
Abstract
Murine macrophages of the J774A.1 line are hydrogen sulphide-producing cells with the primary role of γ-cystathionase (CTH) and secondary role of 3-mercaptopyruvate sulfurtransferase (limited by cysteine availability) and with a negligible role of cystathionine β-synthase (CBS) in H2S generation. J774A.1 cells stimulation with lipopolysaccharide (LPS) or interferon-gamma (IFNγ) resulted in decreased H2S levels after 24 h of incubation; however, they were restored to the control level after 48 h. Negligible CBS expression and activity in J774A.1 cells can result in homocysteine availability for CTH-catalyzed, H2S-generating reactions. This was supported by an increased CTH expression (IFNγ, 24 h and 48 h, and LPS, 48 h) and activity (24 h, LPS) in the stimulated cells. The results confirm the suggested feedback regulation between CBS and CTH.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Methionine is metabolized to homocysteine (HCys) to produce cysteine, which is a substrate of two enzymes—cystathionine β-synthase (CBS) and γ-cystathionase (CTH)—involved in endogenous production of H2S (Scheme 1) [1]. In cells with low CBS expression, H2S can be alternatively generated by CTH-catalyzed condensation reaction between cysteine and HCys or between two molecules of HCys (Scheme 1) [2].

Hydrogen sulfide generation and effect on superoxide levels in cells with low CBS expression. DHLA dihydrolipoic acid, MPST 3-mercaptopyruvate sulfurtransferase, Trx thioredoxin, CBS cystathionine β-synthase, CTH γ-cystathionase.  Limited by CBS expression and activity.
Limited by CBS expression and activity.  Limited by cysteine availability
Limited by cysteine availability
Another endogenous source of H2S is cysteine transamination by cysteine aminotransferase (CAT) and subsequent 3-mercaptopyruvate (3MP) conversion by 3-mercaptopyruvate sulfurtransferase (MPST) [3]. 3MP provides sulfur to the active-site cysteine residue of MPST to produce persulfide, which releases H2S in the presence of thioredoxin (Trx), abundant in cells. Another physiological reducing disulfide is dihydrolipoic acid (DHLA), cofactor for mitochondrial α-ketacid dehydrogenases. Trx and DHLA release H2S from persulfide provided by 3MP at the active site of MPST [4]. The expression of CBS, CTH and MPST are tissue-specific.
Hydrogen sulfide (H2S) plays an important role in inflammation [5] but there is no clear consensus as to its precise role in inflammatory signaling. Macrophages in general play important roles in the initiation and progression of many chronic inflammatory diseases. Li et al. [6] observed biphasic effects of H2S on inflammatory signals in murine lipopolysaccharide (LPS)-treated macrophages, where low H2S levels inhibited LPS-induced synthesis of PGE2, NO, IL-1β, IL-6 and NF-κB activity, but higher NaHS concentrations promoted synthesis of pro-inflammatory mediators.
LPS—a potent activator of macrophages, and interferon-gamma (IFNγ)—the principal macrophage-activating factor, was used to stimulate murine macrophages of the J774A.1 cell line. H2S measured by the zinc acetate-trapping method [7] was used to compare the H2S level in the control, and LPS- and IFNγ-stimulated cells after 24 and 48 h of incubation. The expression and the activity of hydrogen sulfide-generating enzymes, i.e. CBS, MPST, CTH, and TST, were investigated in J774A.1 cells. The results indicate an important role of CTH in the generation of hydrogen sulfide in J774A.1 cells with low activity of the transsulfuration pathway catalyzed by the CBS/CTH enzymatic system. They also confirm a negative feedback regulation between CBS and CSE [8].
The studies presented in this paper were designed to determine the activity and expression of hydrogen sulfide generating enzymes, i.e.: cystathionine β-synthase (CBS), γ-cystathionase (CTH) and MPST in murine macrophages stimulated with LPS and IFN-γ.
