
Abstract
Introduction
Urea is the most commonly used nitrogen (N) fertiliser on Australian dairy pastures, but has low N use efficiency due to high loss potential. Urease and nitrification inhibitors can be used to reduce these losses, however, their efficacy is highly variable and the reported impacts on productivity are inconsistent, with assessments often made using a single inhibitor across all seasons.
Research question
We examined the effectiveness of two commonly used inhibitors, the urease inhibitor N-(n-butyl) thiophosphoric triamide (NBTPT) and the nitrification inhibitor 3,4-Dimethylpyrazole phosphate (DMPP) applied in a strategic program, on pasture responses to urea (U) fertiliser, and fertiliser N use efficiency in a temperate dairy pasture in Southern Australia.
Materials and Methods
Pasture productivity, N uptake, agronomic N use efficiency (NUAE), 15N fertiliser recovery and soil mineral N were monitored. The nitrogen was applied in the urea form at rates of 160, 320, 480 and 640 kg N ha−1 yr−1
Results and Conclusions
The majority of the plant N (64%-85%), assessed over autumn and spring growth periods using 15N, was derived from mineralised soil organic matter (SOM). Neither of the inhibitors significantly altered pasture production or NUAE, most likely because of the major role of SOM in supplying N to plants, and because much of the applied 15N (up to 55%) was retained in the soil (0-40 cm depth) due to microbial immobilisation. Uptake of fertiliser N was highest, up to 29% of applied N, immediately following fertiliser application and then was substantially reduced over subsequent months, reflecting low levels of mineralisation of the previously immobilised N. The total loss of applied 15N was greater following autumn applications of N than spring applications, being 14% - 42% and 2% - 7% respectively and was attributed to volatilisation occurring primarily in autumn, with minimal leaching and denitrification predicted under the experimental conditions.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Intensification of agriculture production systems to boost food production is a consequence of the increased food demand of the growing world population, predicted to exceed 9 billion people by 2050 (Alexandratos and Bruinsma 2012). Increased nitrogen (N) use is one of the major changes associated with this intensification (Smith et al. 2008; Steffen et al. 2015). However globally, losses of N from fertilisers are high, considered to be around 50% on average (Lassaletta et al. 2014; Sprague et al. 2021), and can be much higher (e.g. 80%) in systems with high water inputs (Peoples et al. 1995). These large losses have implications for profitability and the environment, including air pollution, greenhouse gas emissions, depletion of the stratospheric ozone layer, global soil acidification and eutrophication (Coskun et al. 2017). Quantifying N cycling and estimating N loss from fertilisers can be difficult and is complicated by immobilisation and mineralisation processes. Nitrogen fertiliser efficiency metrics provide a measure of N loss and plant utilisation (Antille and Moody, 2021). Agronomic nitrogen-use efficiency (NUAE) can be used to describe N fertiliser efficiency in agricultural systems, and is defined as the ratio of N output (in product) to N input (e.g. fertiliser) (Antille and Moody, 2021, Oenema 2015). Nitrogen fertiliser addition is known to stimulate immobilisation and mineralisation, and plant uptake of non-fertiliser N (the ‘Added nitrogen interaction’, ANI) (Jenkinson et al. 1985), so considering only the recovery of N in the plant (N output) can underestimate the longer-term benefits to productivity from applied fertiliser N. Fertiliser N retained in the soil (immobilised) becomes a store of N for subsequent use, even though the rate of N release from this pool may be slow (Kumar and Goh 2000; Angus et al. 2006). Nutrient efficiency metrics from a fertilisation event should therefore consider cumulative N output over a longer period than just the immediate plant growth cycle. This can be particularly important for short-growth cycle crops such as dairy pastures.
Numerous management approaches can be used to mitigate N fertiliser loss. Coating nitrogen fertilisers with urease or nitrification inhibitors, known as enhanced efficiency fertilisers (EEFs) is one approach (Chen et al. 2008). The most commonly used urease inhibitor is N-(n-butyl) thiophosphoric triamide (NBTPT). Several commercial nitrification inhibitors exist, for example: 3,4-Dimethylpyrazolephosphate (DMPP), dicyandiamide (DCD) and nitrapyrin (N-serve), which are used to varying degrees in different regions of the world. NBTPT reduces the risk of NH3 loss, rapidly transforming into its oxygen analogue, N-(n-butyl) phosphoric triamide, which forms a ligand with the urease enzyme slowing urea hydrolysis for 1-2 weeks (Manunza et al. 1999). Nitrification inhibitors, such as DMPP, suppress the ammonia monooxygenase (AMO) enzyme, and hence the activity of ammonia oxidizing bacteria (AOB) and slow the first step of nitrification, the oxidization of NH4+ to NO2−, reducing the risk of NO3− leaching loss and gaseous denitrification losses (Zhou et al. 2020) as plants have more time to take up the N.
NBTPT can effectively reduce NH3 loss from applied fertilisers (Suter et al. 2013a, 2013b; Drury et al. 2017) and increase crop yield and N plant uptake (Zaman et al. 2014; Suter et al. 2021a, 2021b). The meta-analysis of Abalos et al. (2014) concluded that urease inhibitors could be used to increase crop yields and nitrogen use efficiency (NUE). The nitrification inhibitor DMPP has been reported to reduce NO3− leaching (Pawlick et al. 2019) and N2O emissions (Friedl et al. 2017; Wu et al. 2017; Suter et al. 2020), and increase crop yield and N plant uptake (Rowlings et al. 2016; Thapa et al. 2016). However, contrasting findings are often reported with inconsistent impacts of urease and nitrification inhibitors on yields, N2O emissions and nitrate leaching (Abalos et al. 2014; Dougherty et al. 2016; Koci and Nelson 2016; Silva et al. 2017). This variability is related to the fact that the efficiency of these inhibitors is influenced by soil and environmental factors (Menéndez et al. 2009; Yang et al. 2016) and also, because the experimental design can prevent any benefits being observed. For example, in pasture-based studies, often the same inhibitors (e.g. urease or nitrification inhibitors) are applied continuously over the growing season, whereas we hypothesise that a benefit will only be observed when the appropriate inhibitor is used for the specific losses that are occurring at any time (i.e. use of urease inhibitors when a high risk of ammonia volatilisation exists, use of the nitrification inhibitors when a high risk of leaching and denitrification loss exists). One study on maize has identified that yield benefits can occur if the inhibitors are used strategically (Martins et al. 2017). Inhibitors are often applied at the same, generally regional average, N rate as the conventional fertiliser treatments, and benefits to yield will most likely only occur if lower rates of N are used (Rose et al. 2018). Information is lacking on how strategic use of urease and nitrification inhibitors, at lowered N rates at different times across the growing season will affect the efficacy of urea-based N fertiliser use in rainfed pastures of south-western Victoria, Australia.
The hypotheses of this study were; (i) poor NUAE in pastures does not equate to a large loss of N, and a large amount of applied N will be retained in the soil and slowly released over time, (ii) using strategic application of NBTPT and DMPP across the growing season will optimise the benefit of the inhibitors, and (iii) combining inhibitor use with a reduced rate of N will increase the likelihood of benefits in pasture productivity, N uptake and NUAE.
Materials and methods
Site description, climate and soil properties
A field experiment was conducted on the rain-fed perennial ryegrass (Lolium perenne L. ssp. Multiflorum) dominant pasture at a commercial farm located in a high rainfall zone (HRZ) (743 mm annual rainfall) region at Allansford in south-west Victoria, Australia (38o24’39” S,142o38’28″ E). The experimental site had a history of long-term (>20 years) perennial ryegrass dominant pasture production. The soil is a Melanic-Mottled, Subnatric, Brown Sodosol (Agriculture Victoria 2020) (Solonetz (WRB 2015)) with a loam sand topsoil and a sandy clay at 40 cm depth. The soil was acidic (1:5 (v/v) CaCl2 extract) (Rayment and Higginson 1992) and had high total carbon (C) (4.6%, 0-10 cm) and total N (0.36%, 0-10 cm) (automated dry combustion using Leco FP2000 (MI, USA)) (Table 1).
