
Abstract
Natural regeneration of Norway spruce (Picea abies (L.) Karst.) is a relatively common practice in Norway on medium to low site indices. However, seedling establishment is often hampered by rapid regrowth of competing vegetation in scarified patches. The aim of our study was to examine the effect of coordinating scarification towards an expected seed-fall, by studying germination and seedling establishment in scarified patches of different age (fresh, one- and two-year-old). The experiment was conducted in two stands in southeast Norway that were clear-cut in 2007. Scarification was applied to subplots in autumn 2008–2010. To simulate seed-fall, seeds were sown in fresh scarification patches in spring 2009–2011, in one-year-old patches in 2010 and 2011, and in two-year-old patches in 2011. Both germination and seedling survival were negatively affected by the age of the scarified patches. Germination was higher, and mortality lower, at the small fern woodland site, compared with the bilberry woodland site. Sowing in fresh patches also resulted in increased height and root collar diameter of the seedlings compared with sowing in older patches. It is likely that the competing vegetation both on the site and in the scarification patches affected the growth of the seedlings. In conclusion, the age of the scarified patches affected both germination and mortality, as well as early growth of the seedlings.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Natural regeneration of Norway spruce (Picea abies L. Karst) is a relatively common regeneration practice in Norway on medium to low site indices. According to data from the annual regeneration survey, natural regeneration, or a combination of planting and natural regeneration, was used on about 15% of the total spruce area subjected to final harvest in 2007–2010 (Granhus et al. 2015). Most of these areas were cut using group feelings or small clear-cuts where the seed sources are the trees along the edge of the gaps. However, less than 60% of the naturally regenerated areas were expected to reach the recommended density of established seedlings.
Availability of seeds and microsites, respectively, are two factors that potentially limit recruitment in plant populations (Sarvas 1957; Eriksson and Ehrlen 1992). Several studies have documented the positive impact of mechanical site preparation on establishment and growth of forest regeneration (see e.g. review by Löf et al. 2012). The removal of vegetation and humus reduces competition, and enhances moisture conditions (Winsa 1995) and the seed-substrate contact (Chrosciewicz 1990; Winsa and Bergsten 1994). It also improves temperature conditions (Bjor 1971; Blennow 1998; Langvall et al. 2001) and nutrient availability (Nordborg and Nilsson 2003; Nordborg et al. 2003) as well as reducing pine weevil damage (Örlander and Nilsson 1999; Heiskanen and Viiri 2005; Johansson et al. 2013). However, in clear-cuts the scarified patches will be colonized by herbs and grasses shortly after treatment (Hanssen et al. 2003), so that the positive effect on regeneration is reduced with time. Regrowth of ground vegetation will expectedly happen more slowly on low site indices than on rich soils (Hagner 1962).
In an experiment with Scots pine (Pinus sylvestris L.), Karlsson and Örlander (2000) found that seedling density was higher when soil scarification took place prior to a seed year compared to scarification immediately after cutting. As Norway spruce has less frequent seed years than Scots pine (Sarvas 1957; Koski and Tallqvist 1978) timing is even more important for this species. The probability of a good seed crop in Norway spruce can be estimated in advance, for instance by examining buds during logging in autumn or winter, as the generative buds are formed in the year prior to flowering (Tirén 1935; Owens and Blake 1985).
In general, forest owners are encouraged to regenerate their forests as soon as possible after cutting, which is also convenient from a planning point of view. The Norwegian Regulations on Sustainable Forestry (2006) demand that necessary measures to ensure satisfactory regeneration should be implemented within 3 years after logging. However, when soil scarification is performed shortly after clear-cutting there is a risk of delay until sufficient seed fall is obtained (Sarvas 1957; Drössler et al. 2017). Then, competing vegetation will be established in the scarified spots (Nilsson and Örlander 1999; Béland et al. 2000), making them less valuable as a germination substrate. Thus, there might be a discrepancy between forestry regulations and planning on one hand and the timing of site preparation with good seed years on the other.
To enable better predictions on the likely outcome of natural regeneration in general, there is a need of quantitative data on the proportion of seeds that germinate in different substrates, as well as on the survival and development of seedlings throughout the initial establishment phase. With respect to natural regeneration of Norway spruce after clear-cutting in particular, there is a lack of knowledge about how timing of scarification influences the regeneration success, and how seedling growth is affected by the age of the scarification patches.