Materials and methods
Chemicals
1-Fluoro-2,4-dinitrobenzene, bathophenanthroline-disulfonic acid disodium salt, acetonitrile, pyridoxal phosphate (PLP), β-nicotinamide adenine dinucleotide reduced disodium salt hydrate (NADH), l-lactic dehydrogenase (LDH), 3-mercaptopyruvate acid sodium salt, d,l-dithiothreitol (DTT), N-ethylmaleimide (NEM), dl-propargylglycine (PPG), NaH2PO4·2H2O Na2SO3, chloroform, isopropanol, agarose, sodium hydrosulfide hydrate, NaCl, Folin–Ciocalteu’s phenol reagent, Fe(NO3)3·9H2O, Na2S2O3·5H2O, Na2CO3 and N,N-dimethyl-p-phenylenediamine sulfate salt (DMPPDA) were obtained from Sigma-Aldrich (St. Louis, MO, USA). Trifluoroacetic acid and 2-mercaptoethanol were purchased from Fluka Chemie GmbH (Buchs, Switzerland). Ethanol and 70% perchloric acid 38% formaldehyde, 65% HNO3, 38% HCl, ammonia solution 25%, sodium potassium tarate, copper sulphate pentahydrate (CuSO4·5H2O) potassium dihydrogen phosphate (KH2PO4), ferric chloride (FeCl3), (Zn(CH3COO)2·2H2O), and sodium hydroxide (NaOH) were from Polskie Odczynniki Chemiczne S.A. (Gliwice, Poland). Nε-methyllysine was obtained from Bachem (Bubendorf, Switzerland). DMEM/high glucose (Lonza, Basel, Switzerland), fetal bovine serum and penicillin–streptomycin solution were purchased from Gibco Invitrogen, Life Technologies, (Grand Island, NY, USA). Trizol, ethidium bromide and EDTA–disodium salt dihydrate were obtained from Lab-Empire (Rzeszow, Poland). KCN was obtained from Merck (Darmstadt, Germany). Reverse transcriptase M-MuLV was obtained from Promega (Madison, WI, USA). Polymerase DNA Dream Taq™, Gene Ruler 100 bp DNA Ladder, Oligo(dT)18 primer and dNTP mix were obtained from Abo (Gdańsk, Poland). All the chemicals were analytical grade and HPLC solvents were gradient grade. Water was deionized by passing through an EASY pure RF compact ultrapure water system.
Cell culture
Mouse BALB/c monocyte macrophage of the J774A.1 cell line (ATCC, Manassas, VA, USA) were grown in DMEM medium supplemented with 10% FCS, and antibiotics (100 U/ml penicillin and 100 µg/ml streptomycin), in sterile, plastic culture flasks (BD Falcon, Franklin Lakes, NJ, USA), at 37 °C, in a humidified atmosphere containing 5% CO2. For experiments, the cells were cultured in DMEM medium supplemented with 5% FCS and antibiotics (as above) at 37 °C, in a humidified atmosphere containing 5% CO2, either on sterile, plastic Petri dishes (100 mm in diameter) or on sterile, plastic, 24-well, flat-bottom plates. J774A.1 cells were stimulated for 24 h and 48 h with LPS (100 ng/5 × 105 cells/ml; Sigma Aldrich, St. Louis, MO, USA) or with recombinant IFNγ (100 U/5 × 105 cells/ml; PeproTech, London, UK). When necessary, after a certain time of stimulation, dry cell pellets were frozen and stored at − 80 °C prior to RNA isolation.
The release of TNFα and IL-6 by J774A.1 cells stimulated with IFNγ and LPS
J774A.1 cells (5 × 105 cells/ml/well), cultured on sterile, plastic, 24-well, flat-bottom plates at 37 °C and 5% CO2, were stimulated with either LPS (100 ng/ml) or IFNγ (100 U/ml), as described above. After 24 h, cell-free culture supernatants were collected, frozen and stored at − 20 °C for further detection of cytokine concentrations. TNFα and IL-6 concentrations were measured in ELISA according to procedures delivered by manufacturers with ELISA kits, i.e. mouse TNFα ELISA Ready-SET-Go!® (Bioscience, Inc., San Diego, CA, USA) and mouse IL-6 BD OptEIA™ Set (BD Biosciences, San Diego, CA, USA).
Expression of MPST, CTH, CBS, and rhodanese (TST) in J774A.1 cells
Isolation of total RNA
Total RNA was extracted with Trizol reagent (Lab-Empire, Poland) following the manufacturer’s instructions and RNA quantity and integrity were verified by agarose gel electrophoresis. Isolated RNA was stored at − 80 °C and was used for RT-PCR.
