
Abstract
Auraptene and umbelliprenin are among the most abundant naturally occurring prenyloxy umbelliferone derivatives. Both have been mainly isolated from plants belonging to numerous genera of the Rutaceae (in particular auraptene in Citrus spp.) and Apiaceae (in particular umbelliprenin in Ferula spp.) families, comprising different and widely used medicinal plants and in general plants having beneficial effects to human welfare as well as edible fruits and vegetables. Although known for quite a long time (nearly a century for auraptene and 50 years for umbelliprenin), only in the last two decades the two title natural compounds were revealed to possess valuable and promising pharmacological properties as dietary feeding active cancer chemopreventive, anti-bacterial, anti-protozoal, anti-fungal, anti-inflammatory, neuroprotective, and anti-oxidant agents among the activities best detailed in the recent literature. The aim of this comprehensive review is to outline in detail the effects described in the very last years for auraptene and umbelliprenin and what has been reported about the mechanisms of action underlying the observed pharmacological activities of these oxyprenylated secondary metabolites. In view of the herein described data suggestions on how to address the future research about both natural products in the best ways according to Authors will be also provided.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
During the last decade rare natural products have been subject of intensive research with the objective to better depict their pharmacological profile (Xie et al. 2015). Thanks to these new findings, it can be hypothesized how the observed activities of phytopreparations from medicinal and beneficial plants are indeed due to not only to the main and nowadays fashioned phytochemicals (e.g. polyphenols, terpenes, glycosides, etc.), but to a synergy of action between the most and the less abundant components of phytocomplexes. Notable and recent examples to this concern are represented by extracts of Gingko biloba L. (Fam. Gingkoaceae) (Cheng et al. 2020), of Allium fistulosum L. (Amaryllidaceae) (Zolfaghari et al. 2020), of Anneslea fragrans Wall. (Fam. Pentaphylacaceae) (Omar et al. 2019), and of Trigonostemon heterophyllus Merr. (Fam. Euphorbiaceae) (Xi et al. 2019), of Wasabia japonica (Miq.) Matsum (Fam. Brassicaceae) (Kim et al. 2018), among numerous others cited in the literature of the last 5 years. Some interesting reviews on natural products containing rare skeletons and their plant sources have also been reported (Petkowski et al. 2019; Chan and O’Hagan 2012).
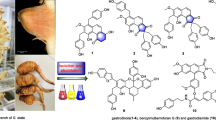
Among the best investigated classes of rare natural products during the last two decades are the oxyprenylated secondary metabolites of plant, fungal, and microbial origin. These include compounds possessing different skeletons like phenylpropanoids, polyketides, and alkaloids. The first group have been subject of more attention for what concerns the phytochemical and pharmacological properties. What was put in evidence for oxyprenylated phenylpropanoids is mainly their efficacy to act as chemopreventive agents of acute and chronic syndromes affecting humans like cancer, inflammation, stroke, neurological disorders, and several others (Fiorito et al. 2019a). The most part of review and research articles appeared in the literature in recent years focused on two oxyprenylated coumarins, both derivatives of the widespread umbelliferone (1). These were auraptene (2,7-geranyloxycoumarin) and umbelliprenin (3,7-farnesyloxycoumarin) (Fig. 1).

Structures of umbelliferone (1), auraptene (2), and umbelliprenin (3)
Data collected in this comprehensive review were taken from the main Internet databases like Scopus, Web of Science, Pubmed, and Google Scholar using the corresponding keyword as the bibliographic search parameter. Analysis of the literature documents indicated that, from the publication of the last review article in August 2019 by Bibak et al., 19 research articles have been reported, while for umbelliprenin from the publication of the last review article covering the widest range of biological activities of this oxyfarnesylated coumarin by Shakeri et al. (2014), 35 research articles appeared.
The aim of the present manuscript is to make a detailed survey about the effects and underlying mechanisms of action reported in the literature covering the very last years for auraptene (2) and umbelliprenin (3). Data outlined herein will be also tentative suggestions to the overall scientific community working in the field on how to stimulate and address future research activities employing both oxyprenylated coumarins in the best ways from phytochemical and pharmacological points of view.
Auraptene
Auraptene (2) is the most abundant geranyloxycoumarin found in nature and its natural sources have been reviewed in 2011 for the first time (Genovese and Epifano 2011). In more recent years its presence has been also documented in the following plants: grapefruit (Citrus x paradisi Macfad) peels (Genovese et al. 2020), Ferula assa-fetida L. rhizome dried latex (Eidi et al. 2020), and pomegranate (Punica granatum L.) seeds (Fiorito et al. 2019b), and Eremurus persicus (Joub & Spach) Boiss crude extract of the whole plant (Mottaghipisheh et al. 2020). Although few, papers on the analysis of auraptene from novel plant sources appeared across the end of 2019 and the mid of 2020 have all substantial elements of novelty. Papers about grapefruit peels and asafetida represent in absolute the first applications of new extractive and analytical techniques for the determination of auraptene and oxyprenylated phenylpropanoids in general: the subcritical butane extraction coupled to UHPLC analysis and the dummy molecularly imprinted solid phase extraction method, respectively. Both methodologies provided significant improvements and represent valid alternatives respect to the state of the art on auraptene extraction from its most common natural sources. These mainly consist in improved extraction yields, shorter operational times, and the use of cheap and easy to build and to handle apparatus. This is particularly true in the case of subcritical butane extraction that was accomplished by means of a handmade and very economic apparatus that proved to be an extraction methodology able to preserve the chemical stability of auraptene respect to heat-based processes like Soxhlet extraction and ultrasound- and microwave-promoted maceration. On the other hand, the paper about pomegranate by Fiorito et al. is the first record of the presence of an oxyprenylated secondary metabolite in the Punicaceae family.
