
Abstract
Purpose
Partial rootzone drying (PRD) typically alternates the dry and irrigated parts of the rootzone, but how plant physiology and soil evaporation respond to this alternation are poorly understood.
Methods
Dwarf tomatoes were grown in small split pots comprising two 250 cm3 compartments and fully irrigated (WW: 100% ETc) or subjected to three deficit irrigation treatments (75% ETc): homogeneous rootzone drying (HRD; irrigation evenly distributed); fixed PRD (PRD-F, irrigation applied to one fixed compartment); alternated PRD (PRD-A: as PRD-F but alternating the irrigated compartment every three days). Stem diameter and evapotranspiration were monitored during alternation cycles. The day after alternating the irrigated side of the root system, whole-plant gas exchange and leaf water potential were measured following step increments of vapour pressure deficit.
Results
Alternation did not affect stem diameter contractions or evapotranspiration, which were lower in HRD than in the two PRD treatments. However, soil evaporation was higher in HRD and PRD-A after alternation than in PRD-F. Following alternation, higher soil evaporation was counteracted by decreased transpiration compared with fixed PRD, despite similar overall soil water content. VPD increments did not change this pattern.
Conclusion
Irrigation placement determined soil moisture distribution, which in turn affected soil evaporation and whole plant gas exchange. Optimising the frequency of PRD alternation to maximise water savings while ensuring productive water use needs to consider how soil moisture distribution affects both soil evaporation and plant water use.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Water-saving irrigation techniques that decrease crop evapotranspiration (ET) can alleviate increasing pressure on water resources for agriculture. However, since crop yield is linearly related to ET and only saturates at high ET (Fereres and Soriano 2007), restricting transpiration (which generally accounts for the greatest proportion of ET) without decreasing yield is challenging. Nevertheless, deficit irrigation techniques that deliberately apply less irrigation volume than potential crop evapotranspiration (ETc) while maintaining yields have been adopted. While most studies of deficit irrigation determine how little water can be applied without affecting yield, irrigation timing and placement within the rootzone can also improve crop water use efficiency at the same irrigation volumes (Dodd et al. 2015; Sebastian et al. 2016).
The partial rootzone drying (PRD) technique was developed to improve crop water use efficiency by changing the placement of deficit irrigation (During et al. 1996). Under PRD, a percentage of ETc water volume is applied locally to one part (usually half) of the rootzone, which maintains leaf water status while decreasing stomatal conductance and shoot growth compared to evenly distributed irrigation at 100%ETc (Kang et al. 1998; Dry and Loveys 1999). Many crops show enhanced xylem concentrations of the antitranspirant hormone abscisic acid (ABA) (Stoll et al. 2000; Liu et al. 2006; Pérez-Pérez et al. 2018), and changes in other phytohormones (Tardieu 2016; Pérez-Pérez et al. 2020) during PRD. ABA-mediated partial stomatal closure restricts transpiration while affecting photosynthesis less (Thompson et al. 2007), thereby increasing instantaneous leaf water use efficiency (the ratio of photosynthesis to transpiration). Sustaining this effect over the growing season allows PRD to improve crop water use efficiency (i.e. their yield per irrigation volume) compared to homogeneous application with the same irrigation volume (Dodd 2009).
However, instantaneous water use efficiency may only transiently increase during PRD because stomata re-opens as the soil of the non-irrigated rootzone further dries (Dry et al. 2000; Stoll et al. 2000). This was attributed to reduced export of xylem-borne ABA from dry roots as their contribution to total sap flow declines with soil water content (Dodd et al. 2008a). This attenuation of PRD-induced signalling requires regular alternation of the irrigated side, to continue to restrict transpiration while maintaining shoot water status and growth. Other physiological measurements may help determine when PRD alternation should occur.
Despite much literature analysing the effects of PRD on different crops (Adu et al. 2018), the timing of PRD alternation is usually empirically based, causing variable effects on crop yield (Dodd et al. 2015). At the same irrigation volumes, alternation usually enhanced yield of field grown crops compared to fixed PRD, associated with higher stomatal conductance of pepper (Kang et al. 2001), but no differences in cotton plant water use (Du et al. 2006; Tang et al. 2005). These measurements were usually made in the middle of the alternation cycle, but few contradictory studies have evaluated the dynamics of physiological responses to alternation. Thus, alternation transiently decreased stomatal conductance of containerised tomato plants (receiving 50% less irrigation) associated with higher xylem ABA concentration (Dodd et al. 2006). Since sustained soil drying likely stimulated root ABA accumulation (Pérez-Pérez et al. 2020), re-watering likely substantially increased ABA export from those roots. However, when the wet side of the rootzone was maintained close to saturation, alternation did not decrease stomatal conductance or increase xylem ABA concentration in tomato (Pérez-Pérez and Dodd 2015; Pérez-Pérez et al. 2020). The intensity of the ABA signal, and hence magnitude of stomatal closure, depends not only on the soil moisture of the dry side of the rootzone but also of the wet side (Dodd et al. 2008b; Puértolas et al. 2016). Thus xylem ABA increments and consequential partial stomatal closure following PRD alternation could be attributed to drying of the former wet side before full rehydration of the newly irrigated side.
Chemical long-distance signals interact with hydraulic signals to mediate shoot responses to water deficits (Tardieu 2016). Since PRD aimed to exploit long distance ABA signalling, potential changes in plant hydraulic conductance following PRD alternation have often been neglected. Re-watering previously dry roots can rapidly increase their hydraulic conductance (Kang et al. 2003) by enhancing aquaporin expression (McLean et al. 2011). Both hydraulic and chemical signalling models can simulate transpiration changes during PRD alternation (Huber et al. 2014), but the relative importance of each mechanism is still uncertain and may vary as evaporative demand changes during the day. Under low evaporative demand, progressive soil drying of half of the rootzone (with the remaining roots in wet soil) maintained leaf water potential, with root-sourced chemical signals triggering stomatal closure (Sobeih et al. 2004). However, under increasing evaporative demand, water supply from the wet rootzone might not meet greater transpirational requirements, thereby decreasing leaf water potential, causing turgor loss in guard cells and eliciting stomatal closure (Buckley 2019). Using soil-root water flow models to analyse the roles of chemical and hydraulic signals suggests that measuring plant transpiration and water potential at several evaporative demands can help discriminate the relative contribution of both mechanisms to stomatal control (Huber et al. 2015), thereby helping to predict the impact of PRD alternation under different environmental conditions.
Transpiration decreases xylem water potential causing water to flow from living vascular tissues into the xylem to partially sustain transpirational flow. Continuously monitoring the extent and velocity of stem diameter contractions, as applied in automated irrigation scheduling (Fernández and Cuevas 2010), can estimate changes in xylem tension (Klepper et al. 1971) between treatments during PRD alternation. Nevertheless, relatively few studies have measured these responses to PRD (e.g. Egea et al. 2011; Elsayed-Farag and Melgar 2015; Consoli et al. 2014).