Reverse transcription of RNA
Total RNA (3 µg) was reversely transcribed with 1 µl of GoScript™ Reverse Transcriptase (160 U/µl; Promega Corporation), 4 µl of GoScript™ 5 × reaction buffer (Promega Corporation), 3 µl of MgCl2, 1 µl of RNase Inhibitor (20 U/µl; Thermo Scientific), 1 µl of dNTP mix (10 mM; Thermo Scientific), and 1 µl of GoScript™ Reverse Transcriptase (160 U/µl; Promega Corporation) in 20 µl of final volume of reaction mixture. RNA was mixed with Oligo d(T) primer and was heated for 5 min at 70 °C. Then, samples were incubated in the reaction mixture for 5 min at 25 °C, 60 min at 42 °C and 15 min at 70 °C.
Polymerase chain reaction (PCR)
PCR was performed using 2 µl of cDNA, 0.2 µM of each primer, 0.04 U/µl of DNA polymerase (Thermo Scientific) in 10 mM buffer Tris–HCl pH 8.8 (supplemented with 1.5 mM MgCl2, 50 mM KCl, 0.1% Triton X-100), 0.2 mM of dNTP mix (Thermo Scientific) and H2O–DEPC in total reaction volume of 25 µl. The primer sequences are described in Table 1.
For CTH, after an initial 5 min denaturation at 94 °C, amplification was performed under the following conditions: 94 °C for 30 s, 57 °C for 30 s, and 72 °C for 2 min for 35 cycles, with a final incubation at 72 °C for 8 min.
For CBS, after an initial 5 min denaturation at 94 °C, amplification was performed under the following conditions: 94 °C for 30 s, 56 °C for 30 s, and 72 °C for 2 min for 35 cycles, with a final incubation at 72 °C for 8 min.
For MPST, after an initial 5 min denaturation at 94 °C, amplification was performed under the following conditions: 94 °C for 30 s, 54 °C for 30 s, and 72 °C for 2 min for 30 cycles, with a final incubation at 72 °C for 8 min [9].
For TST, after an initial 5 min denaturation at 95 °C, amplification was performed under the following conditions: 95 °C for 30 s, 55 °C for 30 s, and 72 °C for 2 min for 34 cycles, with a final incubation at 72 °C for 8 min.
For β-actin, after an initial 5 min denaturation at 94 °C, amplification was performed under the following conditions: 94 °C for 30 s, 55 °C for 35 s, and 72 °C for 35 s for 32 cycles, with a final incubation at 72 °C for 8 min [10].
β-actin was used as an internal standard to normalize mRNA abundance. PCR products were analyzed by 2.0% agarose gel electrophoresis and imaged by UVI-KS 4000i/ImagePC (Syngen Biotech, Poland).
Detection of H2S
H2S produced during the incubation of the cell culture with H2S-releasing compounds was trapped as zinc sulfide in the zinc agarose layer according to [7]. The standard curve was linear at the concentration range of 0–250 µM with a correlation coefficient of 0.994.
Enzymes assay
Cell homogenization
Murine macrophage cells (line J774A.1) (3.5–5 × 106 cells/pellet) were resuspended in 0.1 M phosphate buffer pH 7.5, in the proportion 1 × 106 cells/0.07 ml of the buffer, sonicated 3 × 5 s at 4 °C (Bandelin Sonoplus GM 70). After centrifugation at 1600 × g for 10 min, the supernatant was used for the determination of protein concentration, sulfane sulfur levels and the activity of MPST, CTH and rhodanese. For reverse phase high performance liquid chromatography (RP-HPLC) analyses, the cells were resuspended in 0.1 ml 10% PCA/1 mM BPDS. The sediment was separated by centrifugation at 1400 × g for 10 min, and the supernatant was stored at − 80 °C until analysis.
MPST activity
The MPST activity was assayed according to the method of Valentine and Frankelfeld 1974 [13] with some modification as described by Wróbel et al. [14] and Bronowicka-Adamska et al. [15]. The enzyme activity was expressed as nmoles of pyruvate produced during 1 min incubation at 37 °C/1 mg of protein.