The aim of our study was therefore to investigate how timing of scarification towards an expected seed-fall of Norway spruce influence germination and seedling establishment, and how the age of the scarification patch affects establishment success. The study also addresses to what extent the height and root collar diameter of established seedlings is affected by the timing of scarification relative to seed fall.
Our hypotheses were that (1) increased age of the scarification patches before seed fall would reduce germination, (2) mortality of seedlings will increase with age of the scarification patches at the time of germination, and (3) seedlings germinated in fresh scarification patches will have increased growth compared with seedlings that germinate in one or two-year-old scarification patches.
Materials and methods
Experimental sites
The experiment was established in two clear-cut stands in south-east Norway (Romedal and Stange Commons, Hedmark County). Both stands were located 380–400 m a.s.l on site index class G14, according to the Norwegian H40-system (Tveite 1977). This corresponds to a potential mean annual production of stem wood with bark at 5.5 m3 ha−1. The vegetation types in the two stands were Eu-Piceetum dryopteridetosum and Eu-Piceetum myrtilletosum (Kielland-Lund 1981), and they are hereafter referred to as the small fern woodland and bilberry woodland sites, respectively. The small fern woodland stand (60°57′N 11°29′E) has a north to northwest facing aspect, with inclination varying from 0 to ca. 20%, whereas the bilberry stand (60°59′ N 11° 35′ E) is located on a west facing slope with inclination varying between 5 and 20%. The soil texture is sandy loam (Hodgson 1974) at both sites, however with slightly higher proportions of the clay and sand fractions at the small fern site.
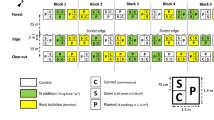
Both stands were originally dominated by Norway spruce and clear-cut in 2007. In September 2008, each stand was divided into three replicates and each replicate was randomly divided into three subplots where soil scarification was performed in October each of the years 2008–2010, respectively (Fig. 1). Scarification was done using a Donaren 870H aggregate mounted on a Valmet 902 forwarder. Aggregate settings were adjusted to create scarification patches of ca. 2.2 m length. Patch width was about 30–40 cm. The same equipment and machine operator was used each year. The distance from the patches used in this study to the edge of the clear-cuts did not exceed 50 m.

Experimental design, illustrated by the plot configuration in the bilberry woodland stand
Seed supply and germination
To simulate seed-fall, 40 stand origin seeds of local provenance (Romedal, seed lot F07-039, tested germination percentage = 100) were sown in each of 20 fresh scarified patches per site and replicate in mid-May each year 2009–2011, in twenty one-year-old patches in mid-May 2010 and 2011, and in twenty two-year-old patches in mid-May 2011. The seeds were sown inside a quadrangle of 25 × 50 cm with exposed mineral soil, and each quadrangle was marked with plastic pegs in each corner shortly after scarification. A total of 120 patches per replicate and site were sown, resulting in 720 patches in the experiment.
Seedling emergence was recorded in late June/early July, and with subsequent registrations of emergence of late germinating seedlings and survival in late September/early October. Each emerged seedling was marked with a small plastic stick, allowing the fate of each individual to be followed throughout the experiment. Germination was also recorded in control patches that were not sown, to be able to quantify natural seed supply. Germination in control patches that were not sown was negligible, thus the results presented are from sown seeds only. Mortality was recorded shortly after snowmelt in May the second year, as well as in late September/early October the second and third years. Height and root collar diameter of the tallest seedling in each patch was recorded after the third growing season.
Climatic variations
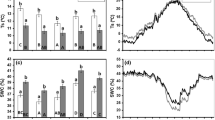
Precipitation and temperature data for the period 7th of May until 30th of October each year 2009–2011 were collected from Hamar meteorological station, which is operated by the Norwegian Meteorological Institute (Fig. 2). In 2009 the spring and early summer were dryer than normal, while 2011 had more precipitation than normal during May–June as well as in August and September.