For what concerns novel biological activities exerted by auraptene both in vitro and in vivo, recent studies enforced the role of this natural coumarin as a neuroprotective agent. Jang et al. deepened the knowledge about the interaction of auraptene with mithochondrial functionality both in cultured cell systems and in vivo model of Parkinson’s disease (Jang et al. 2019). First, the protective effects by auraptene against rotenone- and 1-methyl-4-phenylpiridinium (MPP+)-induced cell death in dopaminergic SN4741 neurons were assessed. While cells not treated with compound (2) exhibited a death pattern in a dose-dependent manner for both rotenone and MPP+, those pre-treated with auraptene, applied in a concentration range between 0.5 and 10 μM, became resistant to neurotoxicity. It is also noteworthy that auraptene had no effects on viability when administered as an individual compound to cells. In the meantime, it provided an about 2.5-fold decrease of the effect of rotenone 0.25 μM on mitochondrial oxygen consumption rate after 24 h (from 67.8 to 24.1%). Furthermore, treatment with auraptene in the same cell line induced the expression of several antioxidant enzymes, thus diminishing the production of reactive oxygen species (ROS) (21.6%). In in vivo experiments, Jang et al. also assessed the extent of the expression of tyrosine hydroxylase in the striatum, the reduction of which is associated to a severe progression of Parkinson’s disease. So, when administered intraperitoneally at a dose of 25 mg/kg to B6 mice 1 day before and 2 days after treatment with the two neurotoxins, auraptene was able to increase (32%) the numbers of neurons expressing this enzyme. In parallel also the astrocyte activation was diminished. As the final part of their study, Jang et al. determined how auraptene was also able to ameliorate in an appreciable manner the overall movements and related coordination of animals exposed to rotenone and MPP+.
In the same year Furukawa et al. used rat C6 glioma cells to investigate their release of the glial cell line-derived neurotrophic factor (GDNF) (Furukawa et al. 2019). After having assessed that auraptene did not affect cell viability up to 80 μM, these Authors incubated cells with this coumarin at the concentration of 10 μM for 60 h. GDNF release was induced along this period in a time- and dose-dependent manner. The same pattern was observed for the expression of GDNF mRNA. As a continuation of their studies, Furukawa et al. (2020) determined also that auraptene had a deep effect on the release and expression of the brain-derived neurotrophic factor (BDNF) in mouse neuroblastoma 2a cells (Furukawa et al. 2020).
Both studies clearly indicate that auraptene exert significant effects at the central nervous system (CNS) level, thus having a great potential as a neuroprotective agent. The main target of this geranyloxycoumarin appear to be neuronal mitochondria although more studies are needed to get further insights to this concern.
Properties of auraptene to modulate the production of ROS has been better investigated by Afshari et al. (2019). These Authors observed a dual behavior in cultured glioblastoma multiforme (GBM) cells. When incubated with compound (2) at concentrations of 50, 100, and 200 μM, the ROS production until 6 h diminished respect to controls of about 50% while after 8 h the same tended to increase around 40% more respect to controls. This action was completely reversed by the application of N-acetylcysteine (NAC). In the same paper, Afshari et al. also measured the apoptotic levels in the same cell line. qRT-PCR analysis revealed an up-regulation of p21, CXCL3, and a down-regulation of cyclin D1 genes expression and Western blot one recorded an up-regulation of the Bax/Bcl-2 ratio protein levels in a dose-dependent manner. These findings allowed these Authors to hypothesize that the mechanisms of auraptene-evoked GBM cells toxicity and induction of apoptosis is mediated by the Bax/Bcl-2 modulation and genes expression. However, Afshari et al. in their manuscript did not cite any hypothesis about the dual contrasting behavior of compound (2) respect to the production of ROS. This gap was in part filled by Askari et al. (2020). These Authors put in evidence how auraptene, in the concentration range 10–90 μM, in phytohemagglutinin-stimulated human lymphocytes markedly decreased the release of ROS and malondialdehyde (MDA), and the secretion of interleukin 6 (IL-6) and tumor necrosis factor (TNF)-α, while increasing the content of glutathione (GSH) and activities of superoxido dismutase (SOD) and catalase (CAT) (Askari et al. 2020).
Salari et al. found that auraptene can be considered also an effective adjuvant agent in the radiotherapy of cancer (Salari et al. 2020). When assayed in combination with ionizing radiations in cultured CT26 cells, auraptene provided a large increase in the number of apoptotic cells. As a parallelism, when administered to mice affected by malignant forms of colon carcinoma exposed to the same radiations, compound (2) recorded a significant decrease in tumor masses and in some case complete regression of colon cancer. Auraptene was able to down regulate the expression of Cyclin D1 and CD44 with involvement of PI3K-AKT-mTORC signaling pathway and caspase-3.
A similar synergy of action by auraptene with commonly used cancer chemotherapeutics (e.g. reduction of tumor masses in cultured cell models of human breast cancer) was observed by Gkionis et al. (2020).
