
Abstract
Aim
Balanced crop nutrition is key to improve nutrient use efficiency and reduce environmental impact of farming systems. We developed and tested a dynamic model to predict the uptake of P and K in long-term experiments to better understand how changes in soil nutrient pools affect nutrient availability in crop rotations.
Methods
Our RC-KP model includes labile and stable pools for P and K, with separate labile pools for placed P and organic fertilizers including farm yard manure (FYM). Pool sizes and crop-specific relative uptake rates determined potential uptake. Actual crop uptake from labile pools was based on concepts developed by Janssen et al. (Geoderma 46:299-318, 1990). The model was calibrated on three long-term experiments from Kenia (Siaya), Germany (Hanninghof) and the United Kingdom (Broadbalk) to estimate parameter values for crop-specific relative uptake rates and site-specific relative transfer rates.
Results
The model described N, P and K uptake accurately with a Nash-Sutcliff modelling efficiency of 0.6–0.9 and root mean squared errors of 2.6–3.4 kg P ha−1 and 14–20 kg K ha−1. Excluding organic labile pools did not affect model accuracy in Broadbalk in contrast to Hanninghof where Mg deficiencies affected crop uptakes in treatments without Mg or FYM.
Conclusions
This relatively simple model provides a novel approach to accurately estimate N, P and K uptake and explore short- and long-term effects of fertilizers in crop rotations. Interactions between limiting nutrients affecting actual nutrient uptake were captured well, providing new options to include N, P and K limitations in crop growth models.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Reduction of emissions to the environment including greenhouse gases and ammonia to air, nitrate and (ortho)phosphates to ground and surfacewaters are needed in agriculture while increasing or maintaining high yields to feed the growing world population. Improvement of nutrient use efficiency (NUE) is key in the context of sustainable and ecological intensification (Struik and Kuyper 2017) and circular systems (Papangelou and Mathijs 2021). Yield responses to fertilizer, expressed as agronomic efficiency, are notoriously variable between fields and seasons (Ichami et al. 2019; Kihara et al. 2016), caused by mismatches between nutrient demand and supply (Schut and Giller 2020). Hence, a better understanding of the dynamics in soil nutrient pools is needed to prevent mining and build-up of large soil pools, determined by the balance between plant uptake and short- and long-term effects of applied fertilizers.
Uptake of P and K is governed by availability of N. For situations without P and K deficiency, the first year recovery of applied N is fairly well understood. The fraction that is applied but not taken up by the crop in the first year may be available in later years, depending on crop residue management. In long-term experiments with monoculture maize, barley and winter wheat where residues were retained, this long-term N recovery fraction was 0.24 larger than the first year N recovery fraction due to on-field recycling (Vonk et al. 2022). These findings are congruent with values reported by Petersen et al. (2012) and values observed in N15 experiments (Petersen et al. 2010). There are also strong effects of crop rotation on N availability via mineralisation of N rich residues, especially important after legumes. Quantitative knowledge about interactions between macronutrients and their effect on uptake and long-term soil supply is limited. Models may help to understand these dynamics and allow to further fine-tune rotational fertilization strategies.
Assessments of the amounts of plant available P and K remains challenging and are strongly affected by errors in lab and field sampling procedures (Schut and Giller 2020). Simple relationships between crop responses and soil P tests are very noisy (Steinfurth et al. 2022) and may at best indicate whether there is or is not a reasonable chance of a yield response to application (Cate Jr and Nelson 1971). Soil tests for exchangeable K contents are also poor predictors for crop uptake. The strong legacy effect of fertilizer (van der Bom et al. 2017) and manure use (Njoroge et al. 2019) are poorly captured by routine soil tests. Non-exchangeable and subsoil pools also need to be taken into account (Damar et al. 2020). N fertilization increases crop demand for other nutrients and the ability to take up these nutrients (Pasley et al. 2019). This increased uptake capacity from the soil is likely due to more extensive roots that can explore much larger soil volumes in response to nitrate in the soil (Lyzenga et al. 2023). Assessments of soil nutrient availability are further complicated by soil-specific sorption characteristics (Nair 1992) and exchange dynamics between pools. Large amounts of P can be found in soil organic matter, Ca-, Fe- or Al-phosphates that determine the concentration of orthophosphate in soil water, depending on the pH of the soil (Penn and Camberato 2019). Actively weathering soils can release P from apatite and K from K bearing minerals such as K-Feldspar or micas. K is also contained in 2:1 clay. Sandy and strongly weathered, kaolinitic clay soils have limited capacity to adsorb K and are prone to leaching, contrasting with 2:1 clay soils that can contain large amounts of K and may even absorb and incorporate K in the clay lattice. On soils with large P availability, uptake is limited by crop demand that depends on growth potential and other nutrient limitations. On soils where P availability is low, P bioavailability is to a large extent determined by exchanges between the labile and stable pools. The interaction between these two mineral pools and long-term recovery of applied P fertilizer is fairly well understood (Janssen et al. 1987; Wolf et al. 1987).
Current models that account for soil P dynamics typically include a labile and a stable P pool (Wolf et al. 1987) combined with one or more organic labile and stable P pools (Das et al. 2022; Dzotsi et al. 2010). Models for K also include labile and stable pools (Scanlan et al. 2015), although Nguyen (2003) included an additional intermediate pool to model K in paddy rice systems. Current crop growth models that account for N and P limitations explicitly account for ad- and desorption characteristics to estimate the amount of P (as H2PO4−, HPO42− or even PO43− in very small amounts) and K+ in soil solution.
Dynamic crop growth models typically use a very detailed approach to soil P dynamics in mineral and organic pools (Das et al. 2019; Lewis and McGechan 2002; Muntwyler et al. 2023). Organic N and P pools are directly linked to soil carbon, although C:P ratios vary strongly when P is deficient (Yang and Post 2011). Das et al. (2022) defined organic, labile and stable pools, where the stable pool was considered to be 10 times larger than the labile pool. The organic P pool is typically large and includes, amongst others, microbial, active and passive pools (Lewis and McGechan 2002). However, these conceptual pools for organic and mineral P are simplifications of the dynamics in the soil. The exchange rates between soil pools are soil specific and depend on sorption capacity, determined by clay minerology, pH and presence of Fe and Al in the soil (Penn and Camberato 2019). The plant available amounts of P and K are very small compared to the size of the labile and stable pools (Das et al. 2019), and its dynamics were poorly captured by the DayCent model (Muntwyler et al. 2023). Detailed (Langmuir) equations, that describe P adsorption and desorption characteristics of the soil (Lewis and McGechan 2002), can be used to estimate plant available P when assuming that labile pools are in dynamic equilibrium with P in soil solution. However, current models fail to properly capture interactions between plant roots, mycorrhiza and rhizosphere conditions affecting uptake of poorly mobile nutrients in soils with limited fertility (Hinsinger et al. 2011).
Although these detailed dynamic models are highly valuable for further scientific understanding, they also require many crop- and soil specific parameters and are challenging to initialize properly with e.g. soil analysis. However, a detailed mechanistic model may not be needed to better understand carry-over effects and long-term changes in soil P pools (Wolf et al. 1987). In the RC-P model, annual uptake from the labile pool is estimated using a single relative uptake rate that is derived from the additional uptake of fertilized treatments (Wolf et al. 1987). This relative uptake rate is crop-specific and refers to the proportional uptake rate in relation to the labile P pool. The RC-P model does not take effects of crop demand and interactions with the availability of other nutrients on P uptake into account.
The uptake of and yield response to one macronutrient is strongly determined by the soil availability and fertilizer supply of other nutrients. The QUEFTS model (Janssen et al. 1990) has been used to describe and predict relationships between N, P and K fertilizer supply and crop yield. QUEFTS is one of the most frequently used models for fertilizer recommendations and has been calibrated for a wide range of crops (Janssen et al. 1990; Kumar et al. 2018; Setiyono et al. 2010; Xu et al. 2015). The principles behind the relationships between N, P and K uptake and yield are robust and are also underpinning the nutrient equivalent concepts (Janssen 1998, 2017). These concepts allowed us to balance N, P and K uptakes of Cassava on a daily basis (Adiele et al. 2022). However, the original QUEFTS model does not account for long-term effects of fertilizer applications or carry-over effects of previous crops in the rotation.
Our objective was to develop a relatively simple model that allows to describe the effects of organic and inorganic fertilizer treatments and crop rotations on P and K uptake in selected long running experiments with both P and K limitations, while masking the effect of year-to-year variation in growth conditions. We are particularly interested in situations where N, P and K are limited, typical for many tropical farming systems at present and temperate systems in future when N and P supply is likely further restricted to reduce environmental impacts. To this end, we evaluate whether annual plant P and K uptake can be estimated with an extended dynamic RC-P model including organic P and K pools to determine potentially plant available N, P and K in the soil and selected QUEFTS components to determine actual uptake of N, P and K by the crop.
