
Abstract
Background:
Diabetic patients infected with coronavirus disease 2019 (COVID-19) often have a higher probability of organ failure and mortality. The potential cellular mechanisms through which blood glucose exacerbates tissue damage due to severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) infection is still unclear.
Methods and results:
We cultured endothelial cells within differing glucose mediums with an increasing concentration gradient of SARS-CoV-2 Spike protein (S protein). S protein can cause the reduction of ACE2 and TMPRSS2, and activation of NOX2 and NOX4. A high glucose medium was shown to aggravate the decrease of ACE2 and activation of NOX2 and NOX4 in cultured cells, but had no effect on TMPRSS2. S protein mediated activation of the ACE2-NOX axis induced oxidative stress and apoptosis within endothelial cells, leading to cellular dysfunction via the reduction of NO and tight junction proteins which may collectively be exacerbated by elevated glucose. In addition, the glucose variability model demonstrated activation of the ACE2-NOX axis in a similar manner observed in the high glucose model in vitro.
Conclusions:
Our present study provides evidence for a mechanism through which hyperglycemia aggravates endothelial cell injury resulting from S protein mediated activation of the ACE2-NOX axis. Our research thus highlights the importance of strict monitoring and control of blood glucose levels within the context of COVID-19 treatment to potentially improve clinical outcomes.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
COVID-19 is a highly infectious disease caused by the acute respiratory syndrome coronavirus 2 (SARS-CoV-2) [1]. As of November 21, 2022, infections associated with the COVID-19 pandemic has reached 763,740,140 people [2]. Patients with COVID-19 exhibit a systemic inflammatory cytokine storm characterized by the recruitment of inflammatory leukocytes by the vascular epithelium resulting in tissue damage and cytokine release. Consequently, such factors are major contributors of acute respiratory distress syndrome (ARDS), disseminated intravascular coagulation, and cardiovascular complications in COVID-19 [3]. Evidence linking endothelial cells (ECs) to SARS-CoV-2 infection includes the expression and function of its receptor, angiotensin-converting enzyme 2 (ACE2) within the vasculature [4, 5]. SARS-CoV-2 Spike protein (S protein) binds to ACE2 located on the cell membrane facilitating viral binding and fusion of the virus within the cellular membrane for host cell entry [6]. Previous studies have shown that S protein could induce endothelial dysfunction [7], triggering pulmonary immune thrombosis resulting in coagulopathy observed in patients with COVID-19 [8].
A history of diabetes or elevated blood glucose correlates with worse prognosis and higher mortality following COVID-19 infection [9,10,11]. While several studies over the course of the pandemic have reported the association between hyperglycemia and worsened outcomes in COVID-19 patients with and without known diabetes [12, 13], little is known about the cellular mechanisms through which blood glucose exacerbates tissue damage due to SARS-CoV-2 infection [14]. In the present study, we examined the relationship of glucose with S protein on human endothelial cells in culture. Our key findings suggest the synergistic effect of hyperglycemia and SARS-CoV-2 spike protein in promoting endothelial dysfunction via activation of the ACE2-NOX axis, which may account for the exacerbation of COVID-19 infection among hyperglycemic patients.
Materials and methods
Cell culture and plasmids transfection
Human umbilical vein endothelial cells (HUVECs) were purchased from the Cell Bank of the Chinese Academy of Sciences. The cells were cultured in minimum essential medium (HyClone) with 5 ng/ml human recombinant epidermal growth factor (Novus), 10% fetal bovine serum (Gibco), and a penicillin–streptomycin supplement. The cells were maintained in a humidified incubator at 37 °C and 5% CO2.
Normal glucose groups were cultured in 5 mmol/L glucose medium, high glucose groups were cultured in 20 mmol/L glucose medium. ACE2 overexpression plasmids were purchased from Public Protein/Plasmid Library (Nangjing, China). Apocynin (GC16237) was purchased from Gibco.