Four further biological targets and associated mechanisms of action for auraptene have been determined in last months. As many other coumarins, also compound (2) was already known to inhibit platelet aggregation (Teng et al. 1992), but the effective mechanism underlying this effect remained largely unknown. Hsia et al. provided a great insight to this concern. These Authors observed that auraptene was able to selectively inhibit platelet aggregation in human platelets stimulated with collagen in the concentration range 10–100 μM and an IC50 = 35 μM. Moreover, it had no effect in inhibiting platelet aggregation stimulated with thrombin, arachidonic acid, and U46619. Auraptene also evoked ATP release, intracellular Ca+2 mobilization up to 75% at a concentration of 50 μM, P-selectin gene expression induced by collagen by around 65%, and blocked PAC-1 binding to integrin αIIbβ3. Auraptene at concentrations of 35 and 50 μM inhibited also the phosphorylation of some enzymes of the Src-family kinases, namelyLyn, Fyn, and Syk, phospholipase Cγ2 (PLCγ2), protein kinase C (PKC), Akt, mitogen-activated protein kinases (MAPKs; extracellular-signal-regulated kinase(ERK1/2), and c-Jun N-terminal kinase (JNK1/2). Auraptene was also tested in vivo by these Authors as a preventing agent of thrombosis associated to acute pulmonary embolism and related mortality. When administered to mice at 7.5 mg/kg and 15 mg/kg this latter was reduced to 25.0% and 62.5% respectively (Hsia et al. 2019).
Also the interaction of auraptene and the farnesoid X receptor, first put in evidence Epifano et al. (2012), was further characterized studying the hepatoprotective action by auraptene against 17α-ethinylestradiol (EE)-induced cholestasis both in vitro using mice primary hepatocytes and in vivo in C57BL/6 mice (Wang et al. 2019). These Authors first determined that compound (2) can modulate FXR target genes involved in the bile acid transport and synthesis. When applied at concentrations of 2.5, 5, and 10 μM for 24 h, auraptene increased the gene expression levels of bile salt export pump (Bsep) and multidrug resistance-associated protein 2 (Mrp2), and small heterodimeric partner (Shp), while slightly decreased the one of Na+-taurocholate cotransporting polypeptide (Ntcp), of the cholesterol 7α-hydroxylase (Cyp7a1),and sterol-12α-hydroxylase (Cyp8b1),all in a dose-dependent manner. When assayed in vivo upon administration for 7 days at doses of 5, 10, and 20 mg/kg, auraptene prevented body weight decrease associated to EE-induced cholestasis, levels of blood markers like alkaline phoshpatase (ALP) and total bile acids, while not modifying other liver function indicators like transaminases and bilirubin. It also ameliorated bile flow and increased bile acids output both suppressed by EE. In the meantime, auraptene diminished the development of inflammatory processes in liver tissues. The confirmation of the involvement of the FXR in the beneficial effects for the liver functions associated to administration of auraptene, came from the acquisition that all the changes evoked by compound (2) were antagonized by guggulsterone, a well-known antagonist of the FXR.
With the aim to get insights into its neuroprotective mechanism of action, Erdogan Orhan et al. (2019) studied in vitro and in silico the interaction of auraptene with cholinesterases. These Authors, using a concentration value of 100 µM in isolated enzyme assays, found virtually no inhibition on acetyl cholinesterase, while compound (2) was found to moderately and selectively inhibit butyryl cholinesterase (59.2% inhibition). In the same studies Erdogan Orhan et al. provided for the first time some key chemico-physical parameters about auraptene like LogP, blood brain barrier permeation, and plasma protein binding, even if only as a result of docking studies calculations.
Finally, Nabekura et al. studied in detail the interaction of auraptene with glycoprotein P. This membrane carrier, expressed in numerous organs and tissue of human body, is well known as the main responsible of the therapeutic inefficacy of several clinically important drugs like verapamil, digoxin, vinblastine, and others (Elmeliegy et al. 2020). These Authors investigated the interaction of auraptene with glycoprotein P in human intestinal epithelial LS174T cells and a reporter plasmid expressing 10.2 kbp of the upstream regulatory region of MDR1 (Nabekura et al. 2020). At the concentration of 50 µM, compound (2) induced MDR1 promoter activity in LS174T cells. Furthermore, the overexpression of the nuclear receptor human pregnane X receptor gene (NR1I2) favoured the auraptene-induced MDR1 activation. Nuclear factor-kappa B inhibitors, Bay11-7082 and JSH-23, largely decreased such an activation. Western blot analyses revealed that auraptene led to the induction of P-glycoprotein expression in treated LS174T cells. Nabekura et al. concluded that auraptene is able to induce the drug efflux transporter P-glycoprotein in human intestinal cells, and thus has the potential to cause food-drug interactions, also as a part of Citrus phytocomplexes.
These last four papers are of great importance as they greatly deepened the knowledge about already acquired knowledge about auraptene effects providing sharp details into its mechanisms of action.
In an animal model of polycystic ovary syndrome (PCOS), one of the most common endocrine-metabolic disorders, Abizadeh et al. (2020) reported some interesting effects following subcutaneous administration of auraptene to prepubertal naval medical research institute (NMRI) female mice. First, these Authors confirmed the inhibitory effect on TNF-α release and an increase of intracellular GSH levels confirming the findings by Askari et al. (2020) in another cell line and recorded an increase in caspase-3 expression and activity. These beneficial effects resulted also in an amelioration of oocytes maturation and fertilization (more than 30%) and subsequent more normalized embryo and blastocyst formation. This study is a confirmation of the role of auraptene in modulating the endocrine system due to its capacity to interact with hormonal receptors, in particular with estrogen ß-receptor acting as a partial agonist with an IC50 = 7.0 µM (De Medina et al. 2010).
Other notable properties of auraptene, like the anti-inflammatory and the anti-bacterial activities have been recently exhaustively reviewed by Fiorito et al. (2018a, b).