Changing irrigation placement and timing can also affect soil evaporation rates. Since PRD usually waters only half the rootzone, it can reduce evaporation rates thereby increasing irrigation efficiency (defined as yield per irrigation volume unit), compared to homogeneous soil drying (Marsal et al. 2008). However, when irrigation is applied frequently enough, PRD alternation might increase soil evaporation from the whole rootzone. Evaporation increases with the wetted soil surface area (Leib et al. 2006), which increases dramatically immediately after alternating the irrigated side as surface soil moisture of both the newly and former irrigated sides is transiently high. Although some field studies have attempted to estimate whether PRD alters the evaporative component of ET (Leib et al. 2006; Marsal et al. 2008), dynamic changes in evaporation during PRD alternation cycles are poorly understood. While conventional rainfed cropping has emphasised the importance of minimising evaporative water losses while maximising transpiration to increase crop water use efficiency (Hatfield and Dold 2019), relatively few studies have determined how PRD affects of crop evapotranspiration components (evaporation and transpiration) when wet and dry sides are alternated. Such information is essential to determine the potential water savings of the technique.
To better understand the dynamics of water losses from soil and crop during alternate partial rootzone drying, spatial and temporal variation in soil evaporation and plant physiological responses were investigated. Different irrigation treatments (homogeneous rootzone drying, and fixed and alternate PRD) were applied to split-root dwarf tomato (MicroTom) plants to measure whole plant gas exchange responses to changes in evaporative demand, both before and after PRD alternation. Unlike fast growing tall tomato genotypes, this variety dried the soil more slowly, allowing longer (three days) alternation cycles with substantial soil drying comparable to field studies. Diurnal stem contractions were readily monitored in this slow growing genotype, as these are overridden by fast diameter growth in tall tomato (Puértolas et al. 2020a). We hypothesised that
-
1.
PRD alternation limits hydraulic conductance more than other deficit treatments, causing larger stem diameter contractions and restricting stomatal conductance (estimated from whole-plant gas exchange measurements) under increasing VPD.
-
2.
PRD alternation transiently increases soil evaporation from the recently irrigated rootzone, causing greater evaporation from the whole rootzone than other deficit treatments.
Material and methods
Plant culture
Forty-eight seedlings of a dwarf tomato variety (Solanum lycopersicum cv. ‘MicroTom’) were raised in small (50 cm3) pots filled with an organic loam (John Innes No.2, Westland Horticulture, Dungannon, UK) and grown for four weeks, so roots colonised the whole pot volume. By gently extracting and unpacking the soil plug, the root system was carefully divided and transplanted (ensuring the root system was equally distributed) into two square (8 × 8 cm) compartments of 250 cm3 (7 cm in height) of a split-pot. Plants were grown for another two weeks, to ensure root colonisation of each compartment, in a ventilated greenhouse (day average temperature 25 °C, night average temperature 18 °C, average daytime air humidity 50%). During the photoperiod (14 hours), when natural light was below 200 μmol m−2 s−1, high-pressure sodium lamps (Son-T, Phillips, Amsterdam, The Netherlands) provided 400 μmol m−2 s−1 at bench height. For six weeks before starting the treatments, pots were weighed daily (between 17:00-18:00) and irrigated to a pre-determined target weight near to pot capacity (to avoid transient rootzone anoxia). This weight was determined by irrigating pots at the end of the photoperiod until observing drainage, with pots weighed at the beginning of the next photoperiod (to guarantee all the gravitational water drained). Evaporative losses during the night period were not considered to ensure the irrigation target was slightly lower than actual pot capacity. Irrigation was distributed evenly over the entire soil surface, before randomly assigning plants to four irrigation treatments that were applied for 21 days:
Well-watered (WW): plants were watered as in the initial period, receiving an accumulated average of 3.02 L plant−1, at 125 mL plant−1 day−1 (62.5 mL per compartment).
Fixed partial rootzone drying (PRD-F): 75% of the WW average irrigation volume was applied daily to only one compartment.
Alternate partial rootzone drying (PRD-A): Same irrigation as PRD-F, but swapping the irrigated compartment every three days, on days 4, 8, 12, 16 and 20 (Fig. 1).

Experimental schedule. For each group of plants, the timing of side alternation in alternated PRD (black drops) and gas exchange measurements (asterisks) is shown for the 21 days of treatment application
Homogeneous soil drying (HRD): 75% of the WW average irrigation volume was distributed evenly between both compartments.
Frequency of irrigation (daily) and alternation (every three days) was determined from the rate of soil drying in preliminary measurements during the week before the experiment commenced. On average, WW plants lost 30% of the soil water volume in one day. Based on a substrate moisture release curve (see next section), daily irrigation was considered adequate for well-watered plants. After three days, water loss was 65% of the initial total water in the pot (estimated from soil volumetric water content measurements, see next section) representing moderate soil drying suitable for alternating the wet and dry sides. Although field-grown plants grown with unrestricted root systems have longer alternation frequencies, containerised tomato grown under greenhouse conditions usually swapped the wet and dry sides at intervals of less than 6 days (Kirda et al. 2004; Pazzagli et al. 2016; Yang et al. 2021). Thus, the alternation frequency was appropriate for the restricted volume of each compartment. Water was slowly applied using a 60 mL plastic syringe to avoid drainage and to wet the whole surface of the compartment uniformly. No drainage was observed either upon alternation in PRD-A or after watering HRD.
Whole plant gas exchange responses to VPD (see gas exchange measurements section) were measured the day after the first, third and fifth alternation cycle, representing the entire treatment period. These time-consuming measurements (typically 2 hours per plant) allowed only one replicate of each treatment (four plants) each day. Since this would have limited replication (n = 4), three groups of 12 plants (G1, G2, G3) were allocated, with each group starting the treatments on different days to ensure the first, third and fifth days of cycle for each group did not coincide on the same day. Also, since gas exchange measurements required leaf removal to normalise against leaf area, an additional set of 12 plants were kept intact until the end of the experiment to measure stem diameter variation. To avoid overlapping target alternation days, treatments started in G1 three days before G2 and six days before G3. Gas exchange was measured in one plant per irrigation treatment of each group the day after the first, third and fifth PRD-A alternation of the irrigated side (days 5, 13 and 21 after the start of the treatments; Fig. 1). This staggered application of irrigation treatments ensured that three replicates of each could be measured at the same time relative to when the treatments started. Whole-plant gas exchange measurements started two hours after the beginning of the photoperiod and finished before the evening irrigation. Treatments were distributed within each, day ensuring that the three replicates were measured at different times of the day to minimise time-related effects (diurnal changes and soil drying) on gas exchange.
