
Abstract
Aims
The lemon cultivation methods and techniques are crucial to ensure maximum productivity in the face of climate change. Mulching with plastic is commonly used in citrus production for saving water, but some side effects need to be investigated. In our study, we investigated different plastic and biological mulching on lemon trees determining growth and physiological parameters in relation to soil chemical and biological composition.
Methods
The experiment was divided into four different lines with ten trees per treatment, the effect of mulching with white and black plastic film, dry pruning mulching respect to a non-mulched treatment of lemon tree orchard during a crop season. The impact of these treatments on vegetative growth, stomatal gas exchange and mineral nutrition on plant and soil bacterial communities were evaluated.
Results
Our results showed that the type of mulching significantly influenced in the parameters studied. All mulching treatments increased temperature and soil moisture levels; plastic mulching treatments had significantly higher values in terms of intrinsic water use efficiency; while mulching with dry pruning showed higher soil microbial activity, leading to increased water use efficiency and productivity.
Conclusion
The results showed that different methods of mulching affected the physiology of lemon trees interacting in a complex way to determine their growth. Specifically, mulching using dry pruning improved the exchange of gases in the plant and plant nutrition which was related to the biological soil health.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The lemon tree (Citrus x limon) (L.) Osbeck is one of the most important commercial plants of the Rutaceae family, with a worldwide distribution. The main producers are India, Mexico, China, Argentina, Brazil, and Spain (Smilanick et al. 2019). The primary citrus cultivation areas in Spain are in semi-arid regions of the east and south, namely the Region of Murcia and the Valencian Community, where cultivation methods and techniques are crucial to ensuring maximum productivity (Georgiou and Gregoriou 1999). These regions suffer from desertification, a complex and dynamic process of land degradation caused by the interaction between climate change and intensive farming (Prăvălie 2021).
Climate change in the Mediterranean area is causing alterations in precipitation, resulting in lower amounts due to an increase in intensity but shorter durations (Nicholson et al. 2018). Consequently, water availability for irrigation will be drastically reduced. Low water availability has been reported to reduce leaf gas exchange, leaf expansion and nutrient uptake (Bista et al. 2018) leading to reductions in crop growth and yields (Aliche et al. 2018).
The availability of water for agriculture will decrease, and the increase in temperatures will imply a greater need for water, along with an extension of the crop irrigation period (Torelló-Sentelles and Franzke 2022). Currently, several soil moisture conservation techniques are employed in agriculture, such as mulching, generally considered to prevent water loss, weed suppression, that improve and increase crop yield in agricultural production (Lamont 2005; Kader et al. 2017). Mulching techniques used materials such as plastic film, paper, straw, mineral-based material or woodchips for ground cover (Lamont 2005). However, the most commonly used material is plastic, as it is highly resistant compared to others (Crawford and Quinn 2017). Different colours of plastic have various effects on the hydrothermal environment of the soil and crop growth due to their distinct physical properties (Zhang et al. 2023). Black plastic mulching, for instance, absorbed more than 90% of solar radiation, warming the soil (Strik et al. 2006), while white plastic mulching reflected a high proportion of solar radiation, decreasing soil temperature and increased canopy light intensity and air temperature (Andreotti et al. 2010). On the other hand, organic mulching, made of any bulk material placed on the soil surface, has been also used for the purpose to retain and minimize water loss, and they also improved soil physical characteristics, enhancing canopy microclimate (Iqbal et al. 2020). However, little is known about the effect of mulching on soil microbiological and mineral availability changes.
Soil microorganisms drives many aspects of biogeochemical carbon and nutrient cycling, water holding capacity, water purification, pathogen control and climate change mitigation, key aspects of plant development (Dangi 2014). Most soil processes are mediated by the biodiversity of soil microorganisms in direct relation to the physic-chemical properties of their environment (Lemanceau et al. 2015).
Soil bacteria play a crucial role in cultivated soils and are related to crop production (Davison 1988), since the interactions between plants and bacteria in the rhizosphere, the area surrounding plant roots, significantly influence plant health and soil fertility. Rhizobacteria also participate in the process of mineral nutrient solubilization (Hayat et al. 2010), contributing to increased resistance to environmental stress, the stability of soil aggregates and the enhancement of soil structure and organic matter content. Rhizobacteria are effective at retaining more organic nitrogen and various nutrients within the plant-soil system which, in turn, reduces the dependency on nitrogen and phosphorus fertilizers and promotes the release of these essential nutrients (Richardson 2001).
In our study, we investigated the effects of different mulches on drip irrigated lemon trees of the variety 'Fino' (Citrus x limon) with fertirrigation and grown in organic modality. We compared mulch with white/black plastic film and mulch with dry pruning crushed, with no mulching as control. The objective of the research was to determine their effects on the soil temperature, water content and nutrient availability in relation to leaf gas exchange and mineral concentration in lemon leaves on growth. Also, the soil bacteria community was determined and its relationship with the other parameters studied.
Materials and Methods
Location and plant growth conditions
The experimental farm “Cañada Honda” is located near the village of Librilla (Region of Murcia, Spain). It is a semi-arid area (Mediterranean climate), with an average annual rainfall of 300–350 l/m2 and average annual temperatures of 18.8 ºC (Elvira-Rendueles et al. 2019).
Seven-year-old lemon trees of the variety "Fino" (Citrus x limon) with drip irrigation were used. This variety is characterized by a large harvest from November to March. The lemon trees are grown with organic methods; being certified 100% organic in 2021. The soil showed a pH of 8.66 ± 0.02 and an electrical conductivity (EC) of 139.33 ± 23.26 µS cm−1. Each tree was irrigated with 2 drippers twice a week, for 90–120 min and the water flow rate was set at 4 l/h per dripper. The crop was organic with manure and organic fertilization as reported previously (Olmos et al. 2024).
The experiment was divided into four different lines with ten trees per treatment, with a 3 × 5 crop frame, comparing soil cover with white/black plastic film mulch, mulching with crushed dry pruning and an outdoor control over a period of six months. Samples were collected between March-July 2023. To make the sampling representative of the crop, samples were randomly taken from five different trees, avoiding crop edge.
The white and black plastic (130 g m−2) were composed of semi-impermeable polypropylene geotextile; while the dry pruning was obtained from the pruning of lemon trees. Dry pruning was used after open air dried, crushed with a tract, and stored for 3 months in the open air before being used as mulch.
Plant and Physiological analysis
-
a)
Vegetative growth
Ten measurements of the new leaf area were made with an interval of 15 days between each measurement from March to July, selected as points represented on the graph in the months of March, May and July. Three leaves on each plant from each of the treatments were previously selected and marked. This procedure was carried out in situ by drawing the outline of the leaf on a sheet of paper and subsequently using the ImageJ program (Tsung-Luo 2017) to calculate the area, and then obtain the relative growth rate with the following equation: RGR = (Ln A2-Ln A1) / (t1-t2), (cm2 cm−2 day −1). The measurement of lemon fruit growth was calculated by measuring the length and diameter with a caliper.
-
b)
Stomata content
Stomatal printing on the new leaves of the tree was carried out to count how many stomata were open or closed on the leaf per unit area. To do this, the surface of the underside of the leaf was impress on a slide with adhesive tape (composed of cellulose acetate), digested with a drop of acetone. In this way, the entire surface of the leaf was impressed on the adhesive tape. Subsequently, the impressions were observed under an optical microscope (OLYMPUS U-CMAD3, Olympus Corporation, Tokyo, Japan) and a counted with an ImageJ analysis program (Jinn 2017). Five plates of new leaves were obtained for each of five trees per treatment.