The one reported in 2020 by Jalilzadeh et al. is among the first examples of application to auraptene of pharmaceutical technology processes. These Authors successfully accomplished the synthesis of nanoencapsulated particles containing compound (2) with biodegradable copolymers, namely triblock (TB) (PCL-PEG-PCL) and pentablock (PB) (PLA-PCL-PEG-PCL-PLA) ones (Jalilzadeh et al. 2020). They tested the obtained devices as anticancer formulations on cultured HT-29 human colon cancer cells. Over a period of 120 h a very small release of the active principles from nanoparticles occurred (16.74% from PB and 14.29 from TB). The cellular uptake over 24 h incubation reached 76% for TB and 89% for PB. The cytoxicity respect to untreated controls were sixfold higher for free auraptene, but 9- and 13-fold higher respectively for TB and PB. This study is very interesting and indicates how biodegradable nanovehicles for sustained delivery of auraptene have a great potential for treatment of patients affected by different types of cancers.
The first randomized, placebo-controlled, double-blind study in healthy volunteers using auraptene enriched-extracts of peels of Citrus kawachiensis Hort. ex. Y. Tanaka, commonly known in Japan as “Kawachibankan”, has been carried out in 2018 by Igase et al.. The test comprise 84 participants divided in 2 experimental groups: 42 participants received the auraptene-enriched C. kawachiensis juice (equivalent to 6.0 mg/day of auraptene) and the remaining 42 were administered the placebo juice. After 24 weeks two parameters were measured, namely the mild cognitive impairment (MCI) Screen using the 10-word immediate recall test, and the mini-mental state examination (MMSE). Results of this interesting and promising study on one hand indicate that auraptene after 24 weeks did not ameliorate in an appreciable manner cognitive functions in the treated group of volunteers respect to the control one, but a great difference in the percentage change in cognitive function between the test and placebo groups (6.3 ± 18.9 vs. −2.4 ± 14.8, P < 0.05) was recorded. Multiple regression analysis showed a significant independent relationship between the percentage change in the 10-word immediate recall test score and test juice consumption including baseline 10-word immediate recall test score in all volunteers. This study confirmed the neuroprotective effects of auraptene, first highlighted by Epifano et al. (2008) in in vitro models and later also put in evidence in animal models, providing an appreciable effect of prevention of cognitive decline and suggest how auraptene-enriched phytopreparations like those commonly and easily obtainable from several edible fruits like Citrus ones can be effectively considered as safe dietary supplements for the prevention of cognitive diseases (Igase et al. 2018). What is substantially lacking in this study, however, is a detailed pharmacokinetic profile of auraptene after per os administration. The only biochemical parameters recorded were common blood exam values, mostly expressing liver functionality, that in the treated group of volunteers interestingly were not significantly different from values recorded in the control one. To this context it is noteworthy how the only two studies reported in the literature about pharmacokinetic of auraptene are still represented by those obtained in animal models, namely dogs and rats, and published respectively by Yuan et al. (2012) and Ye et al. 2016, the details of which can be found in previously published review articles on auraptene (Bibak et al. 2019; Genovese et al. 2015). So, it is desirable that in the next future the pharmacokinetic profile of auraptene in humans could be traced to largely contribute to the determination to its safety for humans.
The last acquisition in the literature about auraptene is not strictly a pharmacological one. Ference et al. in fact found that a massive increase in the biosynthesis of this geranyloxycoumarin occurred in Citrus leaves upon exposure to the fungus Xanthomonas citri (Ference et al. 2020). This is an important note underlining that the biosynthesis of auraptene in plants may represent a defensive mechanism against microbial and fungal attacks, that auraptene has been selected during evolution of living organisms to be able to interact with biomolecular targets, and finally this study represents the first report about the ecological role in the plant kingdom of this oxyprenylated coumarin.
Umbelliprenin
Umbelliprenin (3) was thought to be among the rarest oxyprenylated coumarins until 90’s. Soon after several analytical studies appeared in the literature highlighting their presence in numerous plant species, the list of which has been reviewed by Shakeri et al. (2014). More recently discovered natural sources of umbelliprenin include Ferula sinkiangensis K. M. Shen (Li et al. 2015), Peucedanum palustre (L.) Moench. (Fam. Apiaceae) (Yrjönen et al. 2016), Amaranthus retroflexus L. (Fam. Amaranthaceae) (Fiorito et al. 2017), Anethum greaveolens L., Pimpinella anisum L., Angelica archangelica L. (Apiaceae) (Taddeo et al. 2017), Melaleuca alternifolia (Maiden & Betche) Cheel (Fam. Myrtaceae) (Scotti et al. 2018), Punica granatum L. (Fiorito et al. 2019b), Spinacia oleracea L. (Fam. Amaranthaceae), Lycium barbarum L. (Fam. Solanaceae), Chenopodium quinoa Willd (Fam. Amaranthaceae) ((Fiorito et al. 2019c), and Artemisia vulgaris L. (Fiorito et al. 2020). All these analytical studies were carried out by conventional techniques, namely HPLC and UHPLC, and part of which were the same used for auraptene. The substantial novelties of some of these studies were the application of innovative and greener extraction processes (use of inclusion compounds like cyclodextrins, ionic liquids, and heterogenous solids) and the fact that the presence of umbelliprenin was generalized for some plant families like Amaranthaceae with 3 examples provided by Epifano et al. in the literature of the last 3 years. Such findings allow to hypothesize that this compound could represent a chemotaxonomic marker for the Amaranthaceae family. Umbelliprenin was also for the first time in other plant families like Lythraceae, Myrtaceae, and Solanaceae. Also, these acquisitions could prompt related studies for other species belonging to the same families, considering that all these comprise several edible, medicinal, and beneficial plants. Finally, the presence of umbelliprenin other than auraptene in pomegranate seed extracts, accompanied by the presence of other structurally related O-prenyl coumarins with partial oxidation pattern in the side chain, revealed how prenylation is a putative novel biosynthetic pathways for this plant and related species.