Materials and methods
The model
The RC-KP model described here is an extension of the RC-P model by Wolf et al. (1987) combined with only step II calculations from QUEFTS. The original QUEFTS model includes four calculation steps (Janssen et al. 1990; Sattari et al. 2014). Step I determines potential soil supply using empirical equations describing relationships between soil test values for Olsen P, exchangeable K, pH and soil organic carbon contents and plant available N, P and K in the soil. Step I calculations introduce a lot of uncertainty due to the relatively poor repeatability of soil tests (Schut and Giller 2020); the differences between the equations used (Shehu et al. 2019; Tabi et al. 2008); and uncertainty in the recovery parameters for fertilizer N, P and K. The dynamic labile pools described with RC-KP replace these step I calculations in QUEFTS. Step II calculations determine actual uptake from potential soil supply which are more robust. The step III and IV calculations determine possible yield ranges and yield, these were not included as our objective was to first model N, P and K uptakes, not yield.
The RC-KP model is a dynamic state-rate model that quantifies the long-term changes in soil pools of P and K and associated nutrient uptakes by crops with and without organic or synthetic fertilizer applications. Nutrient uptake refers in this work to nutrients that end up in grains or straw/stover of cereals or potato tubers and is equivalent to nutrient offtake and removal from the field. The model was developed to improve quantitative insights in nutrient transfers between soil pools, and the potential and actual uptake of N, P and K and associated long-term recovery of fertilizers in crop rotations on a seasonal basis. The model does not include detailed processes that determine the dynamics of nutrient pools, nutrient transfers and uptake within a growing season. The model was implemented in R software (R Core Team 2021), version 4.1.0. The ordinary differential equations were solved with the powerful ‘deSolve’ package (Soetaert et al. 2010), using the “lsoda” integration method to handle discrete events. The integration procedure uses a variable time-step and can produce output for any time-step desired.
The RC-KP model includes labile and stable pools for P and K (Fig. 1). A crop-specific proportion of P and K in the labile pools can be taken up by the crop. There is a continuous exchange between labile and stable pools to attain equilibrium. This reflects the processes in the soil with a continuous exchange between P and K in solution that can be taken up, the soil available P or exchangeable K and organic and mineral P and mineral K.

Forrester diagram of the model. Solid lines represent nutrient flows, dashed lines information flows. States are in rectangular shapes, auxiliary variables that determine the available (Av.) amounts in round shapes. Parameters are given above short lines for relative transfer rates (r), fraction of soluble fertilizer (fsol) and relative uptake rates (rur). Red lines show input in discrete events
Labile P includes three pools to capture differences between broadcast and placed P and between organic and mineral P fertilizer. Uptake rates for placed P are much higher than for broadcast P, especially when application rates are low or when soils have a small soluble P pool (van der Eijk et al. 2006). K is much more mobile in the soil than P, hence effects of placement are expected to be small and no placed pool was added for K. Organic P fertilizers are expected to have different dynamics and recovery rates than mineral P fertilizers (Blake et al. 2000), hence the relative transfer rate (r) for P in the farm yard manure (FYM) pool (rPFYM) is expected to differ from the labile pool (rPlabile). In contrast to P (and N), K is not part of soil organic matter and it is expected that rKFYM does not differ from rKlabile.
The stable P pool represents various forms of P in the soil, ranging from apatite, di-hydrogen phosphate, organic P and P in Fe, Al and Ca phosphates. The actual form present in the soil strongly depends on pH and other soil characteristics. The labile pool reflects ortho- and hydrogen phosphates that plants may take up proportionally. The cation K+ behaves very differently as it is typically adsorbed to the cation exchange complex. However, K is also part of the 2:1 clay mineral lattice (Simonsson et al. 2009) and is incorporated in and released from these clay minerals in large quantities as function of long-term K balances (Andersson et al. 2007; Simonsson et al. 2007). The labile pool represents the K that can be taken up proportionally, while the stable K pool reflects K that is either incorporated in these 2:1 clay mineral lattices or very strongly bound to the CEC. Soil pH effects on K in 2:1 clay mineral lattices (Simonsson et al. 2009) were not explicitly accounted for.
Depending on the type of fertilizer and the associated solubility, applied mineral P fertilizer contributes to both labile and stable pools. Mineral K fertilizer is typically applied as muriate of potassium (KCl) which dissolves easily or as animal manures where K is part of the liquid fraction. Therefore, in the RC-KP the applied mineral K fertilizer adds to the labile pool only. The labile component of applied FYM adds to the PFYM and KFYM pools, while the organically bound P adds to the Pstable pool.
Differential equations in the model
The pool sizes are expressed in kg ha−1. The associated rates for labile and stable pools (kg ha−1 y−1) are computed as:
Where \({rP}_{labile}\) and \({rK}_{labile}\) reflect the relative transfer rates (r, y−1) from the labile to stable pool and \({rP}_{stable}\) and \({rK}_{stable}\) the relative transfer rates from stable to labile pools of P and K. The uptake of P from the labile (Puplabile), placed (Pupplaced) and FYM (PupFYM) pools are actual uptakes. These actual uptakes are based on the potential uptake of N, P and K but then adjusted for an imbalanced N, P and K supply using QUEFTS functions. This accounts for the effect of crop demand limitation when not all nutrients are sufficiently available in the soil. The potential uptakes are computed with crop-specific relative uptake rates (rur) for each pool (with a unit y−1).
Potential crop uptakes of N, P and K
The differences in uptake rate between placed P and broadcast P, observed on a wide range of soils when P is strongly limiting yields (de Wit 1953; van der Eijk et al. 2006), decrease with P application level. At higher application rates of about 130 kg P ha−1, differences between broadcast and placed P fertilizer in plant P uptakes disappear (van der Eijk et al. 2006). This application rate corresponds with a maize yield of about 8000 kg ha−1 and an estimated uptake of 23.3 kg P ha−1 when assuming balanced nutrition (Schut and Giller 2020). We assumed that the observed differences between placed and broadcast P were a function of the applied amounts of P fertilizer and the size of the Plabile pool. Hence, a balanced labile pool size (Plabile balance) was determined when potential P uptake was equal to 23.3 kg of P ha−1 y−1 for maize:
There is no difference expected between placed and broadcast applications when the Plabile exceeds this balanced amount. The ratio between Plabile and Plabile balance was used to correct the relative uptake rate of placed P:
This assured that the relative uptake rates from Plabile and Pplaced pools have the same value when the labile P pool exceeds the balanced amount.
The actual uptake is computed using the “step II” equations of the QUEFTS model (Janssen et al. 1990). The potential nutrient supply represents the amount that can be taken up when all other nutrients are available in excess. The potential soil nutrient supply for N (SN), P (SP) and K (SK) are computed per year (y) as:
Where the relative uptake rates (rur, y−1) and N recovery fractions are crop (c) specific. The N uptake from either control or PK treatments (NuptakeREF) is used as reference to estimate how much N can be taken up from the soil when no fertilization was applied. This amount includes N from mineralisation of organic matter, atmospheric deposition and biological N fixation.
The RC-KP model does not include a module to account for mineralisation of soil organic matter (SOM) and nitrogen deposition. However, soil organic matter increases as function of organic inputs and soil N pools will build up under NPK or FYM fertilization (but not without N applications). Parts of the applied N (and P) will become a component of organic pools in the soil that mineralize over time (ten Berge et al. 2019; Vonk et al. 2022), depending on residue management, P and K supply affecting crop growth. To simplify, we estimated the soil N supply from the N uptake of the PK treatment when fertilizers were used or control treatment when not. Any additional soil N supply of other treatments due to internal N recycling was captured by a change in the recovery fraction of applied N over time (see below).
When N uptakes were unknown, these were estimated from a combination of the crop specific rN parameter, determining the minimum N uptake required before yield is formed (Sattari et al. 2014), and the measured yields:
This rN value was set at a value of 5 kg N ha−1 for grain and potato crops (Sattari et al. 2014) and at 0.5 kg N ha−1 for silage maize. In the QUEFTS concept the physiological N use efficiency (PhEN, kg yield kg−1 N uptake), as determined by the relationship between crop nutrient uptake and yield, is bound by the maximum dilution (d) and accumulation (a) of N.