Weekly total precipitation and average temperature at Hamar meteorological station
Vegetation
The vegetation regrowth in 20 unsown scarified patches per replicate and site was recorded with respect to percent cover, species composition and mean height in mid-August following each year of scarification. The recordings were made within the marked quadrangle of 25 × 50 cm. Species were grouped into mosses, grasses, herbs and woody species, which mainly included birch, salix and raspberry. Recording of vegetation was done in the same patches during three consecutive years in subplots scarified during 2008 (in August 2009–2011), during 2 years following scarification in 2009 (in August 2010 and 2011), and 1 year following scarification in 2010 (in August 2011). The amount of vegetation on undisturbed soil between the patches was simultaneously estimated based on destructive harvesting of all aboveground vegetation within five randomly placed quadrangles of 25 × 50 cm per replicate and site. The dry weigh of each sample was recorded after drying at 65 °C for 24–48 h.
Statistical analyses
We used a linear effects model for the outcome Y * srki , germination of seed i or mortality of plant i, in treatment k in replicate r in site s:
where μ 0 is the overall mean, β s is the effect of site s, β r is the effect of replicates, β k is the effect of treatment k, β sr is the effect of replicate r within site s, β sk is the effect of treatment k within site s, and ε srki is the random error term assumed to be independently and identically distributed with a normal distribution with mean zero.
The model in (1) is interpreted as a latent variable model when analysing the germination of individual seeds, the mortality of seed plants and proportion of empty patches, where the observed binary outcome y srki is given as
The probability of a positive outcome is thus given as a probit model and was estimated using maximum likelihood techniques (Wooldridge 2010).
The model in Eq. (1) was also used to analyse to what extent the height and the root collar diameter of established seedlings is affected by timing of scarification relative to the age of the scarified patches at the time of sowing. To compare the different treatments a least significant difference procedure with the Bonferonni test was used. The analyses were conducted using version 9.3 of the SAS/STAT® software (SAS Institute Inc 2014).
Results
Germination
There was a significantly higher overall germination rate (across all patches) at the small fern woodland site (13.8%) compared with the bilberry woodland site (6.0%) (Table 1; Fig. 3). The overall germination rate across both sites and year of sowing declined with age of the scarification patches. In fresh patches 12.9% of the seeds germinated, while in one- and two-year-old patches the average germination rates were 7.8 and 5.0%, respectively (Fig. 3). There were significant differences in germination rate between all combinations of age of patches (p ≤ 0.0468).

Germination of P. abies seeds sown in different years (2009–2011) and in soil scarification patches of different age, on the small fern woodland site (a) and the bilberry woodland site (b). The grey lower parts of the bars represent germination recorded 7–8 weeks after sowing and the entire bars represent the total germination during the first year. Vertical bars indicate the standard errors of the total germination means
Of all germinated seeds, 62% were recorded as germinated in the summer registrations (mean across both sites and patches of different age). The rest, 38%, were recorded as germinated in the autumn registration. However, the percentage of seeds that had germinated at the time of the first registration in summer varied substantially between sowing years (Fig. 3). In 2009 all seeds had germinated by the time of the summer registration, while 67 and 78% of the seeds were germinated by the time of the summer registration in 2011 and 2010, respectively.
When sowing in fresh patches, the year of sowing affected the germination rate significantly (p ≤ 0.0001) (Table 2; Fig. 3), and there was significant interaction between site and year of sowing due to a rank change among years between the two sites. The germination rate in fresh patches was higher at the small fern woodland site compared with the bilberry site, 18.0 and 7.9%, respectively. The germination rate in fresh patches was highest in 2010 with 16.5% (average across both sites) compared with germination rates of 8.4 and 13.8% in 2009 and 2011, respectively (Fig. 3).
Mortality
On average, the mortality among the germinated seedlings was 63.8% after three seasons (mean across different sites, sowing years and age of patches). There were significant differences in overall mortality (across all patches) between sites (Fig. 4; Table 1). The lowest mortality was observed at the small fern woodland site (54.7%) whereas the average mortality at the bilberry woodland site was 72.8% (Fig. 4). There were significant differences in mortality between all combinations of age of patches (p ≤ 0.0191).The highest mortality appeared following sowing in two-year-old patches (81.1%), whereas the average mortality in one-year-old and fresh patches were 67.9 and 55.2%, respectively (Fig. 4).

