examinations were conducted using an Aloka SSD-500 V ultrasound unit and an Aloka Electronic Linear Probe UST-556L-7.5 (Aloka Co., Ltd., Tokyo, Japan) to separate maturing fish from non-maturing fish and place the immature fish back on feed as quickly as possible (Maynard et al. 2007). Fish were reared in these facilities up to 1 month prior to spawning.
Adult releases
The number of fish released was primarily based upon the number of maturing adults in captivity, but in the early years of the program also based in part on expected adult returns to the study streams. Maturing captive Chinook salmon destined for release were fitted with either a Peterson disc tag or spaghetti tag prior to release. Tags were color-coded to identify the brood year and aid in the visual identification of adults for post-release field evaluations.
Mature fish from multiple brood years were pooled by stock for transport to Idaho, and tanks were loaded with approximately 1/4 seawater and 3/4 freshwater (by volume) to begin freshwater acclimation during transport. Once in Idaho, mature captive Chinook salmon were either acclimated in freshwater at EFH or directly released into study sections from early to mid-August in early years (1997–2005), and in early July in later release years (2006–2010). Fish were then released with the aid of a helicopter (distant or inaccessible release sites) in some years or transferred on foot.
Temporary picket weirs were assembled at the downstream end of the study section of each stream (Fig. 1), consistent with three evaluation periods, on the maintstem LEM or tributaries of the LEM (1998–2000), on the WFYF (2001–2003), and the EFSR (2004–2008). In the LEM, all wild Chinook salmon parr were collected from the mainstem LEM, but captive adults were released both into the mainstem (1998 and 200) and two LEM tributaries. Picket weirs not only ensured captive fish did not stray outside of their stock-specific spawning areas, but also remained in study sections where they were readily observed as part of both intensive behavioral surveys and condensed spawner surveys. Trap boxes built into the temporary weirs were used to pass wild Chinook salmon and other species in either direction. Captive fish attempting to move out of the study area were returned to the blocked sections. This prevented some mixture of spawning of captive adults with wild adults in most years, except for years and streams when a picket weir was not utilized.
For each release year, we summarized the age and gender composition of the release group, and we performed several analyses of length at age of captive adults. Mean lengths of released captive adults were calculated for each age (2–5) for each study stream by gender across all years. All mainstem LEM and LEM tributary releases were pooled into a single LEM group for analyses of length at age. Then, for each gender-age group; the following statistics were calculated: minimum, maximum, median, and first and third quartiles. Comparisons of length at age were shown utilizing box and whisker plots containing all the above statistics. Lastly, across all streams and release years, we performed one-tailed Student’s t tests, assuming unequal variance, to determine statistical differences in mean length for each age (ag-3, age-4, and age-5) between females and males; and then between age-4 and age-5 fish.
Length at age of captive adult releases were also compared to length at age of their wild counterparts utilizing data from the adult trap facility on the EFSR, which captures wild adult returns. These comparisons were only available from 2005 to 2009 data, when wild adult lengths were readily available from trapping records concurrently with an appreciable number of captive fish released. A total of 636 lengths were collected from wild Chinook salmon captured at the EFSR adult trap facility in 2005–2009 (63 in 2005, 81 in 2006, 90 in 2007, 207 in 2008, and 194 in 2009). Gender of wild adults was determined by visual inspection of phenotypic traits at the time of capture and ages later assigned from an age at length key developed from fin ray aging of wild adults (Kennedy et al. 2011). The same means and statistics were calculated for all groups and again compared via box and whisker plots as described above. Two-tailed Student’s t tests were performed, assuming unequal variance, to determine statistical differences in length at age between captive and wild fish for each age (age-3, age-4, and age-5).
Spawn behavior
After the release of captive Chinook salmon adults, the main goal was to evaluate whether released adults had the potential to successfully reproduce. Our first objective in evaluating this potential was to quantify captive Chinook salmon habitat use and spawning behavior. The results of the intensive surveys in the WFYF during 2001–2003 have been previously summarized and published by Venditti et al. (2013). In the WFYF, blocking weirs were used to constrain captive releases to smaller study sections in which behavioral observations and spawning success could be more effectively assessed. Habitat use was then quantified by noting whether fish occupied pool, rifle/run, cut bank, and overhead vegetation habitats. Then, spawn behavior was quantified through observations of holding, aggression, courting, moving, and milling; see Venditti et al. (2013) for additional details. Lastly, captive and wild origin fish were distinguished by the presence or absence, respectively, of a Peterson disc or spaghetti tag. Concurrently, these intensive evaluations on the WFYF also recorded the number of redds produced, the spatial and temporal distributions of both wild and captive fish, and the incidence of mating between captive-reared and wild fish.
Reproductive success
Another post-release objective was to estimate captive adult spawning success. Thus, spawner surveys were conducted from mid-July to late September to evaluate spawning success of captive adults released into the LEM and its tributaries (1998–2000), the WFYF (1998, 2001–2010) and EFSR (1999, 2004–2010). These spawner surveys recorded only completed redds, and were condensed relative to the intensive surveys in the WFYF from 2001–2003. Surveys were conducted daily to every third day on a given study section. Completed redds were flagged along the stream bank and fish origin (wild or captive), GPS location, and date of completion were recorded (Stark and Richardson 2011). As in behavioral surveys, captive and wild origin fish were distinguished by the presence or absence, respectively, of a Peterson disc or spaghetti tag. Since fish origin was readily discernible, the incidence of matings (redds) between captive and wild fish was also recorded. Detailed spawn timing data of both captive and wild fish was collected in both the EFSR and WFYF, but only from 2006 to 2010. Reproductive success of released adult captive Chinook salmon was then evaluated based upon the number of redds per female. This measure was calculated by simply dividing the number of captive redds produced by the number of captive adult females released in a given year. Redds were compiled by origin and their temporal (2006–2010) distribution documented.
During 2001–2002 in the WFYF, Venditti et al. (2013) also conducted additional post-spawn evaluations of reproductive success to verify egg deposition, estimate embryo survival to organogenesis, and estimate potential contribution of eyed eggs to the population. For these evaluations, eggs were sampled hydraulically, as described above in collections and rearing, and collected after they had received a minimum of 200 CTU. Eggs were then categorized as either dead or viable, and if viable either fertilized or unfertilized. Lastly, they estimated the number of eyed eggs produced by captive females by multiplying the number of captive redds times the mean egg fertilization rate times an adjusted mean captive female fecundity, see Venditti et al. (2013) for complete methods.
Results
Collections and culture
A total of 2,398 parr were collected from the LEM, WFYF, and EFSR and reared in captivity to adults (Table 1). Survival of parr in captive culture from the time of collection to smolt stage averaged 86.9 % (CI 85.5–88.2 %) for all streams combined; and was 87.8 % (CI 85.4–89.8 %), 88.0 % (CI 85.6–90.1 %), and 84.5 % (CI 81.7–87.0 %) for the LEM, WFYF, and EFSR, respectively (Fig. 2). Survival of parr collections to mature adults averaged 39.7 % (CI 37.8–41.7 %); and was 43.0 % (CI 40.1–46.7 %), 43.7 % (CI 40.3–47.2 %), and 31.3 % (CI 28.0–34.7 %) for the LEM, WFYF, and EFSR, respectively. Drawbacks associated with sourcing parr as rearing groups included: (1) difficulty converting parr from wild diets to hatchery diets, (2) high prevalence of bacterial kidney disease in collected juveniles, (3) and high prevalence of parr carrying parasitic gill copepods and systemic bacteria (Hassemer et al. 1999; Johnson and Heindel 2001) which reduced their overall survival in captivity.

Captive-reared Chinook salmon survival from collection to smolt (a) and collection to maturation (b) for the Lemhi River (LEM), West Fork Yankee Fork Salmon River (WFYF), and East Fork Salmon River (EFSR). Rearing groups were collected as either wild parr (WP) from 1994 to 1999, or wild egg (WE) 2000–2005; with the dashed line representing this change in collection method
A total of 4,867 eyed-eggs were collected from the LEM, WFYF, and EFSR and reared in captivity to adults (Table 1). Survival of eggs in captive culture from the time of collection to smolt stage averaged 91.5 % (CI 90.7–92.2 %) for all streams combined; and was 86.4 % (CI 81.5–90.1 %), 90.7 % (CI 89.4–91.9 %) and 92.7 % (CI 91.6–93.6 %), for the LEM, WFYF and EFSR, respectively (Fig. 2). Survival of egg collections to mature adults averaged 60.3 % (CI 59.0–61.7 %); and was 32.6 % (CI 27.2–38.5 %), 63.1 % (CI 61.1–65.2 %), and 60.9 % (CI 59.0–62.8 %) for the LEM, WFYF, and EFSR, respectively. Survival of captive fish from collection to smolt was slightly higher for fish collected as eyed-eggs (91.5 %, n = 4,867 eggs, n = 4,452 smolts) than fish collected as parr (86.9 %, n = 2,398 parr, n = 2,083 smolts), (z = −6.15, p > 0.0002). However, mean survival from collection to maturity was dramatically higher for fish collected as eyed-eggs (60.3 %, n = 4,867 eggs, n = 2,937 mature adults) than fish collected as parr (39.7 %, n = 2,398 parr, n = 952 mature adults); (z = −16.59, p < 0.0002). In addition to providing a hedge against introducing disease in the program, eyed-egg collections provided the additional advantage of knowing the approximate number of families that contributed to the development of annual rearing groups (although the degree of polygyny and polyandry was unknown).
EFH in coordination with the MRS was able to successfully rear Chinook salmon, collected from the wild, to maturity in captive culture. Survival of captive fish from collection to maturation generally increased with each successive brood year (Fig. 2), as improvements were made in culture methods (Heindel et al. 2005). However, despite better survival over time, in nearly every brood year age 2 fish comprised a substantial portion of mature fish and the rate of precocity did not lessen over time (Table 1). It is important to note that all of these early maturing fish were precocial males.