Under balanced nutrition the dilution factor df will be 0.5. At maximum dilution when df is 1.0, N concentration in harvested products and above-ground residues are at their lowest possible value, resulting in the highest possible PhEN value. This only occurs when N is strongly limiting with P and K in excess supply. N accumulates in products and/or residues when P and K are limiting, when df is between 0 and 0.5. The df value was set to 0.85 and 0.99 for control and PK treatments respectively as the N dilution is stronger when P and K are in ample supply. The amounts of SOM and mineralisation of N differ between treatments in long-term experiments. Some of the applied N becomes part of SOM and is released in later years, while SOM for control and PK treatments is expected to decline due to N limitations on crop growth with decreasing amounts of crop residue to compensate SOM mineralisation. This extra N mineralisation from soil organic matter in N fertilized treatments was captured by assessing a long-term recovery of applied N. The recovery values for N from synthetic fertilizer (Nrec and NrecFYM) and manure (FYMNrec) varied per crop and per year and were estimated from the trial data.
The long-term N recovery of applied mineral N was expected to be slightly lower in treatments with FYM when compared to treatments without FYM due to the larger total N supply and slightly larger losses and were calculated as:
The recovery from N in FYM was determined as:
N uptake in fertilized treatments is expected to increase over time, while N uptake in PK treatments is expected to decrease. Hence, these computed recovery values reflect long-term recoveries that increase over time and are typically much greater than the recovery value in the first year (Vonk et al. 2022).
Actual crop uptakes of N, P and K
As indicated above, QUEFTS functions were used to determine the actual uptake from the given potential SN, SP and SK supply. Following the QUEFTS approach, the potential supply of each nutrient is compared in pairs using crop-specific values for accumulation (a) and dilution (d) (Janssen et al. 1990; Sattari et al. 2014). These values for aN, dN, aP, dP, aK and dK were derived from the experimental data that was available and expressed per kg dry matter (DM), taking the maximum ratios between uptake of P to N and K to N into account when estimating dP and dK. When only fresh crop yields were measured, dry matter contents of 35% for potato and 88% for cereal crops were assumed.
As shown above in the equation for SP, P can be taken up from three pools. Actual uptakes as calculated with QUEFTS are lower than potential uptake. The proportion of the potential P supply that is taken up by the crop is calculated as:
We assumed that actual uptake was proportional to potential supply for all pools:
For K, a similar approach was followed, although it can be argued that K from animal manure is fully mineral and part of the liquid fraction. However, we observed that the first-year recovery rates of K and P when applied as animal manures differ from those of mineral fertilizers, even when application rates are well below optimum. Hence, we expect that the crops differ in relative uptake rates and that these rates differ between labile, placed and FYM pools. Defining separate pools for each type of application allowed to evaluate these differences when FYM and mineral fertilizers are combined.
Fertilizer applications
Fertilizer applications are modelled as discrete events. At an event, values of states are changed instantly. Fertilizers were applied in one time-step, at the indicated application moments of FYM and artificial fertilizers. When P is added as triple super phosphate, 80% was considered labile (Wolf et al. 1987). Most P in animal manures is labile as orthophosphate and up to 88% is water soluble (Li et al. 2014). Hence, a proportion of 0.88 was considered labile and assigned to the PFYM pool. The labile P fraction of mineral P fertilizer was added to the Plabile or Pplaced when placed. The remaining stable fractions of mineral and organic fertilizers were added to the stable P pool. The applied K was fully assigned to the Klabile or KFYM pools.
Experimental data
Experimental data was derived from three long-term experiments. The first experiment was conducted in Siaya, Kenia. In this region, two crops per year are grown. The on-farm trial in Siaya was a nutrient omission trial, replicated on 23 farms that ran for 11 seasons, starting at the long rainy season in 2013 up to and including the long rainy season in 2018. A standard maize crop was grown in all seasons. This trial allows the study of crop uptake in soils with strongly depleted or imbalanced soil nutrient pools (Njoroge et al. 2017, 2019). The fields on these farms varied strongly in soil fertility at the start of the trial. The trial included two distinct phases. Treatments in phase I included a control, PK, NK, NP and NPK. In phase II, 16 farms remained. On 4 farms, PK, NP, NK and NPK were superimposed on every plot creating 20 plots per field. On the other 12 farms NPK was applied on all plots. More details on this experiment can be found in Njoroge (2019). Nutrient uptakes in grain and stover were available for the long rainy season in 2016 and 2018 only. In total, there were recorded uptakes in 105 plots in 2016. In 2018, uptakes were recorded on 9 fields only, including 5 fields with only NPK applied (25 records), and 4 fields with treatments with superimposed NK, NP, PK and NPK resulting in 70 records. Some crops did not yield: unbalanced treatments superimposed on strongly depleted control plots resulted in failed crops were uptakes could not be determined.
The Hanninghof experiment has been running since 1958 with 14 treatments, including a control, N, PK, NK, NP, NPK, NPKMg treatments, in combination with and without manure (FYM) for all treatments. The crop rotation was potatoes, winter rye and oats in this specific order during the studied period from 1961 to 2009. A total of 25 t ha-1 FYM was applied as pig manure to potatoes only about 10 days before planting, adding 175, 73 and 149 kg N, P and K ha-1 respectively. More details of this experiment can be found in Jate (2010; 2012). For each year, 14 uptake measurements of P and K in grains, straw and potatoes were available. In later years in the experiment, NPKMg yields were higher than NPK yields, suggesting that Mg deficiency was present in some treatments, more so for potato than for oats and winter rye (Jate 2012; Jate and Lammel 2022).
The Broadbalk wheat experiment is well known and has been running since 1843 (Johnston and Poulton 2018). The fertilizer experiment started in 1852 and includes a range of treatments including a control, N, NP, NK, and NPK (in strips) with or without Mg (Rothamsted Research 2018). P is applied as triple superphosphate, K as potassium sulphate and N as ammonium nitrate, the amounts that were applied were adapted during the course of the experiment. In autumn, a total of 35 t FYM ha-1 y-1 was applied annually as cattle manure in separate strips, with or without NPK. This FYM application added on average 250, 48 and 335 kg N, P and K ha-1 y-1 respectively. Winter wheat has been grown as monocrop until 1968 (Glendining et al. 2021). In 1968 a crop rotation including potatoes, oats, beans, winter wheat and silage maize started. The original trial was split into 10 sections where treatment strips were maintained (Glendining et al. 2021). Five sections remained in continuous winter wheat (0, 1, 6, 8 and 9), the other five in an arable rotation including winter wheat (Sects. 2–5 and 7). The NK treatment was only available for Sects. 0 and 1 with continuous wheat. Data from the years 1968 up to and including 2015 were downloaded from the ERA portal (www.era.rothamsted.ac.uk), including all available sections and all strips. Data of yields and N, P and K contents of harvested products (including potatoes, grain and stover) from Sect. 1 (only 1968 and 1969 to initialize the model), 2–5 and 7 were used to compute crop uptakes and account for annual variability (see below). Note that from 1976 to 1985 only N contents were measured, P and K contents were not available. Crop rotations and estimated uptakes from Sect. 4 were used to evaluate the RC-KP model including strips 1 (FYMNPK), 2.2 (FYM), 3 (Control), 5 (PKMg), 6 (N1PKMg), 7 (N2PKMg), 10 (N), 11 (NP), 13 (NPK), 14 (N4PKMg), 16 (N5PKMg) and 20 (NKMg) (Rothamsted Research 2018). The names for the treatments in parenthesis are used for illustration purposes in this manuscript as amounts of fertilizer applications varied during the course of the experiment. This choice of Sect. 4 was not very important: the differences between the sections with a rotation (2–5 and 7) are small as treatments had the same inputs, only rotations shifted one year. This Sect. 4 included various rotations: wheat : potato : beans until 1978; potato : winter wheat : fallow until 1986; and three years winter wheat:: fallow : potato until 1998. From 1999 onwards, the rotation became oats : silage maize : winter wheat with initially three years of continuous wheat but later only one season of wheat. For fallow years, fertilizers were applied but no crops were grown. Oats did not receive N between 1996 and 2017.
Accounting for annual variability
Nutrient uptakes and yields strongly vary over time. Here, we were interested in the changes in uptake over a longer period of time to study the effect of pool sizes and their interactions on uptakes of crops in a rotation. Hence linear regression models were used to remove annual variability in yields, N recovery and P and K uptakes.
The following explanatory factors and interactions were included in the fitted linear regression models: fertilizer regime × treatment + crop × (year + treatment) for N, P and K uptake; and fertilizer regime + crop × year for Nrec, NrecFYM and FYMNrec. Fertilizer regime reflects a factor to account for the changes in the amounts of fertilizer that were applied to the crops over the course of the experiment. Interactions were only included when significant.