Mortality of P. abies seedlings following sowing in different years (2009–2011) and in soil scarification patches of different age. Results from the small fern woodland site (a) and the bilberry woodland site (b). Dark grey represents first winter mortality, light grey represents second summer mortality, and the upper white part represents mortality the second winter and the third summer. Vertical bars indicate the standard errors of the total germination means
Comparing mortality after sowing in fresh patches, the results show that the year of sowing affected the mortality significantly. However, the year of sowing influenced the mortality of seedlings sown in fresh patches differently at the two sites (significant interaction between site and year of sowing, Table 2). At the bilberry site, the highest mortality occurred in 2009, while at the small fern woodland site mortality was highest in 2011. At both sites, the lowest mortality occurred following sowing in 2010. There was significantly higher mortality after sowing in fresh patches in 2011 (62.6%) compared with sowing in fresh patches in 2010 (42.6%) (p < 0.0001).
When sowing in fresh patches, a higher part of the total mortality occurred during the first winter, compared with sowing in one- or two-year-old patches (Fig. 4). An exception to this is the one-year-old patches sown in 2009 at the bilberry woodland site, where no mortality occurred after the first winter. Generally, only limited mortality occurred during the second summer, especially when sowing in fresh patches. There were no noteworthy differences in total amount or timing of mortality between early versus late germinated seedlings.
Empty patches
At the small fern woodland site, the percentage of empty patches three growing seasons after sowing increased with the age of the patches at the time of sowing (Fig. 5a). At the bilberry woodland site the percentage of empty patches was generally high, but sowing in fresh patches in 2010 and 2011 resulted in the lowest number of empty patches (Fig. 5b).

Percentage (± SE) of patches without seedlings of P. abies following sowing in different years (2009–2011) and in soil scarification patches of different age. Results from the small fern woodland site (a) and the bilberry woodland site (b)
Seedling growth
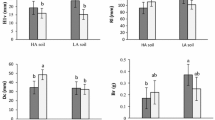
Three growing seasons after sowing, seedlings sown in fresh patches were significantly (p ≤ 0.0226) taller (mean height 11.1 cm) compared with seedlings sown in one-year-old (8.2 cm) or two-year-old (7.3 cm) patches (mean values for the tallest seedlings in each patch measured 3 years after sowing) (Table 3; Fig. 6a). Seedlings sown in fresh patches also had significantly (p ≤ 0.0076) larger root collar diameters (mean diameter 1.8 mm), compared with seedlings sown in one-year-old (1.3 mm) or two-year-old patches (1.1 mm) (Table 3; Fig. 6b). Neither for seedling height nor root collar diameter were the differences between one- and two-year-old patches significant. No significant differences appeared between seedlings grown at the small fern and bilberry woodland sites.

Height (a) and root collar diameter (b) of P. abies seedlings after three growing seasons, following sowing in soil scarification patches of different age
Comparing seedling growth after sowing in fresh patches, the results shows that year of sowing also affected seedling growth significantly. Seedlings sown in 2011 were significantly (p ≤ 0.0121) shorter (mean height 9.6 cm) compared with seedlings sown 2010 (12.3 cm) or 2009 (11.0 cm). Root collar diameter also differed significantly (p ≤ 0.0395) among years of sowing, with largest root collar diameters in seedlings sown in 2009 (mean diameter 2.2 mm), compared with seedlings sown in 2010 (1.8 mm) or 2011(1.4 mm).
Vegetation cover
The vegetation cover in the scarified patches increased substantially the second summer after soil scarification, independently of site and year of scarification (Fig. 7). Regrowth of the patches was slightly higher at the bilberry woodland site (Fig. 7b) compared with the small fern woodland site (Fig. 7a). Species of grasses were dominating in the scarification patches at both sites (Table 4). The cover of woody species increased both with the age of the clear-cuts and the age of the patches.

Vegetation cover in scarification patches recorded in August one, two and three years after soil scarification on the small fern woodland site (a) and the bilberry woodland site (b)
On undisturbed ground between the scarification patches the vegetation cover was high already 2 years after the clear-cutting (Table 5), and species of grass dominated. On the small fern woodland site the total dry weigh of the aboveground vegetation was 813, 1390 and 1303 kg ha−1 in 2009–2011, respectively, while the corresponding values were 2541, 3080 and 3115 kg ha−1 site on the bilberry woodland site.
