
Abstract
Purpose
Perennial crops have been suggested as a more sustainable alternative to the currently most common cropping systems. Compared with annual plants, perennial plants produce more biomass and have deeper roots, and are expected to lead to higher soil organic carbon (SOC). This hypothesis, however, has not been well tested for grain crops.
Methods
Using perennial intermediate wheatgrass (IWG, Thinopyrum intermedium) and annual winter wheat (Triticum aestivum) as focal species, and native grassland as reference, we quantified the SOC accumulation via a process-based model, describing water and heat exchanges and carbon-nitrogen cycling in the canopy and soil to a depth of 2 m. The model includes C fixation via photosynthesis, plant biomass growth and litter production, physical protection of SOC, depolymerisation, C mineralisation, nitrification, denitrification, microbial growth, and necromass turnover in the soil. While of general applicability, we considered a sandy loam under warm-summer humid continental climate.
Results
Following a conversion from native grassland, IWG reduced SOC losses by at least 38%, especially in the particulate organic carbon (POC) pool, within the top 2 m of soil, compared with annual wheat. Soil microbial biomass and soil respiration were higher in IWG than annual wheat. Shifting from annual wheat to high photosynthetic capacity IWG increased SOC by about 33 g C m−2 y−1 (averaged over a 4-year continuous IWG cropping), with a large fraction of SOC gain stemming from restoring POC.
Conclusion
Compared with annual grains, perennial grains can increase soil carbon sequestration and maintain SOC at levels nearer to that of native grasslands.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
The loss of soil organic carbon (SOC) from the conversion of native ecosystems to agricultural land can compromise the ability of soils to provide ecosystem services important for sustaining food production and biodiversity (Lehmann et al. 2020). SOC can enhance crop productivity through water and nutrient retention, maintain good water quality by retaining or removing pollutants through adsorption and biotic transformation, and mitigate climate change through carbon (C) sequestration (Lal 2016). Accumulating or restoring SOC while maintaining primary production of food, feed and fibre is a key target for sustainable agriculture.
The production of staple food is currently heavily dependent on annual crops. There is a growing interest in exploring perennial grain crops as a more sustainable alternative (Wagoner and Schaeffer 1990; Crews et al. 2018; Glover et al. 2010; Kim et al. 2022). Although current breeds of perennial grain crops typically yield less grain than annuals, there has been a continuous research endeavour by plant breeders and agronomists in recent years to enhance their yield potential (Bajgain et al. 2022), making perennial grain crops more economically viable. As progress is made, the adoption of perennial grain crops could offer significant benefits for sustainable agriculture. Compared with annual species, perennial plants can produce more biomass and have deeper, more extensive and developed root systems (Vico et al. 2016; Sprunger et al. 2018). These plant characteristics can lead to higher litter inputs and root exudation, making perennial crops a better candidate for SOC accumulation than annuals (Crews and Rumsey 2017; Ledo et al. 2020). Indeed, SOC increased after conversions of annual croplands to perennial grasslands or perennial bioenergy crops (Kaempf et al. 2016; Ledo et al. 2020; Anderson-Teixeira et al. 2009; Qin et al. 2016; Crews and Rumsey 2017). In the case of perennial grain, only a few studies quantified the soil C stock change (Sprunger et al. 2018, 2019; Pugliese et al. 2019; van der Pol et al. 2022; Rakkar et al. 2023; Taylor et al. 2023). Among these, one study compared particulate organic matter contents in soils cultivated with annual winter wheat and perennial intermediate wheatgrass (IWG, Thinopyrum intermedium) over a 4-year period in Michigan (Sprunger et al. 2018). The researchers found no significant differences, but this was not surprising given the sandy texture of the soil and the short duration of the cropping system. Negligible or marginal increase in SOC under perennial grain cropping was also observed in two other recent short-term field experiments (Rakkar et al. 2023; Taylor et al. 2023). Detecting substantial changes in total SOC within a timeframe less than 5 years is challenging (Necpálová et al. 2014), and hence, relying solely on field studies to assert the potential of IWG to enhance SOC within a realistic timeframe could be misleading. In contrast, another study comparing annual grains and IWG across three paired sites, where IWG had been cultivated for 5–17 years, suggested the potential for IWG to increase SOC at depth, particularly in the form of particulate organic matter (van der Pol et al. 2022). These contrasting results underscore the necessity of employing mathematical models to comprehensively explore the capacity of perennial grain crops to enhance SOC within a timeframe that aligns with practical cropping practices.
It is difficult to predict how SOC stock differs between annual and perennial grain cropping systems because the mechanisms at play have opposite effects. A deeper and more extensive root system can lead to higher amounts of SOC at a greater depth through root exudation in the short-term and root mortality and decomposition in the long-term (Thorup-Kristensen et al. 2020). But increased root exudates in some cases promote rhizosphere priming by stimulating microbial activity and mobilising mineral-associated (protected) organic matter, potentially resulting in a net loss of SOC (Bengtson et al. 2012; Keiluweit et al. 2015). In parallel, increased microbial growth leads to necromass formation, which tends to attach onto mineral surfaces or “stick” within soil aggregates, forming mineral-associated organic matter that is relatively stable and has long turnover time (Cotrufo et al. 2015; Kallenbach et al. 2016). Annual and perennial grains differ also in water use patterns and canopy-atmosphere coupling (Vico and Brunsell 2018; Sutherlin et al. 2019; Vico et al. 2023), which can affect soil water availability that in turn mediate microbial activity and the C cycle. Because the feedbacks between plants, soil microbes, and abiotic soil processes are highly non-linear, the net effect of these intricate interactions in perennial grain cropping systems remains largely unknown.
We investigated the role of annual and perennial grain cropping on SOC stock and C cycling via a plant-soil mechanistic model, representing water and heat flows along a continuous canopy-soil profile and accounting dynamically for crop development. The model also includes coupled C-N cycles that explicitly describe SOC protection, depolymerisation, C mineralisation, nitrification, and denitrification. The model also represents the microbial dynamics of five microbial functional groups, capturing soil temperature and moisture effects on microbial growth and necromass turnover. We hypothesised that (1) perennial grain cropping can maintain a SOC level nearer to a multi-species perennial C3 grassland than the annual crop; and (2) perennial grain crops can increase SOC in a soil that has a history of continuous annual wheat cropping. While the model is of general applicability, we use annual winter wheat (Triticum aestivum) and perennial wheat undergoing domestication (intermediate wheatgrass, IWG, Thinopyrum intermedium) as the model crops, and a multi-species perennial C3 grassland as the reference. We parameterised and benchmarked the model for annual wheat and the perennial IWG, assuming both systems were subject to the same management practices. To facilitate the comparison with the grassland system, we assumed no till, rainfed cropping, and realistic fertilisation based on common wheat cropping practices.
Methods
Model description
Model overview
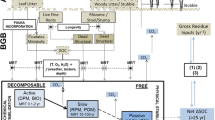
The model simulates the energy, water, C, and nitrogen (N) flows within the coupled plant-soil system, via four coupled modules (Fig. 1a): (i) the canopy module, simulating crop C assimilation and respiration, and transpiration, as a homogenous single crop layer (‘big leaf’; Luan and Vico 2021); (ii) the crop growth module, simulating the allocation of assimilated C to five different crop compartments; (iii) the soil C and N cycles module, considering above and belowground litter inputs, root exudation, depolymerisation, C mineralisation, nitrification, denitrification, and microbial growth and turnover (Pasut et al. 2021); and (iv) the vadose zone module (Maggi 2019), simulating soil water and heat dynamics over a multi-layer soil column. The model was solved numerically, at a sub-daily time step, except for the crop growth module, which was solved at a daily time step.
The vadose zone and soil C-N cycle modules included in this model have been previously tested in several field to global-scales studies (Ceriotti et al. 2020; Pasut et al. 2021; Guglielmo et al. 2021), whereas the canopy and crop growth modules were developed based on existing models (Farquhar et al. 1980; Fatichi et al. 2012; Luan and Vico 2021), resting on well-established principles.

a Scheme of the plant-soil coupled model and b the simulated scenarios. IWG-H and IWG-L refer to intermediate wheatgrass with high and low photosynthetic capacity, respectively
Canopy module: crop C and water exchange
The canopy module determines the canopy C, water, and energy fluxes, accounting for the environmental conditions (air temperature and humidity, solar radiation, soil water content). The net CO2 assimilation and dark respiration were estimated using Farquhar photosynthesis model (Farquhar et al. 1980). The kinetic parameters controlling photosynthesis rate (i.e., the maximum rates of carboxylation, \({V}_{cmax,25}\), and electron transport, \({J}_{max,25}\), at 25˚C) were modelled as functions of canopy temperature (Bernacchi et al. 2001) and soil water availability (Vico and Porporato 2008). The module is described in detail in Luan and Vico (2021). We report here and in the Supplementary Information only the modifications implemented for the coupling to the multi-layered vadose zone module.