These regression models were evaluated with an analysis of variance, using standard functions in R and RStudio software. The significance of factors was evaluated using an F-test, the effects were considered significant when p < 0.05. These model-predicted values were used as reference data for calibration and evaluation purposes.
Model initialization
The calculations of the initial sizes of the P and K pools are the same, hence only calculations for P are shown here. The initial Plabile pool size can be calculated from the P uptake in the NK treatment in the first year of the experiment.
The initial size of the stable pool is determined by assuming that the transfers from stable to labile pool equals P uptake and transfer from labile to the stable pool. The transfer from stable to labile pools is considered to be in balance and fully compensated crop uptake and the transfer from labile to stable pools (Wolf et al. 1987). As uptake and labile pools sizes are known, these transfer rates and hence pool sizes can be calculated:
This assumption of balance, where transfer from stable to labile pool matches the sum of crop uptake and transfer from labile to stabile pool, was not valid for the experimental trials. For Hanninghof, K uptakes in the first rotation were much larger than in second and later rotations. For Siaya and Broadbalk, the first year with measured nutrient uptakes (yf) was not the first year (y0) in the experiment. Multiple years of imbalanced fertilization were expected to result in imbalanced pools, where a net transfer from e.g. stable to labile P pools can be expected for the NK treatment. These estimates of the pool sizes in yf were therefore corrected. The pool sizes in yf were used to calculate the pool sizes after n years of NK treatment for P pools or NP treatment for K pools for each field, where n refers to the number of years between y0 and yf. The pools in yf were then corrected accordingly, shown here for P only:
The ratios reflect the proportional change in pool sizes after n years of NK fertilization for this example.
In the Siaya experiment, uptakes were only available for the 7th season and the initial pool sizes had to be estimated. This was done using a combination of backcasting and pool correction. The known uptakes in the NK and NP treatments at the end of phase I, in 2016, were used to determine pool sizes. These pool sizes were set as initial values and the model was run backwards, thus subtracting the rates of change from the pools. Subsequently, these estimates of initial pool sizes were corrected to account for the disbalance in pools after 7 seasons of NK or NP applications.
The Hanninghof experiment had data on crop nutrient uptakes from the start of the experiment in 1958. Yet crop uptakes revealed very unbalanced pools in the first years with very large K uptakes, likely reflecting use of fertilizers before the experiment. Therefore it was decided to skip the first 3 years of the rotation. Initial pools were estimated from uptakes in the NK and NP treatments in 1961. With these initial pool estimates, a full rotation was run to correct for remaining disbalance in the soil pools after 3 years of rotation.
The Broadbalk experiment was running for a long period of time under a monocrop of wheat and has been fertilized since 1852. Strips received different fertilizers in the wheat experiment since 1926 up to 1968. However, N, P and K uptakes were only available after rotations started in 1968. Pool sizes in 1968 were derived from the NKMg and NP treatments of Sect. 1. These pools were adjusted for differences in pools that can be expected after 42 years of unbalanced NP or NK fertilization. To this end, ratios of pools before and after 42 years of fertilization were computed using the equations above, to estimate the initial pools in 1926. With these estimates, various treatments were applied for 42 years to arrive at reasonable pool estimates for the different strips in 1968.
Model calibration/parameterization
The relative transfer rate from stable to labile pools were set at a fixed value of 1/30, reflecting an average residence time of 30 years for the Pstable and Kstable pools (Wolf et al. 1987). Model parameters related to relative transfer rates from labile to stable pools and crop specific relative uptake rates for P and K were calibrated. The model can be run with two options for SN. The first option is to use SN as explained above with an estimated NuptakeREF value derived from the measured N uptake or yields in the control or PK treatment in combination with estimates for the recovery of N in fertilizer and manure. Alternatively, SN can be set to actual N uptakes of any treatment as second option. Errors in the estimated SN, will directly affect uptake of P and K. Hence, this second option with measured N uptake values replacing SN were used for calibration (i.e. for the Hanninghof and Broadbalk experiments). Note that for Siaya, measured N uptakes were only available for two seasons.
To calibrate, the “optim” function of the ‘Rfast’ package was used in R software (Tsagris and Papadakis 2022). The value that was minimized was a combination of the root of the mean squared errors for P uptake (RMSEP) and K uptake (RMSEK). The RMSEK was about four times larger than RMSEP and hence the cost function was determined as 4 × RMSEP + RMSEK.
In the first calibration step, all selected parameters (relative transfer rates for labile and FYM pools and relative uptake rates) were calibrated in one calibration run including all treatments for each experiment. In a second step, the crop-specific relative uptake rates (rurPFYM, c and rurKFYM, c ) that determine uptake from the PFYM and KFYM pools were set to the calibrated values of rurPlabile,c and rurKlabile,c to test if separate rur values were required. For Broadbalk, this did not affect model accuracy, in contrast to Hanninghof. Hence, in a third step, the rPFYM and rKFYM parameters were recalibrated for Hanninghof. This revealed model accuracy after calibration step 1 and 3 for P and K uptake did not differ for both the Hanninghof and Broadbalk dataset and that co-linearity between parameters was the main cause of the observed differences in the estimates of transfer rates for labile and FYM pools. Hence, we concluded that separate parameters were not required and the model was adapted accordingly. This adapted model is shown in Fig. 1. Only the calibrated parameter values and model results after this third calibration step are presented here. The mass balances for P and K were evaluated for all runs and they all added up to 0 as expected.
The model was evaluated with computed RMSE values and the Nash-Sutcliff modelling efficiency (NSE). The NSE for uptake of nutrient i was computed as:
To evaluate model accuracy after calibration, calibrated parameter values were used to predict uptake for each year and treatment combination, using measured or estimated N uptake from the soil, and values for NSE and RMSE for N, P and K uptake were computed per experiment. For the Siaya dataset, all NK and NP treatment plots were left out for the long rainy season in 2016 for P and K uptakes, as these treatment plots were used for model initialisation.
Results
Fitted linear regression models
The fitted linear regression models for uptake of N, P and K explained between 55 and 82% of total variation (Table 1). The models captured a larger proportion of variation in the Broadbalk dataset when compared to Hanninghof, indicating that the annual variability was larger for Hanninghof. The interactions between crop and year were strongly significant for P and K uptake in both experiments, but not for N uptake in Hanninghof. Interactions between crop and treatment were strongly significant for N, P and K uptake in both experiments, as expected. The interaction between crop and year was not significantly affecting the recovery of applied N in the Broadbalk experiment in contrast to Hanninghof. The linear models captured only 30–74% of the variation in the recovery of N. A larger proportion of the variation in the recovery was captured for applied mineral N fertilizer when compared to organic N from FYM or to mineral N in combination with FYM.
Parameter calibration
The calibrated parameters for relative transfer rates \({rP}_{labile}\) and \({rK}_{\text{l}\text{a}\text{b}\text{i}\text{l}\text{e}}\) differed between experiments and differed between labile and FYM pools (Table 2).
For Broadbalk, separate FYM pools did not resulting in better model estimates for P or K uptake, contrasting with Hanninghof. This may be understood when considering that FYM was applied as cattle manure in Broadbalk annually in autumn, whereas pig manure was applied once every 3 years only, 10 days before planting potatoes, in Hanninghof. These differences in input amounts resulted in PFYM and KFYM pools that were much larger than the Plabile and Klabile pools in Broadbalk. In Hanninghof the KFYM and PFYM pools were much smaller than the Plabile and Klabile pools, also due to additional uptake by oats and rye following potatoes.
Surprisingly, the calibrated value of \({rurP}_{placed}\) was the same as \({rurP}_{labile}\) in Siaya (Table 3), although Plabile and PPlaced pools strongly differed and showed different dynamics. The differences in relative uptake rates are fully explained by the differences in crop uptake rates and soil pool sizes. The relative uptake rates can be directly computed from the ratio between estimated uptake and labile pool size for omission treatments (Fig. 2). This means that for new crops, the rur can directly be estimated when uptake relative to another crops is known. For example, a crop with 20% lower P or K uptake than e.g. potato should also have a 20% lower rur than potato. From Table 3, it can be derived that P uptake for winter rye in Hanninghof is a factor 1.8 higher than for potato, while K uptake in winter rye is only 0.72 of K uptake in potato.

