
Abstract
The study compares lyophilized broccoli sprouts and florets in terms of their chemical composition, cytotoxic and proapoptotic potential against hepatocellular carcinoma HepG2, colorectal cancer SW480, and skin fibroblast BJ cells. Sinapic and isochlorogenic acids were predominant phenolics in the sprouts and florets, respectively. The amount of sulforaphane in the sprouts was significantly higher vs. florets. Oleic and linoleic acids dominated in the sprouts, while caproic, stearic and oleic acids in the florets. Broccoli sprouts were selectively cytotoxic on HepG2 and SW480 cells, with proapoptotic effect for the latter, while the florets were less selective, but more active, with profound proapoptotic effect for HepG2 cells (77.4%). Thus, lyophilized broccoli sprouts may be effectively used in dietary chemoprevention.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Broccoli (Brassica oleracea L. var. italica, Brassicacae) are a good source of isothiocyanates, with most important sulforaphane (SF), known for its chemopreventive properties [1], but are also rich in vitamins (C, K, β-carotene), dietary fibre, polyphenols and fatty acids [2], with considerable health beneficial effects [3]. Broccoli marketable sprouts, florets, flour, fiber, flakes, powder, crisps, etc. are under vast interest nowadays, due to their preventive role in non-communicable diseases, like hypertension, atherosclerosis [2, 4] or cancer (lung, breast, prostate and pancreatic) [1]. Consumption of Brassica vegetables may decrease the risk of gastric cardia and esophageal adenocarcinomas [5], but also colon and colorectal cancers [6] in humans. The so far described phytochemical composition of broccoli concerns mostly fresh material, while many studies discuss the influence of processing methods (cutting, boiling, steaming, etc.) on the loss of active substances, especially SF [7, 8]. Moreover, fresh broccoli are difficult to store for more than a few weeks and, except from the storage in argon or helium atmospheres [9], the data on the alternative methods of preservation is scarce. The studies on the impact of storage conditions on phytochemical content of fresh and lyophilized broccoli products are still limited. Therefore, the aim of our study was qualitative and quantitative analysis of lyophilized broccoli sprouts and florets, focusing on polyphenols (flavonoids, phenolic acids), SF, and fatty acids. Moreover, we also studied broccoli products’ effect on the viability and apoptosis induction in human cancer and normal cells, to determine not only the cytotoxic potential, but also selectivity of the tested samples and the mechanism of cell death.
Material and Method
Plant material and Freeze - Drying Method
Broccoli seeds (Brassica oleracea L. var. italica) were obtained commercially [10], detailed information about the material and freeze - drying method is described in supplementary material.
Extracts Preparation
Extraction process was described in details in supplementary material. SF and lipids were extracted according to [11, 12].
HPLC Determination of Flavonoids and Phenolic Acids
The analysis was performed according to [13] and is described in supplementary material.
UPLC/MS Determination of Sulforaphane Content
The analysis was performed according to [14] and is described in supplementary material.
GC Determination of Fatty Acids Profile
The analysis was performed according to [12] and is described in supplementary material.
Cytotoxicity Assay, Apoptosis and Necrosis Analysis
The analysis was performed according to [15,16,17] and is described in supplementary material.
Statistical Analysis
Statistical differences were determined using Student’s t-test. P < 0.05 was considered to be statistically significant. The hierarchical principal component analysis was used to reveal the correlation structure between the investigated parameters [14, 18] and is described in supplementary material.