The canopy water and energy balances were solved at an hourly time step, forced by observed atmospheric conditions. The model partitions the global solar radiation into the near-infrared and ultraviolet-visible wavebands and calculates their absorptions by crop canopy and the total shortwave radiation reaching the soil surface (see Supplementary Information, Text S1). To account for the effect of soil temperature, we determined the net longwave (thermal) radiation absorbed by the canopy as the difference between the incoming thermal radiation from the sky (downward) and soil (upward), and the outgoing thermal radiation from the canopy to the sky (upward) and soil (downward), following Tuzet et al. (2003) (see Supplementary Information, Text S1). The longwave radiation reaching the soil surface is the sum of the downward sky thermal radiation not absorbed by the canopy and the thermal radiation emitted by the canopy (see Supplementary Information, Text S1). The radiation absorbed by the canopy was then combined with latent and sensible heat fluxes to determine the canopy temperature as in Luan and Vico (2021). For simplicity, we set gross photosynthesis to zero when solar radiation is below 10 W m-2, air temperature below 0 oC, or average water content along the soil profile below the near-wilting point (i.e., at soil water potential of -1.4 MPa), but respiration and transpiration rates were solved for also under these conditions.
The transport of water through the soil-plant-atmosphere continuum (SPAC) was modelled as gradients of total water potential across a series of conductances (Manzoni et al. 2013), from soil to root, root to leaf (i.e., xylem), and leaf to the atmosphere (i.e., stomatal). We calculated the soil to root conductance \({g}_{sr}\) along the soil profile by considering the layer-specific soil hydraulic conductivity, root length density and volume fraction (Lhomme 1998) (see Supplementary Information, Text S2). Root to leaf conductance was assumed to decline with leaf water potential, from a maximum value, \({g}_{p,max}\), dependent on leaf area index (LAI). Leaf to atmosphere conductance was determined as the series of stomatal conductance, estimated based on an optimality principle (Vico et al. 2013), with canopy boundary layer conductance depending on wind velocity (Campbell and Norman 1998), and aerodynamic bulk conductances considering the roughness length and diabatic correction for momentum (Webber et al. 2016). Knowledge of the conductances along the SPAC allows determining canopy transpiration rate and temperature (Luan and Vico 2021).
The crop transpiration, absorbed radiation, and canopy temperature drive the subsequent iteration of the vadose zone module providing soil water and heat dynamics over a multi-layer soil column. At each time step, the simulated soil temperature and water content feedback into the canopy module to regulate canopy temperature and stomatal conductance.
Crop growth module
The net primary productivity (NPP, i.e., the net C gain from photosynthesis) was calculated as the gross C assimilated through photosynthesis minus the sum of maintenance and growth respiration of the whole plant. The leaf maintenance respiration rate (i.e., the sum of light and dark respiration) was obtained from the Farquhar model described above, while the maintenance respiration of roots and stems was calculated as a function of their N content following Fatichi et al. (2012). The growth respiration of the whole plant was determined as a fraction of the remaining C after deducting the cost of maintenance respiration as in Fatichi et al. (2012) (see details in Supplementary Information, Text S3).
NPP was allocated, depending on the growth stage (a function of growing degree days, GDD), to five compartments: leaf, stem, root, grain, and labile C used for root exudation. The size of the compartments and their attributes (e.g., LAI and root length density) evolve dynamically over time, as the result of newly allocated C and senescence of leaves, stems, and roots. The dead tissues return to the soil feeding the soil C and N cycles, while grain and parts of the leaves and stems were harvested for food and feed. The dynamics of crop growth were solved at a daily time-step.
We also modelled the dynamic of N content in each compartment based on a compartment-specific reference C:N ratio. Hourly N uptake by crops depends on N demand and availability in soil (see details in Supplementary Information, Text S3).
Soil C and N cycle
To simulate the dynamics of soil organic C and N, we used a simplified version of C-N coupled reaction network (Biotic and Abiotic Model for SOM-version 4, BAMS4; Pasut et al. 2021). The original BAMS4 includes modules for methane and sulfur cycles, but they were switched off here. The C-N reaction network consists of nine pools, including two SOC polymer pools, PolyC (representing lignin, cellulose, and hemicellulose) and PolyCN (representing peptidoglycan), and two low-molecular weight SOC pools, MonoC (representing monosaccharides, organic acids, fatty acids, and phenols) and MonoCN (representing amino acids, amino sugars, and nucleotides). Finally, we represented five microbial functional groups (Supplementary Information, Figure S2). The decomposition of SOC pools and the transformation of N into various oxidation states depend on microbial activity, which is affected by soil temperature, soil water availability, oxygen content, and nutrient availability. The dynamics of microbial functional groups were explicitly modelled, including microbial assimilation of C and N, growth, mortality, and necromass turnover. The root and straw litters decompose into simpler polymers and low-molecular weight organic C (Riley et al. 2014), while root exudates contain only low-molecular weight organic C (Grayston et al. 1997; Warren 2016). Urea input via fertilization is hydrolysed into NH4+, which is then taken up by crops, assimilated by microorganisms, and nitrified to NO2- and NO3-. The latter compounds were modelled explicitly to describe denitrification and production of NO, N2O and N2.
SOC pools (PolyCN, MonoC, and MonoCN), urea, NH4+, and NO3- can become physically protected by soil minerals and aggregates. The protected forms (or known as mineral-associated) are assumed to be inaccessible to microbial activity and crop uptake. All aqueous chemicals can undergo advection, diffusion, and gas dissolution. Details of the C-N reaction network are reported in Text S4 and Table S3 of the Supplementary Information.
Vadose zone module
We used the general-purpose multi-phase and multi-component BioReactive Transport Simulator, BRTSim v5.0a (Maggi 2019) to solve for the dynamics of water, heat, and biogeochemistry in the vadose zone. BRTSim simulates, over a one-dimensional variably saturated soil column, the water, gas, and heat flows, the transport of aqueous and gaseous chemical species, and the kinetics of biotic and abiotic processes included in the C-N coupled reaction network. The water flow is modelled using the Richards equation (Richards 1931) with the Brooks–Corey relative permeability-water potential-saturation relationship (Brooks and Corey 1964). The advection of dissolved chemicals is described by Darcy’s equation, while the diffusion of aqueous and gaseous compounds is based on Fick’s law. Chemical and biochemical kinetic reactions are described using Michaelis-Menten-Monod kinetics (Michaelis and Menten 1913; Monod 1949). The aqueous complexation and gas dissolution are represented based on the mass action law. The protection of SOC pools is modelled using Langmuir kinetics, while the protection of NH4+, NO3-, and urea is modelled using Freundlich’s adsorption isotherms. Detailed description of the equations used to model the above processes are presented in the User Manual and Technical Guide of BRTSim v5.0a (Maggi 2021), with the key equations used here reported in Text S4 and S5 of the Supplementary Information.
Boundary conditions are water, heat, and gas exchanges between the soil and the air and canopy at the top of the soil column, while a single large element with minimal heat and gas exchange is used to represent free drainage at the bottom. The model does not include horizontal runoff and the interaction with groundwater. Water input occurs via precipitation (and possible ponding) and losses via soil water evaporation and plant transpiration. Losses via soil water evaporation occur only in the first soil layer and depend on the potential evapotranspiration, soil saturation, and the canopy cover fraction (Raes et al. 2009). The transpiration flux calculated by the canopy module is split over the soil profile proportionally to the root fraction distribution profile calculated as in Schenk and Jackson (2002) (see Supplementary Information, Text S3).
Vegetation parameters: annual and perennial wheats, and native grassland
While the model is of general applicability, we parameterised it for winter wheat (Triticum aestivum) as our model annual crop and intermediate wheatgrass (Thinopyrum intermedium) as the perennial counterpart. The perennial Thinopyrum intermedium is undergoing de novo domestication as a crop species and produces grain sold under the trademark Kernza®. Based on literature data, we expect the two crops to differ in canopy characteristics, photosynthetic activity, biomass allocation patterns, and root characteristics, as detailed next. Values of the parameters are reported in Table S2 of the Supplementary Information.
Compared with winter wheat, IWG could achieve a greater LAI and a higher canopy height at maturity (Tinsley 2012) with thicker leaves (i.e., lower specific leaf area, SLA; Jaikumar et al. 2013). IWG also allocated more biomass belowground, but less to grains (Sprunger et al. 2018). Its roots were deeper (Sprunger et al. 2018) and slightly coarser (i.e., had larger average root diameter; Duchene et al. 2020). The growing degree days (GDD) required for a certain phenological stage, defined as the sum of daily temperatures above 0 °C, were generally greater for IWG than annual wheat, based on data from Jungers et al. (2018) for IWG and from USDA (2021) for winter wheat.
There was more variability in terms of photosynthetic capacity among perennial species (Jaikumar et al. 2014; Vico et al. 2016). We thus tested two IWG, differing in their photosynthetic capacity, i.e., either higher (IWG-H) or lower (IWG-L) than the winter wheat. IWG-H was assumed to have 36% and 53% higher maximum carboxylation and electron transport rate at 25 °C (\({V}_{cmax,25}\) and \({J}_{max,25}\)) than annual wheat, based on gas exchange measurements on IWG (Jaikumar et al. 2014). IWG-L was assumed to have 11.3% lower \({V}_{cmax,25}\) and \({J}_{max,25}\) than annual wheat, based on the average differences between congeneric perennials and annuals (Vico et al. 2016).
Plant conductance under well-watered conditions (\({g}_{pmax}\)) was determined from a reference value estimated for annual wheat (Lawlor 1973); subscript ref), after rescaling by the time-dynamic LAI of each modelled crop, i.e., \({g}_{pmax}\)=\({g}_{pmax,ref}\times \text{L}\text{A}\text{I}/{\text{L}\text{A}\text{I}}_{ref}\), with \({\text{L}\text{A}\text{I}}_{ref}=3\)mleaf2 mground-2.