A very recent review article by Shahzadi et al. (2020) well covers the anti-cancer and closely related properties of umbelliprenin. However, some other important and unique properties for this compound has been discovered during the last 3 years. These refer to the strong and valuable modulatory properties of umbelliprenin on melanin biosynthesis. This was first put in evidence by Fiorito et al. (2018c). These Authors screened the properties of compound (3) and another series of natural prenyloxycoumarins in non-tumorigenic murine melanocyte Melan-a cell line. While having no cytotoxicity againts this strain when applied at a concentration of 40 µM along a period of 72 h, umbelliprenin was able to diminish the melanin content of more than 70% respect to untreated control melanocytes. In the mean time this farnesyloxycoumarin had a deep impact on key components of the melanogenic machinery. With the only exception of tyrosinase, the expressions of the enzymes tyrosinase-related protein (TRP)-1, and TRP-2, of the transcription factor melanocyte inducing transcription factor (MITF), and the transporter Rab27a were inhibited (twofold respect to controls). Based on the so ascertained whitening properties, umbelliprenin was also assayed for its capacity to interfere with the natural regulators of skin pigmentation, namely UV radiations and α-melanocyte stimulating hormone (α-MSH). Thus, when Melan-a cells were exposed to both factors for 48 with and without the concomitant administration of umbelliprenin, this latter completely abolished cell pigmentation induced by UV and/or α-MSH. In this same study Fiorito et al. hypothesized an involvement of an antagonistic effect of umbelliprenin towards ER-ß, that in turn is well known to play a pivotal role in melanin biosynthesis (Ohata et al. 2008). In view of a potential use for cosmetic purposes and for preclinical trials, the chemical stability of umbelliprenin was investigated by the same research group under forced degradation studies following the International Council for Harmonisation of Technical Requirements for Pharmaceuticals for Human Use (ICH) guidelines, with a particular attention to its exposure to UV radiations (254 nm), sunlight, and heat in alcoholic solution and in the solid state and sampling times of 15 min, 1 h, and 1–4 days. Residual umbelliprenin was then quantified by HPLC analysis (Genovese et al. 2017). Compound (3) revealed to be largely sensitive to UV radiations and sunlight in solution reaching a maximum of 91.6% degradation after exposure for 4 days to direct sunlight. On the other hand, umbelliprenin was quite stable to heat being only chemically degraded by 34% after 4 days of exposure at 80 °C. In the solid state after the same period, the quantity of this farnesyloxycoumarin decreased by 64.7% and 59.4% respectively after exposure to sunlight and UV 254 nm radiations. The mechanism of umbelliprenin degradation under these experimental conditions (formation of coumarin dimers and of an intramolecular [2 + 2] cycloaddition between the terminal carbon–carbon double bond of the farnesyl side chain and the α,ß unsaturated one) was also clearly outlined. As the production of a pharmaceutical and/or cosmetic depigmenting preparation containing chemically stable umbelliprenin may be of importance in the next future also for commercial purposes, Genovese et al. in the same study investigated the photoprotective effect of antioxidants and UV filters for compound (3). Thus, umbelliprenin solutions alone and/or in combination with antioxidants, UV-A, or UV-B filters were kept under sunlight exposure for 3 and 6 days and analyzed for photodegradation by thin layer chromatography (TLC). Tocopherol acetate, used as the antioxidant, and octinoxate, selected as the UV-B filter, completely failed to protect the oxyprenylated coumarin from photodegradation. On the other hand, UV-A filters like avobenzone and diethylaminohydroxybenzoyl hexyl benzoate (DHHB) largely prevented by 88% and 98% chemical disruption of this natural compound after 6 days. It is noteworthy that in the control vial solution after the same period only 9% of umbelliprenin was detected. This latter study is of importance in shading light into the influence of environmental factors on the stability of compound (3) and thus on the factors of its proper storage as a potential ingredient of pharmaceutical (to be used for the treatment of several skin disorders) and cosmetic formulations (depigmenting agents).
More recently in 2019, the same research group, considering that umbelliprenin is particularly abundant in plants belonging the family of Apiaceae like A. graveolens, P. anisum, and F. campestris, investigated the depigmenting properties of phytopreparations and umbelliprenin-containing extracts of these species adopting the same experimental conditions for biological tests as mentioned above (Taddeo et al. 2019). A parallelism between the content of compound (3) and the observed whitening melanocytes effect (60% for the alcoholic extract of F. campestris was the best value) was assessed for all plant extracts when administered to cell cultures at a concentration of 100 µg/mL. Thus, these latter findings include also phytocomplexes from the above cited plant species and/or their enriched extracts as potential ingredients of formulations for cosmetic purposes as well. It is noteworthy to consider in this context that extracts from the three above listed plants are already part of skin depigmenting lotions (Shojaii and Fard 2012; Sohm et al. 2011; Ding et al. 2017).