Results and Discussion
Phenolic Acids and Flavonoids
Flavonoids and phenolic acids are phytochemicals widespread in brassica vegetables [19]. Qualitative HPLC analysis of broccoli sprouts revealed chlorogenic, p-coumaric, ferulic, gentisic and sinapic acids, and also robinin, with traces of myricetin, luteolin, quercetin and apigenin. In broccoli florets only caffeic, isochlorogenic and sinapic acids and also traces of myricetin were detected. Although some peaks on HPLC chromatograms remained unidentified, we decided to determine and compare the content of the recognized polyphenols in both extracts by means of quantitative HPLC analysis. The results (Table 1) revealed vast differences in the amounts of the phenolic acids of the tested broccoli sprouts and florets extracts. Sinapic, gentisic, and ferulic acids predominated in broccoli sprouts, with chlorogenic and p-coumaric acids in smaller amounts. In contrast, isochlorogenic acid, not detected in sprouts, was present in the highest amount in broccoli florets, accompanied with sinapic and caffeic acids. A range of phenolic acids have been identified so far in broccoli sprouts including chlorogenic, gallic, sinapic, ferulic, benzoic, vanillic, protocatechuic or p-coumaric acid [20,21,22,23], while neochlorogenic and chlorogenic acids, together with sinapic and ferulic acids derivatives, were detected in the florets. The results of Pająk et al. [20] indicated similar qualitative phenolic acids profile, but with quantitative differences: ferulic (7.66 mg/100 g dw), p-coumaric (2.04 mg/100 g dw) and chlorogenic (11.49 mg/100 g dw) acids were in lower, while sinapic acid (548 mg/100 g dw) was in higher amounts, in comparison to our results. For broccoli florets, our results are similar to those by Fernández-León et al. [21] for sinapic acid in some Monaco cultivars. Our results indicated robinin, a kaempferol glucoside, to be the only flavonoid identified in broccoli sprouts. Similar results were reported by Pająk et al. [20] and Gawlik-Dziki et al. [22], indicating kaempferol as the major compound in broccoli sprouts after hydrolysis. Other authors identified also luteolin, apigenin, quercetin, and kaempferol, contributing altogether to 2.77–14.66 mg/100 g dw [20, 21].
Sulforaphane Content
The content of SF is given in Table 1. Broccoli sprouts were significantly richer in SF than the florets, and the results are consistent with Nakagawa et al. [23], who indicated 10 times higher amount of SF in the sprouts in comparison to mature plants. Our results are also similar to López-Cervantes et al. [24], while the results of Moon et al. [2] for 6 days sprouts (222.6 mg/kg fw) and Nakagawa et al. [23] for sprouts from the market (1153 mg/100 g dw) were higher than ours. Such differences may be due to the plant variety or growing and sprouting condition. Our results for SF content in broccoli florets comply with Nakagawa et al. [23] for broccoli from Japan (69.1–171.3 mg/100 g dw) and Campas-Baypoli et al. [25], for lyophilized florets from Mexico (31–45.4 mg/100 g dw). Many studies, including our results, proved that broccoli sprouts are better source of SF than the florets [23, 24], which implies their use as dietary prevention of different health problems.
Fatty Acid Profile
Fatty acid analysis was performed by means of gas chromatography. Preliminary qualitative GC analysis of broccoli sprouts extracts indicated the presence of caproic, myristic, palmitic, stearic, arachidic, lignoceric, palmitooleic, oleic, linoleic, α-linolenic, eicosenoic, erucic and docosadienoic acids, while in broccoli florets caproic, capric, palmitic, stearic, lignoceric, oleic, linoleic, eicosenoic and erucic acids were detected. For detailed comparison of the tested materials, the percentage of saturated (SFAs) and unsaturated (UFAs) fatty acids was further determined by GC. SFAs in broccoli sprouts and florets consisted 10.5 and 77% of total pool of fatty acids, respectively, with palmitic (5.7%) and stearic (2.8%) acids dominating in sprouts, and caproic (34.9%), stearic (15.9%) and palmitic (12%) acids in florets (Table 1). Qualitative fatty acids profile in both evaluated materials was similar to Ahmed et al. [26] for Brassica juncea and López-Cervantes et al. [24] for broccoli sprouts. We found the relative content of UFAs in the analyzed broccoli sprouts to be 88%, with the predominant oleic acid (45.5%) but linoleic (20.8%) and α-linolenic acids (17.0%) on similar level. In broccoli florets oleic acid was also the main UFA (12.4%), but in significantly lower amount in comparison to the sprouts. The results of López-Cervantes et al. [24] indicated linoleic (26.31–31.88%) and α-linolenic acids (21.34–28.69%), with lower amount of oleic acid (5.17–11.70%) in broccoli sprouts. What is particularly important, our results revealed significantly higher total UFAs content in the sprouts in comparison to the florets, with very low amounts of harmful erucic [27] acid in sprouts (0.5%) and florets (2%), in comparison to the broccoli seeds (38% - data not shown). Another study of three South Korea broccoli cultivars revealed much lower content of total SFAs, and higher amount of UFAs, with the predominance of palmitic and linolenic acids, in the florets, when compared to our results [28].