We considered a native grassland as reference. We used vegetation parameters corresponding to those of a C3 grassland (see details in Table S2 of the Supplementary Information). All the other parameters were assumed to be the same for all systems.
Environmental conditions
The model was run based on environmental conditions observed at W.K. Kellogg Biological Station in Hickory Corners, MI, USA (42˚24’N, 85˚24’W), for easy comparison with the wealth of observations available for this site. The site has a warm-summer humid continental climate. Time series of hourly global solar radiation, air temperature, precipitation, and relative humidity from 2012 to 2020 were obtained from the U.S. Climate Reference Network (Diamond et al. 2013). The monthly potential evapotranspiration was sourced from the Climatic Research Unit time-series datasets (Harris et al. 2020). Wind speed was assumed to be constant at the annual average value (Diamond et al. 2013) of the modelled site. Atmospheric deposition of NH4+ and NO3− was set to an annual rate of 0.5 g N m−2 y−1 (Woodmansee 1978).
Soil properties for the location of W.K. Kellogg Biological Station were extracted from gridded data. The site has a sandy loam soil. We used SoilGrids2.0 dataset (Poggio et al. 2021) for the soil bulk density and organic C content, while soil porosity was obtained from the SoilGrids1.0 dataset (Hengl et al. 2017). Soil permeability, pore volume distribution index and air-entry suction of the Brooks–Corey model, soil heat capacity, and heat conductivity were obtained from Dai et al. (2019), and soil residual liquid saturation from Zhang et al. (2018). Soil properties at different soil depths are reported in Table S4 of the Supplementary Information.
Simulations
To compare the impacts of annual and perennial grain crops on SOC dynamics, we contrasted two cropping systems (Fig. 1b): (i) the annual winter wheat system, corresponding to an annual winter wheat monoculture with bare soil outside the growing season; and (ii) the perennial IWG system. No tillage was assumed for both systems, so that soil characteristics (e.g., porosity and bulk density) were considered time invariant and homogeneous within each layer including the top one (i.e., 0–15 cm).
We initialised the biogeochemical fluxes and pools based on a 1000-year spin-up simulation, reaching near steady state conditions, for an unmanaged native C3 grassland, beginning with SOC extracted from SoilGrids2.0 (Poggio et al. 2021). The climatic conditions for the spin-up were those from 2012 to 2020, with years re-occurring in randomised order. The simulated above and belowground biomass of the grassland was within ranges reported for grasses at the W.K. Kellogg Biological Station (Lei et al. 2021), suggesting the modelled status at the end of the spin-up simulation represents conditions of a native grassland.
To investigate the role of land-use history on SOC dynamics in annual and perennial systems, we considered two scenarios (Fig. 1b). In Scenario 1, we assumed the native grassland was converted to winter wheat and IWG without tillage, i.e., we initialised the model with the soil conditions at the end of the 1000-year spin-up simulation and ran it with parameters relative to winter wheat and IWG for 4 growing seasons, corresponding to climatic conditions observed during 2012–2016. The IWG stand age was limited to 4 years to minimise the impact of grain yield decline as the plant ages (Tautges et al. 2018), which was not accounted for in our model. In Scenario 2, we assumed that 4 years of the annual winter wheat cropping were followed either by an additional 4 years of the same crop or by IWG (converted from the annual system without tillage), i.e., the soil conditions at the end of the 4th growing season of annual winter wheat were used to initialise both winter wheat and IWG in this second scenario. Climatic conditions corresponded again to 2012–2016. In both scenarios, land conversion was modelled as occurring without tillage. In practice, a no-till conversion can be challenging and may require herbicide applications and a one-year transition period, in particular the conversion of grassland to cropland.
Sowing and harvesting dates were set based on Sprunger et al. (2018). In both scenarios, winter wheat was sown every year in early-October (i.e., on the day of the year, DOY 273), while IWG was sown only once in the first year on the same day. Winter wheat was harvested in mid-July (on DOY196), while IWG was harvested in late August (on DOY 238) every year. Both crops were harvested for grains and straws. At harvest, IWG was cut to a canopy height of 0.2 m and a LAI of 0.5 mleaf2 mground−2, with all cut straw removed from the field. For winter wheat, a portion of leaves and stems equivalent to LAI of 0.5 mleaf2 mground−2 was assumed to be left on the ground and dead roots were left in the soil without tillage. Both winter wheat and IWG systems were fertilised with urea at an annual rate of 90 kg N ha−1 y−1. Details of sowing, harvesting, and fertilisation practices are reported in Text S6 of the Supplementary Information.
Model output analyses
For the analyses, we consolidated the various SOC pools into three main groups: (i) particulate organic carbon (POC), comprising PolyC(s) and PolyCN(s) in solid phase, (ii) dissolved organic carbon (DOC), comprising MonoC(aq) and MonoCN(aq) in aqueous phase, and (iii) mineral-associated organic carbon (MOC), comprising PolyCN(p), MonoC(p) and MonoCN(p) in the protected form.
In Scenario 1, changes in SOC pools (∆G SOC) were calculated with reference to the initial condition of the grassland, whereas, in Scenario 2, ∆W SOC was expressed with reference to the soil condition of the winter wheat system at the end of the 4th year after the conversion from grassland.
Results
Crop biomass and leaf area index
IWG-H and IWG-L had higher cumulative annual NPP (Figure S4, Supplementary Information), straw and root biomass at harvest and LAI at flowering than winter wheat, but lower grain biomass (Fig. 2). Between the two IWGs, IWG-H had slightly higher cumulative annual NPP and root and straw biomass than IWG-L, thanks to its higher photosynthetic capacity.
Under the same environmental conditions, the modelled LAI and biomass of root, straw, and grain of winter wheat and IWG matched well those observed at the W.K. Kellogg Biological Station (Sprunger et al. 2018; Tinsley 2012; Fig. 2). Although the modelled above and belowground biomass were in line with field observations, our estimated NPP (500–700 g C m−2 y−1 for winter wheat and 650–1170 g C m−2 y−1 for IWG; Figure S4) was slightly higher than values commonly estimated by remote sensing and other process-based models for winter wheat in the USA (350–600 g C m−2 y−1; Li et al. 2014) and higher than the net ecosystem exchange measured in IWG cropping fields based on eddy covariance fluxes (-200 – -810 g C m−2 y−1; de Oliveira et al. 2018; Wiesner et al. 2022). This is because our estimation included C allocation to root exudates, which was generally neglected in remote sensing-based estimates and process-based models. After deducting the contribution of root exudates, our modelled average annual NPP for winter wheat, IWG-H, and IWG-L was 560 g C m−2 y−1, 940 g C m−2 y−1 and 817 g C m−2 y−1 respectively, closer to the empirical estimates.

a root, b straw, and c grain biomass of winter wheat (annual) and intermediate wheatgrass (IWG, perennial) at harvest, and d leaf area index (LAI) at flowering, for Scenario 1. The bars represent the range of modelled values for winter wheat across 4 growing seasons and for matured (older than one year) IWG systems during their respective growing seasons. The diamond markers indicate the modelled values for IWG systems in their first year. IWG-H and IWG-L are intermediate wheatgrass with high and low photosynthetic capacities respectively. For comparison, the biomass of root, straw, and grain of winter wheat and IWG observed at the end of the 4th growing season (Sprunger et al. 2018) and LAI of IWG at the flowering of the 2nd year (Tinsley 2012) are reported (circle and triangle respectively)
SOC balance
SOC inputs
IWG-H and IWG-L provided about 5–6 times higher SOC input than winter wheat in both scenarios, with IWG-H having the highest total input (Fig. 3, top row, grey bars). More than 95% of the SOC input in IWG systems came from belowground inputs (i.e., root litter and root exudates), whereas belowground inputs constituted 67% of the total SOC input in the annual system. In both winter wheat and IWG systems, POC accounted for approximately 60% and 40% of the total C input, respectively, and was primarily originated from root litter. Root exudates contributed to about 14% and 34% of the SOC input in winter wheat and IWG systems, respectively, in the form of DOC.
Our model suggests that winter wheat returned approximately 20% of its NPP to the soil, with only about 2% of its NPP allocated to root exudates. These values are slightly lower than those emerging from a meta-analysis, which found that winter wheat allocates an average of 7% of its NPP to rhizodeposition, and about 26% of its NPP is returned to the soil when the majority of aboveground biomass is harvested (Bolinder et al. 2007). Modelled IWG allocated 6–14% of its NPP to root exudates, starting from the second year after sowing (i.e., Year 2–4; see Figure S5 in Supplementary Information). Our estimates of NPP allocation to root exudates in IWG systems are lower than the 17–30% measured in perennial forage fields (Bolinder et al. 2007). In another field study using multiple-pulse labelling with 13CO2 and 14CO2, IWG net rhizodeposited C during one growing season in the top 1.5 m of soil was about 160 g C m−2 on average, with a range between 70 and 250 g C m−2 (Peixoto et al. 2022). In a separate long-term field experiment conducted in Montana, USA, the rhizodeposition of IWG in the top 1.2 m of soil was estimated to be about 41 g C m−2 (Sainju and Allen 2023). In comparison, our model estimated an average of 190 g C m−2 y−1 and 160 g C m−2 y−1 of root exudation in the top 2 m of soil for IWG-H and IWG-L, respectively. Our modelled root exudation accounted for about 33% of the total belowground C input, matching well another field experiment, where net rhizodeposition of IWG was 12–44% of the total belowground C input across different soil layers down to 2.4 m (Peixoto et al. 2020). It is worth noting that rhizodeposition includes root exudates and other organic materials such as sloughed-off cells, dead roots, and mucilage, which were not included in our model.