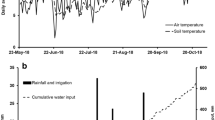
The typical annual rainfall pattern at the site is winter dominant, with 70% falling from April (autumn) to October (spring) (Bureau of Meteorology 2021) (Station 90,060). The pasture production growing season under rain-fed (dryland) conditions occurs from April to early November, being limited by low soil moisture outside these times. The mean annual monthly minimum and maximum temperatures are 8.6 °C and 19.0 °C respectively (Bureau of Meteorology 2021) (Station 90,186). On-site climate was measured as follows: solar radiation, air temperature and relative humidity were measured at a height of 3 m using a 240-100 Fristschen type net radiometer (NovaLynx, USA) (radiation) and a humidity/temperature probe (RotronicHycroClip HC2-xx, Rotronic AG, Switzerland), rainfall was measured with an CS700-L automated tipping bucket with a resolution 0.254 mm (Hydrological Services, Australia).
Experimental design and management
The field experiment commenced on 26th October 2016, following a 2-month period of exclusion of grazing animals, and finished on 22nd November 2017. The experiment was a small plot trial (3 m × 3 m per plot), with nine treatments replicated five times in a randomised block design. Nitrogen (N) response and agronomic indicators were studied from application of no nitrogen (C), granular urea (U; 46% N), and an EEF program comprised of granular urea with either the urease inhibitor N-(n-butyl) thiophosphoric triamide (NBTPT) (applied with urea as the product Green UreaNV® containing 0.105% NBPT per unit of urea-N) (GU) or the nitrification inhibitor 3,4-Dimethylpyrazole phosphate (DMPP) (applied with urea as the product Urea with ENTEC®, containing 15% DMP active per tonne of urea) (EU). Urea was applied at rates of 20, 40 (regional standard practice), 60 and 80 kg N ha−1 (U20, U40, U60, U80) to encompass the expected full N response curve, and the inhibitor products were applied at 20 and 40 kg N ha−1 only (EEF20, EEF40) to determine the benefits of reduced N rates combined with inhibitors on agronomic responses. The choice of inhibitor varied between fertiliser application events depending on the expected loss pathways (Table 2). Fertiliser was broadcast onto the plots manually after each pasture harvest.
Phosphorus (P) and potassium (K) fertilisers were applied as required at rates representative the regional farming practice, from 22 kg ha−1 to 35 kg ha−1 annually for P and from 50 kg ha−1 to 150 kg ha−1annually for K.
Harvest of the pasture occurred at 3-leaf stage. Biomass was collected from an area of 3.0 m2 within each plot (two strips of 0.5 m width and 3 m length located centrally) using a manual mower set to 6 cm harvest height to simulate grazing. The fresh weight of the harvested pasture was recorded and pasture dry matter production (DM) was determined from a subsample of approximately 0.2 kg of fresh pasture which was oven-dried to constant weight at 70 °C. Remaining pasture on the plots was harvested and discarded so there were no pasture returns to the plots. Yield (t ha−1) was calculated on a dry weight basis for each harvest and summed across the seasons. Pasture N removal (kg N ha−1) was calculated by multiplying dry matter production (DM, t ha−1) by the pasture N content (%). Nitrogen content in soil and pasture was analysed by automated dry combustion using Leco FP2000 (MI, USA). Agronomic N use efficiency (NUAE) of the fertiliser for each harvest was calculated according to Suter et al. (2013a, 2013b) as follows;
where DM is the pasture dry matter production (t ha−1), Nx is the rate of N applied from 20 to 80 kg N ha−1, and N0 is no N (control). Values of average NUAE were determined for each harvest, season and the full growing season.
Soil nitrogen
Soil samples (0-10 cm) were collected on a monthly basis initially immediately after harvest and before fertilisation (from 21st of November 2016 to 6th of June 2017) and then two weeks after each fertilisation (from 16th of August to 6th of December 2017) with the change in collection timing resulting from assessment of the initial data collected. Samples were collected using a corer (2.5 cm internal diameter), with a composite made from 5 cores per plot. Samples were analysed for gravimetric soil moisture (θg), and soil mineral nitrogen (ammonium (NH4+-N) and nitrate (NO3−-N)). Soil samples for mineral N analysis were dried (40 °C) immediately after they were collected in the field. Soil mineral N was extracted from dried and sieved (< 2 mm) soil using 2 M KCL (1:10 ratio with 1 h of shaking, and filtration through Whatman 42) and analysed colorimetrically for NH4+- N and NO3 - N plus nitrite (NO2−-N) using a segmented flow automated analyser (Skalar SAN ++). The combined NO3−-N and NO2−-N analysis is considered here as NO3−-N due to the rapid oxidation of NO2− in soil.
Soil physical analysis
Bulk density was determined on field samples collected from representative locations (Haynes and Goh, 1978). Field capacity (FC) and permanent wilting point (PWP) were determined from intact cores using a pressure plate apparatus at −10 (FC) and − 1500 (PWP) kPa).
Fertiliser 15N recovery and source of plant N
The recovery of N from a single fertilisation event was determined over the long-term (>a single growth cycle) using two sets of PVC microplot cylinders (23.7 cm inner diameter × 25 cm depth, inserted 20 cm into the ground). Each set was established, with pasture cut to grazing level, and then fertilised once, with fertiliser application on the same day as establishment, on 4th of April 2017 (autumn, set 1) and 14th of September 2017 (spring, set 2), to measure the fate of urea applied in two different seasons. Granular 15N urea (46%N, 10.2 atom %) was applied at 20 kg N ha−1 and 40 kg N ha−1 rates to the microplots, replicated three times. Pasture was harvested from the microplots regularly following the main plot harvest regime (Table 2) and analysed for biomass (t ha−1), total N (%) and 15N enrichment (%). Both sets of microplots were removed on 1st of May 2018 (after the completion of the N response trial, with the area remaining free of grazing animals till that time), representing a total time of twelve and eight months for the autumn and spring applications respectively. The soil was then collected from the microplots in 10 cm increments to 20 cm depth, and duplicate cores (9.2 cm internal diameter) were collected at two further depths (20-30 and 30-40 cm) below each microplot. Pasture samples from each harvest were separated into above- and belowground (shoots and roots) biomass, soil was cleaned from the roots and then all plant samples were dried at 70 °C to constant weight. Soil samples were dried at 40 °C. For total N and 15N analysis, dried samples were ground (<50 μm) and analysed with isotope ratio mass spectrometry (IRMS) (Hydra 20-20, SerCon). Recovery of 15N in the plant and soils was calculated according to Malhi et al. (2004) as follows;
Where %N plant is the percentage of total N in the pasture (%), DM is the dry matter production (kg ha−1), atom % 15N excess of total N in the plant = (atom % 15N of total N in the plants of the fertilised plots) – (atom % 15N of total N in the plants of the control plots), Rate of N is the rate of applied N (kg N ha−1), and atom % 15N excess of total N in fertiliser = (atom % 15N of fertiliser) – (atom % 15N natural abundance);
Where %N soil is the percentage of total soil N, SW is the weight of dry soil (kg ha−1), atom % 15N excess of total N in the soil = (atom % 15N of total N in the soil of the fertilised plots) – (atom % 15N of total N in the soil of the control plots), and Rate of N and atom % 15N excess of total N in fertiliser are as described above.
The percentage of total plant N derived from fertilisers (%Ndff) was calculated as described by Malhi et al. (2004);
and the total plant N derived from fertilisers (%Ndfs) was calculated as described by Malhi et al. (2004);
Environmental measures
Soil moisture (θv, cm3cm−3) and temperature (°C) were monitored continuously at 10 cm intervals over 80 cm depth using three logging capacitance probes (EnviroPro®, Australia; MAIT industries) placed across the site, and values were averaged.