CTH activity
The cystathionase activity was determined by Matsuo and Greenberg [16] with modifications described by Czubak et al. [17]. The difference between the initial value of absorbance (before adding LDH) and the lowest value (after adding LDH) corresponded to the amount of alpha-ketobutyrate formed in the course of the cystathionase reaction. The cystathionase activity was expressed as nmoles of α-ketobutyrate formed during 1 min incubation at 37 °C/1 mg of protein.
CBS activity
The activity of CBS was examined in cell homogenates in the presence of dl-homoserine as a substrate after 15 min incubation at 37 °C according to the description by Bronowicka-Adamska et al. [18]. The level of cystathionine was determined using the HPLC method described by Bronowicka-Adamska et al. [19]. The CBS activity was expressed as pmoles of cystathionine formed during 1 min incubation at 37 °C/1 mg of protein.
Sulfane sulfur level
Sulfane sulfur was determined by the method of Wood [20] following the procedure described by Bronowicka-Adamska et al. [15]. The method is based on cold cyanolysis and colorimetric detection of ferric thiocyanate complex ion. The sulfane sulfur level was expressed as nmoles of SCN− produced/1 mg of protein.
Protein level
Protein concentration was determined by the method described by Lowry et al. [21] using crystalline bovine serum albumin as a standard.
RP-HPLC (reverse phase high performance liquid chromatography)
The levels of cystathionine in the incubation mixtures were determined using the RP-HPLC method Dominik et al. [22] with modifications [19].
Statistical analysis
All the results from at least three independent experiments, 15–25 individual determinations, were expressed as mean ± SD. The significance of the differences between the controls and investigated samples were calculated using is a nonparametric Mann–Whitney U test (p < 0.05).
Results
The release of TNFα and IL-6 by J774A.1 cells stimulated with IFNγ or LPS
After stimulation with both factors, macrophages released proinflammatory mediators. The effects of LPS stimulation was weaker than IFNγ (Table 2).
H2S levels in control, and LPS and IFNγ-stimulated J774A.1 cells
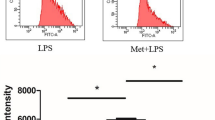
The level of hydrogen sulphide was significantly lower after 24 h in case of both factors (Fig. 1). It was restored to the control levels after 48 h of incubation.

H2S production in LPS and IFNγ-stimulated J774A.1 cells after 24 h and 48 h of stimulation. *p < 0.05 LPS, IFNγ versus the controls (Mann–Whitney U test). The control levels were 7.2 ± 1.5 µM H2S/h for 24 h and 6.3 ± 1.08 µM H2S/h for 48 h
The expression of MPST, CTH, CBS and TST in murine macrophage cells (J774A.1 cells)
After 24 h incubation the control cells, not stimulated (Fig. 2a, line 1), express γ-cystathionase (CTH) and MPST, two H2S-forming enzymes. Low TST expression and only trace expression of cystathionine β-synthase (CBS) was found. Similarly after 48 h (Fig. 2a, line 4) with the exception of higher expression of TST, as compare to 24 h, and lower expression of CTH.

RT-PCR analysis of MPST, CTH, CBS, TST in J774A.1 cells. a One set of representative results is shown. β-Actin was used as the internal control. 1 Control 24 h, 2 100 U IFNγ, 24 h, 3 100 ng/ml LPS, 24 h, 4 control, 48 h, 5 100 U IFNγ, 48 h, 6 100 ng/ml LPS, 48 h. b The relative expression level of MPST, CTH, CBS, TST in J774A.1 cells. Densities of bands were normalized using β-actin, the mean of which was taken as equal to one. *p < 0.05 LPS, IFNγ versus control (Mann–Whitney U test). #The density of CBS after 24 h was out of detection
CBS expression did not change significantly in the LPS-stimulated cells and it seemed even to drop in the IFNγ-stimulated cells (Fig. 2a, b). The expression of MPST significantly decreased during incubation with 100 U IFNγ and 100 ng/ml LPS. IFNγ, in turn, up-regulated CTH expression in J774A.1 cells after both 24 h and 48 h stimulation, in comparison to the control cells (Fig. 2a, b).