SOC losses
While having a higher SOC input to soil, IWG systems also had a higher annual rate of SOC loss than the winter wheat system in both scenarios (Fig. 3, bottom row). The balance between input and loss resulted in a net SOC loss in the top 2 m of soil in the IWG systems in Scenario 1 and a net gain in IWG-H system in Scenario 2, whereas the winter wheat system experienced a net loss in both scenarios (Fig. 4).
In both scenarios, the mineralisation of DOC was the primary process governing the SOC loss in the top 2 m of soil in both annual and IWG systems, accounting for more than 90% of the loss (Fig. 3). IWG systems had substantially higher microbial degradation of N-containing DOC than the annual system. Leaching of DOC to below 2 m was the second largest contributor to SOC loss, contributing an average annual loss of 8–50 g C m−2 y−1. Our estimates of DOC leaching to below 2 m are higher than the average DOC leaching rate (1.53 g C m−2 y−1) measured in the same site for which the model is parameterised (Hussain et al. 2020), but for some years our estimates are within the measured range (0.2–3.8 g C m−2 y−1). We note that DOC leaching rates varies greatly over time (Figure S6 in Supplementary Information). It is worth noting that DOC leaching to below 2 m does not imply a C loss because it can contribute to deep C sequestration.

Average annual soil organic carbon (SOC) input rates (top) and average annual rates of SOC losses within the top 2 m of soil (bottom), averaged over 4 growing seasons after the conversion of grassland to winter wheat and IWG (a, c; Scenario 1), and the conversion of winter wheat to IWG (b, d; Scenario 2). POC: particulate organic carbon; DOC: dissolved organic carbon. Depoly: depolymerisation; Min: mineralisation
SOC content in the top 2 m of soil
Scenario 1: winter wheat and IWG cropping following a native grassland
After the conversion of grassland to winter wheat or IWG (Scenario 1), SOC stock within the top 2 m of soil decreased relative to the stocks of the long-term grassland (Fig. 4a, grey bars). Annual winter wheat system lost the most SOC and IWG-H the least.
Changes in SOC varied in time and along the soil profile. The greatest SOC loss occurred within the top 50 cm of soil and in the first two years after the conversion (Figure S7a and S8a, Supplementary Information). Over 4 growing seasons, in the top 30 cm, SOC stock in annual wheat decreased about 4%, by 330 g C m−2 (i.e., 82 g C m−2 and 1% per year), but less than 3% in IWG systems, by about 83 g C m−2 and 150 g C m−2 in IWG-H and IWG-L respectively (i.e., 21 g C m−2 y−1 and 37 g C m−2 y−1; Figure S7a). Our estimates of SOC loss aligned well with field observations of losses of 22–25 g C m−2 y−1 in the top 20 cm of soil after converting grassland to cropland (Oberholzer et al. 2014). However, our estimated SOC loss rates were slightly lower than values reported in a meta-analysis that found an average SOC loss of about 36% in the top 30 cm after 20 years of conversion from grasslands to croplands, i.e., about 1.8% loss per year (Poeplau et al. 2011).
In all systems, POC and MOC constituted more than 99% of the SOC, with less than 1% being DOC (Figure S9), as commonly observed (Bai and Cotrufo 2022). PolyCN and PolyCN(p) that represent peptidoglycan derived from microbial necromass made up about 70% of SOC in the top 30 cm of soil, in line with the observed 23 – 74% necromass contribution to SOC in the topsoil of global grasslands receiving an annual precipitation of 800–900 mm, similar to our modelling site (Bai and Cotrufo 2022).
Not all SOC pools experienced a net loss (Fig. 4a, colours). In both annual and perennial systems, there was a net gain in mineral-associated PolyCN(p) (i.e., the protected form of peptidoglycan derived from microbial necromass). In IWG systems, there was also a net gain in the aqueous and protected forms of N-containing low-molecular weight SOC (MonoCN(aq) and MonoCN(p), representing amino acids, amino sugars, and nucleotides). Among the pools with a net loss, winter wheat displayed the greatest decrease in PolyC(s) that represents lignin, cellulose, and hemicellulose derived from root and straw litters, contributing to about 49% of the total net SOC loss. Mineral-associated MonoC(p) that represents the protected forms of monosaccharides, organic acids, fatty acids, and phenols also showed a decrease in all systems.
Over the soil profile, more than 80% of losses in POC occurred in the top 100 cm independent of the cropping system (Figure S10a – d, Supplementary Information). Losses in mineral-associated MonoC(p) occurred mostly in soils below 45 cm in winter wheat, while the IWG systems had a slight gain in MonoC(p) between 1 and 2 m.

Change in soil organic carbon (SOC) content in the top 2 m of soil at the end of the 4th-year after the conversion of a grassland to winter wheat and IWG (reference: near steady state grassland), and b winter wheat to IWG (reference: after 4 years of winter wheat). POC: particulate organic carbon; DOC: dissolved organic carbon; MOC: mineral-associated organic carbon. X(s), X(aq), X(p) denotes carbon pool X in solid, aqueous, and protected phases respectively
Scenario 2: winter wheat and IWG cropping following 4-year winter wheat cropping
Four years after the conversion of the winter wheat monoculture to IWG (Scenario 2), IWG-H had led to a 132 g C m−2 gain in the top 2 m of soil, corresponding to approximately 33 g C m−2 y−1 (Fig. 4b). The estimated SOC gain in IWG-H is slightly lower compared to the average annual gain rates of 75–100 g C m−2 y−1 reported in meta-analyses for the conversion of agricultural land to grassland (Conant et al. 2001; Bai and Cotrufo 2022). However, it aligns reasonably well with the estimate of 40 g C m−2 y−1 of SOC gain in IWG field reported in a study comparing three IWG and annual paired sites, despite those sites have lower NPP than our modelled site (van der Pol et al. 2022). The relatively low SOC gain estimated in this study could potentially be attributed to the sandy nature of the soil. However, this SOC gain did not offset the loss caused by the conversion of grassland to annual cropland.
IWG-L did not lead to a gain in SOC, although it had more than 10 times lower C loss than annual winter wheat that was not converted to IWG. The winter wheat system continued to experience SOC loss in the top 2 m of soil at an annual rate of 106 g C m−2 y−1, with more than 70% of the losses contributed by mineral-associated MonoC(p), of which 55% occurs in deeper soils (1 to 2 m, Figure S10e – h, Supplementary Information). This result is in contrast to a field comparison of SOC in IWG and spring wheat, showing higher SOC content in spring wheat at a soil depth of 60–90 cm (Taylor et al. 2023), whereas our modelled results indicated that winter wheat system experienced a higher SOC loss at the same depth compared to IWG systems (Figure S10e – h, Supplementary Information).
Biomass of soil microbial functional groups
The modelled total microbial biomass within the 2 m soil profile ranged over 35–120 g C m−2, of which 60–70% in the top 30 cm (i.e., 30–85 g C m−2). These estimates are consistent with the microbial biomass measured (59–100 g C m−2) in the top 20 cm of soil in a fertilised annual wheat field with no cover crop at the Kellogg Biological Station (Willson et al. 2001).
After the conversion of grassland to winter wheat and IWG (Scenario 1), the biomass of fungi and heterotrophic bacteria decreased over time, with the winter wheat system showing the sharpest decline (Fig. 5, left column). This pattern is in line with reductions in microbial biomass and enzymatic activity following the conversion of grassland to cropping fields measured in field experiments (Raiesi and Beheshti 2014; Tang et al. 2022). After 4 growing seasons, the heterotrophic bacteria biomass in the winter wheat system was approximately 2 times lower compared with that in the IWG system. IWG could sustain a relatively high biomass of fungi and heterotrophic bacteria, to a level just slightly lower than that of grassland. Conversely, the conversion of winter wheat to IWG-H (Scenario 2) did not immediately restore fungi and bacteria biomass (Fig. 5, right column). It took more than 4 growing seasons for the bacteria biomass in IWG systems to stabilise to a new value, which was however lower than the values in Scenario 1.

Biomass of fungi (top row) and heterotrophic bacteria (bottom row) over 4 growing seasons in Scenario 1: the conversion of grassland to winter wheat and IWG (left column) and Scenario 2: the conversion of winter wheat to IWG (right column). The blue circles represent the endpoint of Scenario 1 in the left column, which coincides with the start-point of Scenario 2 (right column)
Soil CO2 emissions
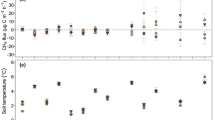
IWG systems had higher soil CO2 emissions than the winter wheat (Fig. 6). There was no substantial difference in the emissions between the two IWG with different photosynthetic capacity. After the second season following the conversion from grassland (Scenario 1), IWG systems reached an annual CO2 emission rate of about 600 g C m−2 y−1. Conversely, winter wheat stabilised around 300 g C m−2 y−1 annual emission after 4 years of conversion (Fig. 6a). When winter wheat was converted to IWG (Scenario 2), CO2 emission increases over the years and reaches about 600 g C m−2 y−1 after the third growing season (Fig. 6b).