-
c)
Photosynthetic parameters
Photosynthetic capacity (An, μmol CO2 m−2 s−1), stomatal conductance (Gs, mol H2O m−2 s−1), internal content of CO2 (Ci, μmol mol⁻1) were measured in fully-expanded new leaves using a TPS-2 Portable Photosynthesis System (PP Systems, Inc., Amesbury, MA, USA). Intrinsic water use efficiency (WUEi μmol CO2 mol−1 H2O) was calculated by dividing the net photosynthetic rate and the stomatal conductance. Five new leaves were measured for each of the five trees in each treatment.
-
d)
Leaves macro and micro mineral content
The macro and micro mineral contents were analysed using Inductively Coupled Plasma-Optical Emission Spectrometry (ICP-OES) on a Thermo ICAP 6500 Duo instrument (Thermo Fisher Scientific, Waltham, MA, USA). Leaves were collected, dried, and ground into a fine powder. A total of 200 mg of each sample was added to a 25 mL tube along with a mixture of 4 mL of 68% purity HNO3 and 1 mL of 33% purity H2O2 for digestion. Additionally, a Teflon reactor contained 300 mL of high-purity de-ionized water, 30 mL of 33% purity H2O2, and 2 mL of 98% purity H2SO4 was added. The microwave heating digestion program consisted of three steps: starting at 20 ºC and 40 bar, increasing by 10 bar per minute for30 min until reaching 220 ºC, and maintaining the temperature at 220 ºC for 20 min. After cooling, the mineralized samples were transferred to 10 mL (for micro minerals) and 25 mL (for macro minerals) double gauge tubes, and the volume was adjusted using high-purity de-ionized water. Calibration standards were prepared using a multi-mineral standard solution supplied by SCP Science (Quebec, Canada).
Soil analysis
-
a)
Soil macro and micro mineral content
It was obtained by using the same above method that the one for macro and micro mineral content of leaves.
-
b)
Soil available Carbon, Nitrogen and Phosphorus
Available carbon and nitrogen in the soil samples was determined through elemental analyzer (C/N Flash EA 112 Series- Leco Truspec). Eight grams of soil samples previously sieved through 2 mm mesh were extracted with 100 mL of distilled water for 1 h in an automatic shaker. After that it was centrifuged at 3500 rpm for 10 min and the supernatant was filtered through Whatman No. 2 filter paper to obtain a clear filtrate that it was used to do the determination of available carbon and nitrogen.
Available phosphorus in the soil samples was determined using the Olsen method. Two grams of sieved air-dried soils was extracted with 100 mL of 0.5 M sodium bicarbonate (NaHCO₃) solution at pH 8.5. The mixture was shaken for 30 min, followed by filtration through Whatman No. 2 filter paper. The phosphorus concentration in the clear filtrate was quantified colorimetrically using the molybdenum blue method, with absorbance measured at 880 nm. Calibration was performed using standard phosphorus solutions to ensure accurate quantification.
-
c)
pH and apparent densit
Soil pH was measured in a 1:10 (w/v) water-soluble extract, after soil shaking for 60 min, using a pH meter (Crison pHmeter, micropH 2001). The dry apparent density was determined from the Spanish standard UNE-EN 13041–2012, by means of gravimetry and particle density was measured by calcination at 540ºC.
-
d)
Soil temperature and moisture
The soil temperature was measured using a precision thermometer (Precision Plus, ETI Ltd, Worthing, West Sussex, United Kingdom) obtaining six measurements (three being in the superficial part of the soil under the tarp and the other three at 15 cm from depth) from each of the selected sampling point of each of the five selected trees in each treatment, at 15 day-intervals between each measurement from March to July.
To determine the moisture content, a determined quantity of soil was weighted and placed in an oven with temperature range of 110 ± 5 ºC for about 24 h. After that, the difference in the wet mass and dry mass of the soil was the water content of the soil. It was determined by triplicate per treatment.
-
e)
Soil biological parameters
For this purpose, dehydrogenase activity and Bacterial community was conducted exclusively at the sampling time of July, coinciding with the culmination of the growing season and the harvest of the lemon fruits, a period that marks the end of the primary physiological activities of the trees. Additionally, July typically experiences the highest temperatures, which can induce significant stress on both soil biological parameters and tree physiology. These conditions were expected to provide the most informative snapshot of the microbial dynamics under peak environmental and physiological stress conditions.
Dehydrogenase activity
Dehydrogenase activity in the soil was determined using a colorimetric procedure according to (von Mersi and Schinner 1991). Briefly, 2 mL of 2-(4-iodophenyl)-3-(4-nitrophenyl)-5-phenyltetrazolium chloride (INT, 0.015 M) were added to 2 g of soil and then homogenized and incubated at 25 ºC for 4 h in the dark. Subsequently, 8 mL of acetone were added to all samples and put them on an orbital shaker (250 rpm) for 1 h in the dark. Iodonitrotetrazolium formazan (INTF) was determined in the centrifuged extracts by measurement at 485 nm spectrophotometrically. The dehydrogenase activity was expressed as nmol INTF g−1 h−1.
Bacteria community
This parameter comprises the soil DNA extraction, amplification of the representative bacterial sequences and the further deep analysis of them by taxonomic characterization and grouping them by using through alpha and beta diversity and Principal Coordinates Analysis (PCoA) analysis. Their relationship with the rest of physical, physico-chemical and chemical soil parameters were analyzed by using the Distance-based redundancy analysis (db-RDA).
DNA was extracted from soil samples (500 mg) using the DNeasy Power Soil Pro Kit (Qiagen) following the manufacture’s protocol. The quantity and quality of the DNA extracts were quantified using a Nano Drop 2000 fluorospectrometer (Thermo Fisher Scientific, Waltham, MA, USA).
As stablished in the Molecular Biology Service at the University of Murcia, purified DNA was used as the template for generating a 16S rRNA gene library. The oligonucleotide primers used for this experiment were 5′-TCGTCGGCAGCGTCAGATGTGTATAAGAGACAGCCTACGGGNGGCWGCAG-3′ and 5′-GTCTCGTGGGCTCGGAGATGTGTATAAGAGACAGGACTACHVGGGTATCTAATCC3′, where the underlined regions are the Illumina adapter overhang nucleotide sequences, while the non-underline sequences are locus-specific sequences targeting conserved regions within the V3 and V4 domains of prokaryotic 16S rRNA genes (Klindworth et al. 2013). The amplified fragments were quantified with the Qubit dsDNA HS Assay Kit (Invitrogen, Merelbeke, Belgium) on a Qubit 2.0 Fluorometer prior to sequencing. Paired-end sequencing of the library was performed on an Illumina MiSeq sequencer (San Diego, CA, USA) using the MiSeq Reagent Kit (v3) with the longest read length set to 2 × 300 base pairs (bp). Library qualities were estimated using the Bioanalyzer High Sensitivity DNA Analysis Kit (Agilent).
The 16S-V4 sequencing library was first reviewed with FastQC for overall quality assessment, and the libraries were processed in R package DADA2 (v.1.8.0) (Callahan et al. 2016). Reads were quality trimmed with the “filterAndTrim” function with “maxEE (2,5)”. Reads below 165 bp after the trimming process were discarded. Errors learned from all samples were used for sample inference with the dada2 algorithm by employing an evaluation of 1E8 bases. Forward and reverse reads are merged below to generate a table of sequences, and the resulting Amplicon Sequence Variants (ASVs) were subjected to de novo chimera detection, using DADA2 and any artifacts were removed.
For bacteria taxonomic assignment, ASVs were queried against the SILVA database v.132 using IDTAXA (Murali et al. 2018) implemented in the R package DECIPHER (Wright 2016) with a threshold 40. Sequences identified as non-bacterial were discarded. Similarly, to numerous recently published studies, we chose to forego rarefaction of our samples as it increases uncertainty in relative abundances (McMurdie and Holmes 2013).