Discussion
In order to succeed with natural regeneration of spruce after clear-cutting, sufficient seed supply as well as a suitable germination substrate are needed. As the effective dispersal range of Norway spruce seeds is limited, the maximal distance from surrounding edge that may allow sufficient seed supply is limited to ca. one-two times the height of the edge trees (Hanssen 2003). Although this imposes obvious limits on the maximal size of clear-cuts, for the majority of the available seedbeds the conditions affecting germination, early survival and growth will differ markedly from those that prevail when regenerating the stand by using small gaps or methods that maintain a uniform canopy cover. Important factors that are strongly affected by gap size and the degree of canopy cover include e.g. the occurrence of temperature extremes (Langvall and Löfvenius 2002), establishment and amount of competing vegetation (Nilsson et al. 2002), as well as the rate of regrowth in scarified patches (Béland et al. 2000; Hanssen et al. 2003).
In the current experiment, the dynamics of early seedling establishment of Norway spruce after clear-cutting and soil scarification was studied by simulating natural seed fall. A controlled number of seeds were sown in scarified patches of different age at two sites with different vegetation types, and in three consecutive years. We were able to quantify the proportions of Norway spruce seeds that actually germinated and survived throughout the critical establishment phase in scarified patches with different site and weather conditions during the 3 years. Ultimately, such quantitative estimates are needed to improve our understanding of the factors leading to success or failure when applying different silvicultural systems and regeneration methods that depend on seeding or natural regeneration.
Previous studies conducted on similar site types as those in the current study have reported rapid vegetation regrowth of scarified spots (Hanssen et al. 2003), which was confirmed by our results. Our results also clearly showed that scarification should be carried out towards an expected seed year, to facilitate germination before competing vegetation establishes in the scarified patches. Reduced germination and increased mortality with age of the scarification patches confirmed hypotheses 1 and 2. This corroborates findings in scarified seed tree stands of Scots pine by Karlsson and Örlander (2000). A regeneration survey previously conducted in scarified clear-cut stands in the same area as this study, also showed that the number of seedlings established from natural seed fall diminished rapidly with increasing time between the soil scarification and the subsequent seed year (Granhus and Fløistad 2010).
In this study, we observed considerable differences in germination and mortality between the two sites, with higher germination and a lower overall mortality on the small fern site type compared to the bilberry site. Generally, when the stand is not scarified, the small fern site type is considered easier to regenerate naturally, due to more favourable soil conditions (i.e. a thinner humus layer with better capillary properties) and usually more stable moisture conditions (Larsson and Søgnen 2003). Although there were no true replicates of the site type in this study, our results indicate better conditions for germination and early survival on the small fern site type even when the seedlings are allowed to establish on a substrate of mineral soil. In the previously mentioned regeneration survey in scarified stands, densities of naturally regenerated spruce were also considerably higher on this site type compared with bilberry woodland. It should be noted however, that in our study the aspects of the two stands were also different, with the bilberry site being located in a west facing slope which was likely more prone to desiccation than the small fern site, which has a predominantly north facing aspect. These differences were probably also important to the outcome, and may explain some of the interactions observed. For example, the differences in germination between the two sites were less pronounced in 2011 than in 2009 and 2010, which led to a significant site*year interaction that most likely can be attributed to a higher precipitation during spring and early summer in 2011 than in the two previous years. At the more drought prone bilberry site, the best germination results were observed this year.
Also for mortality in fresh patches we observed a significant site*year interaction, which can be due to site differences in moisture regime. Our observations during the field registrations in autumn 2011 indicate that several seedlings drowned in the scarified patches at the small fern site that year, due to the high precipitation in late summer, which may also have caused soil erosion (Fig. 2). These observations are also reflected in the mortality of seedlings sown in fresh patches in 2011, for which first-year mortality was substantially higher at the small fern site than at the bilberry site, while it was opposite in 2009 and 2010. Thus, annual variation in rainfall was most likely the main factor affecting germination and mortality in this study, besides the effect of age of the scarified patches. Annual variations in precipitation and temperature may also have influenced the large between-year variation in the percentage of seedlings that had germinated at the time of the first registration in mid-summer, which varied from 100% in sowings made in 2009, to 67% in sowings made in 2011.