Antibodies and western blot analysis
The antibodies to ACE2 (#21115), TMPRSS2(#14437), NOX2(#19013), NOX4(#14347), and GAPDH (#60004) were obtained from Proteintech (Rosemont, IL). The antibodies to NOX1 (#DF8684), NOX3(#AF9134), ZO-1(#AF5145), and Occludin (#DF7504) were obtained from Affinity Biosciences (Cincinnati, OH). Total protein was extracted from HUVECs. Equal amounts of proteins were separated by SDS-PAGE and transferred to a nitrocellulose membrane (Millipore). After blocking with 5% nonfat milk, the membrane was incubated with specific antibody. All Western blotting analyses were repeated five times. Image J was used to quantify the protein expression.
DHE determination of ROS production
Freshly prepared HUVECs were incubated with freshly prepared dihydroethidium (DHE) (2 μmol/L) (Cat#: D7008, Sigma-Aldrich, St. Louis, MO, USA) in the dark for 30 min at room temperature. After washing three times, and fluorescent images were captured using a fluorescence microscope.
Intracellular no measurement by confocal microscopy
HUVECs or ACE2-overexpressing HUVECs were seeded and treated with the indicated agents within their respective group. Cells were washed twice with PBS and incubated with 5 μM DAF-FM DA at 37 °C for 25 min under a dark condition. The cells were then washed twice with cold PBS and analyzed via confocal microscopy at an excitation wavelength of 495 nm and an emission wavelength of 515 nm.
Flow cytometry
After treatment, 10^6 cells were collected, centrifuged and the supernatant was discarded. Cell death was detected by a commercialized apoptosis detection kit (C1062M, Beyotime). ROS was detected by a commercialized ROS kit (40778ES50, Yeasen). Fluorescence was analyzed using a flow cytometer.
Statistical analysis
All numerical data was expressed as mean ± standard error or median and interquartile range. Independent sample t-tests and Mann Whitney U tests were used to compare the continuous variables. Statistical analyses were performed using Statistical Package for the Social Sciences (version 22.0), and GraphPad Prism software (version 8.0). P < 0.05 was considered to be statistically significant.
Results
SARS CoV-2 spike membrane recombinant fusion protein downregulates ACE2 and increases NOX
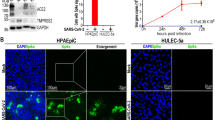
We first examined the effect of S protein exposure on ACE2 and TMPRSS2 in endothelial cells. HUVECs were treated with an increasing concentration gradient of S protein for 24 h. As shown in Fig. 1A-C, HUVECs treated with increasing S protein for 24 h exhibited a gradual decrease in ACE2 and TMPRSS2 protein expression.

Protein expression in different doses of S protein induced HUVECs. A. ACE2 and TMPRSS2 protein expression relative to GAPDH analyzed by western blot in different doses of S protein induced HUVECs. B. Relative expression of gray values for ACE2. C. Relative expression of gray values for TMPRSS2. D. NOX1 protein expression relative to GAPDH analyzed by western blot in different doses of S protein induced HUVECs. E. Relative expression of gray values for NOX1. F. NOX2 protein expression relative to GAPDH analyzed by western blot in different doses of S protein induced HUVECs. G. Relative expression of gray values for NOX2. H. NOX3 protein expression relative to GAPDH analyzed by western blot in different doses of S protein induced HUVECs. I. Relative expression of gray values for NOX3. J. NOX4 protein expression relative to GAPDH analyzed by western blot in different doses of S protein induced HUVECs. K. Relative expression of gray values for NOX4
Western blot analysis of NADPH oxidase (NOX) isoform expression indicated S protein induced NOX dependent oxidative stress in endothelial cells. The western blot results also showed that HUVECs exposed to an increasing S protein concentration gradient led to up-regulation of NOX2 and NOX4 expression but had no effect on NOX1 or NOX3 (Fig. 1D-K).