Relationship between computed relative uptake rates (rur) and calibrated rur parameter values for a P in treatments with NK and b K in treatments with NP fertilizer. Relative uptake rates were computed as the mean of predicted uptakes divided by the mean of the size of the labile pool over all years
These relative uptake rates can also be used to asses recovery of applied fertilizer in the year of application, i.e. the extra uptake of every kg applied when accounting for the proportion that is going to the labile pool. For example, adding 1 kg ha−1 of superphosphate-P increases the Plabile pool with 0.8 kg ha−1 and the Pstable pool with 0.2 kg ha−1. When N and K are not limiting, this extra P will result in an extra uptake of 0.165 × 0.8 = 0.132 kg P ha−1 for winter wheat grown in the Broadbalk field (reflecting a P recovery of 0.132).
Are separate pools for FYM required?
After step 1 of the calibration procedure, the rur for the FYM labile pools in Hanninghof were higher than those for the labile P and K pools for all crops. These higher rur for FYM pools were needed to match the larger P and K uptakes observed in treatments with NPK and FYM when compared to treatments with only NPK. However, extra uptake can be realised with a higher rur or with a larger pool resulting from a lower r value. This suggests some co-linearity between parameters: similar crop uptakes can be realised with different combinations of r and rur values. This co-linearity is also evident in Fig. 2: if the rur is lowered and the uptake remains the same, then the labile pool size needs to decrease and a higher relative transfer rate from a labile to stable pool is required. Setting the rur and r for the FYM pools to those of the labile pools did not affect model errors for Broadbalk and hence rur and r values for FYM pools were set to the values for these parameters matching the respective labile pools for P and K. For Hanninghof, setting the rur of FYM pools to those of the labile pools strongly increased model errors and required re-calibration of the rPFYM and rKFYM parameters. The RMSE values after recalibration (step 3) slightly increased to from 2.3 to 2.6 and from 20.5 to 21.3 kg ha−1 while the NSE decreased from 0.81 to 0.76 and from 0.70 to 0.67 for P and K respectively, mainly due to treatments combining FYM and inorganic fertilizer. These results show that separate parameters for rur associated with the FYM pools are not required.
In contrast to Broadbalk, separate values for rPFYM and rKFYM were required for Hanninghof. A reason for this difference may be found in other growth limitations that are only present in treatments without FYM, such as Mg. Adding Mg increased uptakes of P and K in treatments without FYM with about 8% at Hanninghof, indicating that Mg deficiency was present. For the Broadbalk experiment, the difference between treatments with or without Mg was not consistent across sections in the experiment and hence no clear deficiency was observed.
Model accuracy
The overall accuracy of the calibrated model was high, with a NSE of about 0.8 for N, P and K uptake (Fig. 3). The overall RMSE values were 15.6, 3.0 and 19.6 kg ha−1 for N, P and K uptake respectively.

Measured versus predicted uptakes for N, P and K. The black line represents the 1:1 line. Measured uptakes were corrected for annual variation. The root mean squared error (RMSE) was computed for all experiments combined. Points in panels reflect all possible combinations of years and crops
There were some differences between datasets, although differences between runs with measured and estimated N uptakes were relatively small (Table 4). The calibrations with measured N uptakes resulted in high overall NSE values of 0.6–0.9 (Table 4). RMSE values for P (1.9–3.1 kg P ha−1) and K uptake (15–20 kg K ha−1) were relatively small. When estimates for the N uptake (based on N uptakes in control or PK treatment combined with N fertilization and LM predicted recovery values) were used, errors for N uptake varied between 15 and 16 kg N ha−1. With these estimates for N uptakes, the model errors for the predicted P and K uptakes increased slightly with 0.2–0.7 kg P ha−1 and 0-5.6 kg K ha−1 when compared to the scenario with measured N uptakes. The largest deviations for N and K uptake were present in the Hanninghof dataset. These were associated with the FYM and FYM + PK treatments. For these treatments, FYM provided also extra N to the following crops in the rotation. This was not accounted for in the estimated soil available N (this was derived from the PK treatment), hence N and also P and K uptake predictions for rye and oats were too low for these treatments (Figure S1 in the supplementary materials). Secondly, in the first years, potato suffered from cyst nematodes before a more resistant cultivar was used, reducing nutrient uptake. Consequently, soil P and K pools for these treatments were building up, resulting in larger P and K uptake in later years, especially potato with more available N from FYM that resulted in an overestimation of especially K uptake.
Siaya
The model initialization, followed by backcasting to estimate initial pools and forecasting with NK and NP applications to 2016 resulted in minor errors (< 0.2 kg P and K ha−1). P uptakes were overpredicted in 2016 and underpredicted in 2018, likely due to the relatively low yields that were observed in 2016. Also K uptakes were overpredicted in 2016 for control and PK treatments, suggesting that N uptake in 2016 was overestimated. The large changes in the supplied amounts of P and K after the LR 2016 were captured well in the estimated uptakes, suggesting that the rebuilding of soil P and K pools after depletion was described properly.
Hanninghof
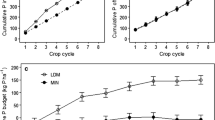
The model predictions for the Hanninghof trial were accurate with a RMSE of 2.6 kg P ha−1and 20.4 kg K ha−1 (Table 4). Overall, uptakes were simulated well for the crop rotation (Fig. 4). Differences between treatments were captured well, although P and K uptake were overestimated for the control treatment while only K uptake was overestimated for the N treatment. This overestimation was counterbalanced by an underestimation for other treatments, most strongly for treatments including Mg or FYM (Fig. 4). The overestimation became stronger in later years, evidenced by the increasing absolute error with accumulating uptakes. Overestimation of P and K uptake in the N only treatment most likely resulted from overestimation of N uptake. For the control this could not be the case, as for this treatment N uptake did not depend on estimated recovery. Over time, measured yields in the control and PK treatment increased while measured uptakes of N decreased. This suggests that crops more strongly diluted N over time, due to a higher nutrient harvest index or due to lower N concentration in grain or tubers. The model did not account for a change in this internal N use efficiency over time. A second factor is the Mg deficiency that may have reduced yields. When compared to the NPK treatment, measured cumulative uptakes of P and K in the NPK treatment with Mg added were on average 8% higher in the rotation. There were no differences between the NPK and NPKMg treatments when FYM was also applied. However, the yield difference due to Mg fertilization resulted in a larger prediction error for the NPKMg when compared to the NPK treatment (Fig. 4).

Predicted vs. measured cumulative uptake of P and K for the full crop rotation in the Hanninghof experiment. The solid black line represents the 1:1 line
Broadbalk
The RMSE was 3.4 and 19.2 kg ha−1 for P and K uptake (Table 4), with a NSE of 0.8. The model overestimated P and K uptake in the FYMNPK treatment and underestimated P uptake in the FYM treatments (Fig. 5). Measured uptakes of P and K were largest for the treatments with FYM. The observed differences between treatments were present in all crops. P uptake in the NP treatment was strongly overestimated for potato, reflecting a higher sensitivity to small K pools and K deficiency than other crops in the rotation. The errors differed most strongly between treatment for spring beans. For non-legume crops, soil N supply is limited, however for legumes more N can be supplied by fixation when demand for N exceeds N supply. The estimate of N that was supplied by organic matter mineralisation, deposition and fixation was based on PK treatments. This likely resulted in a small overestimation of N availability for other treatments when P or K deficiencies were present. However, the cumulative uptakes for spring beans were small compared to uptakes of especially potato and winter wheat, hence these errors had a relatively small effect on the estimated uptakes in the rotation as a whole.

Measured vs/ predicted cumulative uptake of P (a) and K (b) for the full crop rotation in the Broadbalk experiment. The solid black line represents the 1:1 line
Estimated pool sizes
The size of both labile and stabile P and K pools increased over the course of the experiments when treatments included P or K fertilization (Table 5). Labile pools strongly increased in the first years and were more or less constant in later years, while stable pools kept increasing. Treatments including FYM resulted in the largest increase in soil P. In Hanninghof, applications of FYM before planting of potato only resulted in some mining of soil K, while P pools increased. This mining of K was amplified when FYM was combined with NP fertilizer. FYM with NK was much better balanced, resulting in an increase in soil P pools of 419 kg P ha−1, although soil K increased with 2736 kg ha−1.
The continued annual FYM applications in the Broadbalk experiment further increased soil P and K pools in the crop rotation phase, although increased N applications in later years of the experiment better balanced N, P and K and this resulted in a moderation of the increase in stable pools. The initial pools for the FYM treatment were already high in the first year, as FYM applications started already in 1928. Yet, there was still a net increase. The large increase in Pstable and the slight increase in the Kstable pool indicate that FYM was an unbalanced source of nutrients and did not provide enough N to the crops. Note that the decrease in KFYM for the FYM treatment can be understood by the change from winter wheat to crops in rotation including potato with an associated increase in crop K demand. The total P in all soil pools were strongly and linearly related to the measured total P total values in 2000 (R2 of 0.98, slope of 1.07, Fig. S2 in the supplementary materials). The FYM applications in the FYM + NPK treatment stopped after 2001, resulting in a slight decrease in Plabile and more or less balanced Pstable pool.