Adult releases
From 1998 to 2010, IDFG has released 2,819 adult captive Chinook salmon across three Idaho study drainages, from as few as 20 (2010) when the program was winding down to as many as 573 (2007) fish per year, when the program reached its peak (Table 2). In 2002, the program was able to release substantially more adults than in prior years, and greater release numbers were sustained for several years thereafter. During the late 1990s and early 2000s, due to forecasted low adult returns to particular streams in a given year, fish were not released but spawned in the hatchery as a safety-net (LEM: 2001–2002; WFYF: 1999–2000; EFSR: 1998, 2000–2001). Overall, survival of fish to maturity (not all mature fish were released) was 53.5 % (CI 52.4–54.7 %). Post-release adult survival was not estimable, and could only be inferred as high from spawning surveys. The age of released adults varied considerably over the years. Early release years were dominated by younger age fish, predominantly precocial males; while later releases consisted of more age 4 and 5 fish, predominately females (Table 3). The sex ratio of released adults also varied considerably over the years (Table 3).
Analyses of length at age of released captive adults revealed several findings. Lengths of released females were consistently larger than males of the same age; age-3 (t = −1.92, df = 39, p < 0.03), age-4 (t = −5.62, df = 210, p < 0.0001), and age-5 (t = −2.21, df = 21, p < 0.02); (Fig. 3). We also found that age-5 females attained sizes (e.g. lengths) significantly larger than age-4 females (t = −3.00, df = 172, p < 0.002), but age-5 males were not significantly larger than age-4 males (t = −0.37, df = 20, p = 0.36). This suggests that growth rates reached a plateau in captive culture from age 4 to age 5 males. Lastly, in the EFSR from 2005 to 2009 captive fish averaged 13.4, 23.1, and 41.6 cm (FL) shorter than wild fish at age-3, age-4, and age-5; respectively (Fig. 4). Furthermore, these differences in mean length at age between captive fish and their wild counterparts were statistically significant at p < 0.0001 for age-3 (t = −21.36, df = 204), age-4 (t = −56.64, df = 783), and age-5 (t = −29.94, df = 29).

Length at age (cm FL) of male and female adult captive-reared Chinook salmon released into the Lemhi River (LEM), West Fork Yankee Fork Salmon River (WFYF), and the East Fork Salmon River (EFSR), averaged across all years. Median lengths are represented by the middle lines, means by diamonds, first and third quartiles by the boxes, minimum and maximum lengths by the whiskers, and samples sizes of each respective age above each maximum. A total of 101 fish were either not measured (n = 66) or their gender undetermined (n = 35) pre-release

Spawn timing (redd completion) of captive-reared and wild Chinook salmon in the East Fork Salmon River (EFSR) and West Fork Yankee Fork Salmon River (WFYF), 2006–2009
Spawn behavior
From intensive spawning surveys conducted in the WFYF (2001–2003), Venditti et al. (2013) identified that captive fish behaved very similarly to their wild counterparts; but, captive males were less aggressive than natural males. Captive fish were found to show similar habitat selection during redd construction as wild fish, given release section constraints (Venditti et al. 2013). With regard to courtship behavior, captive males were found to display similar behavior as wild males as expressed by frequency of quivers; before, during, and after spawning. Additionally, captive females exhibited similar spawning behavior as wild females as expressed by digging frequency (Venditti et al. 2003). Overall, both male and female captive Chinook salmon released to spawn naturally demonstrated all of the courtship and spawning behaviors wild Chinook salmon exhibited (Venditti et al. 2013).
Reproductive success
While captive fish were generally smaller in size, captive adults spawned successfully in the wild and produced viable eggs and offspring. Redds constructed by captive Chinook salmon were generally distributed throughout areas where fish were able to access. The number of redds per female was highly variable both across streams and years (Table 2). The mean number of redds per female was highest in Bear Valley Creek (BVC) and lowest in the WFYF. The most redds produced in a given year was in the EFSR in 2007, when a total of 313 fish were released (200 males, 113 females). Over all streams and years, a total of 343 redds were produced by captive Chinook salmon adults released to spawn in natal streams (Table 2). With a total of 1,343 known captive females released into all three study streams, an average of 0.29 redds were produced per captive female released. In comparison, wild Chinook salmon generally achieve one redd per female (Murdoch et al. 2009, Gallagher and Gallagher 2005, Neilson and Bradford 1983). The number of redds per female was likely influenced by habitat quality of the study streams, with spawning habitat generally the best in the LEM drainage, intermediate in the EFSR, and poorest in the WFYF. The WFYF was especially degraded through excessive siltation following the ‘Potato’ wildland fire of 2006. Also, some captive x wild and wild x captive redds were observed in years when a picket weir was not installed, and both origins were allowed to intersperse.
Surveys of spawn timing in the EFSR and WFYF during 2006–2009, found captive spawn timing was generally delayed relative to wild Chinook salmon (Fig. 4). On average, redd construction by captive adults started in late August/early September with peak spawning occurring in mid-September and lasting through late September (but this was highly variable both within and across streams and years). Peak spawning for wild Chinook salmon was generally 2–3 weeks in advance of peak captive spawning. However, spawn timing by captive Chinook salmon in the WFYF was earlier and less delayed, relative to wild Chinook salmon spawn timing (Fig. 4). Not coincidently, during these same years (2006–2009) on the WFYF, captive Chinook salmon were not isolated from wild fish via a temporary weir. Furthermore, in the EFSR after the picket weir was lost in a high water event and many captive fish escaped below (2008) and when a picket weir was not installed (2009); captive spawning was also less delayed and overlapped considerably with wild Chinook salmon spawning (Stark et al. 2009; Stark and Gable 2010).

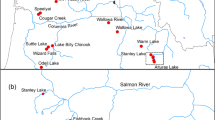
Location of drainages in the Gulf of Maine Distinct Population Segment for endangered Atlantic salmon, Maine
Additional post-spawn evaluations by Venditti et al. (2013) in the WFYF in 2001–2002, found eggs in 22 out of 26 captive redds. In addition, mean egg viability was 54.7 % in 2001 and 34.6 % in 2002, and from this potential production by captive females in the WFYF was 7,500 eggs in 2001 and 11,500 eggs in 2002, for a total of 19,000 eyed-eggs to the population (Venditti et al. 2013). These preliminary investigations have indicated that captive fish may be producing returning adults.
Atlantic salmon in Maine
Background
Maine Atlantic salmon are iteroparous and on average have a 5 year life cycle (Fay et al. 2006). Adults migrate into their rivers of origin between May and October and will hold in deep pools or dead water habitat (Fay et al. 2006). Spawning typically begins in mid—October and lasts through mid—November. Fry begin to emerge in mid—May and move into rearing habitats consisting of cobble boulder substrates (Fay et al. 2006). Typically juvenile Atlantic salmon spend 2 years in riffle run habitat before smolting and migrating to the ocean. Then they spend 2 years in the ocean and migrate as far north as the West Greenland Sea before returning to their natal rivers (Danie et al. 1984; Baum 1997; Fay et al. 2006).
Atlantic salmon within the Gulf of Maine (GOM) Distinct Population Segment (DPS); (Fig. 5) were first listed as endangered under the U.S. Endangered Species Act in 2000 [ESA; National Marine Fisheries Service (NMFS); National Oceanic Atmospheric Administration (NOAA); U.S. Fish and Wildlife Service (USFWS), 2000]. Originally the DPS included eight populations found in the Dennys, East Machias, Machias, Pleasant, Narraguagus, Sheepscot, and Ducktrap, and Cove Brook drainages (Fig. 5). In 2009, the DPS was expanded to include the Penobscot, Kennebec, and Androscoggin River drainages. Current goals for GOM Atlantic salmon are preventing extinction and preserving the current metapopulations structure throughout the DPS (Bartron et al. 2006). Federal (NOAA Fisheries, USFWS), State of Maine (Department of Marine Resources), and Penobscot Indian Nation fisheries biologists collaborate to adaptively manage the Gulf of Maine DPS of Atlantic salmon with the assistance from non-government organizations (NGO’s) and other groups.
GOM Atlantic salmon are adaptively managed and several life stages have been historically cultured and released over the last 100 years to enhance DPS river-specific stocks including unfed fry, age-0 +parr, and smolts. Since 1990 the primary hatchery release strategy from captive broodstock production has been outplanting un-fed fry during May and June. While good at producing large numbers of juveniles and maintaining river specific brood numbers and genetic diversity, the smolt-to-adult return (SAR) rates from fry origin smolts releases has been low (0.72–2.03 % USASAC 2012). Age-0 +parr releases have also been attempted but hatchery capacity limits, and return rates for SARs from parr origin smolts is even lower than fry (0.0286–0.0574 %); (USASAC 2012). Smolts releases have been used since the 1970s in the Penobscot River (USASAC 2012) to enhance populations and they produce over 85 % of all returns to the GOM DPS (USASAC 2012). However, when smolt stocking releases are discontinued, the number of returning adult spawners diminishes (USASAC Database), suggesting that smolt releases will not succeed in restoring self-sustaining runs and smolts require additional costs and rearing space above the other release strategies.
The release of captive-reared, gravid adults in Maine came about from a desire to make effective use of fish deemed excess to broodstock spawning needs, hatchery capacity, and to provide for opportunities for wild exposure and natural selection of progeny. Another similar attempt used sea-caged reared adults and released them into their river of origin. In sea-cage rearing, eggs are hatched and reared to smolt stage in freshwater facilities and then transported to cages moored inshore along the coast of Maine where the salmon are grown to maturity. Experimental outplanting of sea-cage reared adults occurred as early as 1996, but results were difficult to quantify. These early releases suggested that these adults built redds but emergence was low (Mackey and Atkinson 2003). However, evaluation of these studies was incomplete. It was unclear as to the reason for low success and possible contributors could be poor gamete viability or other factors such as handling stress, late outplanting timing or culture methods not evaluated during the time period (Mackey and Brown 2003). Despite the low success with sea-cage reared salmon, Sweka et al. (2006; USASAC 2009) hypothesized that utilizing either planted eggs or captive adults would be the best option for ensuring that parr re-captured for broodstock have experienced the longest period of freshwater selection and thus would have a better chance of returning adults. Therefore, the Maine Atlantic Salmon Technical Advisory committee agreed to adopt the use of captive adult releases into their rivers of origin within suitable habitat as an additional management strategy and encouraged managers to investigate their potential to produce salmon that were better equipped for life in the wild (Trial et al. 2010).