Estimating the evaporative fraction of evapotranspiration
Daily pot evapotranspiration on day n (ET) was measured as:
where PWn is pot weight measured on that day (before irrigation), PWn-1 is pot weight measured on the previous day (before irrigation) and IVn-1 the irrigation volume applied after weighing the pot the day before (assuming a water density of 1 g L−1).
Relative ET with respect to WW average (ETrel) was calculated for each pot of the deficit irrigation treatments.
To estimate the evaporation component of ET, a separate experiment utilised fifteen single compartment pots without plants, each identical to a single compartment of the split-pots used in the main experiment. They were filled with the same substrate, placed in the same environment, and distributed between three treatments to assess the effect of the different irrigation procedures applied to individual compartments.
-
WW: pots were watered to pot capacity every other day (as soil evaporative water losses over two days were similar to evapotranspirational losses from a single day in the main experiment).
-
WH: Water was withheld for nine days to replicate the irrigation procedure in the dry compartment of PRD-F, allowing soil to dry to the lowest level observed in that treatment.
-
RW: Pots were rewatered every two days but with 75% of the water applied to WW, to replicate HRD deficit irrigation and when a compartment was rewatered during PRD alternation.
Pots were weighed and soil volumetric water content (θvsensor) measured with a soil moisture dielectric probe (ML2 Thetaprobe, Delta-T, Burwell, UK) immediately before each rewatering and θv calculated as described above. Evaporation (E) was calculated as the difference in weight between two consecutive days and the relative E (Erel) compared to the average of WW was calculated for each day and pot of the WH and RW treatments. Since the θv vs Erel relationship did not differ between both WH and RW treatments, a unique sigmoid function was fitted to the data to estimate Erel from θvsensor measurements taken in plants instrumented with stem diameter sensors (see next section) before irrigation during the main experiment. Erel was estimated separately for each compartment and the average Erel was then calculated for the whole pot as the mean value of both Erel.
Soil and plant water status
All pots without stem diameter sensors were weighed daily immediately before irrigation to estimate whole-pot soil volumetric water content. For the homogeneous irrigation treatments (WW and HRD), compartments were assumed to have similar water content. In the two PRD treatments, soil volumetric water content (θvsensor) of each compartment was instantaneously measured with a Thetaprobe before and after each irrigation.
At the end of the experiment, soil moisture in both compartments of all plants was measured with the probes and the soil weighed after removing the roots. Soil was dried at 105 °C for 24 h to determine water mass in each compartment and calculate actual soil water content (θv) assuming a volume of 250 cm3 for each compartment. A linear regression model between θvsensor and θv was fitted (P < 0.001; r2 = 0.91), and the equation (θV = 2.06*θvsensor) used to transform measured θvsensor to actual θv. In plants that were not measured with the probes, whole pot volumetric and gravimetric water content was also calculated retrospectively in all treatments from daily pot weight measurements, considering dry soil, pot and plant fresh weight at harvest (plant fresh weight growth considered negligible). Only volumetric values are shown for consistency with probe measurements, but gravimetric values were used to estimate average soil water potential (Ψsoil) from θg values using a previously determined moisture release curve (Puértolas et al. 2020b).
Before the last PRD alternation (around noon) in each group, half of the plants were sealed in a plastic bag and left in the dark for at least 24 hours to ensure soil and plant water potential equilibrated. Leaf water potential at null transpiration (ΨE0) was measured as a proxy of soil water potential averaged across the entire rootzone. The remaining plants in each treatment group were bagged the following morning (or after finishing gas exchange measurements in selected plants), once PRD was alternated the evening before. Leaf area was measured in all plants after ΨE0 measurements.
Measuring stem diameter and gas exchange
A linear variable displacement transducer (LVDT) (DF/2.5, Solartron Metrology, Bognor Regis, UK) was mounted on a metal alloy frame of low thermal expansivity (invar) and attached to the stem of four plants per irrigation treatment in G1. Each LVDT was connected to a multiplexer (AM416) + datalogger (CR1000) logging system (Campbell Scientific, Logan, UT, USA), which recorded stem diameter every 30 seconds. The stem swelled during the night and sharply contracted from the beginning of the photoperiod to around midday, as lamps were activated during that period (Fig. 2). After that, stems contracted at a lower rate and even temporarily swelled again when the greenhouse construction shaded the plants. Stem diameter consistently grew after irrigation and during the night period. To assess daily stem variations on the day of PRD alternation and the day after, maximum daily contraction (MDC) was calculated as the difference between the predawn maximum diameter and the minimum before watering, while afternoon contraction (AC) was the difference between the diameter at the end of the morning supplementary light period and the minimum before watering. Since the logging system failed to record during the first two alternation cycles in G1, the third and fourth cycles were analysed. To calculate water losses in the five WW plants with LVDT sensors, they were placed on portable balances (Scout Pro, Ohaus, Parsippany, NJ, USA) to continuously display pot weight. Weight at pot saturation was recorded right after installing the sensor and placing the pot on the balance.

Example of stem diameter variation from a HRD plant between 19:00 and the same hour on the following day. Stem diameter is shown as variation with respect to the value at 19:00 on the first day (marked with a horizontal dashed line). The shaded and stripped areas denote the nocturnal and afternoon period respectively. Double-headed arrows show the maximum daily contraction (MDC), and the afternoon contraction (AC) while a single-headed arrow indicates when the plant was irrigated
Whole-plant gas exchange responses to step changes in air humidity were measured three times during the experiment in each group. For each group, one plant per treatment was measured the day after the first, third and fifth (last) alternation (Fig. 1). Initially, the whole pot and plant were placed in the custom-built whole-plant gas exchange system described previously (Jáuregui et al. 2018) with details summarised in S1:Appendix 1. The plant was acclimated for approximately 30 min to the light conditions of the chamber. Air relative humidity (RH) inside the chamber was initially set to ∼70% by passing dry air (RH = 6%; [CO2] =420 ppm) through a humidifier compartment. Then the chamber was opened (for less than 30 s) to excise a fully expanded leaf to measure leaf water potential (Ψleaf), then determine leaf area (Model Li-3100, Li-Cor Inc., Lincoln, Nebraska, USA). After closing the chamber again, gas exchange was allowed to stabilise again (typically 10 min), and then RH inside the system was reduced to 50% by introducing a mixture of dry and humidified air to the chamber. After the following RH level was achieved (typically 3 min), plant gas exchange was allowed to stabilise (typically 20-30 min). Each plant was exposed to four sequentially decreasing humidity levels achieved by increasing the ratio of dry to humid air (70%, 50%, 35%, 6%), approximately corresponding to vapour pressure deficit (VPD) values of 1, 1.5, 2 and 3 kPa (measurements taken at ambient temperature of ~25 °C). VPD levels were controlled accurately, with a maximum standard error of 0.15 kPa for each treatment and date combination. At the lowest VPD level, Ψleaf and area were again measured.