Statistical analysis
Data were analysed by ANOVA and differences between treatments were considered significant at the p < 0.05 level using Fisher’s significance test. No transformations were required to improve the residual normality, as tested using the Shapiro-Wilk test. Statistical analysis was performed using GenStat 17th edition.
Results
Weather conditions and soil moisture
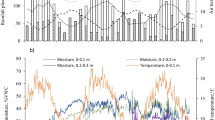
During the study, from November 2016 to May 2018, 1093 mm of rainfall fell on the site, but precipitation distribution was not uniform. The monthly precipitation was above the long-term (50 yr) (Bureau of Meteorology 2021) (Station 90,060) monthly mean from January to April 2017, and below for the remainder of the experiment except March 2018 (Fig. 1). The daily temperature fluctuations were typical of seasonal conditions in this region ranging from 0.1 °C (August 2017) to +40.4 °C (January 2018) (data not shown), and mean minimum and maximum monthly temperatures were between +5.4 °C(July) and + 25.4 °C(January), typical of the climate (Fig. 1). The average monthly soil temperature (0-10 cm) ranged from +8.0 °C (July 2017) to +21.9 °C (February 2018).

Monthly rainfall, minimum and maximum air temperature, and soil temperature (0-10 cm) during the experimental period, and long-term (50-yrs) average monthly rainfall (Bureau of Meteorology 2021, Station 90,060)
The surface (0-10 cm) soil moisture, increased steadily from March 2017 through to September 2017 due to the above average rainfall from January to April and the lowered evaporative loss from March onwards as ambient temperatures decreased (Figs. 1 and 2).

Daily soil moisture for 10, 20 and 30 cm depths, and moisture content at field capacity (FC) and permanent wilting point (PWP) for 0-10 cm soil depth during the field trial, from 1st of November 2016 to 1st of May 2018
The soil moisture content in the surface (0-10 cm) and subsurface (PWP for 10-20 = 14%, for 20-30 = 23%) was below that available to plants (<PWP) through summer to around mid-March.
From the end of March, when active pasture growth commences in this region, to late November soil water was available to plants (between FC and PWP), although the subsoil moisture was low until the end of April. Then the soil moisture rapidly increased and from August to September exceeded FC in the 0-10 cm and 20-30 cm layers due to the abundant rainfall and reduced evaporation. After rainfall dropped and temperatures increased from September 2017 onwards, the soil moisture declined over summer, with intermittent increases associated with periodic summer rain, until the autumn opening rainfalls on 12th of March 2018 (Fig. 2).
Pasture dry matter production and plant N uptake
Pasture growth only occurred between April and November, which is typical for rainfed pastures in this region, due to insufficient soil moisture (< PWP) through summer and early autumn (December-March) (Fig. 2). Pasture dry matter production steadily increased from April to August (average of 2.1 and 3.1 t ha−1 respectively across all fertilised treatments) in response to rain and increased soil moisture (r = 0.48 - 0.70) (Table 3, Fig. 2). After this, growth rates declined as soil moisture content was depleted, until the end of the growing season (November).
Use of N fertiliser increased productivity, with the control (non-fertilised) treatment producing the lowest DM production at all harvest events (Table 3). DM production was significantly (p < 0.05) increased by application of N compared to the control, in all harvests except for late November 2017 (Table 3). On this date, correlation between harvest and applied fertiliser was week (r = −0.26). On all other occasions, application of N highly correlated (r = 0.84 - 0.98) with the plant production and resulted in DM increases (from 21st of November 2016 to 1st of November 2017) of 13% (20 kg N ha−1), 46% (40 kg N ha−1), and 62% (60 kg N ha−1), with no significant (p > 0.05) increase above a rate of 60 kg N ha−1 (Table 4). The annual DM production increased relative to the non-fertilised control by 27%, 48%, 59% and 71% with the addition of 160 kg N ha−1, 320 kg N ha−1, 480 kg N ha−1 and 640 kg N ha−1 respectively as urea over the growing season.
DM production at 20 kg N ha−1 was no different with urea or the inhibitor program. Application of inhibitors at 40 kg N ha−1 (EEF40) appeared to increase DM by 5% compared to conventional urea at the same rate, although non-significantly. N fertiliser uptake by pasture increased from April to August matching the increased DM production (r = 0.64-0.91), reaching a maximum on August 1st and then gradually declining to the lowest values recorded on 22nd November (Table 3). Overall, N uptake by plants significantly increased (p < 0.05) with increasing fertiliser rate to 60 kg N ha−1 with no significant further increase to the 80 kg N ha−1 rate (Table 4).
A total of 685.9 kg N ha−1 was removed under the highest treatment (U80), and 362.9 kg N ha−1 under the non-fertilised treatment. Use of inhibitors did not alter the total N uptake relative to conventional urea at the same rate.
The NUAE ranged from1.74 kg DM kg N−1 to 37.8 kg DM kg N−1, with the greatest NUAE seen in September and October, and very low NUAE on 22nd November 2017 (Table 3). Use of inhibitors did not alter the NUAE relative to conventional urea at the same rate, except on the 1st November 2017 harvest where the NUAE for EEF20 was significantly lower than for U20 (Table 3). NUAE often, but not always, decreased with increasing rate of N applied as urea, although not significantly (Table 3). Total NUAE decreased as N application rate increased, from 17.30 kg DM kg N−1 in the U20 to 11.38 kg DM kg N−1 in the U80 treatment, although there was no significant (p > 0.05) difference between N rates or with the use of inhibitors compared to conventional urea (Table 4).
Profile and surface soil mineral N
The background profile soil mineral N (0-40 cm) was 69.7 kg N ha−1 at the beginning of growing season (April 2017) with NO3−-N being the dominant form (93%) (data not shown). The profile mineral N significantly (p < 0.05) increased to 151.9 kg N ha−1 in the U80 treatment by November 2017. There was no significant change in profile mineral N for any other treatment.
In considering the mineral N levels, we expect lower levels prior to the samples collected in June 2017 compared to those collected afterwards due to the sampling protocol described in the materials and methods. Ammonium (NH4+-N) levels fluctuated in the 0-10 cm layer from 0.58 kg N ha−1 to 54.8 kg N ha−1, with maximum values occurring in June and November 2017 (Fig. 3). With the exception of the November 2017 samples, neither N rate nor use of inhibitors affected soil NH4+-N content.

Surface (0-10 cm) soil NH4+-N (a) and NO3−-N (b) concentrations measured prior to fertiliser application (21st of November 2016 to 6th of June 2017) and then two weeks after fertiliser application (from 16th of August). Values represent means (±standard errors) of five replicates
Nitrate (NO3−) levels in the surface layer increased in April-May, possibly due to enhanced mineralisation occurring with elevated soil moisture (Fig. 2) after the summer. The concentration of NO3− then dropped and N rate affected the level of NO3− with higher levels in the 60 and 80 kg N ha−1 rates. A second increase in NO3− levels occurred just for the 80 kg N ha−1 treatment in October, which may have resulted from reduced soil moisture slowing the crop demand for N and a build-up of N from the repeated high inputs over the growing season (Fig. 3). Use of N rates up to 40 kg N ha−1 (regional practice) and use of inhibitors did not affect NO3− levels, however application of higher rates of N (60 and 80 kg N ha−1) significantly (p < 0.05) increased soil NO3−-N levels in the soil surface layer through the April to November 2017 growing season.
15N recovery in pasture and soil
Between 32.6 and 43.5% of the applied 15N was recovered in the pasture aboveground biomass (shoots) (Table 5) over the experimental period. The majority was found in the first harvest immediately after application (20.6%-23.5% (autumn) and 24.5-26.8% (spring)), with progressively less recovered in subsequent harvests (5.0%-5.3% (autumn) and 12.1%-14.1% (spring) at the second harvest and < 1.6% for each subsequent harvest) (Table 5). The total fertiliser 15N recovery over all harvests was slightly higher when less N was applied (35.1% and 43.5% with 20 kg N ha−1 compared to 32.6% and 37.8% with 40 kg N ha−1 for the autumn and spring harvests respectively). Recovery of 15N in the roots ranged from 1.2% to 3.8% across all harvests and no treatment effect was detected (Table 6).