The activity of MPST, CTH, TST and the level of sulfane sulfur in J774A.1 cells after stimulation
CBS activity in both the control and stimulated cells was undetectable. The activity of MPST (24 h) and rhodanese (24 h and 48 h) was significantly decreased after incubation with IFNγ and LPS after 48 h, in comparison to the control cells (Table 3A, B). Significantly increased CTH activity was determined after 24 h and 48 h stimulation with LPS (Table 3A, B) but in case of IFNγ it was below the detection limit after 24 h or significantly decreased after 48 h, as compare to control values. A homeostasis of the level of sulfane sulfur was observed—it remains unchanged independently of the compound used and time of incubation (Table 3A, B).
Discussion
Hydrogen sulfide (H2S) has been identified as a regulator of inflammatory responses [5].
The results have confirmed the expression of CTH and MPST in J774A.1 cells suggesting that macrophages are hydrogen sulphide producing cells. CBS seems to be of a low importance for H2S formation in J774A.1 cells.
Low CBS expression can result in HCys availability for CTH-catalyzed H2S generating reaction [2, 23] (Scheme 1). The IFNγ-stimulated production of NO was detected to result in a decrease in CBS activity [24] and it is attributed to a reduced rate of the transsulfuration pathway, CBS/CTH dependent, converting majority of HCys (Scheme 1). In the IFNγ-stimulated cells, an increased expression of CTH (Fig. 2a, b) after 24 h is correlated with a decreased - as compared to control—level of H2S. This can result from significantly decreased activity of CTH and TST observed after both 24 h and 48 h of incubation with IFNγ and MPST activity after 24 h (Table 3A, B), the activity of which is regulated through a redox-switch regulatory mechanism [25]. However, higher expression of CTH after 48 h, in comparison to 24 h, resulted in an increased level of hydrogen sulfide (Figs. 1, 2a, b).
In case of LPS, a higher production of H2S after 48 h, as compare to 24 h, is also correlated with higher increase in CTH expression after both IFNγ and LPS stimulation. An increased expression of CTH and H2S production in the macrophages stimulated with LPS was earlier reported [26]. A decreased expression and activity of MPST after 24 h (Fig. 2a, b; Table 3A) limits the amount of cysteine converted by this enzyme and make it more available for H2S-generating reactions catalyzed by CTH (Scheme 1). Interestingly, the expected increased level of HCys in J774A.1 cells with low CBS expression and activity, is associated with a high CTH expression. A decreased level of H2S after 24 h stimulation with both IFNγ and LPS was also associated with an increased CTH expression. These observations may confirm the suggested effect of HCys (increased due to low CBS level) and H2S on CTH expression, which is upregulated by high levels of HCys and low H2S levels and a negative feedback regulation between CBS and CTH [8]. Increased HCys level together with the elevated level of CTH (IFNγ, 48 h) can result in cysteine sucking into reactions with HCys and its lower availability for GSH synthesis. Decreased GSH levels result in an increased level of reactive oxygen species which are, in turn, involved in the redox regulation of immune cells [27].
H2S can be regarded an \({\text{O}}_{2}^{{\cdot - }}\) scavenger—it reacts with superoxide radical anion (Scheme 1). H2S is a reducing agent and a weak acid with approximately 4:1 HS−/H2S ratio at physiological pH [28]. HS−, as opposed to H2S, can reduce the metal center of cytochrome c and lead to production of \({\text{O}}_{2}^{{\cdot - }}\) from molecular oxygen [29]. On the other hand, HS−/H2S can reduce Cu2+ to Cu1+ and enhance \({\text{O}}_{2}^{{\cdot - }}\) scavenging activity of the copper–zinc SOD (CuZnSOD) [29, 30]. Sulfide is a potent reversible inhibitor of myeloperoxidase activity [31]. It has also been shown that HCys can induce Trx-1 expression in human monocytes [32] and in this way potentiate the antioxidant protection of cells (Scheme 1). Thus, in cells, H2S can act as a pro- or antioxidant, depending on the H2S/HS− ratio.