Conclusions and future perspectives
In this review article we summarized the latest literature acquisitions about two naturally occurring and biological active oxyprenylated coumarins, auraptene (2) and umbelliprenin (3). Although not so relatively numerous, the ones described herein are surely brand-new data for both compounds in terms of novel biological activities and mechanisms of actions. Taken together, most recent and past findings are quite similar in concept highlighting the great and promising potential of both title molecules for therapeutic and other beneficial uses for humans. To enforce this statement, very recently at least 20 chemical companies worldwide began commercializing auraptene and umbelliprenin either for analytical purposes as high purity chemical standards or for bulk production for uses in synthetic laboratory and/or pilot plant scales. Furthermore, about 250 international patents about auraptene and 240 for umbelliprenin have been issued mainly in the last decade. Consequently compounds (2) and (3) are of current but also hopefully of future great interest and it is highly probable and desirable that following the more and more growing trend in the number of publications on both phytochemicals observed in the last 5 years, other publications dealing with a wider array of potential pharmacological activities in humans and animals and sharper details about the action of auraptene and umbelliprenin at a biomolecular level will appear in the literature for incoming years. To this concern the overall view of the most recent findings appeared from the very beginning of 2019 to June 2020 offered by the present review article will surely be a useful guide to research teams operating in the field to more suitably and properly address their studies. Finally, it can be also suggested how, stating the plethora of data at disposition deriving from in vivo models closely resembling severe syndromes affecting humans (e.g. inflammatory-based diseases, diabetes, neurological disorders, and others) it is possible to underline how the times are suitable to accomplish further tests in humans (after the preliminary and first one accomplished by Igase et al.) if not for individual molecules, at least for plant extracts and related phytopreparations as beneficial food and/or medicinal plant supplements.
References
Abizadeh M, Novin MG, Amidi F, Ali Ziaei F, Abdollahifar MA, Nazarian A (2020) Potential of auraptene in improvement of oocyte maturation, fertilization rate, and inflammation in polycystic ovary syndrome mouse model. Reprod Sci. https://doi.org/10.1007/s43032-020-00168-9
Afshari AR, Jalili-Nik M, Soukhtanloo M, Ghorbani A, Sadeghnia HR, Mollazadeh H, Karimi Roshan MK, Rahmani F, Sabri H, Vahedi MM, Mousavi SH (2019) Auraptene-induced cytotoxicity mechanism in human malignant glioblastoma (U87) cells: role of reactive oxygen species. EXCLI J 18:576–590
Askari VR, Rahimi VB, Zargarani R, Ghodsi R, Boskabady M, Boskabady MH (2020) Anti-oxidant and anti-inflammatory effects of auraptene on phytohemagglutinin (PHA)-induced inflammation in human lymphocytes. Pharmacol Rep. https://doi.org/10.1007/s43440-020-00083-5
Bibak B, Shakeri F, Barreto GE, Keshavarzi Z, Sathyapalan T, Sahebkar A (2019) A review of the pharmacological and therapeutic effects of auraptene. BioFactors 45:867–879
Chan KKJ, O’Hagan D (2012) The rare fluorinated natural products and biotechnological prospects for fluorine enzymology. Method Enzymol 516:219–235
Cheng JT, Guo C, Cui WJ, Zhang Q, Wang SH, Zhao QH, Liu DW, Zhang J, Chen S, Chen C, Liu Y, Pan ZH, Liu A (2020) Isolation of two rare N-glycosides from Ginkgo biloba and their anti-inflammatory activities. Sci Rep 10:5994
De Medina P, Genovese S, Paillasse M, Mazaheri M, Caze-Subra S, Bystricky K, Curini M, Silvente-Poirot S, Epifano F, Poirot M (2010) Auraptene is an inhibitor of cholesterol esterification and a modulator of estrogen receptor. Mol Pharmacol 78:827–836
Ding AJ, Zheng SQ, Huang XB, Xing TK, Wu GS, Sun HY, Qi SH, Luo HR (2017) Current perspective in the discovery of anti-aging agents from natural products. Nat Prod Bioprospect 7:335–404
Eidi S, Iranshahi M, Mohammadinejad A, Mohsenzadeh MS, Farhadi F, Mohajeri SA (2020) Selective isolation of sesquiterpene coumarins from asafetida using dummy molecularly imprinted solid phase extraction method. J Chromatogr B Analyt Technol Biomed Life Sci 1138:121943
Elmeliegy M, Vourvahis M, Guo C, Wang DD (2020) Effect of P-glycoprotein (P-gp) inducers on exposure of P-gp bubstrates: review of clinical drug-drug interaction studies. Clin Pharmacokinet 59:699–714
Epifano F, Molinaro G, Genovese S, Ngomba RT, Nicoletti F, Curini M (2008) Neuroprotective effect of prenyloxycoumarins from edible vegetables. Neurosci Lett 443:57–60