Maine has proactively released captive adults from 2005 to the present. The majority of releases occurred in eastern Maine; a region characterized by forested uplands and sandy barrens in the lowlands. Drainages include the Hobart, Union, Machias, East Machias, and Dennys Rivers (Fig. 5). Captive releases also occurred in southern Maine in the Sheepscot River. There were two objectives for implementing this strategy: (1) examine spawning behavior, reproductive success, and long term viability and (2) use that information to determine if this would be a better strategy than outplanting un-fed fry. In this paper, we describe the evaluations taken to address these goals from 2005 to 2012.
Methods
Collections and culture
Since 1990, Atlantic salmon captive broodstocks have been managed on a river specific basis to maintain genetic divergence among drainages. Two USFWS hatchery facilities, (CBNFH); (East Orland, ME) and Green Lake National Fish Hatchery (GLNFH); (Ellsworth, ME) are used for propagation and rearing of GOM Atlantic salmon, with river stocks segregated within each facility. For the six stocks released in this study, due to low numbers of wild returns, large parr (1+ or older) are captured via electrofishing from August through October for broodstock, and taken to CBNFH. Here they are raised to adult in freshwater to be primarily spawned for fry production (Bartron et al. 2006).
Collection of juvenile Atlantic salmon for broodstock purposes is guided by two primary objectives: (1) provide a river-specific source for future juvenile stocking events in DPS rivers, and (2) collect hatchery-origin and natural-origin parr from DPS rivers in a manner that represents the genetic variability in those rivers. River-specific broodstock parr collection goals, based on available habitat for fry releases, are set in advance of collections. The number of parr kept for captive broodstocks in each drainage is targeted at 150–250 parr per drainage annually (Bartron et al. 2006; Table 4). In order to target parr collections from natural spawning events, data on the distribution of redds from 2 years previous are used to identify areas where natural reproduction has taken place. Collection locations and target numbers are selected for all accessible river sections where natural reproduction has occurred. Two or more collection locations are typically selected within 1,000 meters of known spawning habitat to account for fry and parr dispersal. In addition, electrofishing sites were selected to recover hatchery-origin parr based on the quantity and quality of riverine habitat. Hatchery-produced fry are stocked into river sections of suitable habitat, and collection efforts focus on these areas to recapture those stocked individuals as parr and incorporate some level of wild exposure into the broodstock through this “river-ranching” strategy. Collection efforts are made in multiple sections throughout each river system. Parr collected for broodstock are generally age-1, although age-2 parr are often captured. On rare occasions, sampling efforts may elect to collect age-0 parr, especially if there is reason to expect difficulties in capturing many parr in the following year.
The collection of parr is not without risks and disease outbreaks are a serious concern. Prospective broodstock from each river are either transported directly to CBNFH following collection, or held for several days in an oxygenated tank at the Maine Department of Marine Resources (DMR) Jonesboro headquarters (due to distance from hatchery), then transferred to CBNFH. The short-term survival for both acclimation methods is approximately 100 % for the captured parr (Bartron et al. 2006). At CBNFH, the parr are brought to the hatchery’s receiving building, and within 24 h receive a prophylactic 250 ppm formalin treatment for 1 h. After the treatment, parr from each river are placed in a fish rearing isolation unit for a quarantine period of 10–12 months. During this time period, fish are observed for abnormal behavior (such as due to disease), and are non-lethally sampled for bacterial pathogens at the 10 % assumed incidence level, using inoculation and culture from vents. After the quarantine period, each fish is implanted with a uniquely coded passive integrated transponder (PIT) tag and a fin clip is taken for genetic analysis. PIT tags facilitate the identification of an individual to river of origin, available genetic data, and spawning history. All tagging information including PIT tag codes, marking date, and capture date are entered into the CBNFH broodstock database and are used in conjunction with genetic pedigree (or relatedness) data to later identify spawning contributions of each fish for the captive broodstock (Bartron et al. 2006).
Adult releases
Annually, there are sexually mature adult salmon that are not essential to the genetic and demographic egg production needs at CBNFH. These adults are defined as individuals that have previously spawned, and are not needed or prioritized for additional spawn crosses in the current year (Bartron et al. 2006). A smaller proportion of the releases can be from maiden fish (not spawned) but also not essential for captive broodstock production needs. As part of the restoration strategy for the GOM DPS, the use of these adults is intended to increase the amount of natural selection exposed to the populations during their life cycle, increase the reproductive potential of these individuals, and restore a historical spawning component to the natural ecosystem (Bartron et al. 2006). To be able to use these fish for targeted restoration actions as spawners in the wild, they must be identified at the hatchery in a timely manner that is also biologically relevant (evidence of sexual maturation is observed just prior to spawning). These fish are identified as sexually mature typically by mid-October, and only at that point are numbers available to managers to determine stocking potential through evaluation of previous genetic contribution, estimates of the number of first-time spawners, and comparison of spawning potential to egg and fry production requests.
Release numbers were not only determined by available fish but Conservation Spawning Escapement (CSE) indices were also used to guide release targets. CSE is calculated as the number of females needed for an egg deposition of 2.4 eggs per m2 of rearing habitat divided into an average fecundity of 7,600 eggs per female then multiplied by 2 to include males (Eq. 1); (Baum and Meister 1971; Beland 1996). Early in the program (2005–2008), a CSE of 1.0 was targeted (e.g. the number of female and male salmon at a 1:1 ratio needed for replacement. Later, spawner escapements of 1.5–2 times CSE were targeted to exceed replacement as calculated by available habitat in outplantings (Baum and Meister 1971).
The timing of releases varied over the 7 years from early to late October to coincide with the timing of wild Atlantic salmon spawning. From 2005 to 2009 adults were released into habitat reaches that were segregated by geologic features to minimize adverse interactions with wild adults even if it meant lower quality spawning and rearing habitats. For example, reaches in the upper Mopang Stream, a tributary of the Machias River, had downstream barriers such as beaver dams which contained fish for easier monitoring and observation while also adding assurance that the fish did not stray out of the spawning areas. In 2009, adults were placed into higher quality reaches of contiguous habitat (namely the Northern Stream, a tributary of East Machias River) with no barriers to movement. This shift from segregated habitat to open habitat came about from positive results in juvenile production in the more segregated reaches and an attempt to compare wild production more equally to this strategy. This change was applied over several drainages but in Northern Stream, a more thorough study design was outlined to examine movement, timing of spawning, distribution, and reproductive success as described below.
Spawn behavior
Throughout the period of Atlantic salmon captive adult releases (2005–2011), spawner surveys were conducted immediately after release, either on foot or by canoe. Redd locations were recorded with a handheld GPS. Spawning structures were defined either as a test pit, where the redd was not complete or was abandoned, or as a completed redd, as defined by Burner (1951). At least one complete survey of all streams with captive adults was conducted after spawning activities ended but weekly surveys were conducted in a selected reach of the Northern Stream to better quantify and timing and duration of spawning for outplanted adults. Annual spawner assessments were also conducted in a wild index tributary to the Machias River (the Old Stream) in 2009–2010 to document the timing and duration of wild spawning. Then, these data are used to compare spawn timing between wild and captive fish in the Old Stream and Northern Stream, respectively during 2009–2010.
To address other aspects of spawning behavior besides timing, several methods were employed to address the movement and distribution of released adults. The objectives of this evaluation were to document the behavior of fish post-release and whether they stayed within the spawning areas. Upstream and downstream dispersal from the release location was summarized from 2005 to 2011 simply by documenting redd locations in relation to the release point.
However, due to uncertainties of spawner fidelity to the release reach and possible wild spawners influencing results, more definitive methods were needed to document fidelity and spawning events from released adults. In 2010, an acoustic tracking network was deployed in Northern Stream and the greater East Machias Drainage (Fig. 6). Twenty-one receivers were placed throughout the Northern Stream and the greater East Machias drainage (Fig. 6). In this year, 40 age-5 repeat spawners were released at 1:1 female to male ratio. Due to the number of tags available, only 10 of the 20 released males but all 20 of the released females were tagged with VEMCO V16 acoustic tags. Their movements were then tracked with VR2 VEMCO acoustic receivers (Bruchs 2012). In addition, all 40 adults were tagged with Carlin dangler tags differentially colored by sex. All tagging activities took place at the CBNFH a week prior to release. These fish were then released the first week in October. Survival of adults from release to early May of 2011 were estimated based on number of acoustically tagged adults surviving to this time point. In the fall of 2011 and 2012, the acoustic telemetry equipment was no longer available, so a single PIT array consisting of two antennas was installed in the Northern Stream to track movements into and out of the stream, determine the spawning ratio of adults, and obtain site fidelity data. Fish were uniquely tagged with Biomark 32 mm 1/2 duplex PIT tags stream-side prior to release (Bruchs 2012; Bruchs and Atkinson 2013). The number of adults present and spawning ratio were determined from known fish still determined to be in the Northern Stream at the end of November, when the array was removed. Estimating survival rates was not deemed possible with this type of antennae since it was a single transect and active surveys were not feasible.