Cytotoxic Activity
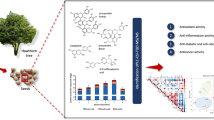
Cytotoxic impact of the tested broccoli products was performed by MTT assay against cancer HepG2 and SW480, but also normal BJ cells, to determine the potential selectivity. The tested samples varied in their influence on the examined cell lines, in dose dependent manner. Both broccoli sprouts and florets caused significant decrease in SW480 and HepG2 cancer cells viability (Fig. 1), with more profound effect for the former and less susceptibility of the latter. The cytotoxic effect of broccoli florets was stronger for both cancer cell lines in comparison to the sprouts. It is worth to note that the latter were also more selective against cancer cells, as opposed to the florets, affecting also normal fibroblasts. Such effects may result from the differences in phytochemical content of both products. Broccoli sprouts with the predominance of polyphenolics and SF, known from their antioxidant and protective effects [1, 3], actually did not affect normal fibroblasts, while the opposite results were observed for the florets, with lower content of the mentioned protective compounds. Stronger cytotoxic effect against cancer cells by broccoli florets, especially in higher concentration, may result from the presence of SFAs, and the effect was also observed by Lima et al. [29]. Similar experiment on cytotoxic activity of broccolini (Brassica oleracea Italica x Alboglabra), a hybrid between broccoli and Gai Lan, on four human cancer cell lines (SW480, HepG2, Hela, and A549) was performed by Wang & Zhang [30] and the results showed a dose-dependent antiproliferative properties of broccolini leaves extract. Cytotoxic activity of broccoli sprouts was previously studied on human leukemic cell line (HL-60), with no significant effects on proliferation or viability [31] and on two murine prostate cancer cell lines, AT2 and Mat-Ly-Lu, with interesting effect, correlated with antioxidant activity [22]. Broccoli florets extracts were tested on human ovary (OVCAR-5), breast (MCF-7), colon (Colo-205) and prostate (PC-3) cancer cells [23] and significant effect on human lung cancer cells A-549 was also described [32]. All the results indicate that broccoli sprouts and florets may be considered as potent cancer chemopreventive agents.

Cytotoxic activity of methanol broccoli sprouts (a) and florets (b) extract on BJ, SW480, HepG2 cells (left two diagrams) and apoptosis/necrosis induction of the 2.5 mg/mL extracts (right two diagrams). Cells were cultured in the presence (0.25–2.5mg/mL) of dry broccoli extracts. Cell viability was expressed as % of control (untreated) cells. Values represents means ±SEM; each experiment was done in triplicate. Means with the same symbols on each subdiagram differ significantly (p < 0.05)
Apoptosis and Necrosis
To further investigate the impact of the tested broccoli sprouts and florets on the cells, their proapoptotic potential was evaluated by means of flow cytometry (Fig. 1). Strong cytotoxic effect of broccoli florets on SW480 cells, described above, proved to be due to necrosis (46.6%) rather than apoptosis (32.7%), while the opposite effect was observed for HepG2 (20.9 and 51.9%, respectively) and for normal fibroblasts (2.1 and 51.9%, respectively). On the contrary, the tested broccoli sprouts, with minor cytotoxic effect, stimulated apoptosis in SW480 (77.4%) and necrosis (50%) in HepG2 cancer cells, and their slight impact on normal fibroblasts was due to proapoptotic effect (20.9%). Our findings for broccoli sprouts were also proved for SW480 cells by Wang & Zhang [30] and Hudson et al. [33], but also for human bladder carcinoma cells [34]. Our study is probably the first to demonstrate significant differences between mechanism of cell death between dry broccoli sprouts and florets. Such strong effect of broccoli sprouts on SW480 cells in comparison to florets may expand dietetic recommendation for chemoprevention of colorectal cancer, though further in vitro and in vivo studies are needed. The statistically significant HPCA model