The abundance matrix, the taxonomy assignment and the metadata obtained from samples were merged and imported with the phyloseq v3.12 package (McMurdie and Holmes 2013) to produce the alpha and beta bacterial diversity. The "prune_taxa" function was used to keep only subsystems with absolute abundance > 0.01%. Counts were normalized in each sample using the median sequencing depth, and phylum and class level plots were created using the ggplot2 and ggpubr packages. Alpha diversity was calculated in R using the phyloseq package, and several alpha indices were generated, such as Shannon and Simpson, using the function “plot_richness”. Beta diversity was calculated using weighted and unweighted Unifrac distances (Lozupone and Knight 2005). To test for significant differences in community composition among different seasons, permutational multivariate analysis of variance using distance matrices (PERMANOVA) was conducted using the Adonis function in the R package vegan with 999 permutations, and the results were visualized by Principal Coordinates Analysis (PCoA).
UpSet plots were created using the upsetR package version 1.4.0. This was done by transforming the data frame of average counts for each soil sample into a data frame that exclusively contained 0 and 1 values. Subsequently, the data was arranged based on the frequency of intersection size.
Finally, db-RDA analysis was conducted to identify soil physicochemical properties with a significant impact on soil bacterial communities across different factors using the dbrda function of the R vegan package vegan v2.6–1. Parameters that significantly explained variation in the bacterial community were identified using forward selection (the ordistep function of the vegan package) with p value < 0.05.
-
f)
Statistical analysis
Statistical analyses were performed using the SPSS 29.0.0.0 (241). Significant differences between the values from all parameters were determined at p ≤ 0.05, according to a one-way ANOVA followed by Duncan’s test. For the studies of alpha diversity, also Student test was used and for the beta diversity, also Adonis was used. All the results are presented as the mean ± SE.
Results
Vegetative growth
The Relative Growth Rate (RGR) of leaves in March did not show significant differences between treatments (Fig. 1a), but in May, the RGR of black plastic mulching showed a significant increase respected to the dry pruning and control, but not of white plastic. In any time, dry pruning mulching did not show significant differences respect to the control. However, in July, RGR for both plastic treatments black and also white showed a significant decrease compared with control and dry pruning treatments.

a Leaf Relative Growth Rate (RGR) (cm2 cm−2 day−1) in the different treatments (control, black plastic, white plastic and dry pruning) in March, May and July, and (b) number of fruits per tree in July. The statistics in RGR were performed at each time, for study the mulching (represented with lowercase letters) and for study the time on a specific treatment (represented with uppercase letters). Significant differences were determined between the values of the treatment parameters at p ≤ 0.05, according to a one-way ANOVA followed by Duncan's test
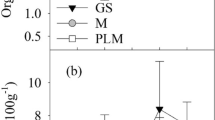
The number of fruits per tree harvested in July (Fig. 1b) showed a significant increase in dry pruning and black plastic compared with the white plastic and control treatments, which exhibited similar lower values. Furthermore, when the fruit relative growth was measured in July, no differences between treatment were found (data not shown).
Stomata content and WUEi
The total stomata per mm2, percentage of open-closed stomata and leaf humidity were measured. The number of stomata (Fig. 2a) did not show significant differences in March; but in May, a significant increase was observed in the dry pruning treatment compared with the black plastic treatment, obtaining no significant differences between black plastic, white plastic treatment and control. In July, the number of stomata was significantly increased in the treatments with black and white plastic compared with control, but no significant differences were observed with dry pruning.

Results of (a) total stomata mm−2, b % leaf humidity, c % open and % closed stomata in the different treatments (control, black plastic, white plastic and dry pruning) in March, May and July. The statistics were performed at each time, for study the mulching (represented with lowercase letters) and at each treatment, for study the time (represented with uppercase letters); Significant differences were determined between the values of the treatment parameters at p ≤ 0.05, according to a one-way ANOVA followed by Duncan's test
Regarding to leaf humidity (Fig. 2b), no significant differences in the months of March and May were observed. However, some significant differences were observed among all treatments in July. In general, white plastic mulching exhibited the highest value of % leaf moisture while the black plastic treatment displayed the lowest value.
Respected to the percentage of open stomata respect to the total (Fig. 2c) different behaviour was observed attending to the sampling time. In March, it showed a significant decrease in black plastic compared with the other treatments, including control, which did not show differences between themselves, In May, itincreased in dry pruning compared to the others treatments, which did not show differences among them. In July, dry pruning and white plastic mulching showed a significant increase on this parameter compared with control and black plastic treatments, which did not show differences between them. Finally, regarding the closed stomata as calculated from the same microscope frame, the values were the opposite than open stomata (Fig. 2c).
Leaf gas exchange parameters
The photosynthesis (An), stomatal conductance (Gs), CO2 internal concentration (Ci) were measured and intrinsic water use efficiency (WUEi) was calculated. In March, the An (Fig. 3a) showed a significant increase in the dry pruning mulching respect to the other three treatments which did not show differences between black plastic and control. In May, all treatments showed a significant increase respect to March, and a different trend was observed where the higher values were obtained in black and white plastic compared with control and dry pruning treatments. In July, a significant decrease was observed in control compared with dry pruning treatment, obtaining no significant differences between white and black plastic treatments.

Results of (a) net photosynthetic rate An (μmol CO2 m−2 s−1), b stomata conductance Gs (mol m−2 s−1), c internal CO2 content Ci (μmol mol⁻1) and (d) intrinsic water use efficiency (WUEi) (μmol CO2 mol−1 H2O) in the different treatments (control, black plastic, white plastic and dry pruning) in March, May and July. The statistics were performed at each time, for study the mulching (represented with lowercase letters) and at each treatment, for study the time (represented with uppercase letters); Significant differences were determined between the values of the treatment parameters at p ≤ 0.05, according to a one-way ANOVA followed by Duncan's test
Regarding Gs, it was observed a significant increase in March respect to the other sampling times, showing control and white plastic treatment, similar An significant lower values. In May, Gs did not show significant differences between all treatments. In July, a significant increase was observed in dry pruning compared with the rest. In March, the Ci (Fig. 3c) showed a significant decrease in the dry pruning, white and black plastic treatments, compared to control. In May, control maintained the higher value with significant decreases with dry pruning and while plastic (being this later one the lowest value). In July, only significant decreases were observed in white plastic compared with the rest. Finally, the WUEi in March and May, did not show significant values, while in July a significant increase was observed in black plastic compared to control (Fig. 3d).
Soil pH and apparent density
These two parameters showed expected values from the Mediterranean soil (Rashid and Ryan 2004), for the case of the pH ranged from 7.69 to 7.7 in the soil covered with plastic that it was a slightly higher than control with 7.87 and the soil with dry pruning mulching was 8.55 but in any of the cases were significantly different. The apparent density was a usual value for a Mediterranean and the treatments did not produce any significantly change ranged from 1.16 to 1.26 g cm3.
Temperature and moisture of soil
Temperature and soil moisture were determined (Table 1). The temperatures measured in the different distances from the mulching (under 15 cm into soil, on top of the mulching and 1 m over the mulching) were increasing from March to July, being similar in March and May. All the temperatures were higher in black plastic compared with the rest of mulching treatments. Furthermore, comparing the other treatments, in the month of March, the temperature on soil surface or mulching was similar between white plastic, dry pruning mulching and control; but in May, white plastic showed significantly higher temperatures than control and dry pruning; and in July, the white plastic and dry pruning were significantly lower than control. The temperature at 15 cm into soil was stable at March and May respecting to all treatments. However, in July, white plastic and dry pruning, temperature was significantly lower than control and black plastic. Temperature at 1 m above soil or mulching surface was similar in March and May for control, white and dry pruning. In July, a gradual increase was observed from black plastic, dry pruning, white plastic and control, being the highest significant value for the black plastic treatment, followed by white plastic, dry pruning and control treatments.