High glucose concentration aggravates S protein induced increase of ACE2 dependent NOX
In order to verify the regulatory relationship between ACE2 and NOX, HUVEC’s where treated with either an empty plasmid, an ACE2 overexpression plasmid or a NOX inhibitor in either the presence or absence of S protein for 24 h. Results showed that ACE2 overexpression significantly inhibited the increase of NOX expression, while NOX inhibitors had no effect on ACE2 protein expression. TMPRSS only decreased by the presence of S protein (Fig. 2A-E). Moreover, as shown in the Fig. 2F-J, results from HUVECs treated with normal glucose (NG), high glucose (HG) and S protein indicated that high glucose aggravated the change of ACE2-NOX axis caused by S protein. TMPRSS only responded a decrease to the presence of S protein.

Protein expression in S protein and different glucose induced HUVECs. A. Protein expression analyzed by western blot in different HUVECs induced by S protein. B–E. Relative expression of gray values for protein. F. Protein expression analyzed by western blot in different glucose induced by S protein. G–J. Relative expression of gray values for protein
High glucose aggravate ros and cell apoptosis caused by s protein
We next used DHE staining methods for the detection of reactive oxygen species (ROS) produced by S protein in different glucose mediums. Following 24 h of culture in HG medium, HUVECs showed no significant difference in ROS production compared to NG medium. However, after the addition of S protein, HUVEC’s exposed to HG produced a stronger red fluorescent signal, indicating higher ROS quantity (Fig. 3A). Flow cytometry results suggested statistically significant higher ROS quantity in HG treated cells compared to NG following the addition of S protein (Fig. 3BC). Moreover, detection of apoptosis via flow cytometry indicated no difference in the baseline apoptotic rate of HUVECs in either HG or NG conditions. Following the addition of S protein, the apoptotic rate of HG exposed cells was significantly higher than that in NG (Fig. 3DE).

ROS and apoptosis analysis for S protein and different glucose induced HUVECs. A. ROS analysis by DHE for different glucose induced by S protein. B. ROS analysis by flow cytometry for different glucose induced by S protein. C. Analysis of data from five independent experiments of B. D. Apoptosis analysis by flow cytometry for different glucose induced by S protein. E. Analysis of data from five independent experiments of D
High glucose aggravates endothelial cell dysfunction caused by s protein
Endothelial dysfunction coincides with a reduction in Nitric Oxide (NO) levels. Data generated from NO fluorescence probe in endothelial cells indicated that S protein led to significant reduction in NO concentration. Furthermore, ACE2 overexpression and inhibition of NOX coincided with downregulation of NO (Fig. 4A). The tight junction between HUVECs facilitates the formation of barriers and maintains normal barrier function. S protein reduced tight junction proteins through the ACE2-NOX axis (Fig. 4B-D). Results demonstrated that HUVESs cultured in HG medium showed significantly reduced production of NO in the presence of S protein via the ACE2-NOX axis, and accelerated degradation of tight junction proteins ZO-1 and Occludin (Fig. 4E–H).

Endothelial dysfunction analysis for S protein and different glucose induced HUVECs. A. NO production analysis for different HUVECs induced by S protein. B. ZO-1 and Occludin protein expression relative to GAPDH analyzed by western blot in different HUVECs induced by S protein. C, D. Relative expression of gray values for protein. E. NO production analysis for different glucose induced by S protein. B. ZO-1 and Occludin protein expression relative to GAPDH analyzed by western blot in different glucose induced by S protein. G, H. Relative expression of gray values for protein
Similar S protein mediated activation of the ACE2-NOX axis was shown in variable glucose and high glucose models
In order to simulate glycemic variability (GV) in vitro, we changed the medium of HUVEC’s every eight hours. Results showed no significant difference between GV, HG, and NG in their baseline values. Following the addition of S protein, ACE2-NOX axis activation was observed. Furthermore, the GV model showed similar results observed with continuous high glucose exposure and the presence of S protein, i.e., ACE2 expression was significantly decreased and NOX was significantly increased. Additionally, TMPRSS expression exhibited only S protein dependence, which was not related to glucose concentration (Fig. 5A-E).