In Siaya, the 23 farms differed strongly in pool sizes. The NPK treatment resulted in an increase of Pstable but a decrease of Klabile and Kstable for farms with a large initial pool sizes, whereas both P and K pools increased on farms with small initial pool sizes.
Discussion
The three long-term datasets analysed here included measurements of N, P and K uptake and yield. Such datasets are very valuable and essential to advance our understanding of nutrient interactions (Das et al. 2019) and what proportion of inputs are recovered in the long-term (Vonk et al. 2022). Availability of N, P and K in the soil have a strong effect on N recovery of applied fertilizer (Janssen et al. 1990) and N losses and hence are key to understand how best to minimize fertilizer inputs and greenhouse gas emissions. Currently few models include N and P limitations, such as APSIM and DDSAT (Das et al. 2022), although those are not yet tested on a wider range of datasets. There are few modelling attempts that account for K limitations. Yet, mining of soil K is common practice in many areas of the world (Das et al. 2021). Hence, proper K fertilization needs more attention in sustainable intensification efforts to reduce greenhouse gas emissions and improve N efficiency.
The RC-KP model was able to capture short- and long-term differences in P and K uptakes for very contrasting treatments that strongly differed in P and K supply. Overall modelling efficiency was high (0.8) with RMSE values of 3.0 kg P ha−1 and 19.6 kg K ha−1, indicating that the model was accurate. To initialise the model, measured uptakes for P and K are required for treatments without N. We have shown that various corrections can be used to estimate initial pool sizes when these uptakes are only measured later in an experiment, using combinations of back-casting, pool corrections and initial model runs to capture fertilization history. Differences between soils were captured with different relative transfer rates. Although relative uptake rates differed between crops and sites, they were fully explained by the differences in uptake of P and K between crops and sites. The relatively simple description of soil pools allowed to account for differences in placed and broadcast applications of artificial fertilizer. Strong changes in crop uptakes in response to drastic changes in fertilizer applications were captured well. The model was able to describe the large differences in unbalanced treatments on strongly depleted soils in Siaya, and the large increases in crop uptakes following fertilization after long periods of soil mining.
The RC-KP model was built following concepts in Wolf et al. (1987) and uses labile and stable pools for P and K, similar to other dynamic models. However, plant P and K availability is considered proportional to the labile P pool size, contrasting with other crop growth models where the relationship between labile P and plant available P has to be defined explicitly. The Broadbalk experiment showed that additional pools for FYM are in principle not required, although they helped to understand how growth limitations due to Mg deficiency affected uptakes in Hanninghof. Yield reductions of 12, 5 and 3% were observed for potato, oats and winter rye in the Hanninghof experiment where no Mg was applied (Jate 2012; Jate and Lammel 2022). These Mg limitations also reduced P and K uptake, by about 8%. These effects were not accounted for in the RC-PK model and caution is needed when using the model for sites where other constraints or deficiencies are present.
The soil pools for P and K continued to increase for all experiments when P or K was added, more strongly when uptake was limited by another nutrient. Treatments including FYM and NPK in both the Hanninghof and the Broadbalk experiment resulted in the strongest increase in P and K pools, more so for the stable than for the labile pools. Soil tests in Broadbalk showed a modest increase in Olsen P values over time (Blake et al. 2000). The modelled pools sizes for P in this study were very strongly related (R2 of 0.98) to measured total P for Broadbalk in 2000, while the relatively small quantities of P leached (Heckrath et al. 1995) were not accounted for in the RC-KP model. In response to this strong increase in total P and associated losses, in 2001 P fertilization was stopped for some treatments, while FYM was no longer applied in the FYM + NPK treatment. In our simulations, soil labile P pools declined and stable P pools were more or less steady and no longer increasing after 2001.
The FYM without NPK treatment resulted in a steady build-up of soil P pools, while labile and stable K pools decreased only slightly for Hanninghof. Treatments in this experiment with mineral P fertilization increased soil P status, and all treatments with mineral K increased soil K status (Jate 2010, 2012; Jate and Lammel 2022). This indicates that the treatments were not optimal and oversupplied P and/or K compared with plant demand, more so in earlier years. Nutrient applications were better balanced after 1980 when a nematode resistant potato cultivar was used and N applications increased. Also all other treatments including FYM increased soil P status in Hanninghof, despite the moderate applications of 25 t ha−1 pig manure once every three years (Jate and Lammel 2022). The annual applications of 35 t ha−1 cattle manure in the Broadbalk experiment provided excessive amounts of P and K, far exceeding crop uptake that resulted in a strong increase in soil pools. These increased soil P pools for all FYM treatments strongly increasing P leaching risks on both locations in the longer term. The N applications of 250 kg N ha−1 in the Broadbalk experiment exceeded the maximum of 170 kg N ha−1 that is allowed in the EU to limit nitrate leaching. From these experiments it is clear that FYM applications should also be limited to reduce P leaching risks when soil pools are saturated, as is the case in many areas of Europe (Fischer et al. 2017; Schoumans and Chardon 2015).
In the Broadbalk experiment, large differences in soil C pools were present. Soil C increased from 80 to 90 t C ha−1 in the period of 1960–2000 for treatments with FYM, while treatments without FYM were more or less constant at about 27 t C ha−1 (Johnston and Poulton 2018). Also in Hanninghof, soil C contents increased for all treatments with FYM with 4–14% or 1.7–5.9 t C ha−1 in the 50 years up to 2008, while treatments without FYM were more or less constant (Jate 2012; Jate and Lammel 2022). This indicates that only a small fraction of the P applied as FYM was locked-up in soil organic matter, increasing by about 118 (Hanninghof) and 250 (Broadbalk) kg C ha−1 y−1 and 1.2–2.5 kg P ha−1 y−1 when assuming a C:P ratio of 100:1, suggesting that separate organic P pools may not be needed.
Interactions between N, P and K were very important and significantly affected short- and long-term recovery of applied fertilizers. These interactions were captured well by the QUEFTS principles, as explained. A better understanding of these interactions is required to maximise agronomic efficiency of applied N and reduce GHG emissions from agriculture. The P and K uptake for treatments including N from FYM or synthetic fertilizer was estimated well. The uptake of N in control and PK treatments was overestimated, and this overestimation increased over time when pools were more strongly depleted. A complicating factor was the increased availability of N and strongly increased yield over time, also for the treatments without N additions (Johnston and Poulton 2018).
Current soil P models include detailed processes yet poorly describe P availability for plants (Hinsinger 2001; Muntwyler et al. 2023), caused by poor quantitative understanding of the uptake efficiency of exploited soil volumes by roots and associated mycorrhiza (Hinsinger et al. 2011) and plant-species specific root-soil interactions through e.g. extrudates affecting nutrient availability in the soil (Richardson et al. 2009). Further orthophosphate and K in solution also depend on soil pH, soil water content dynamics and plant uptake (Xu et al. 2023). The RC-KP does not include detailed processes, and ignores e.g.: annual variability related to weather conditions; the role of soil characteristics on P solubility (Saentho et al. 2022); and plant characteristics and N availability effects on P solubility and uptake (Barrow 2017). All plant-soil interactions were captured by field and crop specific parameters to test if main interactions affecting crop N, P, K uptake were properly captured.
The RC-KP model can be used to assess P and K uptakes that vary from year to year, although we were not directly focused on describing annual variation. Upon testing where N supply was set to the measured annual N uptake, errors in P and K uptake estimates did not increase strongly when compared to the scenario with estimated N uptakes using linear regression models. In our approach, N uptake was a forcing variable, i.e. it was provided as a value for each year or it was estimated from estimates of soil N supply and N recovery. This N supply by the soil is notoriESously difficult to assess accurately without measured crop uptakes from unfertilized crops. However, dynamic crop growth models that include a soil N module, or standard N supply estimates for soil types are typically available. With this, N uptake can be estimated for rainfed or irrigated crops without P and K limitations and these can be used as input for the RC-KP model to predict N, P and K uptakes under nutrient limited conditions. Hence, combining or integrating the RC-KP model with a crop growth model provides a novel new approach to account for weather variability and N, P and K limitations. In other work, we have shown that the nutrient equivalent concepts (Janssen 1998, 2017), that also builds on the QUEFTS approach, allowed us to balance N, P and K uptakes on a daily basis (Adiele et al. 2022). The underlying principles that determine relative crop N, P and K demand seem therefore scalable, i.e. they are useful to describe interactions between N, P and K that determine uptake on time-scales of a day, a season and a period of 50 years.