The soil moisture, in March, showed no significant differences between treatments, while in May and July the mulching treatments showed significantly higher soil moisture than control. In July, a significant decrease was observed compared to the other months for the four treatments.
Soil and leaves mineral content
Soil
In Table 2, the significant soil macronutrients and micronutrients are shown Values are shown in the same units than in leaves for easier comparation. In March, Ca showed a significant decrease when dry pruning mulching was compared to control, while no differences were found for both plastic treatments. For white plastic, potassium exhibited a decrease in May and an increase in July. Magnesium showed an increase with dry pruning in March and July and with black plastic in May. Phosphorus showed only an increase with all treatments in July. Total N level was significantly increased across all treatments in July, being also remarkable the significant higher N in white plastic mulching in March. Total organic carbon (TOC) showed significantly higher values in the black plastic and dry pruning treatments compared to control for the three sampling times. However, the white plastic treatment did not exhibit significant differences. According to micronutrients, they showed slight alterations in certain times with any of the treatments as B that showed an increase in May and July with dry pruning, Cu increased in March with white plastic and in May with black plastic. Also, Fe exhibited only an increased in July with dry pruning, Mn showed an increase in May and July with black plastic and Zn increased in March with white plastic and in May and July with black plastic. Macronutrients and micronutrients not listed show no significant differences.
Leaves
In Table 3, the significant leaf macronutrients and micronutrients are shown. In May, Ca showed a significant increase with dry pruning compared to the rest of mulching treatments. July displayed a significant increase in black plastic, a decrease in white plastic, both compared to control and dry pruning treatments. Potassium exhibited a significant increase in May with dry pruning compared to the control, black and white plastic treatments, and a significant increase in July with white plastic compared to the control, black and dry pruning treatments, along with a decrease in black plastic compared to the same treatments. In March, Mg showed a significant increase with dry pruning compared to black and white plastic treatments; while only in May and July showed this increase also respect to whole treatments. Phosphorus showed a significant increase with dry pruning in May, and white plastic, and dry pruning in July. Sulphur had a significant increase in May with dry pruning and in July with black plastic. Total N had a significant increase in May with black plastic compared to white plastic, and no significant differences with control and dry pruning. In July, a significant increase was observed with white plastic compared to control, black plastic, and dry pruning, with similar values for control, black plastic, and dry pruning. Carbon did not show significant differences with any of the treatments.
In terms of micronutrients, B showed a significant increase in March with dry pruning, and in July, a significant increase was observed in black plastic. Cupper had a significant increase in May with dry pruning and in July with white plastic, while Fe had only in July a significant increase with black plastic and Mn in May with dry pruning and in July, in black plastic white plastic, and dry pruning. Zinc showed a significant increase in May with dry pruning and in July with black plastic Macronutrients and micronutrients not listed show no significant differences.
Dehydrogenase activity
Dehydrogenase activity was measured once the lemon fruit were harvested at the last sampling time in July (Fig. 4). This enzymatic activity of the different mulching treatments showed a significant increase respect to the control. The dry pruning treatment showed a significant difference respect to the black plastic but not to the white one, while no significant differences were observed between both white and black plastic.

Results of dehydrogenase activity (nmol INTF g−1 dry soil h−1) in the different treatments (control, black plastic, white plastic and dry pruning) in July. The statistics were performed individually for each parameter; significant differences between the values from all parameters were determined at p ≤ 0.05, according to a one-way ANOVA followed by Duncan’s test
In Table 4, the water soluble organic carbon and nitrogen and the available phosphorus in July are shown. Soil organic carbon (SOC) showed a significant increase when dry pruning mulching was compared to control, white and black plastic treatments. Available nitrogen (TN) did not show significant differences among the treatments. Available P showed a significant increase when white plastic, dry pruning and black plastic mulching were compared to control. Apparent density and pH did not show significant differences.
Effect of different mulches on bacterial phyla or Bacterial community composition of different mulching treatments
In the same way than the above soil biological parameter, the bacterial community was analysed from the soil sampling in July (Fig. 5). The predominant identified bacterial phyla, classes and orders are presented in Fig. 5a, b and c respectively. The prevailing phylum under the different covers was Proteobacteria, accounting for an average of 39%, succeeded by Actinobacteria (26%) and Choloroflexi (now named Chloroflexota, 11%). At the class level, Alphaproteobacteria exhibited the highest relative abundance (27.2% in average), encompassing detected orders such as Rhizobiales, Rhodospirillaes (also known as Azospirillales), Rhodobacterales and Sphingomonadales. Actinobacteria (12.62%) constituted the second most abundant class in this bacterial community, featuring identified orders like Micrococcales, Gaiellales and Solirubrobacterales. Lastly, Gammaproteobacteria (12.85%) stood as the third most abundant class, with Pseudomonadales being one of the identified order.

Relative abundance (%) of bacterial community at the phylum (a), class (b) and order (c) in soil samples of the treatments (control, black plastic, white plastic and dry pruning) in July
Alpha diversity indices, including richness and Shannon, are presented in Fig. 6a as regards the first two indices, the dry pruning treatment showed significant higher values compared to the control group, while no significant changes were observed between the white and black plastic treatments and the control.

Alpha diversity indices (a) and beta diversity PCoA (b) based on weighted UniFrac distances of bacterial community found in the soil samples of the different treatments (control, black plastic, white plastic and dry pruning) in July. For the studies of alpha diversity, also Student test was used and for the beta diversity, also Adonis was used
In terms of beta diversity, bacterial PCoA based on weighted (Fig. 6b) UniFrac distances showed that bacterial community structure was altered depending on the applied mulching. As regards the weighted UniFrac data, the two principal PCoA coordinates explained 46.6% of the variations (32.2% and 14.4%, respectively) and exhibited significant changes between the different mulching treatments and the control group (PERMANOVA, p ≤ 0.05).
Bacterial sequences were assigned to 2483 ASVs (Fig. 7), with 1123 of the ASVs, corresponding to 45.23% of the total, shared amongst all treatments. The two groups covered by plastic and the dry pruning treatment shared 308 ASVs with each other, which represents 12.40% of the total. The control group harboured the lowest proportion of unique ASVs (2), corresponding to 0.08%, while the other three groups showed the same number of unique ASVs (8), that corresponded to 0.32% of the total of bacterial sequences.

UpSet diagram illustrating the distribution of ASVs in the bacterial community found in the soil samples of the treatments (control, black plastic, white plastic and dry pruning) in July. The total size of each treatment is represented on the left barplot
Redundancy analysis (RDA) was performed to determine the relationship between physicochemical properties of the soil of the different treatments and the bacterial community (Fig. 8). The first and second axis (RDA1 and RDA2) explained 72% of the variation in the bacterial community composition in the soil analysed. While the white plastic treatment did not show a correlation with any examined parameter, the black plastic and dry pruning treatments showed correlation with subsoil temperature and N content, the soil control only showed correlation with soil water content.

Redundancy analysis (RDA) of bacterial community and physicochemical properties (arrows) of the different treatments (control, black plastic, white plastic and dry pruning)
Discussion
Vegetative growth
The environment as temperature, radiation and humidity influenced the physiological processes of the roots, such as the absorption of water and mineral nutrients (Dodd et al. 2000). In our experiments, in March, none of the mulching treatments affected significantly vegetative growth of the trees because at early spring the trees did not start its vegetative development in accordance with low temperatures and short photoperiods (Kozlowski and Pallardy 2002). In the following sampling corresponding to May, the ambient temperature increased in the Mediterranean area and favoured vegetative growth (Camarero et al. 2021), which showed significant differences according to the type of mulch used. In this time, the black and white plastic treatment showed the highest significant values of vegetative growth probably related to the higher water retention of soil covered that it would avoid evaporation (Sinmidele et al. 2015). Accordingly, a mulching with grass clippings, was reported to present lower soil moisture than plastic mulching (Kader et al. 2017 2013). In our experiment, in July, there was a general growth decrease, probably due to the stress caused by the high temperatures recorded, which negatively affect the vegetative growth of lemon trees (Pérez-Pérez et al. 2009). According to the treatments, in July, the opposite results were obtained compared to May, showing control and dry pruning higher vegetative growth values than black and white mulching. This fact was not related to the number of fruits per tree where the higher number was obtained in black and dry pruning mulching. This, revealed the complexity of the citrus growth that not only were affected by ambient temperatures, but also on soil factors as we will be discuss in this section.