Stability of Ψsoil was assumed during the gas exchange measurements, as the total volume of water transpired was less than 10 g in well-watered plants (with θV = 0.60 cm3cm−3) and even less in drier plants (typically around 5 g for plants with θv = 0.20 cm3cm−3). Thus θv decreased by 0.02 and 0.01 cm3cm−3 respectively, corresponding to Ψsoil decreasing from −0.010 to −0.012 and from −0.70 to −0.78 MPa, respectively. Such changes were considered negligible in the context of the treatments imposed.
Net photosynthesis (An), transpiration rate (T), water use efficiency (WUE), and canopy conductance (Gc) were calculated from the difference in [CO2] and [H2O] of the air entering and exiting the chamber and the air flow as described in S1:Appendix 2.
Statistical analyses
Whole pot θv, ETrel and estimated Erel were analysed by repeated measures ANOVA. Since WW values of θv greatly exceeded the deficit irrigation treatments (P < 0.001), a second analysis included only the three deficit irrigation treatments. Treatment differences were determined by a Tukey test (P < 0.05). Maximum stem diameter contraction (MDC) and afternoon contraction (AC) were analysed by a mixed linear model, with irrigation treatment and time with respect to alternation (the day of, and after, alternation) as fixed factors and cycle (third or fourth cycle) and plant as random factors. Treatment differences were assessed by pairwise comparisons using the Bonferroni adjustment. The response of T, Gc, An, WUE, and Ψleaf to VPD on the day after alternation was analysed by a mixed model, with cycle, VPD step and treatment as fixed factors (VPD step as repeated factor), and plant as a random factor. Treatment and time with respect to alternation effects on ΨE0 were assessed by two-way ANOVA and differences between treatments with a post-hoc test (Tukey, P < 0.05). All these analyses were performed with SPSS 24 (IBM, Armonk, NY, USA). The significance of the relationship between θv and Gc measured at the second VPD level (1.5 kPa) was determined by linear regression using Sigmaplot 12.0 (Systat Software Inc., San Jose, CA, USA).
Results
Estimating the evaporative fraction of evapotranspiration
Soil volumetric water content (θv) was sigmoidally related to evaporation relative to WW plants (Erel). Relative evaporation (Erel) of pots with θv > 0.4 was close to 1, and similar to WW pots (Fig. 3). Predicted Erel fluctuated around 0.7 in PRD-A and HRD treatments, and continued to decrease in PRD-F reaching 0.5 at the end of the experiment. Then, Erel of the wet and dry compartments were ~ 0.9 and ~ 0.1 respectively (Fig. 4).

Relationship between compartment soil volumetric water content (θv) and soil evaporation relative to the average of WW compartments for pots from which water was withheld (WH) or rewatered (RW). The sigmoid function fitted for the whole dataset (no statistical differences between WH and RW) is depicted, together with its equation and the correlation coefficient

Daily estimated evaporation (E) relative to WW plants (n = 4; mean ± s.e.) from each side of the pot (side A, black circles, solid lines; Side B, hollow circles, dashed lines) in PRD-Alternated (A), PRD-Fixed (B) and HRD (C) treatments. The dashed vertical lines denote when sides were alternated in PRD-A
In PRD-A, the alternation of irrigated sides dictated fluctuations in predicted Erel, which increased the day after each alternation but progressively decreased before the next alternation (Fig. 5a). Predicted Erel was higher in HRD and PRD-A than in PRD-F. In all treatments, it started to decrease 3 to 6 days after applying treatments (two first PRD-A cycles).

Daily estimated evaporation (E, A) and evapotranspiration (ET,B) relative to WW plants, and whole pot average soil volumetric water content (θv, C) (n = 8 for A, B, n = 4 for C; mean ± s.e.). The dashed vertical lines denote when sides were alternated in PRD-A (last alternation on day 20 not shown as half of the plants were harvested before irrigation). WW: hollow triangles, HRD: solid triangles, PRD-F: hollow circles, PRD-A: solid circles
Actual relative evapotranspiration (ETrel) was lowest in HRD, with intermediate, similar values for the two PRD treatments (Table 1). Following homogenous root drying, ETrel of HRD plants initially remained close to 1 then dropped sharply 3 days after withholding water, but increased again the day after. ETrel progressively decreased in HRD, but remained consistently below the PRD treatments at 0.6 (Fig. 5b). Evapotranspiration of the PRD treatments progressively decreased during the first week, then stabilised until the end of the experiment at 0.75. Overall, PRD alternation did not affect ET, although it transiently increased the day after the third alternation.
Soil and plant water status
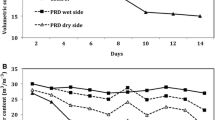
Before irrigation, whole pot θv of WW plants remained at 0.5 cm3 cm−3 (Ψsoil = −0.05 MPa) throughout the experiment. In the three deficit irrigation treatments (which were statistically similar, P = 0.55), it steadily decreased from that value to around 0.23 cm3 cm−3 (Ψsoil = −0.88 MPa) during the first ten days and remained stable thereafter (Fig. 5c). In PRD-A, θv before irrigation generally dropped to 0.2 cm3cm−3 in the dry side, recovered to 0.4 cm3cm−3 the day after irrigation, and gradually increased to 0.5 cm3cm−3 before the next change of irrigation side, as daily irrigation replenished pot capacity (Fig. 6a). Before irrigation, θv of the wet side of PRD-F was stable at 0.5 cm3cm−3 throughout the experiment. Withholding water from the dry side rapidly decreased θv to 0.3 cm3cm−3 in the first three days, then slowly decreased θv to 0.12 cm3cm−3 in the following 11 days before it stabilised (Fig. 6b). The 3 deficit treatments receiving the same irrigation volumes varied in their spatial distribution of soil moisture.

Soil volumetric water content (n = 5, error bars not shown for clarity) estimated from soil moisture probe readings in each compartment taken immediately before and after each irrigation in the two partial rootzone drying treatments (alternated (A) and fixed (B)). Side X (solid line) is the initially irrigated side (dry side after alternation in alternated PRD), while Side Y (dotted line) is the initially dry side. The dashed vertical lines denote when sides were alternated in PRD-A
Pot average θV of WW plants was always higher than in the three deficit irrigation treatments (0.57 vs 0.40 cm3cm−3). However, θv of the wettest side was significantly higher in WW and PRD-F (0.62 and 0.58 cm3cm−3) than in PRD-A and HRD (0.42 cm3cm−3 for both treatments). Water potential at null transpiration (ΨE0) was significantly (P < 0.001) higher in WW plants than the three statistically similar deficit irrigation treatments (Table 1). Neither alternation nor its interaction with treatment (P = 0.52 and 0.50 respectively) affected ΨE0. Thus, the two groups of PRD plants had similar access to soil water both before and after alternation in PRD-A.