Illustration of the Northern Stream study location within the East Machias River, Maine with location of the 2010 acoustic receiver array. Northern Stream is shaded in grey. Additionally, location of the passive integrated transponder (PIT) receiver antennae used in 2011–2012 and the location for adult Atlantic salmon releases for all years (2010–2012) are noted
Reproductive success
As stated above, redd surveys were performed from 2005–2011 to document the distribution of fish but also as way to quantify reproductive success. The number of redds per female released was based on the number of observed redds in a reach divided by the known number of females released into that reach. The exception was the Northern Stream, where redds per female was calculated from either the acoustic receiver data (October 2010–May 2011) or the PIT array data (2011 and 2012). Then, the redds per female ratio was regressed against release date, for all streams combined. The number of redds per female was also analyzed for all age 5 repeat spawners compared to the locations where a mix of age 3, 4 and 5 females were released by regressing the proportion of age 5 females outplanted in a stream against the redd per female ratio in those streams.
The emergence timing of fry produced from captive adults released to spawn naturally was also evaluated in the spring of 2005 in the Mopang Stream a tributary of the Machias River, and in 2009 in the Sheepscot River. First, in reaches where emergence was to be evaluated, redds were marked with white landscaping rocks in a cross pattern for completed redds and in a line parallel to flow for a test pit during the previous fall while conducting spawner surveys. Then, emergence traps were placed over completed redds produced by captive adults (Phillips and Koski 1969; Mackenzie and Moring 1988). Since a complete egg count would be impossible due to intragravel movement, the focus was on timing and relative abundance instead of intragravel survival. Emergence nets consisted of fine mesh nylon netting stretched over PVC pipe frames with a cod end located downstream connected to a trap box by a 3.81 cm inside diameter PVC pipe. Emergent fry were collected in the trap box, and enumerated. Traps were monitored daily; and fry counts, water temperature (Celsius), and stream discharge were recorded.
Lastly, an estimate of adult returns resulting from the use of captive reared adults was conducted as each cohort completed their life cycle, based on the assumption of adults homing to their natal streams (Stabell 1984). For example, eggs deposited by adults outplanted in the fall of 2005 would have emerged in the spring of 2006. These juveniles would typically reside in freshwater for 2 years before out migrating as smolts during the spring of 2008. After 2 years at sea these salmon would return to their natal stream to spawn in the fall of 2010. Factors such as straying adults from other reaches, environmental conditions (Tetzlaff et al. 2005, 2008) or obstructions such as newly constructed beaver dams affect this estimate, but estimating relative return rates still provides an index of success for this management strategy. Thus far, progeny from captive adult releases have only returned as adults in the Chase Mill Stream (2010–2012), Hobart Stream (2011, 2012), and Mopang Stream (2010–2012).
Fry release versus adult release evaluations
One of the main goals of these evaluations was to evaluate whether the release of captive adults was a better strategy than fry releases. In the Northern Stream, electrofishing was conducted during September, 2010–2012 to evaluate growth and abundance of young of the year (YOY) and parr as well as collect biological data such as length, weight, and scales for aging. A combination of multiple-pass depletion estimates (Carle and Strub 1978) and Catch per Unit of Effort (CPUE) sampling (Bateman et al. 2005; Sweka et al. 2006) was used to estimate juvenile abundance within study reaches. Using these data, survival estimates from egg to fry and to age-0 +parr were calculated. Fecundity data collected at CBNFH and published in annual USASAC reports (USASAC, 2005–2012) were used to estimate egg deposition. An egg to fry survival rate of 25 % (Legault 2004) was used to estimate densities for adult origin fry at emergence in an attempt to normalize results for comparison between fry of released adults and stocked fry. The electrofishing results were then used to compare both YOY and parr densities between fry stocked populations (2006–2008) and adult origin populations (2010–2011) resulting from captive adult outplanting activities (2009–2011) using a Kruskal–Wallis rank sum test (Quinn and Keough 2002). These densities were also compared to concurrent fry stocked densities available for other drainages and reaches as well as for previous fry outplanting actions within study reaches using Kruskal – Wallis rank sum tests, with a significance p < 0.05 (Quinn and Keough 2002).
Results
Collections and culture
As mentioned previously, collections for this captive rearing strategy were primarily taken for the broodstock production needs. From 2003 to 2010, a total of 9,060 parr were collected with an average annual collection of 1,133 ±81 parr (Table 4). In most years, drainage targets were met or exceeded. For 2005–2010 collections, survival to maturity in captivity averaged 87 % across all stocks, which provided for 5,813 fish to be spawned or released throughout these years to meet both broodstock needs and release captive adults into the natural environment.
Adult releases
From 2005 through 2011, Maine has released 2,199 adult Atlantic salmon, with actual spawner escapement (ASE) rates ranging from 1.2 to 18.5 times CSE (Table 5) throughout the six major drainages. The numbers of Atlantic salmon adults released was variable depending on availability of adults in excess of broodstock needs, but an effort was made to maintain a 1:1 female to male ratio. The sex ratio varied across years and ranged from 0.6 to 3.0 males to females (Table 5). A minimum of 35 adults were released in the Mopang Stream in 2005 to a maximum of 288 adults in the West Branch Union River in 2011 (Table 5). Over all years, 93.3 % of captive adult releases were age-5 repeat spawners; the remaining adult releases were maiden fish (never previously spawned) and composed of 2 % age-3 and 4.7 % age-4 of all released adults, respectively. The only releases of maiden fish occurred in the Dennys River and West Branch Union River in 2011. Release dates varied from October 3 to November 7, with a mean release date of October 21.
Spawn behavior
Spawning surveys revealed that the distribution of captive adult redds, in relation to the release point, varied from 17.4 km upstream to 10.4 km downstream of the release point with a mean overall dispersal of 3.1 km upstream and downstream of the release point and maximum dispersal of 18 km (Table 6). Median upstream dispersal was 0.6 km and median downstream dispersal was 3.0 km. Release locations did have an effect on movement direction due to obstructions that limited movement. Adults outplanted in the Sheepscot River were outplanted below a dam preventing upstream distribution. Adults outplanted in the Northern Stream were placed in the middle of the reach and distribution extended upstream 6 km and downstream into the East Machias River. In the Dennys River, salmon were released 2.0 km above the head of tide, and fish distributed as far as 17.4 km upstream (Table 6).
Spawning surveys in the Northern Stream indicated that captive adult Atlantic salmon redd construction generally began in the middle of October, with observation of completed redds occurring around the first week of November and all activity ceased by the end of November. This timing was consistent with the spawn timing of wild adults in the Old Stream (10/14–11/17) in 2009 but was later than the spawn timing observed in the Old Stream in 2010 (Fig. 7). Despite these temporal differences, the timing of redd construction was within the range of those reported by Beland et al. (1982). Redd locations across all drainages were also in expected locations such as the tails of riffles or the head of pools in 2–25 cm gravel (Beland et al. 1982; Danie et al. 1984; Fleming 1998).

Timing of spawning in Old Stream, Machias River (Sea-run wild Atlantic salmon) and Northern Stream, East Machias River (captive reared gravid adult Atlantic salmon) for 2009 and 2010 from selected reaches surveyed weekly. Values are proportion of total redds for selected reach by week
Acoustic telemetry data from the Northern Stream in the fall of 2010 found males spent 2–3 weeks on spawning habitat while females spent only 1 week (October 19–30 November 30). Captive adults of both genders moved out of the study reach prior to spawning but there was a high fidelity among females (84 %) and males (86 %) to return to the study reach (Bruchs C, personal communication). Survival through the spawning period was 55 % for females and 50 % for males. After spawning, adults moved into deeper habitat and lakes (Bruchs 2012). Acoustic telemetry data suggests that adults surviving as post-spawners used lakes to overwinter with estimated over-winter survival of 78 % for females and 60 % for males through May of 2011. Overall survival from release (October 2010) through emigration to the estuary (May 2011) was 33 % for females and 30 % for males. One female was even detected by Ocean Tracking Network collaborators in Minas Passage in the inner Bay of Fundy (Broome et al. 2009). These kinds of behaviors are consistent with Atlantic salmon life history given their iteroparity (Danie et al. 1984). Repeat spawners are rare but are occasionally documented in Maine Rivers (USASAC 2013).
Reproductive success
The number of redds per captive female varied from 0 to 3.09 with a mean spawning rate of 0. 94 (±0.34) redds per female. These spawning rates were lower than those found by Fleming (1998) for Atlantic salmon; 3–5 per female in the 1 kg range, to 14 per female in the 6–7 kg range. Overall, the timing of release had an effect on spawning rate (R2 = 0.25, df = 23, p = 0.01) (Fig. 8). Outplanting early fall (October 1st) produced a higher spawning rate, which gradually decreased over time (Fig. 8). This suggests that earlier releases may improve reproductive success; however, it is not known if early releases are possible since fish are not identified for outplanting until late September. The relationship between spawning rate and the proportion of age-5 females in the releases was also examined across years and drainages, and no effect was found (R2 = 0.009, df = 16, p = 0.72); however, this analysis was heavily influenced by a small sample size of age-3 and age-4 captive fish released (7 % of all released adults).

Relationship of redd to female ratio of Atlantic salmon outplanted in Maine between 2005 and 2011 across various streams by outplanting date
Emergence data was collected in two study streams; the Mopang Stream, a tributary of the Machias River in 2006, and the Sheepscot River in 2009. Emergence occurred over 13 and 22 days for the Mopang Stream and Sheepscot River, respectively. Emergence in the Mopang stream began on May 6 and was completed by May 19, with a total of 354 fry trapped. In the Sheepscot emergence started on May 7 and ended by June 13, with 499 fry trapped (Fig. 9). These dates are consistent with emergence timing estimated by Gustafson-Marjanen and Dowse (1983) in the Old Stream, Machias River, Maine. The differences in the onset and termination of emergence were probably due to temporal and geographical differences between the two streams. The emergence numbers are minimum estimates since we did not attempt to prevent intra-gravel alevin movement and largely provide information regarding emergence timing within the study streams.