Leaf humidity of the different studied treatments (Fig. 2) increased in May and July compared to March which could indicate that seasonal strategy for saving water in leaves affected by temperature and light (Ribeiro and Machado 2007). The reduction of stomata has been reported as an adaptive mechanism used by plants to reduce water loss (Karimi et al. 2015), but the number of open and closed stomata indicated a short term regulation. In this way, the changes in the different seasons were not high, but appeared with treatments in May and July. Hence, in the month of July, black and white plastic mulching had higher number of stomata than control and dry pruning, but this was not corresponding with % leaf humidity or % stomata closed. As higher leaf humidity and higher number of stomata open appeared in white plastic and dry pruning mulching, should correlate with higher water transport through leaves indicating the need of transport either water and/or nutrients. In other way, the WUEi has been associated to crop productivity in agricultural ecosystems (Ono et al. 2013). As the decreased observed from March to July in our results, this parameter could be related to the need of fruit production.
It has been reported that plant photosynthesis is regulated by several climate factors, such as temperature, solar radiation and water availability (Nemani et al. 2003). In our experiments, a considerable increase in May and a decrease in the July was observed, but in a different relationship between treatments; pointing to soil influencing parameter rather than to only photosynthetic seasonality due to temperature variation (Garonna et al. 2018). The most significant changes should have appeared in the month of May and July, where the temperature was moderate and excessively high respectively, but only slight changes occurred with no relation to growth. Furthermore, there were only small increases of stomatal conductance from March to May in control, black and white plastic. The observed changes during season bring the assumption that this parameter was not dependent on temperature or light intensity as it was previously described (Allakhverdiev et al. 2008). Also, the fact that the values were higher in March and July in dry pruning mulching, bring the possibility that changes were occurred by other non-studied mechanisms. Under these conditions, plants altered leaves internal concentration of CO2 during different seasons at similar rate than Gs and An. They showed decreases in white plastic compared to black plastic that seemed to be not related to the rest of the parameter but that could show a higher water diffusion in through membranes (Martinez et al. 2011) as it occurred in white plastic trees.
Temperature and humidity of soil
The soil temperature recorded in both the upper and lower soil layers, as it was expected, increased in July respect to May and March (Costa et al. 2023). It is a factor that depended on the climatic conditions during the season and it was also affected to the physical soil properties, it has been reported that higher amplitude of the daily temperature has been related to the sand and clay composition in the soil (2012; Sremac et al. 2021). Accordingly, as our soils had similar composition, the differences were only related to mulching treatment. Therefore, the black plastic showed the highest soil temperature as it absorbs more solar radiation, which has been reported to turn into plant growth, while the white plastic mulches reflected a high proportion of solar radiation (Andreotti et al. 2010), which decreased soil temperature as significantly lower values growth values obtained in May and July compared to black plastic.
The highest significant moisture records were obtained in the month of May in all the treatments studied due to the accumulated rainfall in that period (100 L m−2), while the lowest moisture data were recorded in the month of July due to a decrease of rainfall (2 L m−2). However, soil moisture was significantly higher in the white plastic and dry pruning treatments than in the black plastic and control treatments. Water availability for plants is closely related to the efficiency of photosynthesis (Cheng et al. 2011) as it regulated stomatal conductance, affecting both the entry of CO2 into the mesophyll and the release of H2O by leaf transpiration (Ribeiro and Machado 2007). As soil water content has been positively and directly correlated with photosynthetic rate and growth (Lamptey et al. 2020), we attempted to relate these two parameters in our experiments but we found no direct relation. Therefore, other parameters should be influencing gas such as exchange parameters. Either, on the contrary as reported, there was a lack of relation between WUEi and soil moisture (Pandey et al. 2015) and temperature (Robinson et al. 2020). It has been also reported that mulching is one of the water management practices for increasing water use efficiency in crops located in semi-arid regions (Yaghi et al. 2013). However, we only found increases in black plastic mulching compared to control because higher temperatures provide more kinetic energy to water molecules for evaporating (Zeppetello et al. 2019).
Soil and leaves mineral content
The results obtained in the mineral concentration of the lower soil layer of the different treatments studied indicated that in May, the greatest significant increase in the concentrations of macronutrients and micronutrients of all the treatments studied appeared. This could be due to the high values of both temperature and humidity (Kader et al. 2017) since it has been related to the greater decomposition of added organic matter fertilization and consequently the greater the release of nutrients available to plants (Lenka et al. 2019). However, in July, the opposite occurred, with significantly lower concentrations. This tendency could be due to the increase in temperature and decrease soil humidity. It should be noted that in terms of differentiation between treatments, there is a significant increase of the macronutrients N, Ca and Mg; and of the micronutrients B, Cu and Fe for the dry pruning treatment, followed by the white and black plastic treatments, with respect to the control treatment, in the month of July. This fact could be due to the occurred pruning residue mineralization (Blagodatskaya and Kuzyakov 2008), favouring nutrient release (Manzoni et al. 2008) as a key component of nutrient availability and plant productivity (Kaspari et al. 2008). The fact that also white and black mulching also showed higher reported mineral content than control fits with the high values of both temperature and humidity that could be related to greater decomposition of added organic matter fertilization as alluded previously (Lenka et al. 2019).
Regarding the mineral content of leaves (Table 4), first of all, the nutrient levels in plant tissues of the plant vary over time, depending on the growth stage of the plant and the part analysed (Hu et al. 2023). Citrus trees change mineral nutrient absorption during growth and development processes (Bui et al. 2020). Therefore, increasing root zone temperature, affects water and nutrient uptake by accelerating metabolic activity (Lee et al. 2005) and promoting increased root volume and absorption area (Hussain and Maqsood 2011). However, these parameters were not related to the treatments that favoured the high leaf concentration of nutrients. In fact, higher N, Ca, Fe and Mn were found in leaves of trees grown under black plastic and dry pruning mulching which were the treatments that provide higher number of fruit.
Soil biological activity
The determination of soil microbiology was done in July once lemon crop season occurred and the decrease in water availability and the high sun irradiation were mostly affecting growth and photosynthetic processes (Flexas et al. 2012). Also, soluble organic carbon (SOC), available nitrogen (TN) available phophorus (P Olsen) apparent density and pH were also determined in July for correlation. In this way, firstly, soil dehydrogenase activity was determined as an indicator of the microbial redox system (Yang et al. 2003). Dehydrogenase activity of the mulching treatments was significantly higher to the control, being the dry pruning group which obtained the higher values. This may be attributed to the input of C and N resulting from the decomposition of crop residues (as related to SOC results) added to the soil as mulch, probably due to the whole season of pruning residues in contact to the soil that it should be increased due to temperature, humidity and microbial activity (Boyero et al. 2011). In the treatment involving white and black plastic mulch, the significant increase observed compared to the control, but lower compared to the dry pruning mulching, could be attributed to the documented rise in temperature and humidity, that it would also increase soil mineralization from the natural organic matter, as previously reported (Luo et al. 2019). Accordingly, the composition of the soil microbial community has been reported to be temperature dependent (Creamer et al. 2015), and related to mineralisation of soil organic matter (Huygens et al. 2011), which could be related to the high availability reported in July of our study. Therefore, the fact that higher values of Gs and An were observed in trees grown under dry pruning could be related to this increase in mineralization (Yessoufou et al. 2023; Geng et al. 2017).