Knowledge of annually varying uptake of P and K is in particular important to account for uptake limitations when crop growth is limited, e.g. by water deficiency or reduced by pest and diseases. Our model was able to account for rotation effects and can be used to evaluate fertilization strategies and adapt applications to varying uptake. This enables a more efficient use of resources in both organic and conventional farming systems.
Conclusion
The novel RC-KP model is a deterministic and dynamic model that uses a simple approach to predict annual N, P and K uptake in crop rotations. The model describes the dynamics in labile and stable soil P and K pools in response to organic and synthetic fertilizer inputs and crop uptakes. The model can be run for situations with known N uptakes but also for situations where only soil N supply and crop N recovery are known. This relatively simple RC-KP model was able to describe accurately N, P and K uptakes in crop rotations in the three long-term datasets. The Nash-Sutcliff modelling efficiency ranged from 0.6 to 0.9, with a root mean squared error of 2.6–3.4 kg P ha−1 and 14–20 kg K ha−1. The model needs site-specific parameters for relative transfer rates from labile to stable pools and crop specific relative uptake rates for P and K. These required crop-specific relative uptake rates are well understood and link annual P and K uptakes to soil pool sizes. This model can be used to explore the short and long-term effect of different application rates and composition of fertilisers in crop rotations. As such it is an important tool to ensure balanced nutrition of crops and efficient use of added nutrients, with important implications for reducing nutrient losses to the environment.
Data availability
The processed data for Broadbalk and Siaya and the R code for the RC-KP model are available under a EUPL-1.2 license at models.pps.wur.nl/models. The Hanninghof dataset can be provided upon request with permission from Mr Melkamu Jate.
References
Adiele JG, Schut AGT, Ezui KS, Giller KE (2022) LINTUL-Cassava-NPK: a simulation model for nutrient-limited cassava growth. Field Crops Res 281:108488
Andersson S, Simonsson M, Mattsson L, Edwards AC, ÖBorn I (2007) Response of soil exchangeable and crop potassium concentrations to variable fertilizer and cropping regimes in long-term field experiments on different soil types. Soil Use Manag 23:10–19
Barrow NJ (2017) The effects of pH on phosphate uptake from the soil. Plant Soil 410:401–410
Blake L, Mercik S, Koerschens M, Moskal S, Poulton PR, Goulding KWT, Weigel A, Powlson DS (2000) Phosphorus content in soil, uptake by plants and balance in three european long-term field experiments. Nutr Cycl Agroecosyst 56:263–275
Cate RB Jr, Nelson LA (1971) A simple Statistical Procedure for partitioning soil test correlation data into two classes. Soil Sci Soc Am J 35:658–660
Damar H, Ziadi N, Lafond J, Parent LE (2020) Potassium transformation in clay soil with contrasting K budgets in long-term experiment. Agron J 112:5180–5192
Das B, Huth N, Probert M, Condron L, Schmidt S (2019) Soil phosphorus modeling for Modern Agriculture requires Balance of Science and practicality: a perspective. J Env Qual 48:1281–1294
Das D, Dwivedi BS, Datta SP, Datta SC, Meena MC, Dwivedi AK, Singh M, Chakraborty D, Jaggi S (2021) Long-term differences in nutrient management under intensive cultivation alter potassium supplying ability of soils. Geoderma 393:114983
Das BT, Schmidt S, Biggs J, Lester DW, Bourne N, Huth NI (2022) Simulating long-term phosphorus, nitrogen, and carbon dynamics to advance nutrient assessment in dryland cropping. Field Crops Res 285:108590
de Wit CT (1953) A physical theory on placement of fertilizers. Versl Landbouwk Onderz A 59:1–71. https://edepot.wur.nl/345397
Dzotsi KA, Jones JW, Adiku SGK, Naab JB, Singh U, Porter CH, Gijsman AJ (2010) Modeling soil and plant phosphorus within DSSAT. Ecol Model 221:2839–2849
Fischer P, Pöthig R, Venohr M (2017) The degree of phosphorus saturation of agricultural soils in Germany: current and future risk of diffuse P loss and implications for soil P management in Europe. Sci Total Environ 599–600:1130–1139
Glendining MJ, Poulton PR, Macdonald AJ (2021) Broadbalk Wheat Experiment plan and cropping 1968–2017. Electronic Rothamsted Archive. Rothamsted Research, Harpenden
Heckrath G, Brookes PC, Poulton PR, Goulding K W T (1995) Phosphorus leaching from soils containing different phosphorus concentrations in the Broadbalk experiment. J Env Qual 24:904–910
Hinsinger P (2001) Bioavailability of soil inorganic P in the rhizosphere as affected by root-induced chemical changes: a review. Plant Soil 237:173–195
Hinsinger P, Brauman A, Devau N, Gérard F, Jourdan C, Laclau J-P, Le Cadre E, Jaillard B, Plassard C (2011) Acquisition of phosphorus and other poorly mobile nutrients by roots. Where do plant nutrition models fail? Plant Soil 348:29–61
Ichami SM, Shepherd KD, Sila AM, Stoorvogel JJ, Hoffland E (2019) Fertilizer response and nitrogen use efficiency in african smallholder maize farms. Nutr Cycl Agroecosyst 113:1–19
Janssen BH (1998) Efficient use of nutrients: an art of balancing. Field Crops Res 56:197–201
Janssen BH (2017) Crop yields and NPK use efficiency of a long-term experiment on a former sea bottom in the Netherlands. Wageningen University, Plant Production Systems, Wageningen
Janssen BH, Lathwell DJ, Wolf J (1987) Modelling long-term crop response to fertilizer phosphorus II. Comparison with field results. Agron J 79:452–458
Janssen BH, Guiking FCT, Eijk D, Smaling E, Wolf J, van Reuler H (1990) A system for quantitative evaluation of the fertility of Tropical Soils (QUEFTS). Geoderma 46:299–318
Jate M (2010) Long-term effect of balanced mineral fertilizer application on potato, winter rye, and oat yields; nutrient use efficiency; and soil fertility. Arch Agron Soil Sci 56:421–432
Jate M (2012) Impact of mineral fertilizer integration with farmyard manure on crop yield, nutrient use efficiency, and soil fertility in a long-term trial. In: Sharma P, Abrol V (eds) Crop Production Technologies. IntechOpen, Online
Jate M, Lammel J (2022) Effect of balanced and integrated crop nutrition on sustainable crop production in a classical long-term trial. In: Meena VS, Choudhary M, Yadav RP, Meena SK (eds) Sustainable Crop Production. IntechOpen, Rijeka. p. Ch. 2
Johnston AE, Poulton PR (2018) The importance of long-term experiments in agriculture: their management to ensure continued crop production and soil fertility; the Rothamsted experience. Eur J Soil Sci 69:113–125
Kihara J, Nziguheba G, Zingore S, Coulibaly A, Esilaba A, Kabambe V, Njoroge S, Palm C, Huising J (2016) Understanding variability in crop response to fertilizer and amendments in sub-saharan Africa. Agric Ecosyst Environ 229:1–12
Kumar P, Dua VK, Sharma J, Byju G, Minhas JS, Chakrabarti SK (2018) Site-specific nutrient requirements of NPK for potato (Solanum tuberosum L.) in western indo-gangetic plains of India based on QUEFTS. J Plant Nutr 41:1988–2000
Lewis DR, McGechan MB (2002) SW—Soil and water: a review of field scale phosphorus dynamics models. Biosyst Eng 82:359–380
Li G, Li H, Leffelaar PA, Shen J, Zhang F (2014) Characterization of phosphorus in animal manures collected from three (dairy, swine, and broiler) farms in China. PLoS ONE 9:e102698
Lyzenga WJ, Liu Z, Olukayode T, Zhao Y, Kochian LV, Ham B-K (2023) Getting to the roots of N, P, and K uptake. J Exp Bot 6:1784–1805. https://doi.org/10.1093/jxb/erad035
Muntwyler A, Panagos P, Morari F, Berti A, Jarosch KA, Mayer J, Lugato E (2023) Modelling phosphorus dynamics in four european long-term experiments. Agr Syst 206:103595
Nair KPP (1992) Measuring P buffer power to improve routine soil testing for phosphate. Eur J Agron 1:79–84
Nguyen MH (2003) Soil potassium dynamics under intensive rice cropping. A case study in the Mekong Delta, Vietnam. Wageningen University, Wageningen