The 15N recovered in the soil ranged from 24.5% to 54.7% (Table 6). At the lower N rate, the recovery of 15N in the soil was similar in autumn and spring (48.0-50.0%), while at the higher rate there was more N recovered in the soil in spring (54.7%) compared to autumn (24.5%).
By the end of the experiment, twelve months after the autumn application and eight months after the spring application, 86.9% (autumn) and 94.7% (spring) of the applied 15N was recovered in the plant-soil system with the lower urea rate, and 58.7% (autumn) and 96.3% (spring) with the higher urea rate (Table 6). As shown in Fig. 4, increasing the rate of fertiliser application significantly (p < 0.05) increased the amount of unaccounted for N in both seasons, but most noticeably in autumn, with 2.9 kg N ha−1 (14%) and 17 kg N ha−1 (43%) of N applied at 20 and 40 kg N ha−1 respectively unaccounted for, while only and 0.4 kg N ha−1 (2%) and 2.8 kg N ha−1 (7%) respectively were unaccounted for in spring.

The fate of 15N-labeled fertiliser in plant-soil system as affected by rate and season a) autumn and b) spring at the Allansford dryland site. Values represent means (±standard errors) of three replicates
The majority of the plant N taken up by over the entire experimental period was derived from the soil (Ndfs) (≥65%) (Table 6). Applying a higher rate of N increased the amount of plant N derived from the fertiliser (i.e. 34.8% (autumn) and 26.1% (spring) at 40 kg N ha−1 compared to 15.3% (autumn) and 17.0% (spring) at 20 kg N ha−1), with a significant (p < 0.05) increase in autumn.
At the first harvest, a short time after the fertiliser was applied, the amount of N in the pasture derived from the soil was still large and influenced by the rate of N applied, being nearly 90% (autumn, 20 kg N ha−1), 77% (autumn, 40 kg N ha−1), 88% (spring, 20 kg N ha−1) and 82% (spring, 40 kg N ha−1) of total pasture N (Table 6). Across all treatments and seasons, the majority of the 15N was recovered in the top 0-10 cm soil layer (up to 68% of total recovery) (Table 7).
Discussion
Pasture response to soil mineral N dynamics and urea N
The pasture response was influenced by the pattern of topsoil (0-10 cm) mineral N variation, which was a result of the combined effect of seasonal conditions, particularly rainfall and consequently soil moisture, fertiliser N rates and the plant uptake (Figs. 1, 2 and 3). The N rate increase understandably caused increase of mineral N content in soil in all occasions. During the dry period (December application on 21st of November 2016) combined with limited soil moisture (Fig. 2) led to low levels of soil NH4+ due to slow rates of mineralisation, while an accumulation of NO3− continued over this period due to limited plant growth and N uptake (Fig. 3). During the growth period (April 2017 - November 2017), two peaks in 0-10 cm NH4+ were observed, the first (May 2017) associated with rapid mineralisation of soil organic matter following the higher April rainfall, and the second (November 2017) occurring due to a rapid decline in soil moisture and consequently plant N uptake (Fig.2) and plant growth (Table 3). Similar to this, 2016 to March 2017) the lack of fertiliser N inputs (last limited DM response to fertilisation occurred when soil moisture was low (April and November) and strong responses were observed when there was a good supply of plant available water, with increasing soil and ambient temperatures improving the DM response further when sufficient water was available. The correlation between soil moisture in the top 10 cm and herbage was moderate varying between 0.48 - 0.70 under different urea rates. Similar influences of moisture and temperature on pasture growth have been reported elsewhere, under winter dominant rainfall climates (Smith and Stephens 1976; Suter et al. 2021a, 2021b). Application of fertilise in our experiment resulted in biomass response and an agronomic N fertiliser use efficiency (NUE) of 11.4-17.3 kg of DM for every kg of N applied, which corresponds to previously reported biomass responses under similar climates (Zaman et al. 2013).
We found little benefit of applying more than 60 kg N ha−1 for biomass and NUAE. The observed reduction in NUAE with increasing N application rate is in line with previous studies, reflecting the curvilinear N response curve (Cardenas et al. 2019).
The high levels of soil organic matter, carbon (C) and N at our site (TC 4.6% in the 0-10 cm depth, and a total of 159 t C ha−1 in the 0-40 cm depth; TN of 0.36% in the 0-10 cm depth, and a total of 11 t N ha−1 in the 0-40 cm) likely affected the N fertiliser efficiency by providing N via mineralisation. DM production in the non-fertilised control changed seasonally reflecting differences in mineralisation under changing moisture and temperature conditions, and growing conditions (Table 3). Immediately after the fertiliser was applied, the N derived from the soil was large (>76%) (Table 6) indicating that the soil stored N is an important component of the N budget in these systems.
Pasture response to use of inhibitors
The lack of impact of the inhibitors on DM production and NUAE observed in our study is comparable to previous reports on pasture production systems(Dougherty et al. 2016; Nauer et al. 2018), but contradicts others, and reports from two meta-analyses showing improved yield with urease or nitrification inhibitors (Zaman et al. 2008; Zaman et al. 2009; Abalos et al. 2014; Burzaco et al. 2014; Thapa et al. 2016). A previous study in a similar climate found benefits from using the inhibitors, applied all year rather than strategically, were seasonally driven, for example researches have shown that NBTPT can effectively reduce NH3 loss from temperate grasslands by 29% in Spain (Menéndez et al. 2009), by 70-75% in the UK (Chadwick et al. 2005; Watson et al. 2008), and by up to 80% in southern Australia (Suter et al. 2013a, 2013b; Lam et al. 2019), but these reduced N losses do not necessarily translate into agronomic benefits.
We hypothesise that the lack of a significant effect of the inhibitors on pasture productivity and NUAE in our study was a result of the interaction of several factors including soil organic matter content, soil pH and climate influencing both the magnitude of the N loss and the efficacy of the inhibitors. Saggar et al. (2013) found that soil organic matter was the major determinant of the efficacy of NBTPT in reducing urea hydrolysis and NH3 loss in pastures. The expected time over which the NBTPT is effective in reducing NH3 loss is a few days in high C soils compared to more than two weeks in low C soils (Suter et al. 2011). The lack of response in the top 0-10 cm of soil to N rate and fertiliser type observed in this study (Fig.3) also can be due to rapid urea hydrolysis following the fertiliser application. Rapid urea hydrolysis, often within 1-5 days, is expected in the pasture soils due to high levels of organic matter and urease (Zaman et al. 2009; Suter et al. 2011). The high organic matter content (4.6% C) at our site, and the fact that organic matter is negatively correlated with NBTPT efficacy (Bremner and Chai 1986) may explain the reduced NBTPT efficacy observed. An abundance of urease, the enzyme inhibited by NBTPT (Hongprayoon et al. 1991), can also reduce NBTPT effectiveness in C-rich dairy pasture soils. Some studies demonstrated that SOM is negatively correlated with DMPP effectiveness due to adsorption of the nitrification inhibitor on soil colloids (Volpi et al. 2017). In addition, SOM is a source of energy for heterotrophic microbes that can degrade DMPP and reduce its effectiveness (Fisk et al. 2015). This is in line with the study of Zhu et al. (2019) who showed that higher rates of DMPP were needed for soils with higher clay and organic C content.
The acidic nature of the soil at our site (pHCaCl2 6.1) likely contributed to low efficacy of both NBTPT and DMPP. The urease inhibitor degrades faster in acidic then neutral or alkaline soils, which affects the inhibitor longevity and hence efficacy (Engel et al. 2015; Soares et al. 2012). In addition, under acidic conditions we expect ammonia oxidizing archaea and heterotrophic nitrifiers will be the dominant nitrifiers, both of which are not affected by the nitrification inhibitor (Barth et al. 2001; Stempfhuber et al. 2015).