Cumulative counts of Atlantic salmon fry caught in emergence traps in Mopang Stream, Machias River (2006) and the Sheepscot River (2009)
Adult return data, as measured by the number of redds, from outplanted parents has been limited to Hobart Stream (2011, 2012), Mopang Stream a tributary of the Machias River (2010–2012), and Chase Mill Stream a tributary of the East Machias River (2010–2012) due to short duration of this work relative to the Atlantic salmon lifecycle. Spawning cohorts in the Northern Stream a tributary of the East Machias River will not return until the fall of 2014. The number of redds counted ranged from a high of 10 in Chase Mill Stream in 2011 to a low of 0 in Hobart for all years. In each case, adult to adult return rates varied from 2 to 15 % (mean 5 %).
Fry releases versus adult releases
In the Northern Stream, a tributary to the East Machias River, densities of both YOY and parr were compared between fry stocked origin fish (2006–2008) and captive adult origin fish in 2010–2011 (adults released in 2009–2011). Density of YOY of captive adult origin (12.1/100 m2) was similar to fry stocked origin (15.1/100 m2); (p = 0.32), and survival was similar between captive adult (0.19) and fry origin (0.13) YOY (p > 0.05). Mean captive adult origin parr abundance (6.37 parr/100 m2) was greater than fry stocked origin parr (4.49 parr/m2, p = 0.03), but survival was similar between captive origin (0.53) and fry origin (0.46) parr (p > 0.05); (Table 7).
Juvenile data were also pooled across drainages within the same time frame to compare strategies (Table 8). Densities of fry stocked origin YOY (18.9/100 m2) were greater than captive adult origin YOY (13.1/100 m2; p = 0.01), and densities of fry origin parr (7.9/100 m2) were also greater than captive origin parr (4.8/100 m2, p < 0.05). Survival rates for fry stocked YOY (0.13) and adult origin YOY (0.19) were not different but these values were well below those used by Legault (2004) who presented a mean value of 51.5 % survival from fry to YOY. Likewise, mean fry to parr survival for fry stocked (0.46) and adult origin (0.53) was not different, but these values were greater than Legault (2004) mean survival of 35 %. In each case, comparisons of survival in the Northern Stream or across drainages, it is intriguing that while not significant, mean survival was greater for adult origin YOY and parr. This suggests captive adults may have an advantage over fry stocking, but further investigation is warranted including the influence of estimated fecundities and emergence rates on this comparison.
Discussion
Captive rearing is a relatively uncommon conservation hatchery strategy, this experimental approach has only been used in a few anadromous salmonid populations in North America; coho salmon (Berejikian et al. 1997), Chinook salmon (Berejikian et al. 2001b), and Atlantic salmon (Carr et al. 2004, Dempson et al. 1999, Bruce 1995). Post-release evaluations of Chinook salmon have been limited to captive adults in artificial channels (Berejikian et al. 1997, 1999, 2001b), and only recently, spawn behavior in the wild (Venditti et al. 2013). Several studies have evaluated captive reared Atlantic salmon released into the wild, but findings are mixed. Bruce (1995) found captive adults released into the Big Salmon River, New Brunswick, Canada exhibited a high rate of spawning success, including producing redds. Releases of sea-caged adults into the Bay d’Espoir (estuary) Newfoundland, Canada, found high survival and homing of captive adults to the Conne River (Dempson et al. 1999). Conversely, Carr et al. (2004) found captive reared adults did not contribute to juvenile production; however, they noted their evaluations were likely confounded by release locations and low rate of sexual maturation of released adults. Here, we review the methods and evaluation of this little used tool applied to two programs, Chinook salmon in Idaho and Atlantic salmon in Maine.
Each program approaches the captive rearing concept in slightly different ways, highlighting how captive rearing can be uniquely applied to different species and locations. Some of the differences between these programs may simply be based upon programmatic history and the way in which each program was initially developed. The Chinook salmon captive rearing program in Idaho was designed as an independent experimental strategy with its own monitoring and evaluation. While, the release of captive-reared Atlantic salmon in Maine initially came about opportunistically, when more captive broodstock adults were produced than were needed to spawn. Other differences may be species-specific. For example, a portion of post-spawn adult Atlantic salmon survive to spawn a second time, but all Chinook salmon die post-spawn. The Atlantic salmon program was able to take advantage of iteoparity and release re-conditioned kelts into streams.
The experimental nature of both programs required extensive adaptive management throughout the advancement of each program. For the Chinook salmon program, there was a period of culture development where low survival, primarily due to disease, hampered the success of the program. However, the collection of eyed eggs directly from redds and new bio-security protocols greatly improved survival. The second phase in the Chinook salmon program occurred when greater number of adults were released and the focus of the program shifted to more intensive evaluations of post-release behavior and spawning success. In the Atlantic salmon program, with fewer complications in culture, the early work focused on mechanisms surrounding the use of gravid adults such as redd construction and gamete viability. Atlantic salmon were initially outplanted into less than optimal habitats to avoid possible negative effects on sea-run spawners and their offspring; but once results indicated that these fish could perform similarly to sea-run salmon and juveniles, other study streams were chosen with better habitat access and better rearing conditions.
The culture of captive reared salmon was able to provide a dramatic survival advantage to adults of both programs, with survival from collection to mature adult of 54 and 87 % for Chinook and Atlantic salmon, respectively. The Atlantic salmon program was able to collect and rear parr in captivity, while the Chinook salmon program found higher success with eyed egg collections over parr collections. However, the trade off to increased survival is that captive fish undoubtedly confront a different environment during development and maturation than what wild fish or ‘traditional’ hatchery juveniles released to the wild experience (Hard et al. 2000; Berejikian et al. 2004). As a result of this captive environment there may be some unintended domestication selection and divergent morphological, behavioral, and physiological characteristics compared to their wild counterparts. Proper precautions should be taken to minimize any potential negative impacts during the collection and rearing of captive fish as any alteration may reduce the efficacy of the fish for future supplementation efforts (Araki et al. 2007; 2009). Previous studies suggested differences in behavior and appearance were the result of hatchery domestication in coho (Fleming and Gross 1992, 1993, 1994; Berejikian et al. 1997) and Atlantic salmon (Fleming et al. 1996; Fleming and Einum 1997). In the Chinook salmon programs, we have observed some differences in adult morphology between captive-reared and wild fish, where captive adults were appreciably smaller than wild adults (Venditti et al. 2013). But, as Hard et al. (2000) suggests, these differences may not directly influence spawning behaviors. In fact, captive adult Chinook salmon and Atlantic salmon released to spawn volitionally in their natal streams behaved similarly to their wild counterparts in habitat selection, spawn behavior, and redd construction (Venditti et al. 2013).
Spawning rates for captive adults of each species were below published ranges (Fleming 1998; Gallagher and Gallagher 2005; Murdoch et al. 2009), yet they still contributed production and additional results are encouraging. Reduced spawning rates may be a result of the timing of adult releases for Atlantic salmon or timing of spawning for Chinook salmon (Joyce et al. 1993; Berejikian et al. 2003; Venditti et al. 2013). Further investigations of post spawn success of captive Chinook salmon and Atlantic salmon have also shown positive results. Within-redd survival of progeny from captive Chinook salmon exhibited good survival to eyed-egg (Venditti et al. 2013). Captive origin progeny were also well represented in Maine streams that received adult Atlantic salmon releases, in contrast to findings by Carr et al. (2004). In the Northern Stream, Atlantic salmon parr from captive adult releases were more abundant than parr of fry stocked origin, but both achieved the same survival rate, despite lower emergence densities of captive origin fry relative to fry stocking densities. Life stage evaluations of captive origin Atlantic salmon progeny are ongoing in Maine, but both programs have documented that captive fish successfully reproduce and attain similar survival in early life stages as wild fish.
The final evaluation of captive rearing is not complete; the number of captive offspring that successfully out-migrate and return to natal waters will be the last evaluation of reproductive success. In Maine, redd counts are widely used to estimate adult escapement for Atlantic Salmon streams where there are no weirs or traps in place to enumerate adult returns. To date, adult return data from captive adult release has been limited to the Hobart Stream, Mopang Stream and Chase Mill Stream due to short duration of this work relative to the Atlantic salmon lifecycle (Danie et al. 1984, Baum 1997), and spawning cohorts in the Northern Stream will not return until the fall of 2014. However, initial parentage analyses of Atlantic salmon parr and smolts collected in the Hobart Stream found 38.2 % of females and 38.9 % of males were assigned to parents of captive adults (Bartron unpublished data).
In Idaho, the adult to adult parentage analyses will be used to assess reproductive success of captive adults released into the EFSR. Tissue samples are collected from all adult returns (potential parents or offspring) captured at the EFSR adult trap, and from all captive adults released to spawn naturally (parents). Preliminary parentage analyses of EFSR returns indicate that 21 adults returned from the spawning events in 2006 and 30 fish have returned so far from the spawning events in 2007 (E. Stark unpublished data). Furthermore, initial findings of phenotypic length and age of progeny of captive Chinook salmon initially supports the intimation by Kitada et al. (2011), that some consequences of captive rearing (smaller body size at release) could be eliminated in the first generation after reintroducing hatchery-reared fish. The question of lifetime reproductive success still needs to be examined for both the Chinook salmon and Atlantic salmon programs, but these case studies both show that captive-reared fish clearly contributed to production.
Initial results from these programs suggest that captive rearing may be a viable short term measure to address immediate demographic concerns and to ultimately aid in the recovery of depressed populations. This approach may impart a smaller footprint on the wild population than ‘traditional’ supplementation methods and may be an alternative hatchery rearing strategy in streams where spawning habitat is intact but there are other factors impeding survival of juveniles or adults. Any benefits to natural production may be greatly reduced if spawning habitat is degraded and the released fish are unable to spawn, as we found in the WFYF in Idaho after a forest fire and in some of the initial Maine study streams. Furthermore, while this strategy was performed on very small scales for both of these programs, this strategy could conceivably be scaled-up to larger conservation programs, where more eggs or juveniles could be collected from the study streams. However, existing wild production and goals for the program would determine the scale of collection and a considerable increase in effort and cost may be incurred. Given species-specific generation lengths and programmatic changes over time, it is also imperative that evaluations be performed over long enough periods to determine the efficacy of captive rearing. These two programs, although different in some aspects, share similar methods, tools, and results to date. Final results from both programs will increase our collective understanding of the potential benefits the captive rearing approach offers for addressing demographic and genetics risks, preventing year-class failures, and contributing to the recovery of at-risk populations of salmon in North America.