A substantial proportion of the mortality over the 3-year study period occurred during the first winter (between late September/early October and early May the following year), which was also shown for recently germinated Norway spruce seedlings by Granhus et al. (2008) and for Scots pine by Béland et al. (2000). Although we were not able to identify the actual causes of first winter mortality in the current study, it is likely that several seedlings were lost due to frost heaving (Goulet 1995). The risk of frost heaving may be especially high after deep soil scarification on fine-textured soil, and even more so on scarified clear-cuts where no residual trees are left to reduce the frequency of repeated freeze-thaw cycles (de Chantal et al. 2007).
In addition to physical factors, predation accounts for the loss of many seeds and juvenile seedlings, though studies show great variation (Nystrand and Granström 2000). Carabids, slugs, voles and birds are the dominant predators of seeds and newly germinated seedlings (Bjor 1971; Nilsen 1986; Nystrand and Granström 2000). Silvicultural treatments such as site preparation will affect seed and seedling predation. For instance, Nystrand and Granström (1997a) found a higher seed predation by birds on pure mineral soil than in a moss seedbed. Predation by slugs on juvenile seedlings was highest on moist forest floors (Nystrand and Granström 1997b). In general, however, predation was low in clear-cuts compared to shelterwoods or unlogged stands (Nystrand and Granström 2000). It is therefore likely that physical factors have caused most of the mortality in our study.
Not only germination and survival but also growth of the small seedlings is of importance to a successful seedling establishment. The results clearly show that seedlings established in fresh patches grew faster than seedlings established in one- or two-year-old patches, and thus confirmed our third hypothesis. This corroborates also with a substantial amount of long-term data showing yield gain when competing vegetation is effectively managed (Wagner et al. 2006). The root collar diameter was more clearly affected than seedling height in our study, in accordance with Jylhä and Hytönen (2006). Growth was also clearly affected by the year of scarification, with increased growth the earlier after clear-cutting the scarification was carried out. Vegetation cover increased rapidly in the scarified patches and grasses constituted the main component of the vegetation on both sites. As stated by Balandier et al. (2006), the competitive effect of grasses may be extremely high shortly after seedling establishment. It is likely that both the general regrowth of the site as well as regrowth in the scarification patches affected the growth of the small spruce seedlings.
Initial differences in growth rates would eventually allow the more fast-growing seedlings to gain a competitive advantage relative to competing vegetation, and to faster reach a size where they are more able to withstand lethal damage from herbivores and insects such as pine weevils (Hylobius abietis L.). The effects of these differences on the future stand development may only be assessed through more long-term studies.
Conclusions
Both germination, mortality and seedling growth was negatively affected with increasing time between scarification and year of sowing. This illustrates the need to consider the rapid vegetation dynamics in the scarified patches when regenerating Norway spruce from seeds after clear-cutting and soil scarification. Moreover, the results of the current study and the previously mentioned regeneration survey (Granhus and Fløistad 2010), conducted in the same region, both showed that the probability of obtaining sufficient regeneration was greater on the small fern vegetation type than on bilberry woodland. This emphasizes that the site type in question may need to be carefully taken into account. We consider the results of the current study to be relevant for both natural regeneration and direct seeding on small clear-cuts. When applying natural regeneration, the need to time the site preparation with the availability of seeds implies considerable logistic challenges for forest managers, due to the infrequent seed years in Norway spruce.