According to Lauber et al. (2008), soil microbial community composition is significantly correlated with changes in soil chemical properties. In this study, the C and N soil content play important roles in changes in the bacterial community structure. The dominant taxonomic groups identified in the soil assayed were Proteobacteria, Actinobacteria, Chloroflexi and Acidobacteria, all reported by several other studies related to agricultural ecosystems (Smit et al. 2001; Valinsky et al. 2002). Proteobacteria phylum is one of the most abundant in soil ecosystems, which members occupied the highest richness across all soil samples. The Alphaproteobacteria, comprising orders Rhizobiales, Rhodospirillales, Rhodobacterales and Sphingomonadales, play crucial roles in degradation of inorganic compounds and nitrogen fixation (Fallah et al. 2021) and stimulating plant growth, underscoring their significance from an agricultural perspective (Ceja-Navarro et al. 2010). Actinobacteria is a diverse phylum of Gram-positive bacteria which some of them participate in carbon cycling and have been linked with soil organic matter production (Trinchera et al. 2022). On the other hand, Acidobacteria is a phylum of Gram-negative bacteria which members are physiologically diverse and ubiquitous. They are particularly abundant in soil habitats and can degrade complex and recalcitrant carbon sources (Fierer et al. 2003). Less copious phyla such as the Planctomycetota phylum comprises widely distributed bacteria, with many species capable of anaerobic ammonium oxidation (Kallscheuer and Jogler 2021), which is why they play an important role in global nitrogen and carbon cycles. The occurrence of members within the Cyanobacteria phylum was unexpected, given their aquatic inclination. Nevertheless, recent research, exemplified in the study led by Mhete et al. (2020), has revealed their presence in soil crusts within arid ecosystems, where they are documented as primary producers, demonstrating the ability to fix nitrogen.
The higher species richness observed through alpha diversity analysis in the dry pruning treatment compared to control can be attributed to the augmented availability of carbon for rhizosphere microorganisms. This suggests favourable soil health conditions, potentially leading to positive effects on crop productivity (Nielsen and Winding 2002). The absence of significant values in bacterial communities involving black and white plastic mulches may be attributed to the potential creation of an anaerobic environment, reducing oxygen exchange (Liao et al. 2021).
This could be due to the addition of organic matter from the pruning residues, which might provide more diverse habitats and resources for different bacterial species where dry pruning materials may create a more conducive environment for microbial diversity compared to plastic mulches being affected by numerous various factors, where one of the most important one is organic and nutrient incorporation more than moisture or temperature. Therefore, mulching produce higher fruit production, but if we want to preserve soils quality, pruning residue mulching would be a good choice. The impact of these changes on long-term soil fertility and plant productivity would be an important avenue for further research.
Conclusions
The study explored how mulching treatments influenced growth and physiological processes of lemon trees across different seasons. Mulching enhanced fruit production by the positive effect on tree gas exchange and water and nutrient uptake while preserving soil quality. However, depending on the environmental temperature, as it has been studied from the different sampling times, it would be concluded that rising ambient temperatures favoured vegetative growth, with black and white plastic mulching. This was related to the higher temperature and moisture than in dry pruning mulching and control. According to the mineral nutrients in soil, interestingly, the dry pruning treatment showed a significant increase in mineral concentrations in July, particularly for macronutrients and micronutrients, possibly due to increased mineralization of organic malemotter from crop pruning residue inputs. The soil microbial activity, as it was pointed out by dehydrogenase activity, and soil bacterial communities and diversity, also pointed out how the dry pruning mulching showed higher values, since it could be inhibited in the other plastic mulching by the high temperature. Accordingly, the higher number of fruits was related to the dry mulching. Overall, the study highlights the intricate interplay between environmental factors, mulching treatments, and physiological responses in lemon tree growth, but revealed that the use of dry pruning could be helpful to improve both the lemon crop and soil health, that it would be converted in a sustainable way of lemon production.
Data availability
The datasets generated and/or analysed during the current study are available from the corresponding author on reasonable request.
References
Aliche EB, Oortwijn M, Theeuwen T et al (2018) Drought response in field grown potatoes and the interactions between canopy growth and yield. Agric Water Manag 206:20–30. https://doi.org/10.1016/j.agwat.2018.04.013
Allakhverdiev SI, Kreslavski VD, Klimov VV et al (2008) Heat stress: An overview of molecular responses in photosynthesis. Photosynth Res 98:541–550. https://doi.org/10.1007/s11120-008-9331-0
Andreotti C, Ravaglia D, Costa G (2010) Effects of fruit load and reflective mulch on phenolic compounds accumulation in nectarine fruit. Eur J Hortic Sci 75:53–59 (https://hdl.handle.net/10863/35952)
Bista DR, Heckathorn SA, Jayawardena DM et al (2018) Effects of drought on nutrient uptake and the levels of nutrient-uptake proteins in roots of drought-sensitive and -tolerant grasses. Plants 7:28. https://doi.org/10.3390/plants7020028
Blagodatskaya E, Kuzyakov Y (2008) Mechanisms of real and apparent priming effects and their dependence on soil microbial biomass and community structure: Critical review. Biol Fertil Soils 45:115–131. https://doi.org/10.1007/s00374-008-0334-y
Boyero L, Pearson RG, Gessner MO et al (2011) A global experiment suggests climate warming will not accelerate litter decomposition in streams but might reduce carbon sequestration. Ecol Lett 14:289–294. https://doi.org/10.1111/j.1461-0248.2010.01578.x
Bui T, Vu B, Vy NTT et al (2020) Estimation of the soil to plant transfer factor and the annual organ equivalent dose due to ingestion of food crops in Ho Chi Minh city, Vietnam. Chemosphere 259:127432. https://doi.org/10.1016/j.chemosphere.2020.127432
Callahan BJ, McMurdie PJ, Rosen MJ et al (2016) DADA2: High-resolution sample inference from Illumina amplicon data. Nat Methods 13:581–583. https://doi.org/10.1038/nmeth.3869
Camarero JJ, Rubio-Cuadrado Á, Gazol A (2021) Climate windows of intra-annual growth and post-drought recovery in Mediterranean trees. Agric For Meteorol 308:108606. https://doi.org/10.1016/j.agrformet.2021.108606
Ceja-Navarro JA, Rivera-Orduña FN, Patiño-Zúñiga L et al (2010) Phylogenetic and multivariate analyses to determine the effects of different tillage and residue management practices on soil bacterial communities. Appl Environ Microbiol 76:3685–3691. https://doi.org/10.1128/AEM.02726-09