Abbreviations
- IDFG:
-
Idaho Department of Fish and Game
- LEM:
-
Lemhi River
- WFYF:
-
West Fork Yankee Fork Salmon River
- EFSR:
-
East Fork Salmon River
- MDMR:
-
Maine Department of Marine Resources
References
Araki H, Cooper B, Blouin MS (2007) Genetic effects of captive breeding cause a rapid, cumulative fitness decline in the wild. Science 318:100–103. http://www.sciencemag.org/content/318/5847/100.full
Araki H, Cooper B, Blouin MS (2009) Carry-over effect of captive breeding reduces reproductive fitness of wild-born descendants in the wild. Biol Lett 5:621–624. http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2781957/
Baker DB et al (2006) Captive rearing program for Salmon River Chinook salmon. 2005 Project Progress Report to the Bonneville Power Administration, Contract Number 00004002, Portland, Oregon. https://collaboration.idfg.idaho.gov/FisheriesTechnicalReports/Res06-27Baker2005%20Captive%20Rearing%20Initiative%20for%20Salmon%20River%20Chinook%20Salmon.pdf
Bartron M, Buckley D, King T, King TL, Kinnison MG, Sheehan T, Beland K, Marancik J (2006) Draft Captive Broodstock Management Plan for Atlantic salmon. Craig Brook National Fish Hatchery, East Orland, Maine
Bateman DS, Gresswell RE, Torgersen CE (2005) Evaluating single-pass catch as a tool for identifying spatial pattern in fish distribution. J Freshw Ecol 20(2):335–345. http://connection.ebscohost.com/c/articles/17154376/evaluating-single-pass-catch-as-tool-identifying-spatial-pattern-fish-distribution
Baum E (1997) Maine Atlantic salmon, a national treasure. Atlantic Salmon Unlimited, Hermon, Maine
Baum ET, Meister AL (1971). Fecundity of Atlantic salmon (Salmo salar) from two Maine rivers. Fish Res Board Can 28:764–767. http://www.nrcresearchpress.com/doi/abs/10.1139/f71-106#.Um7bpRB9uTQ
Beland KF (1996) The relationship between redd counts and Atlantic salmon (Salmo salar) parr populations in the Dennys River, Maine. Can J Fish Aquat Sci 53:513–519. http://www.nrcresearchpress.com/doi/pdf/10.1139/f95-216
Beland KF, Jordan RM, Meister AL (1982) Water depth and velocity preferences of spawning Atlantic salmon in Maine rivers. N Am J of Fish Manag 2:11–13. http://www.tandfonline.com/doi/abs/10.1577/1548-8659%281982%292%3C11%3AWDAVPO%3E2.0.CO%3B2#preview
Berejikian BA, Tezak EP, Schroder SL, Knudsen CM, Hard JJ (1997) Reproductive behavioral interactions between wild and captively reared coho salmon (Oncorhynchus kisutch). ICES J Mar Sci 54:1040–1050. http://icesjms.oxfordjournals.org/content/54/6/1040.full.pdf
Berejikian BA, Tezak EP, Schroder SL, Flagg TA, Knudsen CM (1999) Competitive differences between newly emerged offspring of captive-reared and wild coho salmon. Trans Am Fish Soc 128:832–839. http://www.tandfonline.com/doi/abs/10.1577/1548-8659%281999%29128%3C0832%3ACDBNEO%3E2.0.CO%3B2#preview
Berejikian BA, Tezak EP, Park L, LaHood E, Schroder SL, Beall E (2001a) Male competition and breeding success in captively reared and wild coho salmon (Oncorhynchus kisutch). Can J Fish Aquat Sci 58:804–810. http://www.nrcresearchpress.com/doi/abs/10.1139/f01-039#.Um7dNRB9uTQ
Berejikian BA, Tezak EP, Schroder SL (2001b) Reproductive behavior and breeding success of captively reared Chinook salmon. N Am J Fish Manag 21:255–260. http://www.researchgate.net/publication/254310988_Reproductive_Behavior_and_Breeding_Success_of_Captively_Reared_Chinook_Salmon
Berejikian BA, Fairgrieve WT, Swanson P, Tezak EP (2003) Current velocity and injection of GnRHA affect reproductive behavior and body composition of captively reared Chinook salmon (Oncorhynchus tshawytscha). Can J Fish Aquat Sci 60:690–699. http://nativefishsociety.org/conservation/wild_vs_hatchery/annotated_bibliography_on_salmon_hatcheries/fitness/CurrentVelocity.htm
Berejikian BA, Flagg T, Kline PA (2004) Release of captively reared adult anadromous salmonids for population maintenance and recovery: biological trade-offs and management considerations. In: Nickum MJ, Mazik PM, Nickum JG, MacKinlay DD (eds). Propagated fish in resource management. American Fisheries Society, Symposium 44, Bethesda, pp 233–245. http://www.nwfsc.noaa.gov/publications/scipubs/display_doctrack_allinfo.cfm?doctrackmetadataid=1150
Berejikian BA, Gable JT, Vidergar DT (2011) Effectiveness and trade-offs associated with hydraulic egg collections from natural salmon and steelhead redds for conservation hatchery programs. Trans Am Fish Soc 140:549–556. http://www.tandfonline.com/doi/abs/10.1080/00028487.2011.583540#preview
Broome J, Stokesbury M, Redden A, Bradford R (2009) 3-D Acoustic Tracking of Fish, Sediment-Laden Ice and Large Wood Debris in the Minas Passage of the Bay of Fundy. http://members.oceantrack.org/data/discovery/NSP.htm. Accessed 21 February 2013
Bruce H (1995) Big doings on the Big Salmon: volunteers spark special stocking. Atl Salmon Fed J 14:1–2
Bruchs C (2012) CBNFH Pre-spawn Outplant Assessment. Assessments of the demographic benchmarks of the Gulf of Maine Distinct Population Segment of Atlantic Salmon of Mutual Interest to Maine DMR and NOAA. Semi-Annual Project Report. NOAA Grant NA11NMF4720235. Covering the period January 1, 2012 to June 30, 2012. Maine Department of Marine Resources, Sea Run Fisheries and Habitat, Bangor, Maine. http://www.maine.gov/dmr/searunfish/salmonprojects/salmonnoaareports.shtml
Bruchs C, Atkinson E (2013) CBNFH Pre-spawn Outplant Assessment. Assessments of the demographic benchmarks of the Gulf of Maine Distinct Population Segment of Atlantic Salmon of Mutual Interest to Maine DMR and NOAA. Semi-Annual Project Report. NOAA Grant NA11NMF4720235. Covering the period July 1, 2012 to December 31, 2012. Maine Department of Marine Resources, Sea Run Fisheries and Habitat, Bangor, Maine. http://www.maine.gov/dmr/searunfish/salmonprojects/salmonnoaareports.shtml
Burner CJ (1951) Characteristics of spawning nests of Columbia River salmon. U.S. Fish and Wildlife Serv Fish Bull 52:97–110. http://www.nativefishlab.net/library/textpdf/14037.pdf
Carle FL, Strub MR (1978) A new method for estimating population size from removal data. Biometrics 34:621–630
Carr JW, Whoriskey F, O’Reilly PO (2004) Efficacy of releasing captive reared broodstock into an imperiled wild Atlantic salmon population as a recovery strategy. J Fish Biol 65 (Supplement A):38–54. http://onlinelibrary.wiley.com/doi/10.1111/j.0022-1112.2004.00546.x/abstract
Chebanov NA, Riddell BE (1998) The spawning behavior, selection of mates, and reproductive success of Chinook Salmon (Oncorhynchus tshawytscha) spawners of natural and hatchery origins under conditions of joint spawning. J Ichthyol 38:517–526
Connor WP, Marshall AR, Bjornn TC, Burge HL (2001) Growth and long-range dispersal by wild sub yearling spring and summer Chinook salmon in the Snake River basin. Trans Am Fish Soc 130(6):1070–1076. doi:10.1577/1548-8659(2001)130<1070:GALRDB>2.0.CO;2
Copeland T, Venditti DA (2009) Contribution of three life history types to smolt production in a Chinook salmon (Oncorhynchus tshawytscha) population. Can J Fish Aquat Sci 66:1658–1665. http://www.nrcresearchpress.com/doi/abs/10.1139/F09-110#.Uo5t5V_n8iQ
Cuenco ML et al (1993) The use of supplementation to aid in natural stock restoration. In: Cloud JG, Thorgaard GH (eds) Genetic conservation of Salmonid Fisheries, Plenum Press. New York,pp 269–293. http://link.springer.com/chapter/10.1007/978-1-4615-2866-1_24#page-1
Danie DS, Trial JG, Stanley JG (1984) Species profiles: life histories and environmental requirements of coastal fish and invertebrates (North Atlantic)—Atlantic salmon. U.S. Fish and Wildlife Service. FWS/OBS-82/11.22. U.S. Army Corps of Engineers, TR EL-82-4. 19 pp http://www.nwrc.usgs.gov/wdb/pub/species_profiles/82_11-022.pdf
Dempson JB, Pepper VA, Furey G, Bloom M, Nicholls T, Hoskins G (1999) Evaluation of an alternative strategy to enhance salmon populations: cage rearing wild smolts from Conne River, Newfoundland. ICES J Mar Sci 56:422–432. http://icesjms.oxfordjournals.org/content/56/4/422.short-term
Fay C, Bartron M, Craig S, Hecht A, Pruden J, Saunders R, Sheehan T, and Trial J (2006) Status Review for Anadromous Atlantic Salmon (Salmo salar) in the United States. Report to the National Marine Fisheries Service and U.S. Fish and Wildlife Service. 294 pp. http://www.nmfs.noaa.gov/pr/species/statusreviews.htm
Flagg TA, Mahnken CVW (1995) An assessment of the status of captive broodstock technology for Pacific Salmon. Final report to the Bonneville Power Administration (Project No. 93-56), P.O. Box 3621, Portland, 225 pp. http://www.researchgate.net/publication/241975189_An_Assessment_of_the_Status_of_Captive_Broodstock_Technology_of_Pacific_Salmon_1995_Final_Report)