of two significant components was derived. Table 1S shows the details of PCA models for blocks X1 and X2 as well as for overall HPCA model. Block X1 was described completely by only one principal component which accounts for 99.2% of the variation in this block. All variables in this block had similar absolute values of their loadings (around 0.35), but for C16:0, C18:0, C24:0, C20:1 and C22:1 with positive, while for C18:1 and C18:2 with negative signs, which therefore indicates inverse correlation between the last two and the rest of fatty acids as a whole group. Figure 1S shows HPCA final plot for the first two components. The first principal component in this model had positive loadings for APOPT-SW480, SF, sinapic acid, and first principal component from the block X2, which all were interrelated with correlation weighs of ca. 0.13–0.14, and simultaneously correlated negatively with APOPT-BJ, APOPT-HEPG2 and principal component from the block X1 (again with correlation weighs with absolute values in the same range as above). Being in one cluster of variables (APOPT-BJ, APOPT-HEPG2 and principal component from the block X1) means also that APOPT-BJ and APOPT-HEPG2 were positively interrelated with fatty acids C16:0, C18:0, C24:0, C20:1 and C22:1, and negatively with C18:1 and C18:2. Such relationships suggest that sinapic acid, SF and two fatty acids (C18:1 and C18:2) inhibit apoptosis of BJ and HepG2, while other fatty acids promote it. Second principal component in HPCA was solely loaded by second principal component from X2 block predominantly influenced by NECRO-BJ and NECRO-HEPG2. The separation between BS and BF samples in HPCA model was evident in direction determined only by the first principal component of HPCA model. Considering this result, BS samples were characterized by apparently higher values of sinapic acid, SF, C18:1, C18:2, MTT-BJ-2.5, and MTT-HEPG2–2.5, and lower values of APOPT-BJ, APOPT-HEPG2, C16:0, C18:0, C24:0, C20:1 and C22:1 as compared with BF samples. This is consistent with results delivered by t-test analysis.
Conclusions
The results of our study provide new and more precise insights into phytochemical content and potential health benefits of broccoli sprouts and florets. Lyophilized sprouts and florets do substantially differ in their polyphenolic, sulforaphane and fatty acid profile and amounts. Moreover, the examined sprouts exhibited significant, selective cytotoxic and proapoptotic effect against colorectal cancer, without a toxic impact on normal cells. Such properties may be a strong argument for recommending lyophilized sprouts as functional broccoli product, with chemopreventive potential against large intestine tumor.
References
Herr I, Büchler MW (2010) Dietary constituents of broccoli and other cruciferous vegetables: implications for prevention and therapy of cancer. Cancer Treat Rev 36(5):377–383. https://doi.org/10.1016/j.ctrv.2010.01.002
Moon JK, Kim JR, Ahn YJ, Shibamoto T (2010) Analysis and anti-Helicobacter activity of sulforaphane and related compounds present in broccoli (Brassica oleracea L.) sprouts. J Agric Food Chem 58(11):6672–6677. https://doi.org/10.1021/jf1003573
Evans PC (2011) The influence of sulforaphane on vascular health and its relevance to nutritional approaches to prevent cardiovascular disease. EPMA J 2(1):9–14. https://doi.org/10.1007/s13167-011-0064-3
Akhlaghi M, Bandy B (2010) Dietary broccoli sprouts protect against myocardial oxidative damage and cell death during ischemia-reperfusion. Plant Foods Hum Nutr 65(3):193–199. https://doi.org/10.1007/s11130-010-0182-4
Steevens J, Schouten LJ, Goldbohm RA, van den Brandt PA (2011) Vegetables and fruits consumption and risk of esophageal and gastric cancer subtypes in the Netherlands Cohort Study. Int J Cancer 129(11):2681–2693. https://doi.org/10.1002/ijc.25928
Wu QJ, Yang Y, Vogtmann E, Wang J, Han LH, Li HL, Xiang YB (2012) Cruciferous vegetables intake and the risk of colorectal cancer: a meta-analysis of observational studies. Ann Oncol 24(4):1079–1087. https://doi.org/10.1093/annonc/mds601