Whilst temperature is a major factor influencing both inhibitors, the soil temperatures recorded in our study indicates temperature was likely not a key driver of their performance. The soil temperatures recorded during the time of DMPP and NBTPT use were below the critical values for increased degradation rates of 20 °C for DMPP and 25 °C for NBTPT (Bremner et al. 1986; Carmona et al. 1990; Clay et al. 1990; Zerulla et al. 2001). While ambient air temperature during the growing season reached 33.5 °C on some occasions, the maximum surface soil temperature was regulated with the grass cover and did not exceed17.5 °C and20.3 °C respectively in the periods when DMPP and NBTPT were used.
Soil moisture has being identified as a key factor affecting DMPP efficacy, with DMPP found to be less effective in high water content soils due to a predominance of anaerobic conditions leading to reduced rates of nitrification (Walker et al. 2003; Chen et al. 2010; Landesman and Dighton 2010) and increased rates of immobilization (Compton and Boone 2002). Often, especially in winter, the soil moisture at our site was at the upper end and above the optimum moisture conditions for nitrification (60%-65% water filled pore space (WFPS)) (Linn and Doran 1984; Kiese et al. 2008), ranging from 65%-91% WFPS (equivalent of 32-45% volumetric water content (VWC)) (Fig. 2). Hence, the high soil water content could be another factor limiting DMPP’s ability to cause a productivity and NUE response. We do not expect that leaching loss, which can be prevented by DMPP, would be high in our soil due to the low permeability clay subsoil. Lower hydraulic conductivity was evident in the subsoil compared to the topsoil (Table 1). Thus, we hypothesize that leaching was minimal in the investigated soil.
The limited impact observed with the inhibitors in our study is further influenced by the fact that the majority of N supplied to the pasture was derived from mineralisation of organic matter rather than the fertiliser (Table 6), as discussed below.
Uptake and recovery of 15N labelled fertiliser
Of the applied 15N, 35 to 47% was recovered in the biomass over the 8-12 month period, similar to previous studies for pastures in temperate climates (29%-49%) (Haynes 1999; Williams et al. 2000; Suter et al. 2020), 25-55% was recovered in the soil, and1 to 41% was lost. Most of the fertiliser N recovered in the pasture over the 8-12 month period was rapidly taken up in the first (61-63% of the total 15N recovered), and second (14-32% of the total 15N recovered) harvests following fertiliser application (Table 5). This pattern is typically associated with high levels of microbial immobilisation, especially evident in a microbially rich pasture soils (Ledgard et al. 1988). This immobilised N can be considered as a store of N available for subsequent crops. However, the release of this stored fertiliser N was slow and decreased with time, particularly after the second harvest with only 0.01% to 3.4% recovered in subsequent harvests, similar to the range reported elsewhere (e.g. < 1%-10%) (Hart et al. 1993; Suter et al. 2020), with a large proportion (up to 55%) of applied 15N remaining in the soil after 12 months. This low recovery can be due to the fact that only a small amount of the labelled inorganic N incorporated into the soil organic pool was subsequently mineralised and the majority of mineralised N came from older soil stores (Ladd and Amato 1986; Kuzyakov et al. 2000). Our data shown that less than 35% of the plant N was derived from the fertiliser (Ndff) (Table 6) indicating that soil N supplied most of the plant’s N requirement. Supply of mineral N to plants via mineralisation is common in agricultural systems, particularly where high organic matter exists. For example, in a rotation of 2-4 years grass/clover pasture with 4-6 years of wheat, 96%-98% of the N taken up by the wheat plants originated from mineralised soil organic N that built up during the pasture phase (Williams and Haynes 1997). Typically, concurrent with immobilisation of applied N, soil organic N is readily mineralised, supplying N to plants. This process was described by Jenkinson et al. (1985) as the ‘added nitrogen interaction’ (ANI). We suggest that the main mechanism reducing recovery of applied fertiliser N in the plant in our study was immobilisation occurring rapidly after N application, coupled with increasing uptake of mineralised N induced by ANI.
The plant uptake after the first and second harvests, when most of applied 15N was recovered, tended to be higher in spring (36.6% - 40.9%) compared to autumn (25.9% - 28.5%). This is in line with other pasture studies conducted in similar climatic regions (Cookson et al. 2001). This was associated with slower plant growth due to reduced solar radiation in autumn, while other climatic parameters were similar in both seasons, the average ambient temperature at the period was +12.2 °C in autumn and + 12.1 °C in spring, volumetric water content ranged from 27% to 57% in autumn and from 23% to 56% in spring to the top 10 cm layer (Figs. 1 and 2). Interestingly, the total recovery of N in the autumn-fertilised plots (32.6%-35.1%) continued to be lower, albeit non-significantly, than in spring-fertilised plots (37.8%-43.5%).
Most of the N loss occurred after the autumn fertiliser application, increasingly evident with an increase in the N rate (Fig. 4). In agricultural systems, applied fertiliser N can be lost via several pathways, including via NH3 volatilisation which can be as high as 30% in temperate pastures under similar climate conditions (warm weather with some humidity) as experienced at our site in autumn 2017 (Saggar et al. 2013; Suter et al. 2013a, 2013b). Applied N can also be lost via leaching or denitrification after NH4+ is nitrified to NO3−, but we hypothesise that these were minor contributors to N loss in our system. Typically, denitrification losses are expected to be minimal due to the well-developed root system of long-term (>20 year) ryegrass plants competing for the N substrate (NO3−) with microbiota (Hoyle et al. 2011). Nitrate leaching losses are thought to be minimal because of the clay subsoil restricting the hydraulic conductivity (to 9 mm hr.−1) and hence water movement below 50 cm depth (Table 1). Although some lateral flow was possible, but we do not expect it to be significant because of the predominantly flat landscape.
The limited importance of the fertiliser N for plant nutrition immediately following fertilisation helps to explain the lack of impact of the inhibitors on pasture productivity. In autumn, where recovery of N in the plant was low, we conclude this is likely due to high loss of NH3, the immediate impact from a urease inhibitor, which has been shown to significantly reduce NH3 loss in similar systems (Suter et al. 2013a, 2013b), is limited because the soil N store supplies the N for plant growth. In rainfed pasture systems where losses from leaching and denitrification may be low, the use of nitrification inhibitors has been shown to lead to greater retention of N in the soil organic matter pool (Suter et al. 2020) which will increase the efficiency of the system in the longer term but may not be apparent in the short term. Regular fertiliser application is therefore required to build the soil organic N pool to maintain a good supply of N for the subsequent pasture, and the best time to build this store is when the risk of loss is lowest. Future research on the optimal variable fertiliser strategy (timing) to minimise N loss and build the soil N reserves is warranted, as well as a long-term assessment of the inhibitor impacts, to make recommendations for long-term sustainable management of N in rainfed perennial dairy pastures.
Conclusion
Pasture dry matter production variation in the rainfed perennial ryegrass dominant pasture was found to be due to a combination of environmental variables and urea N rates. Soil moisture, influenced by rainfall distribution over the year, was a major factor affecting productivity and NUAE. Soil organic matter was also a key factor affecting fertiliser response and productivity. Applying conventional urea above the rate of 60 kg ha−1 did not lead to increased productivity, but did decrease NUAE, indicating this was a critical limit of N efficiency. The urease inhibitor NBTPT and the nitrification inhibitor DMPP did not have any consistent effect on yield or NUAE, likely due to the large reserves of mineralizable soil N, and the large contribution of soil N to pasture nutrition. Further, limited leaching and denitrification loss likely limited the impact of the nitrification inhibitor. For sustainable management of dairy pasture soils under dryland conditions in temperate environments, regular N fertiliser use is required to build the soil organic matter pool and facilitate continued pasture nutrition, and the optimal application timings should be considered based on the expected seasonal N losses. Under this system where N inputs have been historically high, and N losses are predicted to be minimal, the strategic use of inhibitors did not lead to short-term productivity and NUE benefits. Benefits from using inhibitors should be considered alongside longer-term N management strategies designed to lower the background N reserves.