Flagg TA, McAuley WC, Kline PA, Powell MS, Taki D, Gislason JC (2004) Application of captive broodstocks to preservation of ESA-listed stocks of Pacific salmon: Redfish Lake Sockeye Salmon case example. In: Nickum MJ, Mazik PM, Nickum JG, MacKinlay DD (eds) Propagated fish in resource management. American Fisheries Society, Symposium 44, Bethesda, pp 387–400
Fleming IA (1998) Pattern and variability in the breeding system of Atlantic salmon (Salmo salar), with comparisons to other salmonids. Can J Fish Aquat Sci 55 (Suppl. 1):59–76. http://www.nrcresearchpress.com/doi/pdf/10.1139/d98-009
Fleming IA, Einum S (1997) Experimental tests of genetic divergence of farmed from Atlantic salmon due to domestication. ICES J Mar Sci 54:1051–1063. http://icesjms.oxfordjournals.org/content/54/6/1051.abstract
Fleming IA, Gross MR (1992) Reproductive behavior of hatchery and wild coho salmon (Oncorhynchus kisutch): does it differ? Aquac 103:101–121. http://www.sciencedirect.com/science/article/pii/004484869290405A
Fleming IA, Gross M (1993) Breeding success of hatchery and wild coho salmon (Oncorhynchus kisutch) in competition. Ecol Appl 3(2):230–245. http://www.jstor.org/discover/10.2307/1941826?uid=3739648&uid=2129&uid=2&uid=70&uid=4&uid=3739256&sid=21102848067633
Fleming IA, Gross MR (1994) Breeding competition in a Pacific salmon (coho: Oncorhynchus kisutch): measures of natural and sexual selection. Evolution, 48:637–657. http://www.jstor.org/discover/10.2307/2410475?uid=3739648&uid=2129&uid=2&uid=70&uid=4&uid=3739256&sid=21102848067633
Fleming IA, Jonsson B, Gross MR, Lamberg A (1996) An experimental study of the reproductive behaviour and success of farmed and wild Atlantic salmon (Salmo salar) J App Ecol 33:893–905. http://www.jstor.org/discover/10.2307/2404960?uid=3739648&uid=2129&uid=2&uid=70&uid=4&uid=3739256&sid=21102848067633
Fraser DJ (2008) How well can captive breeding programs conserve biodiversity? A review of salmonids. Evol Appl 1(4):535–596. http://onlinelibrary.wiley.com/doi/10.1111/j.1752-4571.2008.00036.x/full
Gallagher SP, Gallagher CM (2005) Discrimination of Chinook salmon, Coho salmon, and steelhead redds and evaluation of the use of redd data for estimating escapement in several unregulated stream in Northern California. N Am J Fish Manag 25:284–300. http://www.fs.fed.us/psw/publications/4351/Gallagher.pdf
Gilbert CH (1913) Age at maturity of the Pacific coast salmon of the genus Oncorhynchus. Bull Bur Fish 32:1–22. http://aquaticcommons.org/3016/1/age_maturity_pacific_coast_salmon.pdf
Gustafson-Marjanen KI, Dowse HB (1984) Seasonal and diel patterns of emergence from the redd of Atlantic salmon (Salmo salar) fry. Can. J. Fish. Aquat. Sci. 40:813-817. http://www.nrcresearchpress.com/doi/abs/10.1139/f83-106?journalCode=cjfas)
Hard JJ, Berejikian BB, Tezak EP, Schroeder SL, Knudsen CM (2000) Evidence for morphometric differentiation of wild and captively reared adult coho salmon: a geometric analysis. Environ Biol Fish 58:61–73. http://link.springer.com/article/10.1023%2FA%3A1007646332666#page-1
Hassemer PF et al (1999) Captive rearing initiative for Salmon River Chinook salmon. 1998 Project Progress Report to the Bonneville Power Administration, Contract Numbers 97-BI-97538 and 98-BI-63416. Portland, Oregon. https://collaboration.idfg.idaho.gov/FisheriesTechnicalReports/Res99-03Hassemer1998%20Captive%20Rearing%20Initiative%20for%20Salmon%20River%20Chinook%20Salmon.pdf
Healey MC (1991) Life history of Chinook salmon (Oncorhynchus tshawytscha) In: Groot C, Margolis L (eds) Pacific salmon life histories. UBC Press, Vancouver, pp 311–393. http://books.google.com/books?hl=en&lr=&id=I_S0xCME0CYC&oi=fnd&pg=PA313&dq=Life+history+of+chinook+salmon+healey&ots=_vCDrM5ic-&sig=_B_Ipod5tYrfb50trweO0V5Kijk#v=onepage&q&f=false
Hebdon JL, Kline P, Taki D, Flagg, TA (2004) Evaluating reintroduction strategies for Redfish Lake Sockeye Salmon captive broodstock progeny. In: Nickum MJ, Mazik PM, Nickum JG, MacKinlay DD (eds) Propagated fish in resource management. American Fisheries Society, Symposium 44, Bethesda, pp 401–413 (https://collaboration.idfg.idaho.gov/FisheriesTechnicalReports/Evaluating%20Reintroduction%20Strategies%20for%20Redfish%20Lake%20Sockeye%20Salmon.pdf
Heindel JA, Baker DJ, Johnson KA, Kline PA, Redding JJ (2005) A simple isolation incubator for specialized rearing of salmonid eggs and first feeding fry. N Am J Aquac 67:13–17. http://www.tandfonline.com/doi/abs/10.1577/FA04-017.1?journalCode=unaj20#preview
Hess MA, Rabe CD, Vogel JL, Stephenson JJ, Nelson DD, Narum SR (2012) Supportive breeding boosts natural population abundance with minimal negative impacts on fitness of a wild population of Chinook salmon. Mol Ecol 21(21):5236–5250. http://onlinelibrary.wiley.com/doi/10.1111/mec.12046/full
Johnson K, Heindel J (2001) Efficacy of manual removal and ivermectin gavage for control of Salmincola californiensis (Wilson) infestation of Chinook salmon, Oncorhynchus tshawytscha (Walbaum), captive broodstocks. J Fish Dis 24(4):197–302. http://onlinelibrary.wiley.com/doi/10.1046/j.1365-2761.2001.00279.x/abstract
Joyce JE, Martin RM, Thrower FP (1993) Successful maturation of captive chinook salmon broodstock. Prog Fish Cult 55:191–194. http://www.tandfonline.com/doi/abs/10.1577/1548-8640%281993%29055%3C0191%3ASMASOC%3E2.3.CO%3B2#preview
Kennedy P, Copeland T, Johnson J, Apperson KA, Flinders J, Hand R (2011) Idaho natural production monitoring and evaluation. 2009-2010 Project Progress Report to the Bonneville Power Administration, Contract Numbers 40873 and 45995. Portland, Oregon. https://collaboration.idfg.idaho.gov/FisheriesTechnicalReports/Res11-23Kennedy2009-2010 INPMEP Report.pdf
Kitada SH, Kishino H, Hamasaki K (2011) Bias and significance of relative reproductive success estimates based on Steelhead Trout (Oncorhynchus mykiss) data: a Bayesian meta-analysis. Can J Fish Aquat Sci 68:1827–1835. http://www.nrcresearchpress.com/doi/abs/10.1139/f2011-087#.Um_aYBB9uTQ
Kuligowski DR, Ford MJ, Berejikian BA (2005) Breeding structure of Steelhead inferred from patterns of genetic relatedness among nests. Trans Am Fish Soc 134:1202–1212. http://www.tandfonline.com/doi/abs/10.1577/T04-187.1#preview
Legault CM (2004) Salmon PVA: a population viability analysis model for Atlantic salmon in the Maine Distinct Population Segment. U.S. Department of Commerce Northeast Fisheries Science Center Reference Document 04-02; 88 p. Available from: National Marine Fisheries Service, 166 Water St., Woods Hole, Massachusetts 02543-1026. http://nefsc.noaa.gov/publications/crd/crd0402/crd0402.pdf
Lichatowich JA (1999) Salmon without rivers. Island Press, Washington DC http://islandpress.org/ip/books/book/islandpress/S/bo3560726.html
MacKenzie C, Moring JR (1988) Estimating survival of Atlantic salmon during the intragravel period. N Am J Fish Manag 8:45–48. http://www.tandfonline.com/doi/abs/10.1577/1548-8675%281988%29008%3C0045%3AESOASD%3E2.3.CO%3B2#preview
Mackey G, Atkinson E (2003) Summary of emergent fry trapping on the Dennys River in 2001 and 2002: evaluation of reproductive success by pen-reared adult Atlantic salmon. Atlantic salmon freshwater assessments and research. Semi-Annual Project Report. NOAA Grant NA17FL1157. Covering the period May1, 2002 to October 31, 2002. Maine Atlantic Salmon Commission Bangor, Maine http://www.maine.gov/dmr/searunfish/salmonprojects/salmonnoaareports.shtml
Mackey G, Brown N (2003) Appendix 3. Estimation of gamete viability and fecundity of river-specific marine net pen reared Atalntic salmon in Maine. Semi-Annual Project Report. NOAA Grant NA17FL1157. Covering the period May1, 2002 to October 31, 2002. Maine Atlantic Salmon Commission Bangor, Maine http://www.maine.gov/dmr/searunfish/salmonprojects/salmonnoaareports.shtml
Mahnken CV, Ruggerone WG, Waknitz FW, Flagg TA (1998) A historical perspective on salmonid production from Pacific rim hatcheries. North Pacific Anadromous Fish Commission Bulletin, 1:38–53. http://www.npafc.org/new/publications/Bulletin/Bulletin%20No.%201/page%2038-53%28Mahnken%29.PDF
Maynard DJ, McAuley WC, Flagg TA (2007) Snake River spring/summer chinook captive broodstock rearing and research, 2006. Annual report to Bonneville Power Administration, Contract 96-AI-96441, 56 p