Protein expression in different glucose induced HUVECs. A. Protein expression relative to GAPDH analyzed by western blot in different glucose induced HUVECs. B–E. Relative expression of gray values for protein
Discussion
This study demonstrates binding of S protein to endothelial cells requires ACE2, and TMPRSS. Additionally, S protein was shown to activate NOX 2 and NOX 4 in a dose-dependent manner. High glucose conditions significantly aggravate the expression of the ACE2-NOX axis, resulting in increased S protein-induced oxidative stress and apoptosis. These conditions result in reduced NO and tight junction proteins thus contributing to endothelial cell dysfunction (Supplemental Fig 1). Additionally, our research demonstrates that both variable glucose and high glucose concentrations led to significant activation of the ACE2-NOX axis in vitro.
This study sheds light on the mechanism behind the susceptibility of diabetic patients to SARS CoV-2 infection. Following infection, S protein is cut and activated by TMPRSS2 at the S1 and S2 sites, followed by infusion within the cell membrane [15] and subsequent binding of S protein to ACE2, triggering endocytosis [16]. Previous studies have reported endocytosis of S protein into cells is significantly enhanced in patients with diabetes [17]. Our study indicates that ACE2 and TMPRSS2 of endothelial cells are consumed in a dose–response manner after the addition of S protein. Additionally, the high glucose medium significantly increased the intake of ACE2 induced by S protein, with no effect on TMPRSS2 consumption. Based on our findings, we suspect the susceptibility of diabetic patients to S protein may be due to increased interaction between S protein and the extracellular domain of ACE2 after the S protein is cut, thus triggering clathrin-dependent endocytosis of the complex.
NOX catalyzes the deprotonation of superoxide anion into hydrogen peroxide and water, which produces toxic ROS. Relevant research has shown that ROS and oxidative stress produced by NOXs are involved in the pathogenicity of viruses [18]. Additionally, the down regulation of ACE2-ang (1–7)-MAS axis has been shown to play an important role in COVID-19 infection and its complications [17]. Other studies have shown that NOXs and ACE2/Ang (1–7)/Mas receptor axes are highly dependent [19]. Results from the present study demonstrate that S protein can induce the dose–response increase of NOX2 and NOX4 dependent to ACE2 in-vitro. Activation of NOX leads to uncoupling of endothelial NO synthase and the production of excessive ROS, thus reducing the production of NO and increases its consumption within the cell membrane, thereby decreasing bioavailable NO which in turn contributes to endothelial cell dysfunction [20]. Intercellular tight junctions composed of ZO-1, Occludin, and other proteins are critical for building the epithelial barrier and maintaining epithelial polarity [21]. Decreased tight junction proteins is an indicator of endothelial dysfunction. In our present study high glucose conditions is thought to aggravate the ACE2-NOX axis induced reduction of NO and tight junction protein expression indicated in endothelial dysfunction.
The in-vitro high glucose concentration model provides insight into the correlation with poor patient prognosis on the cooccurrence of SARS-CoV-2 infection and diabetes. A previous meta-analysis of 33 studies showed that diabetes may double the mortality and severity of COVID-19 [22]. Our study employed a high glucose density medium to simulate high glucose concentration in vitro, and showed that high glucose concentration facilitates S protein mediated activation of ACE2-NOX axis, ROS production, and endothelial cell dysfunction. Furthermore, in the absence S protein, high glucose concentration lead to only slight increases in ROS, with no significant statistical differences observed after repeated experimentation. In addition, no statistical difference was observed between endothelial cell dysfunction and apoptosis rate between normal glucose and high glucose concentration groups. This difference from our hypothesis may be due to the usage of 20 mmol/l glucose to culture cells for 24 h, as relevant studies have shown that high glucose concentration models with longer culturing time and higher concentration can lead to changes of adhesion molecules and oxidative stress [23,24,25].