Torres-Contreras AM, Nair V, Cisneros-Zevallos L, Jacobo-Velázquez DA (2017) Stability of bioactive compounds in broccoli as affected by cutting styles and storage time. Molecules 22(4):636. https://doi.org/10.3390/molecules22040636
Villarreal-García D, Alanís-Garza PA, Cuéllar-Villarreal MDR, Redondo-Gil M, Mora-Nieves JL, Jacobo-Velázquez DA (2015) Effects of different defrosting methods on the stability of bioactive compounds and consumer acceptability of frozen broccoli. CyTA J Food 13(2):312–320. https://doi.org/10.1080/19476337.2014.971347
Jamie P, Saltveit ME (2002) Postharvest changes in broccoli and lettuce during storage in argon, helium, and nitrogen atmospheres containing 2% oxygen. Postharvest Biol Technol 26(1):113–116. https://doi.org/10.1016/S0925-5214(02)00006-6
Paśko P, Krośniak M, Prochownik E, Tyszka-Czochara M, Fołta M, Francik R, Sikora J, Malinowski M, Zagrodzki P (2018) Effect of broccoli sprouts on thyroid function, haematological, biochemical, and immunological parameters in rats with thyroid imbalance. Biomed Pharmacother 97:82–90. https://doi.org/10.1016/j.biopha.2017.10.098
Alvarez-Jubete L, Valverde J, Patras A, Mullen AM, Marcos B (2014) Assessing the impact of high-pressure processing on selected physical and biochemical attributes of white cabbage (Brassica oleracea L. var. capitata alba). Food Bioprocess Technol 7(3):682–692. https://doi.org/10.1007/s11947-013-1060-5
Pasko P, Gdula-Argasinska J, Podporska-Carroll J, Quilty B, Wietecha-Posluszny R, Tyszka-Czochara M, Zagrodzki P (2015) Influence of selenium supplementation on fatty acids profile and biological activity of four edible amaranth sprouts as new kind of functional food. J Food Sci Technol 52(8):4724–4736. https://doi.org/10.1007/s13197-014-1602-5
Makowska-Wąs J, Galanty A, Gdula-Argasińska J, Tyszka-Czochara M, Szewczyk A, Nunes R, Carvalho IS, Michalik M, Paśko P (2017) Identification of predominant phytochemical compounds and cytotoxic activity of wild olive leaves (Olea europaea L. ssp. sylvestris) harvested in south Portugal. Chem Biodivers 14(3). https://doi.org/10.1002/cbdv.201600331
Paśko P, Okoń K, Krośniak M, Prochownik E, Żmudzki P, Kryczyk-Kozioł J, Zagrodzki P (2018) Interaction between iodine and glucosinolates in rutabaga sprouts and selected biomarkers of thyroid function in male rats. J Trace Elem Med Biol 46:110–116. https://doi.org/10.1016/j.jtemb.2017.12.002
Adach A, Daszkiewicz M, Tyszka-Czochara M, Barszcz B (2015) New oxovanadium (IV) complexes with pincer ligand obtained in situ: experimental and theoretical studies on the structure, spectroscopic properties and antitumour activity. RSC Adv 5:85470–85479. https://doi.org/10.1039/C5RA12561H
Tyszka-Czochara M, Pasko P, Zagrodzki P, Gajdzik E, Wietecha-Posluszny R, Gorinstein S (2016) Selenium supplementation of amaranth sprouts influences betacyanin content and improves anti-inflammatory properties via NFκB in murine RAW 264.7 macrophages. Biol Trace Elem Res 169:320–330. https://doi.org/10.1007/s12011-015-0429-x
Tyszka-Czochara M, Konieczny P, Majka M (2017) Caffeic acid expands anti-tumor effect of metformin in human metastatic cervical carcinoma HTB-34 cells: implications of AMPK activation and impairment of fatty acids de novo biosynthesis. Int J Mol Sci 18(2):462–478. https://doi.org/10.3390/ijms18020462
Janne K, Pettersen J, Lindberg NO, Lundstedt T (2001) Hierarchical principal component analysis (PCA) and projection to latent structure (PLS) technique on spectroscopic data as a data pretreatment for calibration. J Chemom 15:203–213. https://doi.org/10.1002/cem.677
Cartea ME, Francisco M, Soengas P, Velasco P (2010) Phenolic compounds in Brassica vegetables. Molecules 16(1):251–280. https://doi.org/10.3390/molecules16010251
Pająk P, Socha R, Gałkowska D, Rożnowski J, Fortuna T (2014) Phenolic profile and antioxidant activity in selected seeds and sprouts. Food Chem 143:300–306. https://doi.org/10.1016/j.foodchem.2013.07.064