Data Availability
The data that support the findings of this study are available on request from the corresponding author, H. Suter.
Abbreviations
- DMPP3:
-
4-Dimethylpyrazole phosphate
- N:
-
nitrogen
- NBTPT:
-
(N-(n-butyl) thiophosphoric triamide
- NUAE:
-
agronomic N use efficiency
- U:
-
urea
References
Abalos D, Jeffery S, Sanz-Cobena A, Guardia G, Vallejo A (2014) Meta-analysis of the effect of urease and nitrification inhibitors on crop productivity and nitrogen use efficiency. Agric Ecosyst Environ 189:136–144. https://doi.org/10.1016/j.agee.2014.03.036
Agriculture Victoria (2020) SW3. In: Victorian resources online, State Government of Victoria vro.agriculture.vic.gov.au/dpi/vro/vrosite.nsf/pages/vrohome/ accessed 19 march 2018)
Alexandratos N, Bruinsma J (2012) World agriculture towards 2030/2050: the 2012 revision. FAO, ESA working paper no. 12-03, Rome
Angus JF, Bolger TP, Kirkegaard JA, Peoples MB (2006) Nitrogen mineralisation in relation to previous crops and pastures. Aust J Soil Res 44:355–365. https://doi.org/10.1071/SR05138
Antille DL, Moody PW (2021) Nitrogen use efficiency indicators for the Australian cotton, grains, sugar, dairy and horticulture industries. J Environ Sustain 10:100099. https://doi.org/10.1016/j.indic.2020.100099
Barth G, von Tucher S, Schmidhalter U (2001) Influence of soil parameters on the effect of 3,4-dimethylpyrazole-phosphate as a nitrification inhibitor. Biol Fertil Soils 34:98–102. https://doi.org/10.1007/s003740100382
Bremner JM, Chai HS (1986) Evaluation of N-butyl phosphorothioic triamide for retardation of urea hydrolysis in soil. Commun Soil Sci Plant Anal 17:337–351
Bremner JM, McCarty GW, YeomansJC CHS (1986) Effects of phosphoroamides on nitrification, denitrification, and mineralization or organic nitrogen in soil. Commun Soil Sci Plant Anal 17:369–384
Bureau of Meteorology (2021) Climate Data Online. http://www.bom.gov.au/climate/ Accessed 10 March 2020
Burzaco JP, Ciampitti IA, Vyn TJ (2014) Nitrapyrin impacts on maize yield and nitrogen use efficiency with spring-applied nitrogen: field studies vs. meta-analysis comparison. J Agron 106:753–760. https://doi.org/10.2134/agronj2013.0043
Cardenas LM, Bhogal A, Chadwick DR, McGeough K, Misselbrook T, Rees RM, Thorman RE, Watson CJ, Williams JR, Smith KA, Calvet S (2019) Nitrogen use efficiency and nitrous oxide emissions from five UK fertilised grasslands. Sci Total Environ 661:696–710. https://doi.org/10.1016/j.scitotenv.2019.01.082
Carmona G, Christianson CB, Byrnes BH (1990) Temperature and low concentration effects of the urease inhibitor N-(n-butyl) thiophosphoric triamide (nBTPT) on ammonia volatilization from urea. Soil Biol Biochem 22:933–937
Chadwick D, Misselbrook T, Gilhespy S, Williams J, Bhogal A, Sagoo L, Nicholson F, Webb J, Anthony S, Chambers B, (2005) Ammonia emissions and crop N use efficiency. In (ed.) DEFRA, component report for Defra project NT2605 (CSA 6579) pp 71
Chen D, Suter HC, Islam A, Edis R (2010) Influence of nitrification inhibitors on nitrification and nitrous oxide (N2O) emission from a clay loam soil fertilized with urea. Soil Biol Biochem 42:660–664. https://doi.org/10.1016/j.soilbio.2009.12.014
Chen D, Suter HC, Islam A, Edis R, Freney JR, Walker CN (2008) Prospects of improving efficiency of fertilizer nitrogen in Australian agriculture; a review of enhanced efficiency fertilizers. Aust J Soil Res 46:289–301. https://doi.org/10.1071/SR07197
Clay DE, Malzer GL, Anderson JL (1990) Ammonia volatilization from urea as influenced by soil temperature, soil water content, and nitrification and hydrolysis inhibitors. Soil Sci Soc Am J 54:263–266
Compton JE, Boone RD (2002) Soil nitrogen transformations and the role of light fraction organic matter in forest soils. Soil Biol Biochem 34:933–943. https://doi.org/10.1016/S0038-0717(02)00025-1
Cookson WR, Rowarth JS, Cameron KC (2001) The fate of autumn-, late winter- and spring- applied nitrogen fertilizer in a perennial ryegrass (Lolium perenne L.) seed crop on a silt loam soil in Canterbury. New Zealand Agric Ecosyst Environ 84:67–77. https://doi.org/10.1016/S0167-8809(00)00196-1
Coskun D, Britto DT, Shi W, Kronzucker HJ (2017) How plant root exudates shape the nitrogen cycle. Trends Plant Sci 22:661–673. https://doi.org/10.1016/j.tplants.2017.05.004
Dougherty WJ, Collins D, Van Zwieten L, Rowlings DW (2016) Nitrification (DMPP) and urease (NBPT) inhibitors had no effect on pasture yield, nitrous oxide emissions, or nitrate leaching under irrigation in a hot-dry climate. Soil Res 54:675. https://doi.org/10.1071/SR15330
Drury CF, Yang X, Reynolds WD, Calder W, Oloya TO, Woodley AL (2017) Combining urease and nitrification inhibitors with incorporation reduces Ammonia and nitrous oxide emissions and increases corn yields. J Environ Qual 46:939–949
Engel RE, Towey BD, Gravens E (2015) Degradation of the urease inhibitor NBPT as affected by soil pH. Soil Sci Soc Am J 79:1674–1683
Fisk LM, Maccarone LD, Barton L, Murphy DV (2015) Nitrapyrin decreased nitrification of nitrogen released from soil organic matter but not amoA gene abundance at high soil temperature. Soil Biol Biochem 88:214–223. https://doi.org/10.1016/j.soilbio.2015.05.029
Friedl J, Scheer C, Rowlings DW, Mumford MT, Grace PR (2017) The nitrification inhibitor DMPP (3,4-dimethylpyrazole phosphate) reduces N2 emissions from intensively managed pastures in subtropical Australia. Soil Biol Biochem 108:55–64. https://doi.org/10.1016/j.soilbio.2017.01.016
Hart PBS, Powlson DS, Poulton PR, Johnston AE, Jenkinson DS (1993) The availability of the nitrogen in the crop residues of winter wheat to subsequent crops. J Agric Sci 121:355–362
Haynes RJ, Goh KM (1978) Ammonium and nitrate nutrition of plants. Bioll Rev 53:465–510
Haynes RJ (1999) Fate and recovery of 15N-labelled fertilizer urea applied to winter wheat in spring in the Canterbury region of. New Zealand J Agric Sci 133:125–130
Suter H, Lam SK, Walker C, Chen D (2021a) Benefits from enhanced efficiency nitrogen fertilisers in rainfed temperate pastures are seasonally driven. Soil res. Accepted 12th august 2021
Hongprayoon C, Lindau CW, Patrick WHJ, Bouldin DR, Reddy KR (1991) Urea transformations in flooded soil columns: I experimental results. Soil Sci Soc Am J 55(4):1130–1134. https://doi.org/10.2136/sssaj1991.03615995005500040038x
Hoyle F, Murphy D, Baldock J (2011) Soil organic carbon - role in rainfed farming systems with particular reference to Australian conditions. In: Tow P, Cooper I, Partridge I, Birch C (eds) Rainfield farming systems. Springer, pp 339–361
Jenkinson DS, Fox RH, Rayner JH (1985) Interactions between fertilizer nitrogen and soil nitrogen-the so-called 'priming' effect. J Soil Sci 36:425–444. https://doi.org/10.1111/j.1365-2389.1985.tb00348.x