References
Balandier P, Collet C, Miller JH, Reynolds PE, Zedaker SM (2006) Designing forest vegetation management strategies based on the mechanisms and dynamics of crop tree competition by neighbouring vegetation. Forestry 79:1–27. https://doi.org/10.1093/forestry/cpi056
Béland M, Agestam E, Ekö P-M, Gemmel P, Nilsson U (2000) Scarification and seedfall affects natural regeneration of Scots pine under two shelterwood densities and a clear-cut in southern Sweden. Scand J For Res 15:247–255
Bjor K (1971) Forest meteorological, soil climatological and germination investigations. Medd Nor Skogforsøksves 28:429–526
Blennow K (1998) Modelling minimum air temperature in partially and clear felled forests. Agric For Meteorol 91:223–235
Chrosciewicz Z (1990) Site conditions for jack pine seeding. For Chron 66:579–584
de Chantal M, Hanssen KH, Granhus A, Bergsten U, Ottosson Löfvenius M, Grip H (2007) Frost heaving damage to one-year-old Picea abies seedlings increases with soil horizon depth and canopy gap size. Can J For Res 37:1236–1243
Drössler L, Fahlvik N, Wysocka NK, Hjelm K, Kuehne C (2017) Natural regeneration in a multi-layered Pinus sylvestris–Picea abies forest after target diameter harvest and soil scarification. Forests 8:1–14
Eriksson O, Ehrlen J (1992) Seed and microsite limitation of recruitment in plant populations. Oecologia 91:360–364
Goulet F (1995) Frost heaving of forest tree seedlings: a review. New For 9:67–94
Granhus A, Fløistad IS (2010) Naturlig foryngelse etter markberedning på middels bonitet (G14). Forsk skog landsk 1/10: 23 p
Granhus A, Hanssen KH, de Chantal M (2008) Emergence and seasonal mortality of naturally regenerated Picea abies seedlings: effects of overstory density and two site preparation methods. New For 35:75–82
Granhus A, Fløistad IS, Eriksen R (2015) Hogst og foryngelse av granskog: Tilstandsbeskrivelse basert på Resultatkontroll skogbruk/miljø og Landsskogtakseringen. Rapp Skog landsk 04/15: 23 p
Hagner S (1962) Naturlig föryngring under skärm. En analys av föryngringsmetoden, dess möjligheter och begränsningar i mellannorrländskt skogsbruk. [Natural regeneration in shelterwood stands] Medd Statens skogforsk inst 52: 263 p
Hanssen KH (2003) Natural regeneration of Picea abies on small clear-cuts in SE Norway. For Ecol Manage 180:199–213
Hanssen KH, Granhus A, Brække FH, Haveraaen O (2003) Performance of sown and naturally regenerated Picea abies seedling under differet scarification and harvesting regimens. Scand J For Res 18:351–361
Heiskanen J, Viiri H (2005) Effects of mounding on damage by the European pine weevil in planted Norway spruce seedlings. North J Appl For 22:154–161
Hodgson JM (1974) Soil survey field handbook. Soil Survey of England and Wales, Harpenden
Johansson K, Nilsson U, Örlander G (2013) A comparison of long-term effects of scarification methods on the establishment of Norway spruce. Forestry 86:91–98. https://doi.org/10.1093/forestry/cps062
Jylhä P, Hytönen J (2006) Effect of vegetaion control on the survival and growth of Scots pine and Norway spruce planted on former agricultural land. Can J For Res 36:2400–2411
Karlsson C, Örlander G (2000) Soil scarification shortly before a rich seed fall improves seedling establishment in seed tree stands of Pinus sylvestris. Scand J For Res 15:256–266. https://doi.org/10.1080/028275800750015073
Kielland-Lund J (1981) Die Waldgesellschaften SO-Norwegens. Phytocoenologia 9:53–250
Koski V, Tallqvist R (1978) Results of long-time measurements of the quantity of flowering and seed crop of forest trees. Folia For 364:1–60
Langvall O, Löfvenius MO (2002) Effect of shelterwood density on nocturnal near-ground temperature, frost injury risk and budburst date of Norway spruce. For Ecol Manage 168:149–161
Langvall O, Nilsson U, Örlander G (2001) Frost damage to planted Norway spruce seedlings—influence of site preparation and seedling type. For Ecol Manage 141:223–235
Larsson JY, Søgnen SM (2003) Vegetasjon i norsk skog: vekstvilkår og skogforvaltning. Landbruksforlaget, Oslo
Löf M, Dey DC, Navarro R, Jacobs DF (2012) Mechanical site preparation for forest resoration. New For 43:825–848. https://doi.org/10.1007/s11056-012-9332-x