According to our model analysis, the higher soil respiration in the IWG systems was caused by the higher SOC inputs and a higher microbial biomass. The higher soil respiration in IWG cropping is in line with results of a field experiment comparing annual wheat against four perennial wheat genotypes in Central Italy (Baronti et al. 2022). Overall, our modelled soil CO2 emissions fall within the observed ranges for temperate croplands and grasslands (Bond-Lamberty and Thomson 2018). Our modelled CO2 emissions in IWG systems (i.e., 200–700 g C m−2 y−1) closely matched the observations from a long-term IWG cropping experiment in Montana, USA, which recorded a CO2 emission of about 425–774 g C m−2 y−1 (Sainju and Allen 2023), thus supporting our estimates of C losses to atmosphere through heterotrophic respiration.

Annual soil CO2 emissions over 4 growing seasons in a Scenario 1: the conversion of grassland to winter wheat and IWG and b Scenario 2: the conversion of winter wheat to IWG
Discussion
Perennial grain cropping reduces SOC loss when compared with annual cropping
Following a conversion from native grassland, perennial grain crops can reduce at least 38% of the SOC loss that would otherwise occur in the annual grain cropping at all soil depths (0–2 m) at our modelling site in Michigan, USA. This holds even for the perennial with photosynthetic capacity lower than the annual grain. In particular, perennial grains reduced the loss of particulate organic carbon (POC) and mineral-associated non-N containing low-molecular weight organic C (e.g., sugars, organic acids, fatty acids, phenols). The mineral-associated organic carbon (MOC) pools in our model are defined based on molecular composition and thus do not exactly match the definition by size commonly used in previous studies (Benbi et al. 2014; Lugato et al. 2021). Nevertheless, the transformation of C into MOC in our model considered both the microbial (via formation of necromass) and physical (via association with mineral surfaces and protection within aggregates) transfer pathways (Cotrufo et al. 2015), an approach similar to the Millennial model (Abramoff et al. 2018). The mineral associated and protected organic C constitutes a large fraction of SOC stock (Rumpel and Kögel-Knabner 2011) and is expected to be less vulnerable to disturbance than the unprotected particulate matter (Cotrufo et al. 2015). However, the continuous disturbance in croplands can increase the vulnerability of the mineral associated/protected organic carbon (Beniston et al. 2014), causing them to be more sensitive to changes in climatic patterns than that in native ecosystems. Perennial grain cropping has the potential to reduce the loss of MOC and increase the resilience of SOC.
The advantage provided by perennials on SOC, POC and MOC can be explained by IWG larger belowground biomass, and hence a higher root litter input and root exudation. Nevertheless, the advantages of perennial over annual systems in terms of SOC are likely underestimated by our model, because no tillage was assumed for both systems. Perennial grain cropping often reduces soil physical disturbances, compared with tilled annual ones. These disturbances can break soil macroaggregates, exposing the previously protected SOC and leading to increased microbial access (Six et al. 2000). Hence, the perennial grain cropping that reduces physical disturbance may have an even bigger potential to reduce SOC loss compared with tilled annual cropland (Crews et al. 2018). However, to what extent reduced disturbance can affect SOC stock remains uncertain, especially for the deeper soil layers (Ogle et al. 2019). Most of the field experiments reporting positive effect of reduced tillage on SOC stock focused only on the top 30 cm of soil, while the few studies that sampled below the 30 cm depth reported no consistent accrual of SOC (Baker et al. 2007).
Perennial grain crops can restore SOC following annual crops in the long run
We showed that the conversion of winter wheat to IWG with high photosynthetic capacity increased the size of all SOC pools (except MonoC(p) and MonoCN(p)) after the first growing season at our modelling site. At depths above 45 cm, a large fraction of the SOC gain stems from restoring POC (Figure S10, right column). However, the original SOC level of grassland (i.e., the level initialised prior to model run for Scenario 1) was not restored even after 4 years of continuous cultivation of perennial grains.
In our simulation, the perennial cropping system accumulated about 150–250 g C m−2 of SOC more than annual wheat in the soil top 30 cm after 4 growing seasons (Figure S7b). However, this increase corresponds to less than 4% of the total SOC stock, which remains below the minimum detectable difference (Necpálová et al. 2014), and hence could have been missed in field studies (Sprunger et al. 2018). Furthermore, a minimum of seven years was needed to detect SOC differences between two treatments with at least 15 replicates (Necpálová et al. 2014). In fact, a long-term study on IWG cropping that spanned across 10 years had detected a SOC gain of about 100 g C m−2 y−1 in the top 30 cm of soil, higher than our estimates (Sainju et al. 2023). Our modelled SOC gain appears lower than field observations, possibly caused by the sandy soil texture at our modelled site, which tends to have lower SOC protection capacity.
Perennial grains can improve and maintain soil health, not just SOC
Soil microorganisms play an important role in maintaining soil health as they drive many fundamental processes that sustain the stability and productivity of agro-ecosystems such as the cycling of nutrients, biodegradation of pollutants, C sequestration, and formation of soil aggregates (Chantigny et al. 1997; Singh et al. 2011). The conversion of grassland to winter wheat or IWG cropping decreased microbial biomass in all systems, but the decrease in IWG systems was much reduced compared with the annual (Fig. 5, left column). This is because the continuous living roots of IWG provided a high input of root litter and root exudates throughout the year, supplying readily bioavailable C to soil microbes. In contrast to IWG systems that nearly continuously had LAI > 1 mleaf2 mground−2 beyond the first year, the annual system we modelled lacked substantial crop cover for an extended period (i.e., only about 3 months per year have LAI > 1 mleaf2 mground−2). The C input to soil was thus much reduced. Moreover, in the site we modelled, the annual winter wheat system was left bare after harvest in mid-July to the following sowing in early October. Lack of canopy cover in the summer led to high soil temperatures (> 35 ˚C) in the topsoil layers (not shown). Such temperatures exceeded optimal thermal range for microbial growth in temperate soils (25–30 ˚C; Pietikäinen et al. 2005), further decreasing microbial biomass and widening the difference between the annual and perennial systems.
Although the IWG cropping systems can maintain a much higher microbial biomass than the annual one following a conversion from grassland, the restoration of microbial biomass in annual cropland by introducing perennial IWG is a slow process (Fig. 5, right column), which was also observed in a field study across three sites in Southern Minnesota, USA (Rakkar et al. 2023). After 4 years of IWG following an annual cropping, the heterotrophic microbial biomass had only reached about 90% of the level IWG systems can maintain, and the biomass of fungi had not even started to recover. This slow recovery of microbial community is in line with observations in long-term field monitoring of post-agricultural grassland, where recovery required up to a few decades (Bach et al. 2010; Jangid et al. 2011; Rosenzweig et al. 2016). Fungi have a slower growth rate than bacteria (Sun et al. 2017).
Increased SOC together with increased microbial activity can transform soil physical and hydraulic properties such as porosity, permeability, and hydraulic conductivity. These changes generally increase the soil water retention capacity (Saxton and Rawls 2006; Pelak and Porporato 2019), even though the extent of the effect depends on soil texture and the initial SOC content (Rawls et al. 2003). Hence, the perennial grain cropping, by accumulating more SOC, is expected to have a higher soil water retention than the annual cropping and reduce nutrient leaching (Culman et al. 2013; Jungers et al. 2019; Huddell et al. 2023). In addition to the potential to increase water holding capacity, perennial grains have deep root system (Sprunger et al. 2018; Thorup-Kristensen et al. 2020) and can take up water and nutrients from subsoils. They are thus better candidates for future climates projected to have less frequent but more intense rainfall events (Zhang et al. 2013), as well as warmer conditions leading to higher evaporative demands.
Processes affecting the dynamics of SOC and microbial biomass described above are dependent on the interplay between soil properties, climatic conditions (e.g., solar radiation, temperature, rainfall patterns), and management practices. While the model is of general applicability, our results refer to one specific location and management practices. Some conclusions may not be generalisable. For example, in line with Sprunger et al. (2018), the perennial IWG in our modelled systems was harvested in late August while the annual winter wheat was harvested in mid-July, so that the IWG systems had a longer period with higher LAI and greater root exudation as opposed to the annual system that was left bare for 2–3 months. The decrease of microbial biomass in annual system may not be so drastic if the crops were harvested in autumn rather than summer. Furthermore, we considered a no-till monoculture scenario to isolate the effects of crop life habit from other confounding factors, despite such management does not represent best management practices. Incorporating crop rotation and diversity can reduce the dependence on fertilisers and pesticides and can enhance soil health and grain yields (McDaniel et al. 2014; Sprunger et al. 2020). Although management practices in annual system such as cover crops, crop rotation, and organic farming can increase microbial biomass and activity (Wittwer et al. 2021; Garland et al. 2021) and grain yields (Smith et al. 2023) as well as decrease nutrient leaching (Thapa et al. 2018), whether the resulting effects are comparable with those of perennial cropping is yet to be tested.