Kiese R, Hewett B, Butterbach-Bah K (2008) Seasonal dynamic of gross nitrification and N2O emission at two tropical rainforest sites in Queensland, Australia. Plant Soil 30:105–117
Koci J, Nelson PN (2016) Tropical dairy pasture yield and nitrogen cycling: effect of urea application rate and a nitrification inhibitor, DMPP. Crop Pasture Sci 67:766–779. https://doi.org/10.1071/CP15400
Kumar K, Goh KM (2000) Crop residues and management practices: effects on soil quality, soil nitrogen dynamics, crop yield and nitrogen recovery. Adv Agron 68:198–319
Kuzyakov Y, Friedel JK, Stahr K (2000) Review of mechanisms and quantification of priming effects. Soil Biol Biochem 32:1485–1498
Ladd JN, Amato M (1986) The fate of nitrogen from legume and fertilizer sources in soils successively cropped with wheat under field conditions. Soil Biol Biochem 18:417–425. https://doi.org/10.1016/0038-0717(86)90048-9
Lam SK, Suter H, Bai M, Walker C, Mosier AR, van Grinsven H, Chen D (2019) Decreasing ammonia loss from an Australian pasture with the use of enhanced efficiency fertilizers. Agric Ecosyst Environ 283:106553. https://doi.org/10.1016/j.agee.2019.05.012
Landesman WJ, Dighton J (2010) Response of soil microbial communities and the production of plant-available nitrogen to a two-year rainfall manipulation in the New Jersey pinelands. Soil Biol Biochem 42:1751–1758. https://doi.org/10.1016/j.soilbio.2010.06.012
Lassaletta L, Billen G, Grizzetti B, Anglade J, Garnier J (2014) 50 year trends in nitrogen use efficiency of world cropping systems: the relationship between yield and nitrogen input to cropland. Environ Res Lett 9:105011. https://doi.org/10.1088/1748-9326/9/10/105011
Ledgard SF, Steele KW, Feyter C (1988) Influence of time of application on the fate of 15N-labelled urea applied to dairy pasture. New Zealand J Agric Res 31:87–91. https://doi.org/10.1080/00288233.1988.10421368
Linn DM, Doran JW (1984) Effect of water-filled pore space on carbon dioxide and nitrous oxide production in tilled and nontilled soils. Soil Sci Soc Am J48:1267–1272. https://doi.org/10.2136/sssaj1984.03615995004800060013x
Malhi SS, Johnston AM, Gill KS, Pennock DJ (2004) Landscape position effects on the recovery of 15N-labelled urea applied to wheat on two soils in Saskatchewan, Canada. Nutr Cycl Agroecosyst 68:85–93
Manunza B, Deiana S, Pintore M, Gessa C (1999) The binding mechanism of urea, hydroxamic acid and N-(N-butyl)-phosphoric triamide to the urease active site. A comparative molecular dynamics study. Soil Biol Biochem 31:789–796
Martins MR, Sant’Anna SAC, Zaman M, Santos RC, Monteiro RC, Alves BJR, Jantalia CP, Boddey RM, Urquiaga S (2017) Strategies for the use of urease and nitrification inhibitors with urea: impact on N2O and NH3 emissions, fertilizer-15N recovery and maize yield in a tropical soil. Agric Ecosyst Environ 247:54–62. https://doi.org/10.1016/j.agee.2017.06.021
Menéndez S, Merino P, Pint M, González-Murua C, Estavillo JM (2009) Effect of N-(n-butyl) Thiophosphoric triamide and 3,4 dimethylpyrazole phosphate on gaseous emissions from grasslands under different soil water contents. J Environ Qual 38:27–35. https://doi.org/10.2134/jeq2008.0034
Nauer PA, Fest BJ, Visser L, Arndt SK (2018) On-farm trial on the effectiveness of the nitrification inhibitor DMPP indicates no benefits under commercial Australian farming practices. Agric Ecosyst Environ 253:82–89. https://doi.org/10.1016/j.agee.2017.10.022
Oenema O (2015) In: Society CIF (ed) Nitrogen use efficiency (NUE) - an indicator for the utilization of nitrogen in agriculture and food systems, prepared by the EU nitrogen expert panel. Wageningen University, Alterra, PO Box 47, NL-6700 Wageningen, Netherlands, pp 32–47
Pawlick AA, Wagner-Riddle C, Parkin GW, Berg AA (2019) Assessment of nitrification and urease inhibitors on nitrate leaching in corn (Zea mays L.). Can J Soil Sci 99:80–91
Peoples MB, Freney JR, Mosier AR (1995) Minimizing gaseous losses of nitrogen. In: Bacon PE (ed) Nitrogen fertilization in the environment. Marcel Dekker Inc, New York, pp 565–602
Rayment GE, Higginson FR (1992) Australian laboratory handbook of soil and water chemical methods. Inkata Press, Melbourne
Rose TJ, Wood RH, Rose MT, Van Zwieten L (2018) A re-evaluation of the agronomic effectiveness of the nitrification inhibitors DCD and DMPP and the urease inhibitor NBPT. Agric Ecosyst Environ 252:69–73. https://doi.org/10.1016/j.agee.2017.10.008
Rowlings DW, Scheer C, Liu S, Grace PR (2016) Annual nitrogen dynamics and urea fertilizer recoveries from a dairy pasture using 15N; effect of nitrification inhibitor DMPP and reduced application rates. Agric Ecosyst Environ 216:216–225. https://doi.org/10.1016/j.agee.2015.09.025
Saggar S, Singh J, Giltrap DL, Zaman M, Luo J, Rollo M, Kim DG, Rys G, der Weerden TJ (2013) Quantification of reductions in ammonia emissions from fertiliser urea and animal urine in grazed pastures with urease inhibitors for agriculture inventory: New Zealand as a case study. Sci Total Environ 465:136–146. https://doi.org/10.1016/j.scitotenv.2012.07.088
Silva AGB, Sequeira CH, Sermarini RA, Otto R (2017) Urease inhibitor NBPT on ammonia volatilization and crop productivity: a meta-analysis. J Agron 109:1–13. https://doi.org/10.2134/agronj2016.04.0200
Smith P, Martino D, Cai Z, Gwary D, Janzen H, Kumar P, McCarl B, Ogle S, O'Mara F, Rice C, Scholes B, Sirotenko O, Howden M, McAllister T, Pan G, Romanenkov V, Schneider U, Towprayoon S, Wattenbach M, Smith J (2008) Greenhouse gas mitigation in agriculture. Philos Trans R Soc Lond Ser B Biol Sci 363:789–813. https://doi.org/10.1098/rstb.2007.2184
Smith RCG, Stephens MJ (1976) Importance of soil moisture and temperature on the growth of improved pasture on the northern tablelands of New South Wales. Crop Pasture Sci 27:63–70
Soares JR, Cantarella H, Menegale MLC (2012) Ammonia volatilization losses from surface-applied urea with urease and nitrification inhibitors. Soil Biol Biochem 52:82–89. https://doi.org/10.1016/j.soilbio.2012.04.019
Sprague SJ, Lilley JM, Bullock MJ, Virgona JM, Kirkegaard JA, Hunt JR, Hopwood MDA, Faulkner MG, Angus JF (2021) Low nitrogen use efficiency of dual-purpose crops: causes and cures. Field Crop Res 267:1–11. https://doi.org/10.1016/j.fcr.2021.108129
Steffen W, Richardson K, Rockström J, Cornell SE, Fetzer I, Bennett EM, Biggs R, Carpenter SR, Vries W, Wit CA, Folke C, Gerten D, Heinke J, Mace GM, Persson LM, Ramanathan V, Reyers B, Sörlin S (2015) Planetary boundaries: guiding human development on a changing planet. Science 347:736–736