Nilsen P (1986) Loss of seeds and seedlings of Norway spruce (Picea abies (L.) Karst.) after sowing on different vegetation types in mountain forest and lowland forest. Medd Nor Inst Skogforsk 39:130–145
Nilsson U, Örlander G (1999) Vegetation management on grass-dominated clearcuts planted with Norway spruce in southern Sweden. Can J For Res 29:1015–1126. https://doi.org/10.1139/x99-071
Nilsson U, Gemmel P, Johansson U, Karlsson M, Welander T (2002) Natural regeneration of Norway spruce, Scots pine and birch under Norway spruce shelterwoods of varying densities on a mesic-dry site in southern Sweden. For Ecol Manage 161:133–145
Nordborg F, Nilsson U (2003) Growth, damage and net nitrogen uptake in Picea abies (L.) Karst. seedlings, effects of site preparation and fertilisation. Ann For Sci 60:657–666. https://doi.org/10.1051/forest:2003058
Nordborg F, Nilsson U, Örlander G (2003) Effects of different soil treatments on growth and net nitrogen uptake of newly planted Picea abies (L.) Karst. seedlings. For Ecol Manage 180:571–582. https://doi.org/10.1016/s0378-1127(02)00650-3
Nystrand O, Granström A (1997a) Post-dispersal predation on Pinus sylvestris seeds by Fringilla spp: ground substrate affects selection for seed color. Oecologia 110:353–359
Nystrand O, Granström A (1997b) Forest floor moisture controls predator activity on juvenile seedlings of Pinus sylvestris. Can J For Res 27:1746–1752
Nystrand O, Granström A (2000) Predation on Pinus sylvestris seeds and juvenile seedlings in Swedish boreal forest in relation to stand disturbance by logging. J Appl Ecol 37:449–463
Örlander G, Nilsson U (1999) Effect of reforestation methods on pine weevil (Hylobius abietis) damage and seedling survival. Scand J For Res 14:341–354
Owens JN, Blake MD (1985) Forest tree seed production. In: Information report PI-X-53. Canadian Forest Service, 161 p
Sarvas R (1957) Studies on the seed setting of Norway spruce. Medd Nor Skogforsøksves 48:529–556
SAS Institute Inc (2014) SAS/STAT® 9.3 User’s guide. http://support.sas.com/documentation/cdl/en/statug/63962/HTML/default/viewer.htm#titlepage.htm
The Norwegian Regulations on Sustainable Forestry (2006) www.lovdata.no/dokument/SF/forskrift/2006-06-07-593
Tirén L (1935) Om granens kottsättning, dess periodicitet och samband med temperatur och nederbörd. Meddn St Skogförs Anst 28:413–524
Tveite B (1977) Bonitetskurver for gran. Medd Nor Inst Skogforsk 33:1–84
Wagner RG, Little KM, Richardson B, McNabb K (2006) The role of vegetation management for enhanching productivity of the world’s forests. Foresty 79:57–79
Winsa H (1995) Influence of rain shelter and site preparation on seedling emergence of Pinus sylvestris L. after direct seeding. Scand J For Res 10:167–175
Winsa H, Bergsten U (1994) Direct seeding of Pinus sylvestris using microsite preparation and invigorated seed lots of different quality: 2-year results. Can J For Res 24:77–86
Wooldridge JM (2010) Econometric analysis of cross section and panel data, 2nd edn. The MIT Press
Acknowledgements
We wish to thank Frida Skovdahl Hovland, Marit Helgheim, Rasmus Stokkeland, Benedikte Watne Oliver, Håvard Dufseth, Espen Martinsen and Eldar Paulsrud for valuable technical assistance. The study was carried out at the property of Romedal and Stange Commons (RASA), their hospitality and practical assistance are greatly appreciated. We would also like to acknowledge two anonymous reviewers who provided helpful comments on the manuscript. The study was supported by grants from The County Forest Officers in the counties Hedmark, Oppland, Sør-Trøndelag and Nord-Trøndelag, the Development Fund for Forestry (Project 209540), the Norwegian Forestry Fund (Skogtiltaksfondet), the Norwegian Agricultural Agency and Stange municipality.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Fløistad, I.S., Hylen, G., Hanssen, K.H. et al. Germination and seedling establishment of Norway spruce (Picea abies) after clear-cutting is affected by timing of soil scarification. New Forests 49, 231–247 (2018). https://doi.org/10.1007/s11056-017-9616-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11056-017-9616-2