Perennial grain crops exhibit a wide variety of characteristics, some of which are still under selection. We tested the role of photosynthetic capacity because it is key for C fixation and hence biomass accumulation and root exudation. Despite a 1.5-fold change in photosynthetic capacity between the two IWGs modelled, photosynthetic capacity does not markedly affect the ability of IWG to reduce SOC loss and sustain soil microorganisms. Irrespective of their photosynthetic capacity, the IWG we tested had grain yields lower than annual wheat (Fig. 2). Selection is in progress to improve yield (Bajgain et al. 2022). Increased C allocation to grains in future varieties might reduce rhizodeposition and the potential for IWG to accumulate SOC. Nevertheless, a model simulation of an IWG cropping with a 10 times higher grain to shoot ratio showed that a higher C allocation to grain decreased only by about 11% the SOC gain and microbial biomass, and they are still very distinct to those of the annual system (results not shown). The selection for shorter IWG plants to minimise lodging may also affect the potential of IWG to reduce SOC loss.
Conclusions
Perennial grain crops have been proposed as an alternative to annual grains to enhance the sustainability of agriculture. Yet, their potential to accumulate SOC and restore soil functioning to conditions relative to pre-agricultural conversion remains largely unquantified. Using a mechanistic plant-soil model, we showed that the perennial wheat candidate intermediate wheatgrass can accumulate more SOC than annual wheat and maintain a SOC level nearer to a multi-species C3 grassland. The perennial intermediate wheatgrass with high photosynthetic capacity resulted in higher soil C accumulation in a warm-summer humid continental climate. Compared with annual cropping, perennial intermediate wheatgrass reduced more than 38% the SOC loss and sustained 2.5 times higher bacterial biomass following a conversion from native grassland. Perennial grain with high photosynthetic capacity could restore the SOC and microbial biomass in cropland previously under annual wheat, although the restoration was slow and did not achieve the same levels as the native grassland. With higher C inputs to soil and microbial biomass, the perennial system had a higher soil respiration than the annual system. However, the high soil respiration in perennial grains was compensated by their high net primary productivity, resulting in a net C sequestration. Our study shows that perennial grain crops can hold their promises in increasing soil C and improving soil health and functions.
Data availability
The datasets generated during and/or analysed during the current study are available from the corresponding author on reasonable request.
Code availability
BRTSim software is freely available at https://sites.google.com/site/thebrtsimproject/home. Input files for running the model described in this study are available from the corresponding author on reasonable request.
References
Abramoff R et al (2018) The millennial model: in search of measurable pools and transformations for modeling soil carbon in the new century. Biogeochemistry 137(1):51–71
Anderson-Teixeira KJ, Davis SC, Masters MD, Delucia EH (2009) Changes in soil organic carbon under biofuel crops. GCB Bioenergy 1(1):75–96
Bach EM, Baer SG, Meyer CK, Six J (2010) Soil texture affects soil microbial and structural recovery during grassland restoration. Soil Biol Biochem 42(12):2182–2191
Bai Y, Cotrufo MF (2022) Grassland soil carbon sequestration: current understanding, challenges, and solutions. Science 377(6606):603–608
Bajgain P, Crain JL, Cattani DJ, Larson SR, Altendorf KR, Anderson JA, ... DeHaan LR (2022) Breeding intermediate wheatgrass for grain production. Plant Breed Rev 46:119–217
Baker JM, Ochsner TE, Venterea RT, Griffis TJ (2007) Tillage and soil carbon sequestration—what do we really know? Agric Ecosyst Environ 118(1–4):1–5
Baronti S, Galassi E, Ugolini F, Miglietta F, Genesio L, Vaccari FP, ... Gazza L (2022) Agronomic and ecophysiological evaluation of an early establishment of perennial wheat lines in Central Italy. Genet Resour Crop Evol 1–15
Benbi DK, Boparai AK, Brar K (2014) Decomposition of particulate organic matter is more sensitive to temperature than the mineral associated organic matter. Soil Biol Biochem 70:183–192
Bengtson P, Barker J, Grayston SJ (2012) Evidence of a strong coupling between root exudation, C and N availability, and stimulated SOM decomposition caused by rhizosphere priming effects. Ecol Evol 2(8):1843–1852
Beniston JW et al (2014) Soil organic carbon dynamics 75 years after land-use change in perennial grassland and annual wheat agricultural systems. Biogeochemistry 120(1):37–49
Bernacchi CJ et al (2001) Improved temperature response functions for models of Rubisco-limited photosynthesis. Plant Cell Environ 24(2):253–259
Bolinder MA, Bolinder MA, Janzen HH, Gregorich EG, Angers DA, VandenBygaart AJ (2007) An approach for estimating net primary productivity and annual carbon inputs to soil for common agricultural crops in Canada. Agric Ecosyst Environ 118(1–4):29–42
Bond-Lamberty BP, Thomson AM (2018) A Global database of soil respiration data, Version 4.0, s.l. ORNL DAAC, Oak Ridge
Brooks RH, Corey AT (1964) Hydraulic properties of porous media. Hydrology Papers 3. Colorado State University, Fort Collins
Campbell GS, Norman JM (1998) An introduction to environmental biophysics. Springer, New York City
Ceriotti G, Tang FHM, Maggi F (2020) Similarities and differences in the sensitivity of soil organic matter (SOM) dynamics to biogeochemical parameters for different vegetation inputs and climates. Stoch Env Res Risk Assess 34(12):2229–2244
Chantigny MH, Chantigny MH, Angers DA, Prévost D, Vézina L-P, Chalifour F-P (1997) Soil aggregation and fungal and bacterial biomass under annual and perennial cropping systems. Soil Sci Soc Am J 61(1):262–267
Conant RT, Paustian K, Elliott ET (2001) Grassland management and conversion into grassland: effects on soil carbon. Ecol Appl 11(2):343–355
Cotrufo MF, Cotrufo MF, Soong JL, Horton AJ, Campbell EE, Haddix M, Wall DH, Parton WJ (2015) Formation of soil organic matter via biochemical and physical pathways of litter mass loss. Nat Geosci 8(10):776–779
Crews TE, Rumsey BE (2017) What agriculture can learn from native ecosystems in building soil organic matter: a review. Sustainability 9(578):1–18
Crews TE, Carton W, Olsson L (2018) Is the future of agriculture perennial? Imperatives and opportunities to reinvent agriculture by shifting from annual monocultures to perennial polycultures. Global Sustain 1(e11):1–18
Culman SW, Culman SW, Snapp SS, Ollenburger M, Basso B, DeHaan LR (2013) Soil and water quality rapidly responds to the perennial grain Kernza wheatgrass. Agron J 105(3):735–744
Dai Y et al (2019) A global high-resolution data set of soil hydraulic and thermal properties for land surface modeling. J Adv Model Earth Syst 11(9):2996–3023
de Oliveira G, Brunsell NA, Sutherlin CE, Crews TE, DeHaan LR (2018) Energy, water and carbon exchange over a perennial Kernza wheatgrass crop. Agric Meteorol 249:120–137
Diamond HJ, Diamond HJ, Karl TR, Palecki MA, Baker CB, Bell JE, Leeper RD, Easterling DR, Lawrimore JH, Meyers TP, Helfert MR, Goodge G, Thorne PW (2013) US climate reference network after one decade of operations: Status and assessment. Bull Am Meteorol Soc 94(4):485–498
Duchene O et al (2020) Introducing perennial grain in grain crops rotation: the role of rooting pattern in soil quality management. Agronomy 10(9):1254
Farquhar GD, von Caemmerer SV, Berry JA (1980) A biochemical model of photosynthetic CO2 assimilation in leaves of C 3 species. Planta 149(1):78–90
Fatichi S, Ivanov VY, Caporali E (2012) A mechanistic ecohydrological model to investigate complex interactions in cold and warm water-controlled environments: 1. Theoretical framework and plot‐scale analysis. J Adv Model Earth Syst 4(2):M05002
Garland G, Garland G, Edlinger A, Banerjee S, Degrune F, García-Palacios P, Pescador DS, Herzog C, Romdhane S, Saghai A, Spor A, Wagg C, Hallin S, Maestre FT, Philippot L, Rillig MC, van der Heijden MGA (2021) Crop cover is more important than rotational diversity for soil multifunctionality and cereal yields in European cropping systems. Nat Food 2(1):28–37
Glover JD, Glover JD, Reganold JP, Bell LW, Borevitz J, Brummer EC, Buckler ES, Cox CM, Cox TS, Crews TE, Culman SW, DeHaan LR, Eriksson D, Gill BS, Holland J, Hu F, Hulke BS, Ibrahim AMH, Jackson W, Jones SS, Murray SC, Paterson AH, Ploschuk E, Sacks EJ, Snapp S, Tao D, Van Tassel DL, Wade LJ, Wyse DL, Xu Y (2010) Increased food and ecosystem security via perennial grains. Science 328(5986):1638–1639