Finally, we sought to simulated blood glucose variability in vitro by changing the medium every 8 h. Our results demonstrated glycemic variation induced increased activation of the ACE2-NOX axis following addition of S protein, with similar results observed in the high glucose concentration model. This indicates a correlation between glucose variation and poor physiological response following SARS-CoV-2 infection. Though further research on the mechanism is needed, the present results suggest stricter blood glucose control schemes are necessary for the treatment of COVID-19, and that intermittent blood glucose control may not be sufficient to improve the prognosis of COVID-19 patients.
There are limitations to this study. First, our research focused on the response of endothelial cells to S protein. As in clinical practices, SARS-CoV-2 does not only have pathogenic ability via the role S protein, it also demonstrates a subsequent inflammatory cytokine storm and multi organ crosstalk cascade reaction. The present research did not take account into these effects. Additionally, our research did not examine the dynamic changes of such results over time. Lastly, the Glycemic Variation model is only set by changing the medium every 8 h, which fails to more appropriately simulate physiological fluctuations in blood glucose levels in patients.
In conclusion, our study indicates that high glucose intensifies S protein mediated activation of the ACE2-NOX axis, resulting in increased ROS production as well as endothelial cell dysfunction and apoptosis. The GV model observed the same synergistic effect with S protein as the HG in vitro model. Collectively, the study yields insights for possible improvement in the treatment of COVID-19, indicating that blood glucose should be strictly controlled to improve the prognosis.
Data availability
All data generated or analysis during this study are included in this published article [and its supplementary information files].
References
Madhi SA, Baillie V, Cutland CL et al (2021) Efficacy of the ChAdOx1 nCoV-19 Covid-19 vaccine against the B.1.351 variant. N Engl J Med 384:1885–1898
World Health Organization https://covid19.who.int/
Evans PC, Rainger GE, Mason JC et al (2020) Endothelial dysfunction in COVID-19: a position paper of the ESC working group for atherosclerosis and vascular biology, and the ESC council of basic cardiovascular science. Cardiovasc Res 116:2177–2184
Klouda T, Hao Y, Kim H et al (2022) Interferon-alpha or -beta facilitates SARS-CoV-2 pulmonary vascular infection by inducing ACE2. Angiogenesis 25:225–240
Barbosa LC, Goncalves TL, de Araujo LP, Rosario LVO, Ferrer VP (2021) Endothelial cells and SARS-CoV-2: An intimate relationship. Vascul Pharmacol 137:106829
Buchrieser J, Dufloo J, Hubert M et al (2020) Syncytia formation by SARS-CoV-2-infected cells. EMBO J 39:e106267
Bernard I, Limonta D, Mahal LK, Hobman TC (2020) Endothelium Infection and Dysregulation by SARS-CoV-2: Evidence and Caveats in COVID-19. Viruses 13(1):29
McGonagle D, O’Donnell JS, Sharif K, Emery P, Bridgewood C (2020) Immune mechanisms of pulmonary intravascular coagulopathy in COVID-19 pneumonia. Lancet Rheumatol 2:e437–e445
Paolisso P, Bergamaschi L, Rambaldi P et al (2021) Impact of Admission Hyperglycemia on Heart Failure Events and Mortality in Patients With Takotsubo Syndrome at Long-term Follow-up: Data From HIGH-GLUCOTAKO Investigators. Diabetes Care 44:2158–2161
Wu J, Zhang J, Sun X et al (2020) Influence of diabetes mellitus on the severity and fatality of SARS-CoV-2 (COVID-19) infection. Diabetes Obes Metab 22:1907–1914