Cheng T, Rivard B, Sánchez-Azofeifa A (2011) Spectroscopic determination of leaf water content using continuous wavelet analysis. Remote Sens Environ 115:659–670. https://doi.org/10.1016/j.rse.2010.11.001
Costa JM, Egipto R, Aguiar FC et al (2023) The role of soil temperature in mediterranean vineyards in a climate change context. Front Plant Sci 14:1145137. https://doi.org/10.3389/fpls.2023.1145137
Crawford CB, Quinn B (2017) Physiochemical properties and degradation. Microplastic Pollutants. https://doi.org/10.1016/B978-0-12-809406-8.00004-9
Creamer CA, de Menezes AB, Krull ES et al (2015) Microbial community structure mediates response of soil C decomposition to litter addition and warming. Soil Biol. Biochem. 80:175–188. https://doi.org/10.1016/j.soilbio.2014.10.008
Dangi SR (2014) Soil ecology and ecosystem services. Soil Sci Soc Am J 78:335–335. https://doi.org/10.2136/sssaj2013.0005br
Davison J (1988) Plant beneficial bacteria. Nat Biotechnol 6:282–286 https://doi.org/10.1038/nbt0388-282
Dodd IC, He J, Turnbull CG et al (2000) The influence of supra-optimal root-zone temperatures on growth and stomatal conductance in Capsicum annuum L. J Exp Bot 51:239–248. https://doi.org/10.1093/jexbot/51.343.239
Elvira-Rendueles B, Moreno JM, Costa I et al (2019) Pollen calendars of Cartagena, Lorca, and Murcia (Region of Murcia), southeastern Iberian Peninsula: 2010–2017. Aerobiologia 35:477–496. https://doi.org/10.1007/s10453-019-09578-y
Fallah N, Yang Z, Tayyab M et al (2021) Depth-dependent influence of biochar application on the abundance and community structure of diazotrophic under sugarcane growth. PLoS ONE 16:1–19. https://doi.org/10.1371/journal.pone.0253970
Fierer N, Schimel JP, Holden PA (2003) Variations in microbial community composition through two soil depth profiles. Soil Biol Biochem 35:167–176. https://doi.org/10.1016/S0038-0717(02)00251-1
Flexas J, Barbour MM, Brendel O (2012) Mesophyll diffusion conductance to CO2: An unappreciated central player in photosynthesis. Plant Sci. 193–194:70–84. https://doi.org/10.1016/j.plantsci.2012.05.009
Garonna I, De Jong R, Stöckli R et al (2018) Shifting relative importance of climatic constraints on land surface phenology. Environ Res Lett 13:024025. https://doi.org/10.1088/1748-9326/aaa17b
Geng YJ, Chen L, Yang C et al (2017) Dry-season deficit irrigation increases agricultural water use efficiency at the expense of yield and agronomic nutrient use efficiency of Sacha Inchi plants in a tropical humid monsoon area. Ind Crops Prod 109:570–578. https://doi.org/10.1016/j.indcrop.2017.09.022
Georgiou A, Gregoriou C (1999) Growth, yield and fruit quality of “Shamouti” orange on fourteen rootstocks in Cyprus. Sci Hortic (amsterdam) 80:113–121. https://doi.org/10.1016/S0304-4238(98)00232-5
Hayat R, Ali S, Amara U et al (2010) Soil beneficial bacteria and their role in plant growth promotion: A review. Ann Microbiol 60:579–598. https://doi.org/10.1007/s13213-010-0117-1
Hu Y, Zhao M, Liu Z et al (2023) Seasonal changes in gas exchange, water and macro-nutrient content differ between Citrus cultivars. Environ Exp Bot 213:105432. https://doi.org/10.1016/j.envexpbot.2023.105432
Huygens D, Schouppe J, Roobroeck D et al (2011) Drying-rewetting effects on N cycling in grassland soils of varying microbial community composition and management intensity in south central Chile. Appl. Soil Ecol. 48:270–279. https://doi.org/10.1016/j.apsoil.2011.04.012
Hussain S, Maqsood MA (2011) Root zone temperature influences nutrient accumulation and use in maize. Pakistan J Bot 43:1551–1556
Iqbal R, Raza MAS, Valipour M et al (2020) Potential agricultural and environmental benefits of mulches—a review. Bull Natl Res Cent 44:1–16. https://doi.org/10.1186/s42269-020-00290-3
Jinn TL (2017) Using silicon polymer impression technique and scanning electron microscopy to measure stomatal apertures. Bio-Protocol, vol 7. https://doi.org/10.21769/bioprotoc.2449
Kader MA, Senge M, Mojid MA (2017) Recent advances in mulching materials and methods for modifying soil environment. Soil Tillage Res 168:155–166. https://doi.org/10.1016/j.still.2017.01.001
Kallscheuer N, Jogler C (2021) The bacterial phylum Planctomycetes as novel source for bioactive small molecules. Biotechnol Adv 53:107818. https://doi.org/10.1016/j.biotechadv.2021.107818
Karimi S, Yadollahi A, Arzani K et al (2015) Gas-exchange response of almond genotypes to water stress. Photosynthetica 53:29–34. https://doi.org/10.1007/s11099-015-0070-0
Kaspari M, Garcia MN, Harms KE et al (2008) Multiple nutrients limit litterfall and decomposition in a tropical forest. Ecol Lett 11:35–43. https://doi.org/10.1111/j.1461-0248.2007.01124.x
Klindworth A, Pruesse E, Schweer T, Peplies J, Quast C, Horn M, Glöckner FO (2013) Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res 41(1):e1–e1. https://doi.org/10.1093/nar/gks808
Kozlowski TT, Pallardy SG (2002) Acclimation and adaptive responses of woody plants to environmental stresses. Bot Rev 68:270–334. https://doi.org/10.1663/0006-8101(2002)068[0270:AAAROW]2.0.CO;2
Lamont WJ (2005) Plastics: Modifying the microclimate for the production of vegetable crops. Horttechnology 15:477–481. https://doi.org/10.21273/horttech.15.3.0477
Lamptey S, Li L, Xie J et al (2020) Tillage system affects soil water and photosynthesis of plastic-mulched maize on the semiarid Loess Plateau of China. Soil Tillage Res 196:104479. https://doi.org/10.1016/j.still.2019.104479
Lauber CL, Strickland MS, Bradford MA et al (2008) The influence of soil properties on the structure of bacterial and fungal communities across land-use types. Soil Biol Biochem 40:2407–2415. https://doi.org/10.1016/j.soilbio.2008.05.021
Lee SH, Ahn SJ, Im YJ et al (2005) Differential impact of low temperature on fatty acid unsaturation and lipoxygenase activity in figleaf gourd and cucumber roots. Biochem Biophys Res Commun 330:1194–1198. https://doi.org/10.1016/j.bbrc.2005.03.098
Lemanceau P, Maron PA, Mazurier S et al (2015) Understanding and managing soil biodiversity: a major challenge in agroecology. Agron Sustain Dev 35:67–81. https://doi.org/10.1007/s13593-014-0247-0
Lenka S, Trivedi P, Singh B et al (2019) Effect of crop residue addition on soil organic carbon priming as influenced by temperature and soil properties. Geoderma 347:70–79. https://doi.org/10.1016/j.geoderma.2019.03.039
Liao Y, Cao HX, Xue WK, Liu X (2021) Effects of the combination of mulching and deficit irrigation on the soil water and heat, growth and productivity of apples. Agric Water Manag 243:106482. https://doi.org/10.1016/j.agwat.2020.106482
Lozupone C, Knight R (2005) UniFrac: A new phylogenetic method for comparing microbial communities. Appl Environ Microbiol 71:8228–8235. https://doi.org/10.1128/AEM.71.12.8228-8235.2005
Luo S, Wang S, Yao P et al (2019) Soil microbial communities under film mulching and N fertilization in semiarid farmland. Nutr Cycl Agroecosyst 114:157–170. https://doi.org/10.1007/s10705-019-09998-9
Manzoni S, Jackson RB, Trofymow JA, Porporato A (2008) The global stoichiometry of litter nitrogen mineralization. Science 321:684–686. https://doi.org/10.1126/science.1159792
Martínez-Ballesta MC, Rodríguez-Hernández MC, Alcaraz-López C (2011) Plant hydraulic conductivity: In: Lakshmanan Elango (ed) hydraulic conductivity - issues, determination and applications, In: Tech, pp 103–121. https://doi.org/10.5772/18580