Grayston SJ, Vaughan D, Jones D (1997) Rhizosphere carbon flow in trees, in comparison with annual plants: the importance of root exudation and its impact on microbial activity and nutrient availability. Appl Soil Ecol 5(1):29–56
Guglielmo M, Tang FH, Pasut C, Maggi F (2021) Soil-watergrids, mapping dynamic changes in soil moisture and depth of water table from 1970 to 2014. Sci Data 8(1):1–16
Harris I, Osborn TJ, Jones P, Lister D (2020) Version 4 of the CRU TS monthly high-resolution gridded multivariate climate dataset. Sci Data 7(1):1–18
Hengl T, Hengl T, Mendes de Jesus J, Heuvelink GBM, Ruiperez Gonzalez M, Kilibarda M, Blagotić A, Shangguan W, Wright MN, Geng X, Bauer-Marschallinger B, Guevara MA, Vargas R, MacMillan RA, Batjes NH, Leenaars JGB, Ribeiro E, Wheeler I, Mantel S, Kempen B (2017) SoilGrids250m: global gridded soil information based on machine learning. PLoS ONE 12(2):e0169748
Huddell A et al (2023) Nitrate leaching losses and the fate of 15 N fertilizer in perennial intermediate wheatgrass and annual wheat—a field study. Sci Total Environ 857:159255
Hussain MZ, Robertson GP, Basso B, Hamilton SK (2020) Leaching losses of dissolved organic carbon and nitrogen from agricultural soils in the upper US Midwest. Sci Total Environ 734:139379
Jaikumar NS, Snapp SS, Sharkey TD (2013) Life history and resource acquisition: photosynthetic traits in selected accessions of three perennial cereal species compared with annual wheat and rye. Am J Bot 100(12):2468–2477
Jaikumar NS, Snapp SS, Flore JA, Loescher W (2014) Photosynthetic responses in annual rye, perennial wheat, and perennial rye subjected to modest source: sink ratio changes. Crop Sci 54(1):274–283
Jangid K, Jangid K, Williams MA, Franzluebbers AJ, Schmidt TM, Coleman DC, Whitman WB (2011) Land-use history has a stronger impact on soil microbial community composition than aboveground vegetation and soil properties. Soil Biol Biochem 43(10):2184–2193
Jungers JM et al (2018) Growth, development, and biomass partitioning of the perennial grain crop thinopyrum intermedium. Ann Appl Biol 172(3):346–354
Jungers JM, Jungers JM, DeHaan LH, Mulla DJ, Sheaffer CC, Wyse DL (2019) Reduced nitrate leaching in a perennial grain crop compared to maize in the Upper Midwest, USA. Agr Ecosyst Environ 272:63–73
Kaempf I et al (2016) Potential of temperate agricultural soils for carbon sequestration: a meta-analysis of land-use effects. Sci Total Environ 566:428–435
Kallenbach CM, Frey SD, Grandy AS (2016) Direct evidence for microbial-derived soil organic matter formation and its ecophysiological controls. Nat Commun 7(1):1–10
Keiluweit M et al (2015) Mineral protection of soil carbon counteracted by root exudates. Nat Clim Change 5(6):588–595
Kim K, Kim K, Daly EJ, Flesch TK, Coates TW, Hernandez-Ramirez G (2022) Carbon and water dynamics of a perennial versus an annual grain crop in temperate agroecosystems. Agric for Meteorol 314:108805
Lal R (2016) Soil health and carbon management. Food Energy Secur 5(4):212–222
Lawlor DW (1973) Growth and water absorption of wheat with parts of the roots at different water potentials. New Phytol 72(2):297–305
Ledo A, Ledo A, Smith P, Zerihun A, Whitaker J, Vicente‐Vicente JL, Qin Z, McNamara NP, Zinn YL, Llorente M, Liebig M, Kuhnert M, Dondini M, Don A, Diaz‐Pines E, Datta A, Bakka H, Aguilera E, Hillier J (2020) Changes in soil organic carbon under perennial crops. Glob Change Biol 26(7):4158–4168
Lehmann J, Bossio DA, Kögel-Knabner I, Rillig MC (2020) The concept and future prospects of soil health. Nat Rev Earth Environ 1(10):544–553
Lei C, Abraha M, Chen J, Su YJ (2021) Long-term variability of root production in bioenergy crops from ingrowth core measurements. J Plant Ecol 14(5):757–770
Lhomme JP (1998) Formulation of root water uptake in a multi-layer soil-plant model: does van den Honert’s equation hold? Hydrol Earth Syst Sci 2(1):31–39
Li Z, Li Z, Liu S, Tan Z, Bliss NB, Young CJ, West TO, Ogle SM (2014) Comparing cropland net primary production estimates from inventory, a satellite-based model, and a process-based model in the midwest of the United States. Ecol Model 277:1–12
Luan X, Vico G (2021) Canopy temperature and heat stress are increased by compound high air temperature and water stress and reduced by irrigation–a modeling analysis. Hydrol Earth Syst Sci 25(3):1411–1423
Lugato E, Lavallee JM, Haddix ML, Panagos P, Cotrufo MF (2021) Different climate sensitivity of particulate and mineral-associated soil organic matter. Nat Geosci 14(5):295–300
Maggi F (2019) BRTSim, a general-purpose computational solver for hydrological, biogeochemical, and ecosystem dynamics. Preprint at arXiv https://arxiv.org/abs/1903.07015
Maggi F (2021) BRTSim v5.0 release a, a general-purpose multiphase and multispecies computational solver for biogeochemical reaction-advection-dispersion processes in porous and non-porous media user Manual and Technical Guide. The University of Sydney, Sydney
Manzoni S, Vico G, Porporato A, Katul G (2013) Biological constraints on water transport in the soil–plant–atmosphere system. Adv Water Resour 51:292–304
McDaniel MD, Tiemann LK, Grandy AS (2014) Does agricultural crop diversity enhance soil microbial biomass and organic matter dynamics? A meta-analysis. Ecol Appl 24(3):560–570
Michaelis L, Menten ML (1913) Die kinetik der invertinwirkung. Biochem z 49(333):352
Monod J (1949) The growth of bacterial cultures. Annu Rev Microbiol 3(1):371–394
Necpálová M, Necpalova M, Anex RP, Kravchenko AN, Abendroth LJ, Del Grosso SJ, Dick WA, Helmers MJ, Herzmann D, Lauer JG, Nafziger ED, Sawyer JE, Scharf PC, Strock JS, Villamil MB (2014) What does it take to detect a change in soil carbon stock? A regional comparison of minimum detectable difference and experiment duration in the north central United States. J Soil Water Conserv 69(6):517–531
Oberholzer HR, Leifeld J, Mayer J (2014) Changes in soil carbon and crop yield over 60 years in the Zurich organic fertilization experiment, following land-use change from grassland to cropland. J Plant Nutr Soil Sci 177(5):676–704
Ogle SM et al (2019) Climate and soil characteristics determine where no-till management can store carbon in soils and mitigate greenhouse gas emissions. Sci Rep 9(1):1–8
Pasut C, Pasut C, Tang FHM, Hamilton D, Riley WJ, Maggi F (2021) Spatiotemporal assessment of GHG emissions and nutrient sequestration linked to agronutrient runoff in global wetlands. Glob Biogeochem Cycles 35(4):e2020GB006816
Peixoto L, Peixoto L, Elsgaard L, Rasmussen J, Kuzyakov Y, Banfield CC, Dippold MA, Olesen JE (2020) Decreased rhizodeposition, but increased microbial carbon stabilization with soil depth down to 3.6 m. Soil Biol Biochem 150:108008
Peixoto L, Peixoto L, Olesen JE, Elsgaard L, Enggrob KL, Banfield CC, Dippold MA, Nicolaisen MH, Bak F, Zang H, Dresbøll DB, Thorup-Kristensen K, Rasmussen J (2022) Deep-rooted perennial crops differ in capacity to stabilize C inputs in deep soil layers. Sci Rep 12(1):5952
Pelak N, Porporato A (2019) Dynamic evolution of the soil pore size distribution and its connection to soil management and biogeochemical processes. Adv Water Resour 131:103384
Pietikäinen J, Pettersson M, Bååth E (2005) Comparison of temperature effects on soil respiration and bacterial and fungal growth rates. FEMS Microbiol Ecol 52(1):49–58
Poeplau C et al (2011) Temporal dynamics of soil organic carbon after land-use change in the temperate zone–carbon response functions as a model approach. Glob Change Biol 17(7):2415–2427
Poggio L, Poggio L, de Sousa LM, Batjes NH, Heuvelink GBM, Kempen B, Ribeiro E, Rossiter D (2021) SoilGrids 2.0: producing soil information for the globe with quantified spatial uncertainty. Soil 7(1):217–240
Pugliese JY, Culman SW, Sprunger CD (2019) Harvesting forage of the perennial grain crop kernza (Thinopyrum intermedium) increases root biomass and soil nitrogen cycling. Plant Soil 437(1):241–254
Qin Z, Qin Z, Dunn JB, Kwon H, Mueller S, Wander MM (2016) Soil carbon sequestration and land use change associated with biofuel production: empirical evidence. GCB Bioenergy 8(1):66–80
Raes D, Steduto P, Hsiao TC, Fereres E (2009) Aquacrop—the FAO crop model to simulate yield response to water: II. Main algorithms and software description. Agron J 101(3):438–447
Raiesi F, Beheshti A (2014) Soil C turnover, microbial biomass and respiration, and enzymatic activities following rangeland conversion to wheat–alfalfa cropping in a semi-arid climate. Environ Earth Sci 72:5073–5088
Rakkar M, Jungers JM, Sheaffer C, Bergquist G, Grossman J, Li F, Gutknecht JL (2023) Soil health improvements from using a novel perennial grain during the transition to organic production. Agric Ecosyst Environ 341:108164
Rawls WJ, Rawls WJ, Pachepsky YA, Ritchie JC, Sobecki TM, Bloodworth H (2003) Effect of soil organic carbon on soil water retention. Geoderma 116(1–2):61–76
Richards LA (1931) Capillary conduction of liquids through porous mediums. Physics 1(5):318–333