Fernández-León MF, Fernández-León AM, Lozano M, Ayuso MC, González-Gómez D (2012) Identification, quantification and comparison of the principal bioactive compounds and external quality parameters of two broccoli cultivars. J Funct Foods 4(2):465–473. https://doi.org/10.1016/j.jff.2012.02.005
Gawlik-Dziki U, Jeżyna M, Świeca M, Dziki D, Baraniak B, Czyż J (2012) Effect of bioaccessibility of phenolic compounds on in vitro anticancer activity of broccoli sprouts. Food Res Int 49(1):469–476. https://doi.org/10.1016/j.foodres.2012.08.010
Nakagawa K, Umeda T, Higuchi O, Tsuzuki T, Suzuki T, Miyazawa T (2006) Evaporative light-scattering analysis of sulforaphane in broccoli samples: quality of broccoli products regarding sulforaphane contents. J Agric Food Chem 54(7):2479–2483. https://doi.org/10.1021/jf051823g
López-Cervantes J, Tirado-Noriega LG, Sánchez-Machado DI, Campas-Baypoli ON, Cantú-Soto EU, Núñez-Gastélum JA (2013) Biochemical composition of broccoli seeds and sprouts at different stages of seedling development. J Food Sci Technol 48(11):2267–2275. https://doi.org/10.1111/ijfs.12213
Campas-Baypoli ON, Sánchez-Machado DI, Bueno-Solano C, Ramírez-Wong B, López-Cervantes J (2010) HPLC method validation for measurement of sulforaphane level in broccoli by-products. Biomed Chromatogr 24(4):387–392. https://doi.org/10.1002/bmc.1303
Ahmed S, Hassan MH, Kalam MA, Rahman SA, Abedin MJ, Shahir A (2014) An experimental investigation of biodiesel production, characterization, engine performance, emission and noise of Brassica juncea methyl ester and its blends. J Clean Prod 79:74–81. https://doi.org/10.1016/j.jclepro.2014.05.019
Wendlinger C, Hammann S, Vetter W (2014) Various concentrations of erucic acid in mustard oil and mustard. Food Chem 153:393–397. https://doi.org/10.1016/j.foodchem.2013.12.073
Bhandari SR, Kwak JH (2015) Chemical composition and antioxidant activity in different tissues of Brassica vegetables. Molecules 20(1):1228–1243. https://doi.org/10.3390/molecules20011228
Lima TM, Kanunfre CC, Pompeia C, Verlengia R, Curi R (2002) Ranking the toxicity of fatty acids on Jurkat and Raji cells by flow cytometric analysis. Toxicol in Vitro 16(6):741–747. https://doi.org/10.1016/S0887-2333(02)00095-4
Wang B, Zhang X (2012) Inhibitory effects of Broccolini leaf flavonoids on human cancer cells. Scanning 34(1):1–5. https://doi.org/10.1002/sca.20278
Chaudhary A, Rampal G, Sharma U, Thind TS, Singh B, Vig AP, Arora S (2012) Anticancer, antioxidant activities and GC-MS analysis of glucosinolates in two cultivars of broccoli. Med Chem Drug Disc 2(2):30–37
Farag MA, Motaal AAA (2010) Sulforaphane composition, cytotoxic and antioxidant activity of crucifer vegetables. J Adv Res 1(1):65–70. https://doi.org/10.1016/j.jare.2010.02.005
Hudson EA, Howells LM, Gallacher-Horley B, Fox LH, Gescher A, Manson MM (2003) Growth-inhibitory effects of the chemopreventive agent indole-3-carbinol are increased in combination with the polyamine putrescine in the SW480 colon tumour cell line. BMC Cancer 3(1):1–18. https://doi.org/10.1186/1471-2407-3-2
Tang L, Zhang Y, Jobson HE, Li J, Stephenson KK, Wade KL, Fahey JW (2006) Potent activation of mitochondria-mediated apoptosis and arrest in S and M phases of cancer cells by a broccoli sprout extract. Mol Cancer Ther 5(4):935–944. https://doi.org/10.1158/1535-7163
Acknowledgements
This study was supported by the following grants from the Polish Ministry of Science and Higher Education: K/DSC/000037; 001418; 000805, K/ZDS/001294; 005562. The Uniflora Company (Częstochowa, Poland) support idea of investigation of broccoli sprouts and florets in in vitro models.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflicts of Interest
The authors declare that they no conflict of interest.
Electronic Supplementary Material
ESM 1
(DOCX 36 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Paśko, P., Tyszka-Czochara, M., Galanty, A. et al. Comparative Study of Predominant Phytochemical Compounds and Proapoptotic Potential of Broccoli Sprouts and Florets. Plant Foods Hum Nutr 73, 95–100 (2018). https://doi.org/10.1007/s11130-018-0665-2
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11130-018-0665-2