Epifano F, Genovese S, Squires JE, Gray MA (2012) Nelumal A, the active principle from Ligularia nelumbifolia, is a novel farnesoid X receptor agonist. Bioorg Med Chem Lett 22:3130–3135
Erdogan Orhan I, Sezer Senol Deniz F, Ekhteiari Salmas R, Durdagi S, Epifano F, Genovese S, Fiorito S (2019) Combined molecular modeling and cholinesterase inhibition studies on some natural and semisynthetic O-alkylcoumarin derivatives. Bioorg Chem 84:355–362
Ference MC, Manthey JA, Narciso JA, Jones JB, Baldwin EA (2020) Detection of phenylpropanoids in citrus leaves produced in response to Xanthomonas citri subsp. citri. Phytopathology 110:287–296
Fiorito S, Epifano F, Palmisano R, Genovese S, Taddeo VA (2017) A re-investigation of the phytochemical composition of the edible herb Amaranthus retroflexus L. J Pharm Biomed Anal 143:183–187
Fiorito S, Epifano F, Taddeo VA, Genovese S (2018a) Recent acquisitions on oxyprenylated secondary metabolites as anti-inflammatory agents. Eur J Med Chem 153:116–122
Fiorito S, Epifano F, Taddeo VA, Genovese S, Preziuso F (2018b) A survey of the anti-microbial properties of naturally occurring prenyloxyphen ylpropanoids and related compounds. Curr Top Med Chem 18:2097–2101
Fiorito S, Epifano F, Preziuso F, Cacciatore I, di Stefano A, Taddeo VA, de Medina P, Genovese S (2018c) Natural oxyprenylated coumarins as modulators of melanogenesis. Eur J Med Chem 152:274–282
Fiorito S, Epifano F, Preziuso F, Taddeo VA, Genovese S (2019a) Biomolecular targets of oxyprenylated phenylpropanoids and polyketides. Prog Chem Org Nat Prod 108:143–205
Fiorito S, Ianni F, Preziuso F, Epifano F, Scotti L, Bucciarelli T, Genovese S (2019b) UHPLC-UV/Vis quantitative analysis of hydroxylated and O-prenylated coumarins in pomegranate seed extracts. Molecules 24:1963
Fiorito S, Preziuso F, Epifano F, Scotti L, Bucciarelli T, Taddeo VA, Genovese S (2019c) Novel biologically active principles from spinach, goji and quinoa. Food Chem 276:262–265
Fiorito S, Genovese S, Palumbo L, Scotti L, Ciulla M, Di Profio P, Epifano F (2020) Umbelliprenin as a novel component of the phytochemical pool from Artemisia spp. J Pharm Biomed Anal 184:113205
Furukawa Y, Hara RI, Nakaya M, Okuyama S, Sawamoto A, Nakajima M (2019) Citrus auraptene induces glial cell line-derived neurotrophic factor in C6 cells. Int J Mol Sci 21:253
Furukawa Y, Washimi YS, Hara RI, Yamaoka M, Okuyama S, Sawamoto A, Nakajima M (2020) Citrus auraptene induces brain-derived neurotrophic factor in neuro2a cells. Molecules 25:1117
Genovese S, Epifano F (2011) Auraptene: a natural biologically active compound with multiple targets. Curr Drug Targets 12:381–386
Genovese S, Fiorito S, Epifano F, Taddeo VA (2015) A novel class of emerging anticancer compounds: oxyprenylated secondary metabolites from plants and fungi. Curr Med Chem 22:3426–3433
Genovese S, Taddeo VA, Epifano F, Fiorito S, Bize C, Rives A, de Medina P (2017) Characterization of the degradation profile of umbelliprenin, a bioactive prenylated coumarin from a Ferulago species. J Nat Prod 80:2424–2431
Genovese S, Epifano F, Preziuso F, Stefanucci A, Scotti L, Bucciarelli T, di Profio P, Canale V, Fiorito S (2020) A novel and efficient subcritical butane extraction method and UHPLC analysis of oxyprenylated phenylpropanoids from grapefruits peels. J Pharm Biomed Anal 184:113185
Gkionis L, Kavetsou E, Kalospyros A, Manousakis D, Garzon Sans M, Butterworth S, Detsi A, Tirella A (2020) Investigation of the cytotoxicity of bioinspired coumarin analogues towards human breast cancer cells. Mol Divers. https://doi.org/10.1007/s11030-020-10082-6
Hsia CW, Tsai CL, Sheu JR, Lu WJ, Hsia CH, Velusamy M, Jayakumar T, Li JY (2019) Suppression of human platelet activation via integrin αIIbβ3 outside in independent signal and reduction of the mortality in pulmonary thrombosis by auraptene. Int J Mol Sci 20:5585
Igase M, Okada Y, Ochi M, Igase K, Ochi H, Okuyama S, Furukawa Y, Ohyagi Y (2018) Auraptene in the peels of Citrus kawachiensis (Kawachibankan) contributes to the preservation of cognitive function: a randomized, placebo-controlled, double-blind study in healthy volunteers. J Prev Alzheimer’s Dis 5:197–201
Jalilzadeh N, Samadi N, Salehi R, Dehghan G, Iranshahi M, Dadpour MR, Hamishehkar H (2020) Novel nano-vehicle for delivery and efficiency of anticancer auraptene against colon cancer cells. Sci Rep 10:1606
Jang S, Choo H, Lee MJ, Han J, Kim JS, Ju X, Cui J, Lee YL, Ryu MJ, Oh ES, Choi SY, Chung W, Kweon GR, Heo JY (2019) Auraptene mitigates Parkinson’s disease like behavior by protecting inhibition of mithochondrial respiration and scavenging reactive oxygen species. Int J Mol Sci 20:3409
Kim CS, Oh J, Subedi L, Kim SY, Choi SU, Lee KR (2018) Rare thioglycosides from the roots of Wasabia japonica. J Nat Prod 81:2129–2133
Li G, Li X, Cao L, Zhang L, Shen L, Zhu J, Wang J, Si J (2015) Sesquiterpene coumarins from seeds of Ferula sinkiangensis. Fitoterapia 103:222–226