Riley WJ, Riley WJ, Maggi F, Kleber M, Torn MS, Tang JY, Dwivedi D, Guerry N (2014) Long residence times of rapidly decomposable soil organic matter: application of a multi-phase, multi-component, and vertically resolved model (BAMS1) to soil carbon dynamics. Geosci Model Dev 7(4):1335–1355
Rosenzweig ST, Carson MA, Baer SG, Blair JM (2016) Changes in soil properties, microbial biomass, and fluxes of C and N in soil following post-agricultural grassland restoration. Appl Soil Ecol 100:186–194
Rumpel C, Kögel-Knabner I (2011) Deep soil organic matter—a key but poorly understood component of terrestrial C cycle. Plant Soil 338(1):143–158
Sainju UM, Allen BL (2023) Carbon footprint of perennial bioenergy crop production receiving various nitrogen fertilization rates. Sci Total Environ 861:160663
Sainju UM, Allen BL, Lenssen AW (2023) Soil total carbon and nitrogen under long-term perennial bioenergy crops receiving various nitrogen fertilization rates. Agron J 115:2216–2226. https://doi.org/10.1002/agj2.21422
Saxton KE, Rawls WJ (2006) Soil water characteristic estimates by texture and organic matter for hydrologic solutions. Soil Sci Soc Am J 70(5):1569–1578
Schenk HJ, Jackson RB (2002) The global biogeography of roots. Ecol Monogr 72(3):311–328
Singh JS, Pandey VC, Singh DP (2011) Efficient soil microorganisms: a new dimension for sustainable agriculture and environmental development. Agric Ecosyst Environ 140(3–4):339–353
Six JΑΕΤ, Elliott ET, Paustian K (2000) Soil macroaggregate turnover and microaggregate formation: a mechanism for C sequestration under no-tillage agriculture. Soil Biol Biochem 32(14):2099–2103
Smith ME, Vico G, Costa A, Bowles T, Gaudin AC, Hallin S, Bommarco R (2023) Increasing crop rotational diversity can enhance cereal yields. Commun Earth Environ 4(1):89
Sprunger CD, Culman SW, Robertson GP, Snapp SS (2018) Perennial grain on a Midwest Alfisol shows no sign of early soil carbon gain. Renew Agric Food Syst 33(4):360–372
Sprunger CD, Sprunger CD, Culman SW, Peralta AL, DuPont ST, Lennon JT, Snapp SS (2019) Perennial grain crop roots and nitrogen management shape soil food webs and soil carbon dynamics. Soil Biol Biochem 137:107573
Sprunger CD, Martin T, Mann M (2020) Systems with greater perenniality and crop diversity enhance soil biological health. Agric Environ Lett 5(1):e20030
Sun S et al (2017) Soil bacterial and fungal communities show distinct recovery patterns during forest ecosystem restoration. Appl Environ Microb 83(14):e00966–e00917
Sutherlin CE et al (2019) Contrasting physiological and environmental controls of evapotranspiration over Kernza perennial crop, annual crops, and C4 and mixed C3/C4 grasslands. Sustainability 11(6):1640
Tang SM et al (2022) Effects of grassland converted to cropland on soil microbial biomass and community from agro-pastoral ecotone in Northern China. Grassl Sci 68(1):36–43
Tautges NE, Jungers JM, DeHaan LR, Wyse DL, Sheaffer CC (2018) Maintaining grain yields of the perennial cereal intermediate wheatgrass in monoculture v. bi-culture with alfalfa in the upper midwestern USA. J Agric Sci 156(6):758–773
Taylor K, Samaddar S, Schmidt R, Lundy M, Scow K (2023) Soil carbon storage and compositional responses of soil microbial communities under perennial grain IWG vs. annual wheat. Soil Biol Biochem 184:109111
Thapa R, Mirsky SB, Tully KL (2018) Cover crops reduce nitrate leaching in agroecosystems: a global meta-analysis. J Environ Qual 47(6):1400–1411
Thorup-Kristensen K, Thorup-Kristensen K, Halberg N, Nicolaisen M, Olesen JE, Crews TE, Hinsinger P, Kirkegaard J, Pierret A, Dresbøll DB (2020) Digging deeper for agricultural resources, the value of deep rooting. Trends Plant Sci 25(4):406–417
Tinsley SG (2012) An evaluation of perennial wheat and intermediate wheatgrass as dual-purpose, forage-grain crops under organic management. Michigan State University, Michigan
Tuzet A, Perrier A, Leuning R (2003) A coupled model of stomatal conductance, photosynthesis and transpiration. Plant Cell Environ 26(7):1097–1116
USDA (2021) Metadata for Winter/Spring Wheat Growth Stage Models [Online] Available at: https://ipad.fas.usda.gov/cropexplorer/description.aspx?legendid=313®ionid=na. Accessed 25 Sept 2021
van der Pol LK, van der Pol LK, Nester B, Schlautman B, Crews TE, Cotrufo MF (2022) Perennial grain Kernza® fields have higher particulate organic carbon at depth than annual grain fields. Can J Soil Sci 102(4):1005–1009
Vico G, Porporato A (2008) Modelling C 3 and C 4 photosynthesis under water-stressed conditions. Plant Soil 313(1):187–203
Vico G, Brunsell NA (2018) Tradeoffs between water requirements and yield stability in annual vs. perennial crops. Adv Water Resour 112:189–202
Vico G et al (2013) A perspective on optimal leaf stomatal conductance under CO2 and light co-limitations. Agric for Meteorol 182:191–199
Vico G et al (2016) Trade-offs between seed output and life span–a quantitative comparison of traits between annual and perennial congeneric species. New Phytol 209(1):104–114
Vico G, Tang FHM, Brunsell NA, Crews TE, Katul GG (2023) Photosynthetic capacity, canopy size and rooting depth mediate response to heat and water stress of annual and perennial grain crops. Agric For Meteorol 341:109666
Wagoner P, Schaeffer JR (1990) Perennial grain development: past efforts and potential for the future. CRC Crit Rev Plant Sci 9(5):381–408
Warren CR (2016) Simultaneous efflux and uptake of metabolites by roots of wheat. Plant Soil 406(1):359–374
Webber H et al (2016) Simulating canopy temperature for modelling heat stress in cereals. Environ Model Softw 77:143–155
Wiesner S, Duff AJ, Niemann K, Desai AR, Crews TE, Risso VP, Stoy PC, Wiesner S, Duff AJ, Niemann K, Desai AR, Crews TE, Risso VP, Riday H, Stoy PC (2022) Growing season carbon dynamics differ in intermediate wheatgrass monoculture versus biculture with red clover. Agric for Meteorol 323:109062
Willson TC, Paul EA, Harwood RR (2001) Biologically active soil organic matter fractions in sustainable cropping systems. Appl Soil Ecol 16(1):63–76
Wittwer RA, Wittwer RA, Bender SF, Hartman K, Hydbom S, Lima RAA, Loaiza V, Nemecek T, Oehl F, Olsson PA, Petchey O, Prechsl UE, Schlaeppi K, Scholten T, Seitz S, Six J, van der Heijden MGA (2021) Organic and conservation agriculture promote ecosystem multifunctionality. Sci Adv 7(34):eabg6995
Woodmansee RG (1978) Additions and losses of nitrogen in grassland ecosystems. Bioscience 28(7):448–453
Zhang X, Zhang X, Wan H, Zwiers FW, Hegerl GC, Min S-K (2013) Attributing intensification of precipitation extremes to human influence. Geophys Res Lett 40(19):5252–5257
Zhang Y, Schaap MG, Zha Y (2018) A high-resolution global map of soil hydraulic properties produced by a hierarchical parameterization of a physically based water retention model. Water Resour Res 54(12):9774–9790
Acknowledgements
This research was funded by the Swedish Research Council for Sustainable Development FORMAS grant 2018 − 00646 to GV, TEC and NAB. The model simulations in this study were enabled by resources in project SNIC 2021/22–758 provided by the Swedish National Infrastructure for Computing (SNIC) at UPPMAX, partially funded by the Swedish Research Council through grant agreement no. 2018–05973. Additional funding to TEC and NAB was provided through the United States Department of Agriculture (Award #2020-68012-31934). The authors acknowledge Federico Maggi for providing technical assistance on the use of the BRTSim software, which is freely available at https://sites.google.com/site/thebrtsimproject/home. The authors acknowledge Stefano Manzoni for providing constructive feedbacks on the manuscript.
Funding
Open access funding provided by Swedish University of Agricultural Sciences. This research was funded by the Swedish Research Council for Sustainable Development FORMAS grant 2018-00646 to GV, TEC and NAB. Additional funding to TEC and NAB was provided through the United States Department of Agriculture (Award #2020-68012-31934).
Author information
Authors and Affiliations
Contributions
All authors contributed to the study conception and design. Model development was performed by FHMT and GV. The first draft of the manuscript was written by FHMT and all authors commented on previous versions of the manuscript. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors have no relevant financial or non-financial interests to disclose.
Additional information
Responsible Editor: Kenny Png.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Tang, F.H.M., Crews, T.E., Brunsell, N.A. et al. Perennial intermediate wheatgrass accumulates more soil organic carbon than annual winter wheat – a model assessment. Plant Soil 494, 509–528 (2024). https://doi.org/10.1007/s11104-023-06298-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-023-06298-8