McMurdie PJ, Holmes S (2013) Phyloseq: An R Package for reproducible interactive analysis and graphics of microbiome census data. PLoS One 8:e61217. https://doi.org/10.1371/journal.pone.0061217
Mhete M, Eze PN, Rahube TO et al (2020) Soil properties influence bacterial abundance and diversity under different land-use regimes in semi-arid environments. Sci Afr 7:e00246. https://doi.org/10.1016/j.sciaf.2019.e00246
Murali A, Bhargava A, Wright ES (2018) IDTAXA: A novel approach for accurate taxonomic classification of microbiome sequences. Microbiome 6:1–15. https://doi.org/10.1186/s40168-018-0521-5
Nemani RR, Keeling CD, Hashimoto H et al (2003) Climate-driven increases in global terrestrial net primary production from 1982 to 1999. Science 300:1560–1563. https://doi.org/10.1126/science.1082750
Nicholson SE, Funk C, Fink AH (2018) Rainfall over the African continent from the 19th through the 21st century. Glob Planet Change 165:114–127. https://doi.org/10.1016/j.gloplacha.2017.12.014
Nielsen MN, Winding A (2002) Microorganisms as indicators of soil health. Natl Environ Res Inst 388:82
Olmos R, Garcia P, Carvajal M et al (2024) Exploring membrane vesicles in citrus fruits: a comparative analysis of conventional and organic farming approaches. J Sci Food Agric 104:235–248. https://doi.org/10.1002/jsfa.12903
Ono K, Maruyama A, Kuwagata T et al (2013) Canopy-scale relationships between stomatal conductance and photosynthesis in irrigated rice. Glob Chang Biol 19:2209–2220. https://doi.org/10.1111/gcb.12188
Pandey S, Singh J, Maurya IB (2015) Effect of black polythene mulch on growth and yield of winter dawn strawberry (Fragaria × ananassa) by improving root zone temperature. Indian J Agric Sci 85:1219–1222. https://doi.org/10.56093/ijas.v85i9.51634
Pérez-Pérez JG, Robles JM, Tovar JC et al (2009) Response to drought and salt stress of lemon “Fino 49” under field conditions: Water relations, osmotic adjustment and gas exchange. Sci Hortic (amsterdam) 122:83–90. https://doi.org/10.1016/j.scienta.2009.04.009
Prăvălie R (2021) Exploring the multiple land degradation pathways across the planet. Earth-Sci Rev 220:103689. https://doi.org/10.1016/j.earscirev.2021.103689
Rashid A, Ryan J (2004) Micronutrient constraints to crop production in soils with Mediterranean-type characteristics: a review. J Plant Nutr 27(6):959–975
Ribeiro RV, Machado EC (2007) Some aspects of citrus ecophysiology in subtropical climates: Re-visiting photosynthesis under natural conditions. Braz J Plant Physiol 19:393–411. https://doi.org/10.1590/S1677-04202007000400009
Richardson A (2001) Prospects for using soil microorganisms to improve the acquisition of phosphorus by plants. Funct. vol 28Plant Biol, pp 897–906 https://doi.org/10.1071/PP01093
Robinson JM, Barker SLL, Arcus VL et al (2020) Contrasting temperature responses of soil respiration derived from soil organic matter and added plant litter. Biogeochemistry 150:45–59. https://doi.org/10.1007/s10533-020-00686-3
Sinmidele KO, Babatunde IJ, Omolola OE (2015) Response of tomato variety (Roma F) yield to different mulch materials and staking in Kabba, Kogi State, Nigeria. J Agric Stud 3:61. https://doi.org/10.5296/jas.v3i2.7568
Smilanick JL, Erasmus A, Palou L (2019) Citrus fruits. Postharvest pathol fresh hortic prod 3–53. https://doi.org/10.12987/9780300242546-011
Smit E, Leeflang P, Gommans S et al (2001) Diversity and seasonal fluctuations of the dominant members of the bacterial soil community in a wheat field as determined by cultivation and molecular methods. Appl Environ Microbiol 67:2284–2291. https://doi.org/10.1128/AEM.67.5.2284-2291.2001
Sremac AF, Lalic B, Cuxart J et al (2021) Maximum, minimum, and daily air temperature range in orchards: What do observations reveal? Atmosphere (basel) 12:1–22. https://doi.org/10.3390/atmos12101279
Strik B, Righetti T, Rempel H (2006) Black plastic mulch improved the uptake of 15nitrogen from inorganic fertilizer and organic prunings in summer-bearing red raspberry. HortScience 41:272–274. https://doi.org/10.21273/hortsci.41.1.272
Torelló-Sentelles H, Franzke CLE (2022) Drought impact links to meteorological drought indicators and predictability in Spain. Hydrol Earth Syst Sci 26:1821–1844. https://doi.org/10.5194/hess-26-1821-2022
Trinchera A, Migliore M, Warren Raffa D et al (2022) Can multi-cropping affect soil microbial stoichiometry and functional diversity, decreasing potential soil-borne pathogens? A study on European organic vegetable cropping systems. Front Plant Sci 13:952910. https://doi.org/10.3389/fpls.2022.952910
Tsung-Luo J (2017) Using silicon polymer impression technique and scanning electron microscopy to measure stomatal apertures. Bio-Protocol 7:e2449–e2449. https://doi.org/10.21769/bioprotoc.2449
Valinsky L, Vedova G, Scupham AJ et al (2002) Analysis of bacterial community composition by oligonucleotide fingerprinting of rRNA genes. Appl Environ Microbiol 68:3243–3250. https://doi.org/10.1128/AEM.68.7.3243-3250.2002
Von Mersi W, Schinner F (1991) An improved and accurate method for determining the dehydrogenase activity of soils with iodonitrotetrazolium chloride. Biol Fertil Soils 11:216–220. https://doi.org/10.1007/BF00335770
Wright ES (2016) Using DECIPHER v2.0 to analyze big biological sequence data in R. R J 8:352–359. https://doi.org/10.32614/rj-2016-025
Yaghi T, Arslan A, Naoum F (2013) Cucumber (Cucumis sativus, L.) water use efficiency (WUE) under plastic mulch and drip irrigation. Agric Water Manag 128:149–157. https://doi.org/10.1016/j.agwat.2013.06.002
Yang YJ, Dungan RS, Ibekwe AM et al (2003) Effect of organic mulches on soil bacterial communities one year after application. Biol Fertil Soils 38:273–281. https://doi.org/10.1007/s00374-003-0639-9
Yessoufou MWIA, Tovihoudji PG, Zakari S et al (2023) Hill-placement of manure and fertilizer for improving maize nutrient- and water-use efficiencies in the northern Benin. Heliyon 9. https://doi.org/10.1016/j.heliyon.2023.e17823
Zeppetello LR, Battisti DS, Baker MB (2019) The origin of soil moisture evaporation “regimes.” J Clim 32:6939–6960. https://doi.org/10.1175/JCLI-D-19-0209.1
Zhang Y, Huang C, Zhao L et al (2023) Effects of different colors of film mulch on soil temperature and rice growth in a non-flooded condition. J Sci Food Agric 103:6352–6361. https://doi.org/10.1002/jsfa.12709
Acknowledgements
The authors thank Dr Lucía Yepes and Dr. Gloria Barzana for their help with statistics.
Funding
Open Access funding provided thanks to the CRUE-CSIC agreement with Springer Nature. This research was funded by Spanish Ministerio de Ciencia e Innovación and CDTI (MIP-243413).
Author information
Authors and Affiliations
Contributions
All authors contributed to the study conception and design. Material preparation, data collection and analysis were performed by Rafael Olmos-Ruiz, María Hurtado-Navarro, José Antonio Pascual, and Micaela Carvajal. The first draft of the manuscript was written by Rafael Olmos-Ruiz and all authors commented on previous versions of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Additional information
Responsible Editor: Juan Barcelo.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Olmos-Ruiz, R., Hurtado-Navarro, M., Pascual, J.A. et al. Mulching techniques impact on soil chemical and biological characteristics affecting physiology of lemon trees. Plant Soil (2024). https://doi.org/10.1007/s11104-024-06894-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11104-024-06894-2