Njoroge SK (2019) Feed the crop, not the soil! Explaining variability in maize yield responses to nutrient applications in smallholder farms of western Kenya. Wageningen University, Wageningen
Njoroge S, Schut AGT, Giller KE, Zingore S (2017) Strong spatial-temporal patterns in maize yield response to nutrient additions in african smallholder farms. Field Crops Res 214:321–330
Njoroge S, Schut AGT, Giller KE, Zingore S (2019) Learning from the soil’s memory: tailoring of fertilizer application based on past manure applications increases fertilizer use efficiency and crop productivity on kenyan smallholder farms. Eur J Agron 105:52–61
Papangelou A, Mathijs E (2021) Assessing agro-food system circularity using nutrient flows and budgets. J Environ Manage 288:112383
Pasley HR, Cairns JE, Camberato JJ, Vyn TJ (2019) Nitrogen fertilizer rate increases plant uptake and soil availability of essential nutrients in continuous maize production in Kenya and Zimbabwe. Nutr Cycl Agroecosystems 115:373–389
Penn CJ, Camberato JJ (2019) A critical review on Soil Chemical processes that control how Soil pH affects phosphorus availability to plants. Agriculture 9:120
Petersen J, Thomsen IK, Mattsson L, Hansen EM, Christensen BT (2010) Grain yield and crop N offtake in response to residual fertilizer N in long-term field experiments. Soil Use Manag 26:455–464
Petersen J, Thomsen IK, Mattsson L, Hansen EM, Christensen BT (2012) Estimating the crop response to fertilizer nitrogen residues in long-continued field experiments. Nutr Cycl Agroecosystems 93:1–12
R Core Team (2021) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Richardson AE, Hocking PJ, Simpson RJ, George TS (2009) Plant mechanisms to optimise access to soil phosphorus. Crop Pasture Sci 60:124–143
Rothamsted Research (2018) Broadbalk experiment fertilizer and manure treatments 1852–2021. Electronic Rothamsted Archive. Rothamsted Research, Harpenden
Saentho A, Wisawapipat W, Lawongsa P, Aramrak S, Prakongkep N, Klysubun W, Christl I (2022) Speciation and pH- and particle size-dependent solubility of phosphorus in tropical sandy soils. Geoderma 408:115590
Sattari SZ, Ittersum MK, v, Bouwman AF, Smit AL, Janssen BH (2014) Crop yield response to soil fertility and N, P, K inputs in different environments: testing and improving the QUEFTS model. Field Crops Res 157:35–46
Scanlan CA, Huth NI, Bell RW (2015) Simulating wheat growth response to potassium availability under field conditions with sandy soils. I. Model development. Field Crops Res 178:109–124
Schoumans OF, Chardon WJ (2015) Phosphate saturation degree and accumulation of phosphate in various soil types in the Netherlands. Geoderma 237–238:325–335
Schut AGT, Giller KE (2020) Soil-based, field-specific fertilizer recommendations are a pipe-dream. Geoderma 380:114680
Setiyono TD, Walters DT, Cassman KG, Witt C, Dobermann A (2010) Estimating maize nutrient uptake requirements. Field Crops Res 118:158–168
Shehu BM, Lawan BA, Jibrin JM, Kamara AY, Mohammed IB, Rurinda J, Zingore S, Craufurd P, Vanlauwe B, Adam AM, Merckx R (2019) Balanced nutrient requirements for maize in the northern nigerian Savanna: parameterization and validation of QUEFTS model. Field Crops Res 241:107585
Simonsson M, Andersson S, Andrist-Rangel Y, Hillier S, Mattsson L, Öborn I (2007) Potassium release and fixation as a function of fertilizer application rate and soil parent material. Geoderma 140:188–198
Simonsson M, Hillier S, Öborn I (2009) Changes in clay minerals and potassium fixation capacity as a result of release and fixation of potassium in long-term field experiments. Geoderma 151:109–120
Soetaert K, Petzoldt T, Setzer RW (2010) Solving Differential equations in R: Package deSolve. J Stat Softw 33:1–25
Steinfurth K, Börjesson G, Denoroy P, Eichler-Löbermann B, Gans W, Heyn J, Hirte J, Huyghebaert B, Jouany C, Koch D, Merbach I, Mokry M, Mollier A, Morel C, Panten K, Peiter E, Poulton PR, Reitz T, Rubæk GH, Spiegel H, van Laak M, von Tucher S, Buczko U (2022) Thresholds of target phosphorus fertility classes in european fertilizer recommendations in relation to critical soil test phosphorus values derived from the analysis of 55 european long-term field experiments. Agric Ecosyst Environ 332:107926
Struik PC, Kuyper TW (2017) Sustainable intensification in agriculture: the richer shade of green. A review. Agron Sust Dev 37:39
Tabi FO, Diels J, Ogunkunle AO, Iwuafor ENO, Vanlauwe B, Sanginga N (2008) Potential nutrient supply, nutrient utilization efficiencies, fertilizer recovery rates and maize yield in northern Nigeria. Nutr Cycl Agroecosyst 80:161–172
ten Berge HF, Hijbeek M, van Loon R, Rurinda MP, Tesfaye J, Zingore K, Craufurd S, van Heerwaarden P, Brentrup J, Schröder F, Boogaard JJ, de Groot HL, van Ittersum MK (2019) Maize crop nutrient input requirements for food security in sub-saharan Africa. Glob Food Sec 23:9–21
Tsagris M, Papadakis M (2022) Forward Regression in R: from the Extreme slow to the Extreme fast. J Data Sci 16:771–780
van der Bom F, Magid J, Jensen LS (2017) Long-term P and K fertilisation strategies and balances affect soil availability indices, crop yield depression risk and N use. Eur J Agron 86:12–23
van der Eijk D, Janssen BH, Oenema O (2006) Initial and residual effects of fertilizer phosphorus on soil phosphorus and maize yields on phosphorus fixing soils: a case study in South-West Kenya. Agric Ecosyst Environ 116:104–120
Vonk WJ, Hijbeek R, Glendining MJ, Powlson DS, Bhogal A, Merbach I, Silva JV, Poffenbarger HJ, Dhillon J, Sieling K, ten Berge HFM (2022) The legacy effect of synthetic N fertiliser. Eur J Soil Sci 73:e13238
Wolf J, Wit CT, d, Janssen BH, Lathwell DJ (1987) Modelling long-term response to fertilizer phosphorus. I. The model. Agron J 79:445–451
Xu X, Xie J, Hou Y, He P, Pampolino MF, Zhao S, Qiu S, Johnston AM, Zhou W (2015) Estimating nutrient uptake requirements for rice in China. Field Crops Res 180:37–45
Xu D, Zhu Q, Ros GH, Xu M, Wen S, Zhang F, de Vries W (2023) Model-based optimal management strategies to mitigate soil acidification and minimize nutrient losses for croplands. Field Crops Res 292:108827
Yang X, Post WM (2011) Phosphorus transformations as a function of pedogenesis: a synthesis of soil phosphorus data using Hedley fractionation method. Biogeosci Disc 8:5907–5934
Acknowledgements
We gratefully acknowledge Mr Melkamu Jate for his assistance and Yara for providing the data for the trial at Hanninghof, Dr Margaret Glendining for her assistance with the Broadbalk dataset and Dr Samuel Njoroge for providing the Siaya dataset. We thank the Lawes Agricultural Trust and Rothamsted Research for Broadbalk data from the e-RA database. The Rothamsted Long-term Experiments National Capability is supported by the UK BBSRC (Biotechnology and Biological Sciences Research Council, BBS/E/C/000J0300, 2017–2023) and the Lawes Agricultural Trust. We are grateful for the comments provided by three independent reviewers that substantially improved the manuscript.
Author information
Authors and Affiliations
Contributions
Both authors contributed to the study conception, design, data collection and analysis. The first draft of the manuscript was written by Dr Antonius G.T. Schut and W. Reymann commented on all versions of the manuscript. Both authors read and approved the final manuscript.
Corresponding author
Ethics declarations
The authors have no relevant financial or non-financial interests to disclose.
Additional information
Responsible Editor: Adamo Domenico Rombolà.
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Below is the link to the electronic supplementary material.
ESM 1
(DOCX 259 KB)
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Schut, A.G.T., Reymann, W. Towards a better understanding of soil nutrient dynamics and P and K uptake. Plant Soil 492, 687–707 (2023). https://doi.org/10.1007/s11104-023-06209-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-023-06209-x