Li H, Tian S, Chen T et al (2020) Newly diagnosed diabetes is associated with a higher risk of mortality than known diabetes in hospitalized patients with COVID-19. Diabetes Obes Metab 22:1897–1906
Khunti K, Del Prato S, Mathieu C, Kahn SE, Gabbay RA, Buse JB (2021) COVID-19, Hyperglycemia, and New-Onset Diabetes. Diabetes Care 44:2645–2655
Vas P, Hopkins D, Feher M, Rubino F, M BW, (2020) Diabetes, obesity and COVID-19: A complex interplay. Diabetes Obes Metab 22:1892–1896
Liu D, Tang Y, Zhang Q (2021) Admission hyperglycemia predicts long-term mortality in critically Ill patients with subarachnoid hemorrhage: a retrospective analysis of the MIMIC-III database. Front Neurol 12:678998
Hoffmann M, Kleine-Weber H, Schroeder S et al (2020) SARS-CoV-2 cell entry depends on ACE2 and TMPRSS2 and Is blocked by a clinically proven protease inhibitor. Cell 181(271–280):e278
Chen Y, Guo Y, Pan Y, Zhao ZJ (2020) Structure analysis of the receptor binding of 2019-nCoV. Biochem Biophys Res Commun 525(1):135–140
Gross S, Jahn C, Cushman S, Bar C, Thum T (2020) SARS-CoV-2 receptor ACE2-dependent implications on the cardiovascular system: from basic science to clinical implications. J Mol Cell Cardiol 144:47–53
Violi F, Oliva A, Cangemi R et al (2020) Nox2 activation in Covid-19. Redox Biol 36:101655
Damiano S, Sozio C, La Rosa G, Santillo M (2020) NOX-Dependent signaling dysregulation in severe COVID-19: clues to effective treatments. Front Cell Infect Microbiol 10:608435
Meza CA, La Favor JD, Kim DH, Hickner RC (2019) Endothelial dysfunction: is there a hyperglycemia-induced imbalance of NOX and NOS? Int J Mol Sci 20(15):3775
Otani T, Furuse M (2020) Tight junction structure and function revisited. Trends Cell Biol 30:805–817
Kumar A, Arora A, Sharma P et al (2020) Is diabetes mellitus associated with mortality and severity of COVID-19? a meta-analysis. Diabetes Metab Syndr 14:535–545
La Sala L, Pujadas G, De Nigris V et al (2015) Oscillating glucose and constant high glucose induce endoglin expression in endothelial cells: the role of oxidative stress. Acta Diabetol 52:505–512
Quagliaro L, Piconi L, Assaloni R, Martinelli L, Motz E, Ceriello A (2003) Intermittent high glucose enhances apoptosis related to oxidative stress in human umbilical vein endothelial cells: the role of protein kinase C and NAD(P)H-oxidase activation. Diabetes 52:2795–2804
Ceriello A, De Nigris V, Prattichizzo F (2020) Why is hyperglycaemia worsening COVID-19 and its prognosis? Diabetes Obes Metab 22:1951–1952
Acknowledgements
None
Funding
This work was supported by the National Natural Science Foundation (No. 82102298 to Dr Su), Innovation Cultivation Foundation of Wuhan University/Zhongnan Hospital (No. 413000345/CXPY2020017 to Dr Su) and Research Project Foundation of Zhongnan Hospital (No. ZNYB2020013 to Dr Su). National Natural Science Foundation of China, 82102298, Lianjiu Su, Innovation Cultivation Foundation of Wuhan University/Zhongnan Hospital, 413000345/CXPY2020017, Lianjiu Su,Research Project Foundation of Zhongnan Hospital, ZNYB2020013, Lianjiu Su
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Conflicts of interest
The authors have no conflicts of interest to declare relevant to this publication.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Zhang, J., He, L., Huang, R. et al. Synergistic effect of elevated glucose levels with SARS-CoV-2 spike protein induced NOX-dependent ROS production in endothelial cells. Mol Biol Rep 50, 6039–6047 (2023). https://doi.org/10.1007/s11033-023-08504-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11033-023-08504-3