Stem diameter and gas exchange measurements
Maximum daily contraction (MDC) was consistently around 50-100 μm with θv > 0.35 cm3cm3, and increased as average θv decreased below that threshold (Fig. 7a, b). MDC did not statistically differ (P = 0.25) between treatments, with MDC of PRD-A plants remaining low (generally below 100 μm) and not changing with alternation. Afternoon contraction (AC) followed a similar trend as MDC (Fig. 7c, d), but the treatments significantly differed (p = 0.006), with AC significantly higher in HRD than WW, while PRD treatments had intermediate values (not significantly different from HRD or WW; Table 1). Thus, stem diameter measurements in the afternoon during the period of supplementary lighting could distinguish the treatments.

Relationship between soil volumetric water content averaged across both compartments and both maximum daily contraction (A, B) and afternoon contraction (C, D) on the day of (A, C) and the day after (B, D) alternation for the three deficit irrigation treatments. HRD: solid triangles, PRD-F: hollow circles, PRD-A: solid circles. For significant regressions, P values and regression coefficient for the nonlinear (quadratic) regression fitted for each relationship are shown (P = 0.10 for regression in panel , not shown)
When measured after alternation, leaf gas exchange (T, Gc and An) was similar between WW and PRD-F plants and between PRD-A and HRD plants. These three variables were significantly higher (P = 0.006, 0.005, 0.04 respectively) in WW and PRD-F than in PRD-A and HRD treatments (Fig. 8). However, irrigation treatment did not affect Ψleaf and instantaneous water use efficiency (P = 0.13 and P = 0.69 respectively, data not shown).

Transpiration rate (A), canopy conductance (B) and net photosynthesis rate (C) for the four irrigation treatments (WW: hollow triangles, HRD: solid triangles, PRD-F: hollow circles, PRD-A: solid circles) at four incremental VPD steps (1 = 1 kPa, 2 = 1.5 kPa, 3 = 2.5 kPa, 4 = 3 kPa; s.e < 0.15 kPa). Data points represents the average of nine plants (three plants and three measurement dates; after alternation at the end of the first, third and fifth PRD-A cycles)
Canopy conductance and Ψleaf decreased with VPD, while T and WUE increased (P < 0.001 for Gc, Ψleaf, T, and WUE, Fig. 8). None of these variables (An, Gc, Ψleaf and WUE) interacted with treatment (P > 0.35) although T increased with VPD in WW and PRD-F plants more than in PRD-F and HRD plants (P = 0.02 for the Treatment x VPD interaction, Fig. 8a). Net photosynthesis increased from the first to the second VPD step change, and then decreased below the initial value from the third to the fourth VPD level (Fig. 8c).
For the deficit irrigation treatments, whole-pot average θv and Gc measured at the second VPD level (1.5 kPa, maximum difference between treatments as in Fig. 8b) were not related, as PRD-F (with higher Gc) had similar θv to the other two treatments (Fig. 9a). However, Gc significantly increased with θv of the wettest compartment, with both higher in PRD-F (Fig. 9b). Thus, fixed PRD maintained similar whole plant gas exchange to WW plants, while PRD alternation and homogenous soil drying decreased it. Greater stomatal closure caused by PRD alternation, and comparing HRD plants to fixed PRD, limited any VPD-induced increase in transpiration.

Relationship between canopy conductance (Gc) measured at the second VPD level (1.5 kPa) and soil volumetric water content (θv) averaged across the whole pot (A) and of the wettest compartment (B) for the three deficit irrigation treatments. HRD: solid triangles, PRD-F: hollow circles, PRD-A: solid circles. For B panel, P value and regression coefficient for the linear regression fitted is shown (P = 0.18 for the regression in panel A, not shown)
Discussion
Developing a miniaturised model system of PRD allowed precise whole plant measurements of photosynthesis and transpiration at controlled VPDs (Jauregui et al. 2018), while continuously measuring stem diameter as a proxy of shoot water status. Furthermore, this system readily distinguished the impacts of different irrigation treatments on both evaporative and transpiration components of ET. Variable impacts of PRD alternation on these components may account for variation in crop yield and water use efficiency (Dodd 2009; Adu et al. 2018), but most PRD studies have emphasised leaf-level physiological or whole plant agronomic responses without reconciling these effects across scales. Although the components of evapotranspiration were not directly dissected, the differences in both transpiration (measured in the whole-plant gas exchange chamber) and evaporation (measured in a dedicated experiment) suggests that PRD alternation restricts plant water use but enhances evaporative losses. These contrasting effects might explain the lack of net water savings of alternate compared with fixed application of PRD in our model system. (Table 1, Fig. 10). Since fixed PRD maintained photosynthesis relative to well-watered plants and suppressed soil evaporation to a great extent, alternating the wet and dry sides of PRD plants may be unnecessary to maximise water savings. Further field experiments should confirm that these effects are not restricted to the miniaturised crop system used here. Nevertheless, since PRD-F is less complex to operate than alternate PRD, this could facilitate greater farmer adoption. Irrespective, minimising soil evaporative losses in irrigated agriculture (e.g. through mulching) seems essential if gains in leaf-level water use efficiency (stimulated by our enhanced understanding of plant responses to heterogeneous soil drying) are to bring maximal benefits to water scarce cropping systems.

Schematic representation of the relative contribution of the components of irrigation efficiency (Net photosynthesis: An, Transpiration (T), Evaporation (E), Evapotranspiration (ET)) as affected by soil moisture (θ) distribution. Relative size of arrows and letters are proportional to the observed size of effect. For alternated PRD, effects observed after alternation. Sub-indices A and B for E represent the evaporation from either side
The presence of wet soil layers in otherwise dry rootzones (PRD) maintains transpiration compared with uniformly dry soil of the same water content (HRD). This was attributed to the maintenance of high root water potential and hence lower ABA accumulation of roots growing in heterogeneously rather than homogenously dry soil (Puértolas et al. 2017, 2020b), due to hydraulic redistribution from wet to dry roots (Bauerle et al. 2008). Both PRD-F and PRD-A treatments (with different soil moisture distributions but always a wetted part of the root system) had similar leaf water potential at null transpiration (Table 1) and during gas exchange measurements, and consequently stem diameter contractions (Fig. 7). Thus, increased root-sourced chemical signalling after PRD alternation might decrease gas exchange. Under PRD, most water uptake occurs in the irrigated rootzone, where root-sourced signals are presumably low (Puértolas et al. 2015), but alternation rapidly decreases water uptake from this side while increasing it from the formerly dry side (Pérez-Pérez and Dodd 2015). Together with increasing soil drying in the formerly wet side, this allows both sides of the root system to transiently export root-sourced signals. Although ΨE0 is lower in PRD-F than WW plants, their similar gas exchange suggests soil moisture distribution is a key variable in stomatal regulation under mild soil water deficits.