Stempfhuber B, Engel M, Fischer D, Neskovic-Prit G, Wubet T, Schöning I, Gubry-RanginC KS, Schloter-Hai B, Rattei T, Welzl G, Nicol GW, Schrumpf M, Francoi B, Prosser JI, Schloter M (2015) pH as a driver for ammonia-oxidizing archaea in forest soils. Microb Ecol 9:879–883
Suter HC, Lam SK, Walker C, Chen D (2021b) Benefits from enhanced-efficiency nitrogen fertilisers in rainfed temperate pastures are seasonally driven. Soil Res 60(2):147–157. https://doi.org/10.1071/SR21083
Suter HC, Lam SK, Walker C, Chen D (2020) Enhanced efficiency fertilisers reduce nitrous oxide emissions and improve fertiliser 15N recovery in a southern Australian pasture. Sci Total Environ 699:134–147. https://doi.org/10.1016/j.scitotenv.2019.134147
Suter HC, Pengthamkeerati P, Walker C, Chen D (2011) Influence of temperature and soil type on inhibition of urea hydrolysis by N-(n-butyl) thiophosphoric triamide in wheat and pasture soils in southeastern Australia. Soil Res 49:315–319
Suter HC, Sultana H, Davies R, Walker C, Chen D (2013a) Influence of enhanced efficency fertilisation techniques on nitrous oxide emissions and productivity response from utea in a temperature Australian ryegrass pasture. Soil Res 54:523–532
Suter HC, Sultana H, Turner DA, Davies R, Walker C, Chen D (2013b) Influence of urea fertiliser formulation, urease inhibitor and season on ammonia loss from ryegrass. Nutr Cycl Agroecosyst 95:175–185. https://doi.org/10.1007/s10705-013-9556-y
Thapa R, Chatterjee A, Awale R, McGranahan DA, Daigh A (2016) Effect of enhanced efficiency fertilizers on nitrous oxide emissions and crop yields: a Meta-analysis. Soil Sci Soc Am J 80:1121–1134. https://doi.org/10.2136/sssaj2016.06.0179
Volpi I, Laville P, Bonari E, di Nasso NN, Bosco S (2017) Improving the management of mineral fertilizers for nitrous oxide mitigation: the effect of nitrogen fertilizer type, urease and nitrification inhibitors in two different textured soils. Geoderma 307:181–188. https://doi.org/10.1016/j.geoderma.2017.08.018
Walker TS, Bais HP, Grotewold E, Vivanco JM (2003) Root exudation and rhizosphere biology. Plant Physiol 132:44–51
Watson CJ, Akhonzada NA, Hamilton JTG, Matthews DI (2008) Rate and mode of application of the urease inhibitor N-(n-butyl) thiophosphoric triamide on ammonia volatilization from surface-applied urea. Soil Use Manag 24:246–253
Williams PH, Haynes RJ (1997) Recovery of N derived from 15N-labelled grass/clover residues, recently immobilised urine 15N or native soil organic N by a wheat crop following cultivation of a pasture soil. Agric Ecosyst Environ 63:67–72
Williams PH, Rowarth JS, Tregurtha RJ (2000) Recovery of 15N-labelled fertiliser by a perennial ryegrass seed crop and a subsequent wheat crop. Nutr Cycl Agroecosyst 56:117–123. https://doi.org/10.1023/a:1009803008544
WRB, IWG (2015) World Reference Base for soil resources 2014, update 2015. FAO, World Soil Resources Reports, Rome
Wu D, Cárdenas LM, Calvet S, Brüggemann N, Loick N, Liu S, Bol R (2017) The effect of nitrification inhibitor on N2O, NO and N2 emissions under different soil moisture levels in a permanent grassland soil. Soil Biol Biochem 113:153–160. https://doi.org/10.1016/j.soilbio.2017.06.007
Yang M, Fang Y, Sun D, Shi Y (2016) Efficiency of two nitrification inhibitors (dicyandiamide and 3, 4-dimethypyrazole phosphate) on soil nitrogen transformations and plant productivity: a meta-analysis. Sci Rep 6:22075. https://doi.org/10.1038/srep22075
Zaman M, Nguyen ML, Blennerhassett JD, Quin BF (2008) Reducing NH3, N2O and NO3−-N losses from a pasture soil with urease or nitrification inhibitors and elemental S-amended nitrogenous fertilizers. Biol Fertil Soils 44:693–705
Zaman M, Saggar S, Blennerhassett JD, Singh J (2009) Effect of urease and nitrification inhibitors on N transformation, gaseous emissions of ammonia and nitrous oxide, pasture yield and N uptake in grazed pasture system. Soil Biol Biochem 41:1270–1280
Zaman M, Zaman S, Adhinarayanan C, Nguyen ML, Nawaz S, Dawar KM (2013) Effects of urease and nitrification inhibitors on the efficient use of urea for pastoral systems. Soil Sci Plant Nutr 59:649–659. https://doi.org/10.1080/00380768.2013.812940
Zaman M, Zaman S, Quin BF, Kurepin LV, Shaheen S, Nawaz S, Dawar KM (2014) Improving pasture growth and urea efficiency using N inhibitor, molybdenum and elemental Sulphur. Soil Sci Plant Nutr 14:245–257. https://doi.org/10.4067/S0718-95162014005000020
Zerulla W, Barth T, Dressel J, von Locquenghien KEKH, Pasda G, Rädle M, Wissemeier AH (2001) 3,4-Dimethylpyrazole phosphate (DMPP) – a new nitrification inhibitor for agriculture and horticulture. Biol Fertil Soils 34:79–84. https://doi.org/10.1007/s003740100380
Zhu G, Ju X, Zhang J, Müller C, Rees RM, Thorman RE, Sylvester-Bradley R (2019) Effects of the nitrification inhibitor DMPP (3,4-dimethylpyrazole phosphate) on gross N transformation rates and N2O emissions. Biol Fertil Soils 55:603–615. https://doi.org/10.1007/s00374-019-01375-6
Zhou X, Wang S, Ma S, Zheng X, Wang Z, Lu C (2020) Effects of commonly used nitrification inhibitors—dicyandiamide (DCD), 3,4-dimethylpyrazole phosphate (DMPP), and nitrapyrin—on soil nitrogen dynamics and nitrifiers in three typical paddy soils. Geoderma 380:114637. https://doi.org/10.1016/j.geoderma.2020.114637
Acknowledgments
This research is supported by funding from the Australian Government Department of Agriculture, Water and the Environment as part of its Rural R&D for Profit Program, Dairy Australia and The University of Melbourne. The authors would like thank the Croft family from Allansford for their significant contribution and cooperation which made this research possible. The authors also thank the many research assistants involved in the project: Dr. Dona Thushari Wijesinghe, Dr. Arjun Pandey, Dr. Bhawana Bhatta, Dr. Robert Impraim, Dr. MD Shahin Rhaminur, Mr. Andrew Longmire and Dr. Robert Edis for soil physics assessment. Analytical results were provided by the Soils Node of the Trace Analysis for Chemical, Earth and Environmental Sciences (TrACEES) platform at the University of Melbourne.
Funding
Open Access funding enabled and organized by CAUL and its Member Institutions.
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Competing interests
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Additional information
Responsible Editor: Zucong Cai.
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Belyaeva, O., Ward, G., Wijesinghe, T. et al. Pasture productivity benefits from strategic urease and nitrification inhibitor use are limited in rainfed temperate dairy pastures of southern Australia. Plant Soil (2024). https://doi.org/10.1007/s11104-024-06675-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11104-024-06675-x