Maynard DJ, Flagg TA, McAuley WC, Frost DA, Kluver B, Wastel MR, Colt JE, Dickhoff WW (2012) Fish culture technology and practices for captive broodstock rearing of ESA-listed salmon stocks. U.S. Department of Commerce. NOAA Technical Memorandum. NMFS-NWFSC-117, 65 p. http://www.worldcat.org/title/fish-culture-technology-and-practices-for-captive-broodstock-rearing-of-esa-listed-salmon-stocks/oclc/795202047
Murdoch AR, Pearsons TN, Maitland TW (2009) The number of redds constructed per female spring Chinook Salmon in the Wenatchee River Basin. N Am J Fish Manag 29(2):441–446. http://www.tandfonline.com/doi/abs/10.1577/M08-063.1#preview
Naish KA, Taylor JE, Levin PS et al (2007) An evaluation of the effects of conservation and fishery enhancement hatcheries on wild populations of salmon. Advances Mar Biol 53:61–194. http://www.ncbi.nlm.nih.gov/pubmed/17936136
National Research Council (NRC) (1996) Upstream: Salmon and society in the Pacific Northwest. National Academy Press, Washington DC http://www.nap.edu/openbook.php?record_id=4976
Neilson JD, Bradford CE (1983) Chinook salmon (Oncorhynchus tshawytscha) spawner characteristics in relation to redd physical features. Can J Zool 61(7):1524–1531. http://www.nrcresearchpress.com/doi/abs/10.1139/z83-205
NMFS (National Marine Fisheries Service) (1992) Threatened status for Snake River spring/summer Chinook salmon, threatened status for Snake River fall Chinook salmon. Final rule. Federal Register 57:78 (April 22, 1992):14563–14663. http://swr.nmfs.noaa.gov/attach1.htm
Petrosky CE, Schaller HA (2010) Influence of river conditions during seaward migration and ocean conditions on survival rates of Snake River Chinook salmon and steelhead. Ecology of Freshw Fish 19:520–536. http://onlinelibrary.wiley.com/doi/10.1111/j.1600-0633.2010.00425.x/abstract
Phillips RW, Koski KV (1969) A fry trap method for estimating salmonid survival from egg deposition to fry emergence. Fish Res Board Can 26:133–141. http://www.nrcresearchpress.com/doi/abs/10.1139/f69-012#.Um_ixBB9uTQ
Quinn GP, Keough MJ (2002) Experimental design and data analysis for biologists. Cambridge University Press. http://catdir.loc.gov/catdir/samples/cam033/2001037845.pdf
Reisenbichler RR, Rubin SP (1999) Genetic changes from artificial propagation of Pacific salmon affect the productivity and viability of supplemented populations. ICES J Mar Sci 56:459–466. http://icesjms.oxfordjournals.org/content/56/4/459.abstract
Saunders R, Hachey MA, Fay CW (2006) Maine’s diadromous fish community: past, present, and implications for Atlantic salmon recovery. Fisheries 31:537–547. http://ijc.org/files/publications/Endangered%20Species.pdf
Stabell OB (1984) Homing and olfaction in salmonids: a critical review with special reference to the Atlantic salmon. Biol Rev 59:333–388. http://onlinelibrary.wiley.com/doi/10.1111/j.1469-185X.1984.tb00709.x/abstract
Stark EJ et al (2008) Captive rearing program for Salmon River Chinook salmon 2007. Project Progress Report to the Bonneville Power Administration, Contract Number 00029463. Portland, Oregon (https://collaboration.idfg.idaho.gov/FisheriesTechnicalReports/Res08-18Stark2007%20Captive%20Rearing%20Initiative%20for%20Salmon%20River%20Chinook%20Salmon.pdf
Stark EJ et al (2009) Captive rearing program for Salmon River Chinook salmon. 2008 Project Progress Report to the Bonneville Power Administration, Contract Number 00035399. Portland, Oregon (https://collaboration.idfg.idaho.gov/FisheriesTechnicalReports/Res09-04Stark2008%20Captive%20Rearing.pdf
Stark EJ, Gable J (2010) Captive rearing program for Salmon River Chinook salmon. 2009 Project Progress Report to the Bonneville Power Administration, Contract Number 00039364. Portland, Oregon. https://collaboration.idfg.idaho.gov/FisheriesTechnicalReports/Res10-01Stark2009%20Captive%20Rearing.pdf
Stark EJ, Richardson D (2011) Captive rearing program for Salmon River Chinook salmon. 2010 Project Progress Report to the Bonneville Power Administration, Contract Number 00044419. Portland, Oregon. https://collaboration.idfg.idaho.gov/FisheriesTechnicalReports/Res11-03Stark2010%20Chinook%20Captive%20Rearing.pdf
Sweka JA, Legault CM, Beland KF, Trial JT, Millard MJ (2006) Evaluation of removal sampling for basinwide assessment of Atlantic salmon. N Am J Fish Manag 26:995–1002. http://www.tandfonline.com/doi/abs/10.1577/M05-079.1#preview
Tetzlaff D, Soulsby C, Youngson AF, Gibbins C, Bacon PJ, Malcolm IA, Langan S (2005) Variability in stream discharge and temperature: a preliminary assessment of the implications for juvenile and spawning Atlantic salmon. Hydrol Earth Syst Sci 9: 193–208. http://www.hydrol-earth-syst-sci.net/9/193/2005/hess-9-193-2005.html
Tetzlaff D, Gibbins C, Bacon PJ, Youngson AF, Soulsby C (2008) Influence of hydrological regimes on the pre-spawning entry of Atlantic salmon (Salmo salar L.) into an upland river. River Res Appl 24:528–542. http://onlinelibrary.wiley.com/doi/10.1002/rra.1144/abstract
Trial J, Fay C, Kocik J, Marancik J, Beland K, Anthony J, Mackey G (2010) Strategies for using hatchery production in excess of river specific management needs in Maine Rivers. A report to the Maine Atlantic Salmon Technical Advisory Committee
USASAC (United States Atlantic Salmon Assessment Committee) (2009) Annual report of the U.S. Atlantic Salmon Assessment Committee. Report No. 22-2009 Activities. http://www.nefsc.noaa.gov/USASAC/Reports/USASAC2010-Report%2322-2009-Activities.pdf
USASAC (United States Atlantic Salmon Assessment Committee) (2012) Annual report of the U.S. Atlantic Salmon Assessment Committee. Report No. 24-2011 Activities. http://www.nefsc.noaa.gov/USASAC/Reports/USASAC2012-Report%2324-2011-Activities.pdf
USASAC (United States Atlantic Salmon Assessment Committee) (2013) Annual report of the U.S. Atlantic Salmon Assessment Committee. Report No. 25-2012 Activities. http://www.nefsc.noaa.gov/USASAC/Reports/USASAC2013-Report%2325-2012-Activities.pdf
USFWS (U.S. Fish and Wildlife Service), NOAA (National Oceanic and Atmospheric Administration) (2000) Endangered and threatened species; final endangered status for a distinct population segment of anadromous Atlantic salmon (Salmo salar) in the Gulf of Maine; Final Rule. Federal Register 65:223 (November 17, 2000):69459–69483. http://ecos.fws.gov/docs/federal_register/fr3678.pdf
Venditti DA et al (2003) Captive rearing program for Salmon River Chinook salmon. 2001 Project Progress Report to the Bonneville Power Administration, Contract Number 00004002. Portland, Oregon (https://collaboration.idfg.idaho.gov/FisheriesTechnicalReports/Res03-33Venditti2001%20Captive%20Rearing%20Initiative%20for%20Salmon%20River%20Chinook%20Salmon.pdf
Venditti DA et al (2005) Captive rearing program for Salmon River Chinook salmon. 2003 Project Progress Report to the Bonneville Power Administration, Contract Number 00004002. Portland, Oregon. https://collaboration.idfg.idaho.gov/FisheriesTechnicalReports/Res05-06Venditti2003%20Captive%20Rearing%20Initiative%20for%20Salmon%20River%20Chinook%20Salmon.pdf
Venditti DA, James C, Kline P (2013) Reproductive behavior and success of captive-reared Chinook salmon spawning under natural conditions. N Am J Fish Manag 33:97–107. http://www.tandfonline.com/doi/abs/10.1080/02755947.2012.746244#preview
Wang J, Ryman N (2001) Genetic effects of multiple generations of supportive breeding. Conserv Biol 15:1619–1631. http://onlinelibrary.wiley.com/doi/10.1046/j.1523-1739.2001.00173.x/abstract
Acknowledgments
We thank C. Bruchs, D. Baker, P. Kline, D. Schill, and J. Trial for draft manuscript review; M. Peterson, C. Steele, W. Ardren, and D. Venditti for valuable discussion; B. Ayers for data compilation, and K. Plaster for producing the Idaho study map. Funding for the IDFG Chinook salmon program is provided by the Bonneville Power Administration (Project Number 2007-403-00) through the Northwest Power and Conservation Council’s Fish and Wildlife Program. Funding for the MDMR Atlantic salmon program is provided by The National Oceanic Atmospheric Administration (NOAA Grant NA11NMF4720235). Use of trade names does not imply endorsement by the Idaho Department of Fish and Game or The Maine Department of Marine Resources.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Stark, E.J., Atkinson, E.J. & Kozfkay, C.C. Captive rearing for Chinook salmon (Oncorhynchus tshawytscha) and Atlantic salmon (Salmo salar): the Idaho and Maine experiences. Rev Fish Biol Fisheries 24, 849–880 (2014). https://doi.org/10.1007/s11160-014-9346-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11160-014-9346-x