Moreover, alternating wet and dry sides during partial rootzone drying irrigation transiently homogenised soil moisture distribution compared to fixed PRD, as soil moisture of the previously dry side only partially recovered to field capacity (Fig. 6a). Thus, soil water content in both sides of PRD-A was lower than 0.5 cm3cm−3 while the wet side of PRD-F was above that threshold, allowing higher stomatal conductance of PRD-F than PRD-A plants (Fig. 8b). Thus, continuously sustaining irrigation to part of the rootzone while withholding irrigation from the other could maintain whole plant gas exchange and biomass accumulation. Treatment differences in whole-plant transpiration occurred after PRD alternation (higher in WW and PRD-F than HRD and PRD-A - Fig. 8), but all deficit irrigation treatments had lower ET than WW plants, especially the HRD treatment that was lower than both PRD treatments (Fig. 5b). The effect of irrigation placement on evaporation (E) also partly explained variation in ET. Irrigating the whole pot surface in HRD was estimated to increase E compared to irrigating only half of the pot, which might have attenuated treatment differences in ET caused by the low T of HRD than PRD-F. Interestingly, alternation in PRD-A transiently increased E (Fig. 5a) and maintained it higher than PRD-F. This might explain the similar ET of both treatments, even though PRD-F had higher T than PRD-A after alternation (Fig. 10). While the presence of a wet part within the rootzone with heterogeneous soil drying (PRD) enhanced plant transpiration, evaporation from the non-irrigated side of the rootzone was restricted.
Although the three deficit treatments had similar overall soil moisture (Fig. 5c), the more homogeneous soil moisture distribution across the entire rootzone in HRD and PRD-A plants maintained higher overall evaporation than PRD-F. Similar results occurred when comparing PRD with sustained deficit irrigation (DI), and between different dripper distributions under DI (Marsal et al. 2008). The temporal dynamics of E in PRD-A supports a positive relationship between soil moisture homogeneity and evaporation. Alternation tended to equalise soil moisture in both compartments (Fig. 6a), which transiently (for one day) increased Erel (Fig. 5a). Between alternation events, as θv increasingly differed between compartments, predicted Erel decreased since soil water content was non-linearly related to evaporation (Fig. 3, Wythers et al. 1999). Thus, soil evaporation in each side of treatments with homogeneously moderately dry rootzone (HRD and PRD-A) is only slightly lower than the wet side of PRD-F, but much higher than in the dry side (Fig. 4), thereby resulting in higher overall evaporation than in PRD-F (Fig. 5a). Thus, both irrigation placement and timing affected not only plant water use but also soil evaporation, which determines irrigation efficiency.
Although canopy shading of the soil surface may modify the relationship between θv and E by decreasing E, this relationship should still be proportional to the wetted soil volume. Thus, techniques that lower soil evaporation (such as sub-surface irrigation or soil cover) could maximise water savings during deficit irrigation (Abdelraouf and Ragab 2018; Gil et al. 2018; Iqbal et al. 2021). However, applying PRD-F when the irrigated side is covered to minimise soil evaporation might be the most effective technique, as it does not reduce Gc and An either transiently (PRD-A) or consistently (HRD) (Fig. 8). In some production systems, covering the soil of only the irrigated side of the rootzone could save time and money while maintaining high irrigation efficiency. However, in field experiments comparing fixed and alternate application of PRD, the latter consistently outyielded the former (Dodd et al. 2015 and references therein). Daily irrigation of our experiment was unable to maintain optimal water content of the wet side of PRD plants. Since soil moisture levels of the irrigated side explain physiological responses to PRD (Dodd et al. 2008a), more homogeneous water uptake between sides of PRD-A plants (the dry side does not dry as much as in fixed PRD) might explain some favourable results for alternate PRD (Kang et al. 2001). However, it also suggests that effective PRD application should consider an adequate frequency of irrigation.
Irrigation regime determined overall whole-plant gas exchange, but not its response to evaporative demand. Soil moisture content or distribution did not seem to affect stomatal sensitivity to VPD (Fig. 8). Hydraulic models indicate that stomatal conductance is controlled only by changes in leaf water potential, which in turn, depends on both soil water potential and transpiration rate (Buckley 2019). As stomatal aperture also controls transpiration rate, this configures a feedback model that predicts greater differences in Gc between plants with contrasting soil water potential at low VPD levels, which would decrease as VPD increases (Sperry et al. 2017). This occurs because limited water loss at low VPD levels, and differences in Gc, reflect differences in soil water potential. As the VPD increases, plant transpiration in drier soil will be more limited than in wetter soil, inducing proportionally greater stomatal closure. In our experiment, this difference in Gc was similar across the whole VPD range tested (1-3 kPa, Fig. 8b), reinforcing that observed differences in gas exchange in response to the moderate water deficits elicited by these irrigation treatments are not associated to hydraulic constraints. Thus root-sourced signals regulate differences in Gc at low VPD, with increasing VPD enhancing transpiration and decreasing leaf water potential equally across all the treatments, reducing GC similarly.
Conclusions
Hypothesis 1, that alternation would impose hydraulic limitations that increase stem diameter contractions and reduce Gc and An, was not sustained. Instead, alternation transiently decreased Gc and An coincident with the homogenisation of rootzone soil moisture, caused by the disappearance of the wet (close to field capacity) region within the rootzone. Decreasing soil moisture heterogeneity transiently increases fluxes of root-sourced chemical signals to the shoot to elicit stomata closure (Dodd et al. 2008b; Puértolas et al. 2016). However, homogenising soil moisture across the rootzone also transiently increased soil evaporation, confirming Hypothesis 2, thereby offsetting the decreased Gc and resulting in similar crop water losses (Fig. 10). While partial rootzone drying already minimises soil evaporation by irrigating only half the rootzone, further water savings could be achieved by reducing evaporative losses from irrigated areas (by covering them or using sub-surface irrigation) when applying PRD. Since applying PRD-F and covering only the irrigated half of the rootzone could minimise the cost of soil cover, further field trials are needed to empirically assess these concepts at different irrigation frequencies, and the consequences for irrigation efficiency.