Mottaghipisheh J, Kusz BN, Hohmann J, Tsai YC, Csupor D (2020) Isolation of secondary metabolites from the Iranian medicinal plant Eremurus persicus. Z Naturforsch C J Biosci. https://doi.org/10.1515/znc-2020-0059
Nabekura T, Kawasaki T, Kato Y, Kawai K, Fiorito S, Epifano F, Genovese S, Uwai Y (2020) Citrus auraptene induces drug efflux transporter P-glycoprotein expression in human intestinal cells. Food Funct 11:5017–5023
Ohata C, Tadokoro T, Itami S (2008) Expression of estrogen receptor beta in normal skin, melanocytic nevi and malignant melanomas. J Dermatol 35:215–221
Omar AM, Dibwe DF, Tawila AM, Sun S, Phrutivorapongkul A, Awale S (2019) Chemical constituents of Anneslea fragrans and their antiausterity activity against the PANC-1 human pancreatic cancer cell line. J Nat Prod 82:3133–3139
Petkowski JJ, Bains WW, Seager S (2019) Natural products containing ‘rare’ organophosphorus functional groups. Molecules 24:866
Salari H, Afkhami-Poostchi A, Soleymanifard S, Nakhaei-Rad S, Merajifar E, Iranshahi M, Matin MM, Rassouli FB (2020) Coadministration of auraptene and radiotherapy; a novel modality against colon carcinoma cells in vitro and in vivo. Int J Radiat Biol 2020:1–9
Scotti L, Genovese S, Bucciarelli T, Martini F, Epifano F, Fiorito S, Preziuso F, Taddeo VA (2018) Analysis of biologically active oxyprenylated phenylpropanoids in tea tree oil using selective solid-phase extraction with UHPLC-PDA detection. J Pharm Biomed Anal 154:174–179
Shahzadi I, Ali Z, Baek SH, Mirza B, Ahn KS (2020) Assessment of the antitumor potential of umbelliprenin, a naturally occurring sesquiterpene coumarin. Biomedicines 8:126
Shakeri A, Iranshahy M, Iranshahi M (2014) Biologicalproperties and molecular targets of umbelliprenin—a mini-review. J Asian Nat Prod Res 16:884–889
Shojaii A, Fard MA (2012) Review of pharmacological properties and chemical constitutents of Pimpinella anisum. ISRN Pharm 2012:510795
Sohm B, Cenizo V, Andrè V, Zahouani H, Pailler-Mettel C, Vogelgesang B (2011) Evaluation of the efficacy of a dill extract in vitro and in vivo. Int J Cosmet Sci 33:157–163
Taddeo VA, Genovese S, de Medina P, Palmisano R, Epifano F, Fiorito S (2017) Quantification of biologically active O-prenylated and unprenylated phenylpropanoids in dill (Anethum graveolens), anise (Pimpinella anisum), and wild celery (Angelica archangelica). J Pharm Biomed Anal 134:319–324
Taddeo VA, Epifano F, Preziuso F, Fiorito S, Caron N, Rives A, de Medina P, Poirot M, Sylvente-Poirot S, Genovese S (2019) HPLC analysis and skin whitening effects of umbelliprenin-containing extracts of Anethum graveolens, Pimpinella anisum, and Ferulago campestris. Molecules 24:501
Teng CM, Li HL, Wu TS, Huang SC, Huang TF (1992) Antiplatelet actions of some coumarin compounds isolated from plant sources. Thromb Res 66:549–557
Wang J, Fu T, Dong R, Wang C, Liu K, Sun H, Huo X, Ma X, Yang X, Qiang M (2019) Hepatoprotection of auraptene from the peels of citrus fruits against 17α-ethinylestradiol-induced cholestasis in mice by activating farnesoid X receptor. Food Funct 10:3839
Xi Y, An L, Yang X, Song Z, Zhang J, Tuerhong M, Jin DQ, Ohizumi Y, Lee D, Xu J, Guo Y (2019) NO Inhibitory phytochemicals as potential anti-inflammatory agents from the twigs of Trigonostemon heterophyllus. Bioorg Chem 87:417–424
Xie T, Song S, Li S, Ouyang L, Xia L, Huang J (2015) Review of natural product databases. Cell Prolif 48:398–404
Ye XD, Ouyang H, Zhong LY, Li TE, Rao XY, Feng YL, Yang WL (2016) Rapid and sensitive LC-MS/MS method for the determination of auraptene in rat plasma and its application in a pharmacokinetic and bioavailability study in rats. Genet Mol Res 15:gmr8786
Yrjönen T, Eeva M, Kauppila TJ, Martiskainen O, Summanen J, Vuorela P, Vuorela H (2016) Profiling of coumarins in Peucedanum palustre (L.) Moench populations growing in Finland. Chem Biodiv 13:700–709
Yuan J, Ming L, Chen H, Wuliang Y, Xiaoquan L, Yingying L, Yu Z (2012) An HPLC method for the determination of auraptene in dog plasma: application to pharmacokinetic study. Lat Am J Pharm 31:251–256
Zolfaghari B, Yazdiniapour Z, Sadeghi M, Akbari M, Troiano R, Lanzotti V (2020) Cinnamic acid derivatives from Welsh onion (Allium fistulosum) and their anti bacterial and cytotoxic activities. Phytochem Anal. https://doi.org/10.1002/pca.2924
Funding
Open access funding provided by Università degli Studi G. D'Annunzio Chieti Pescara within the CRUI-CARE Agreement.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Fiorito, S., Preziuso, F., Sharifi-Rad, M. et al. Auraptene and umbelliprenin: a review on their latest literature acquisitions. Phytochem Rev 21, 317–326 (2022). https://doi.org/10.1007/s11101-020-09713-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11101-020-09713-5