Change history
17 August 2022
A Correction to this paper has been published: https://doi.org/10.1007/s11104-022-05653-5
References
Abdelraouf RE, Ragab R (2018) Applying partial root drying drip irrigation in the presence of organic mulching. Is that the best irrigation practice for arid regions? Field and modelling study using the saltmed model. Irrig Drain 67:491–507. https://doi.org/10.1002/IRD.2249
Adu MO, Yawson DO, Armah FA et al (2018) Meta-analysis of crop yields of full, deficit, and partial root-zone drying irrigation. Agric Water Manag 197:79–90. https://doi.org/10.1016/j.agwat.2017.11.019
Bauerle TL, Richards JH, Smart DR, Eissenstat DM (2008) Importance of internal hydraulic redistribution for prolonging the lifespan of roots in dry soil. Plant Cell Environ 31:177–186. https://doi.org/10.1111/j.1365-3040.2007.01749.x
Buckley TN (2019) How do stomata respond to water status? New Phytol 224:21–36. https://doi.org/10.1111/nph.15899
Consoli S, Stagno F, Roccuzzo G et al (2014) Sustainable management of limited water resources in a young orange orchard. Agric Water Manag 132:60–68. https://doi.org/10.1016/J.AGWAT.2013.10.006
Dodd IC (2009) Rhizosphere manipulations to maximize “crop per drop” during deficit irrigation. J Exp Bot 60:2454–2459. https://doi.org/10.1093/jxb/erp192
Dodd IC, Theobald JC, Bacon MA, Davies WJ (2006) Alternation of wet and dry sides during partial rootzone drying irrigation alters root-to-shoot signalling of abscisic acid. Funct Plant Biol 33:1081–1089. https://doi.org/10.1071/Fp06203
Dodd IC, Egea G, Davies WJ (2008a) Accounting for sap flow from different parts of the root system improves the prediction of xylem ABA concentration in plants grown with heterogeneous soil moisture. J Exp Bot 59:4083–4093. https://doi.org/10.1093/Jxb/Ern246
Dodd IC, Egea G, Davies WJ (2008b) Abscisic acid signalling when soil moisture is heterogeneous: decreased photoperiod sap flow from drying roots limits abscisic acid export to the shoots. Plant Cell Environ 31:1263–1274. https://doi.org/10.1111/j.1365-3040.2008.01831.x
Dodd IC, Puertolas J, Huber K et al (2015) The importance of soil drying and re-wetting in crop phytohormonal and nutritional responses to deficit irrigation. J Exp Bot 66:2239–2252. https://doi.org/10.1093/jxb/eru532
Dry PR, Loveys BR (1999) Grapevine shoot growth and stomatal conductance are reduced when part of the root system is dried. Vitis 38:151–156
Dry PR, Loveys BR, During H (2000) Partial drying of the rootzone of grape. I. Transient changes in shoot growth and gas exchange. Vitis 39:3–7
Du TS, Kang SZ, Zhang JH, Li FS, Hu XT (2006) Yield and physiological responses of cotton to partial root-zone irrigation in the oasis field of Northwest China. Agric Water Manag 84:41–52. https://doi.org/10.1016/j.agwat.2006.01.010
During H, Loveys BR, Dry PR (1996) Root signals affect water use efficiency and shoot growth. Acta Hortic:1–13
Egea G, Dodd IC, Gonzalez-Real MM et al (2011) Partial rootzone drying improves almond tree leaf-level water use efficiency and afternoon water status compared with regulated deficit irrigation. Funct Plant Biol 38:372–385. https://doi.org/10.1071/Fp10247
Elsayed-Farag S, Melgar JC (2015) Efficiency of tree-based water status indicators at the onset of water deficit in citrus. Hortic Environ Biotechnol 56:305–309. https://doi.org/10.1007/S13580-015-0136-8
Fereres E, Soriano MA (2007) Deficit irrigation for reducing agricultural water use. J Exp Bot 58:147–159. https://doi.org/10.1093/jxb/erl165
Fernández JE, Cuevas MV (2010) Irrigation scheduling from stem diameter variations: a review. Agric For Meteorol 150:135–151. https://doi.org/10.1016/j.agrformet.2009.11.006
Gil PM, Lobos P, Durán K et al (2018) Partial root-zone drying irrigation, shading, or mulching effects on water savings, productivity and quality of 'syrah’ grapevines. Sci Hortic 240:478–483. https://doi.org/10.1016/J.SCIENTA.2018.06.050
Hatfield JL, Dold C (2019) Water-use efficiency: advances and challenges in a changing climate. Front Plant Sci 19. https://doi.org/10.3389/fpls.2019.00103
Huber K, Vanderborght J, Javaux M et al (2014) Modelling the impact of heterogeneous rootzone water distribution on the regulation of transpiration by hormone transport and/or hydraulic pressures. Plant Soil 384:93–112. https://doi.org/10.1007/s11104-014-2188-4
Huber K, Vanderborght J, Javaux M, Vereecken H (2015) Simulating transpiration and leaf water relations in response to heterogeneous soil moisture and different stomatal control mechanisms. Plant Soil 394:109–126. https://doi.org/10.1007/s11104-015-2502-9
Iqbal R, Habib-ur-Rahman M, Raza MAS et al (2021) Assessing the potential of partial root zone drying and mulching for improving the productivity of cotton under arid climate. Environ Sci Pollut Res 28:66223–66241. https://doi.org/10.1007/S11356-021-15259-6
Jauregui I, Rothwell SA, Taylor SH, Parry MAJ, Carmo-Silva E, Dodd IC (2018) Whole plant chamber to examine sensitivity of cereal gas exchange to changes in evaporative demand. Plant Methods 14(1). https://doi.org/10.1186/s13007-018-0357-9
Kang SZ, Liang ZS, Hu W, Zhang JH (1998) Water use efficiency of controlled alternate irrigation on root-divided maize plants. Agric Water Manag 38:69–76. https://doi.org/10.1016/s0378-3774(98)00048-1
Kang SZ, Zhang L, Hu X, Li Z, Jerie P (2001) An improved water use efficiency for hot pepper grown under controlled alternate drip irrigation on partial roots. Sci Hort 89:257–267. https://doi.org/10.1016/S0304-4238(00)00245-4
Kang SZ, Hu X, Jerie P, Zhang JH (2003) The effects of partial rootzone drying on root, trunk sap flow and water balance in an irrigated pear (Pyrus communis L.) orchard. J Hydrol 280:192–206. https://doi.org/10.1016/s0022-1694(03)00226-9
Kirda C, Cetin M, Dasgan Y et al (2004) Yield response of greenhouse grown tomato to partial root drying and conventional deficit irrigation. Agric Water Manag 69:191–201. https://doi.org/10.1016/J.AGWAT.2004.04.008
Klepper B, Browning VD, Taylor HM (1971) Stem diameter in relation to plant water status. Plant Physiol 48:683. https://doi.org/10.1104/pp.48.6.683
Leib BG, Caspari HW, Redulla CA et al (2006) Partial rootzone drying and deficit irrigation of “Fuji” apples in a semi-arid climate. Irrig Sci 24:85–99. https://doi.org/10.1007/s00271-005-0013-9
Liu FL, Shahnazari A, Andersen MN et al (2006) Physiological responses of potato (Solanum tuberosum L.) to partial root-zone drying: ABA signalling, leaf gas exchange, and water use efficiency. J Exp Bot 57:3727–3735. https://doi.org/10.1093/jxb/erl131
Marsal J, Mata M, Del Campo J et al (2008) Evaluation of partial root-zone drying for potential field use as a deficit irrigation technique in commercial vineyards according to two different pipeline layouts. Irrig Sci 26:347–356. https://doi.org/10.1007/S00271-007-0098-4
McLean EH, Ludwig M, Grierson PF (2011) Root hydraulic conductance and aquaporin abundance respond rapidly to partial root-zone drying events in a riparian Melaleuca species. New Phytol 192:664–675. https://doi.org/10.1111/j.1469-8137.2011.03834.x
Pazzagli PT, Weiner J, Liu F (2016) Effects of CO2 elevation and irrigation regimes on leaf gas exchange, plant water relations, and water use efficiency of two tomato cultivars. Agric Water Manag 169:26–33. https://doi.org/10.1016/J.AGWAT.2016.02.015
Pérez-Pérez JG, Dodd IC (2015) Sap fluxes from different parts of the rootzone modulate xylem ABA concentration during partial rootzone drying and re-wetting. J Exp Bot 66:2315–2324. https://doi.org/10.1093/jxb/erv029
Pérez-Pérez JG, Navarro JM, Robles JM, Dodd IC (2018) Prolonged drying cycles stimulate ABA accumulation in Citrus macrophylla seedlings exposed to partial rootzone drying. Agric Water Manag 210:271–278. https://doi.org/10.1016/j.agwat.2018.08.020
Pérez-Pérez JG, Puértolas J, Albacete A, Dodd IC (2020) Alternation of wet and dry sides during partial rootzone drying irrigation enhances leaf ethylene evolution. Environ Exp Bot 176:104095. https://doi.org/10.1016/j.envexbot.2020.104095
Puértolas J, Conesa MR, Ballester C, Dodd IC (2015) Local root abscisic acid (ABA) accumulation depends on the spatial distribution of soil moisture in potato: implications for ABA signalling under heterogeneous soil drying. J Exp Bot 66:2325–2334. https://doi.org/10.1093/jxb/eru501
Puértolas J, Conesa MC, Dodd IC (2016) An empirical model predicting xylem sap ABA concentration from root biomass and soil moisture distribution in plants under partial root-zone drying. Acta Hortic 1112:147–153
Puértolas J, Larsen EK, Davies WJ, Dodd IC (2017) Applying “drought” to potted plants by maintaining suboptimal soil moisture improves plant water relations. J Exp Bot 68:2413–2424. https://doi.org/10.1093/jxb/erx116
Puértolas J, Albacete A, Dodd IC (2020a) Irrigation frequency transiently alters whole plant gas exchange, water and hormone status, but irrigation volume determines cumulative growth in two herbaceous crops. Environ Exp Bot 176:104101. https://doi.org/10.1016/J.ENVEXPBOT.2020.104101
Puértolas J, Pardos M, de Ollas C et al (2020b) Soil moisture heterogeneity regulates water use in Populus nigra L. by altering root and xylem sap phytohormone concentrations. Tree Physiol 40:762–773. https://doi.org/10.1093/treephys/tpaa037
Sebastian B, Lissarrague JR, Santesteban LG et al (2016) Effect of irrigation frequency and water distribution pattern on leaf gas exchange of cv. ‘Syrah’ grown on a clay soil at two levels of water availability. Agric Water Manag 177:410–418. https://doi.org/10.1016/J.AGWAT.2016.08.032
Sobeih WY, Dodd IC, Bacon MA et al (2004) Long-distance signals regulating stomatal conductance and leaf growth in tomato (Lycopersicon esculentum) plants subjected to partial root-zone drying. J Exp Bot 55:2353–2363. https://doi.org/10.1093/Jxb/Erh204
Sperry JS, Venturas MD, Anderegg WRL et al (2017) Predicting stomatal responses to the environment from the optimization of photosynthetic gain and hydraulic cost. Plant Cell Environ 40:816–830. https://doi.org/10.1111/pce.12852
Stoll M, Loveys B, Dry P (2000) Hormonal changes induced by partial rootzone drying of irrigated grapevine. J Exp Bot 51:1627–1634. https://doi.org/10.1093/jexbot/51.350.1627
Tang LS, Li Y, Zhang J (2005) Physiological and yield responses of cotton under partial rootzone irrigation. Field Crops Res 94:214–223. https://doi.org/10.1016/j.fcr.2005.01.005
Tardieu F (2016) Too many partners in root shoot signals. Does hydraulics qualify as the only signal that feeds back over time for reliable stomatal control? New Phytol 212:802–804. https://doi.org/10.1111/nph.14292
Thompson AJ, Andrews J, Mulholland BJ et al (2007) Overproduction of abscisic acid in tomato increases transpiration efficiency and root hydraulic conductivity and influences leaf expansion. Plant Physiol 143:1905–1917. https://doi.org/10.1104/pp.106.093559
Wythers KR, Lauenroth WK, Paruelo JM (1999) Bare-soil evaporation under semiarid field conditions. Soil Sci Soc Am J 63:1341–1349. https://doi.org/10.2136/sssaj1999.6351341x
Yang X, Bornø ML, Wei Z, Liu F (2021) Combined effect of partial root drying and elevated atmospheric CO2 on the physiology and fruit quality of two genotypes of tomato plants with contrasting endogenous ABA levels. Agric Water Manag 254:106987. https://doi.org/10.1016/J.AGWAT.2021.106987
Acknowledgements
We are grateful to Prof. Pilar Pita from Universidad Politécnica de Madrid for lending us the LVDT sensors and logging system. We also thank Prof. Bill Davies for critically reviewing the manuscript. The research was funded by the European Union through the project: “Managing water scarcity in European and Chinese crop systems” (SHui; Project Number: 773903) and by the Global Challenge Research Funding (Project RECIRCULATE, ES/P010857/1).
Funding
Open Access funding provided thanks to the CRUE-CSIC agreement with Springer Nature. This research was funded by the European Union through the project: “Managing water scarcity in European and Chinese crop systems” (SHui; Project Number: 773903) and by the Global Challenge Research Funding (Project RECIRCULATE, ES/P010857/1).
Author information
Authors and Affiliations
Contributions
Both authors contributed to the study conception and experimental design. Material preparation, experimental execution, and data collection and analysis were performed by JP, who also wrote the first draft of the manuscript. ID commented on several versions of the manuscript and both authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors have no relevant financial or non-financial interests to disclose.
Additional information
Responsible Editor: Rafael S. Oliveira.
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
ESM 1
(DOCX 16 kb)
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Puértolas, J., Dodd, I.C. Evaluating soil evaporation and transpiration responses to alternate partial rootzone drying to minimise water losses. Plant Soil 480, 473–489 (2022). https://doi.org/10.1007/s11104-022-05594-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-022-05594-z