In stimulated cells, hydrogen sulphide can react with nitric oxide [33], what can result in decreased H2S levels, especially pronounced after 24 h, and particularly in the IFNγ-stimulated cells with a massive amount of pro-inflammatory mediators released (Table 1). On the other hand, H2S might also exert anti-inflammatory effects by inhibiting NO production [11]. H2S oxidation results in persulfides and thiosulfate formation [34], substrates of mitochondrial rhodanese. It catalyzes the transfer of sulfane sulfur atom from thiosulfate to various acceptors (e.g. GSH), leading to H2S, and allows for recycling H2S without consuming additional cysteine. However, it seems this does not occur due to its significantly decreased activity after 48 h stimulation with both IFNγ and LPS (Table 3A, B). On the other hand, a homeostasis of sulfane sulfur level was observed (Table 3A, B).
Conclusions
J774A.1 macrophage cells are hydrogen sulphide-producing cells with CTH as the main enzyme and MPST playing a supporting role. A low CBS expression and non-detectable CBS activity in the IFNγ-stimulated cells can result in HCys availability for CTH-catalyzed, H2S-generating reactions (Scheme 1). A decreased level of H2S after 24 h stimulation with both IFNγ and LPS associated with an increased CTH expression can confirm CTH upregulation by a high level of HCys (due to low CBS expression) and low H2S and a negative feedback regulation between CBS and CTH.
Abbreviations
- CBS:
-
Cystathionine β-synthase
- CTH:
-
γ-Cystathionase
- HCys:
-
Homocysteine
- IFNγ:
-
Interferon γ
- LPS:
-
Lipopolysaccharide
- MPST:
-
3-Mercaptopyruvate sulfurtransferase
- TST:
-
Rhodanese
References
Wang R (2002) Two’s company, three’s a crowd: Can H2S be the third endogenous gaseous transmitter? FASEB J 16:1792–1798
Sen U, Mishra PK, Tyagi N, Tyagi SC (2010) Homocysteine to hydrogen sulfide or hypertension. Cell Biochem Biophys 57:49–58
Shibuya N, Tanaka M, Yoshida M, Ogasawara Y, Togawa T, Ishii K, Kimura H (2009) 3-Mercaptopyruvate sulfurtransferase produces hydrogen sulfide and bound sulfane sulfur in the brain. Antioxid Redox Signal 11:703–714
Mikami Y, Shibuya N, Kimura Y, Nagahara N, Ogasawara Y, Kimura H (2011) Thioredoxin and dihydrolipoic acid are required for 3-mercaptopyruvate sulfurtransferase to produce hydrogen sulfide. Biochem J 439:479–485
Bhatia M (2012) Role of hydrogen sulfide in the pathology of inflammation. Scientifica. https://doi.org/10.6064/2012/159680
Li M, Whiteman L, Rose P, Tan CH, Parkinson DB, Moore PK (2010) The effect of hydrogen sulfide donors on lipopolysaccharide-induced formation of inflammatory mediators in macrophages. Antioxid Redox Signal 12:1147–1154
Kartha RV, Zhou J, Hovde LB, Belinda WY, Cheung BWY, Schröder H (2012) Enhanced detection of hydrogen sulfide generated in cell culture using an agar trap method. Anal Biochem 423:102–108
Nandi SS, Mishra PK (2017) H2S and homocysteine control a novel feedback regulation of cystathionine beta synthase and cystathionine gamma lyase in cardiomyocytes. Sci Rep 7(1): 3639
Jurkowska H, Placha W, Nagahara N, Wróbel M (2011) The expression and activity of cystathionine-γ-lyase and 3-mercaptopyruvate sulfurtransferase in human neoplastic cell lines. Amino Acids 41:151–158
Brunner M, Gruber M, Schmid D, Baran H, Moeslinger T (2015) Proliferation of macrophages due to the inhibition of inducible nitric oxide synthesis by oxidized low-density lipoproteins. EXCLI J 14:439–451
Oh GS, Pae HO, Lee BS, Kim BN, Kim JM, Kim HR, Jeon SB, Jeon WK, Chae HJ, Chung HT (2006) Hydrogen sulfide inhibits nitric oxide production and nuclear factor-kappaB via heme oxygenase-1 expression in RAW264.7 macrophages stimulated with lipopolysaccharide. Free Radic Biol Med 41:106–119
Shanmugam MK, Jing Z, Bhatia M (2009) Aminooxyacetate inhibits hydrogen sulfide and ammonium synthesis and protects mice in acute pancreatitis. Int J Integr Biol 8:7–14
Valentine WN, Frankelfeld JK (1974) 3-Mercaptopyruvate sulfurtransferase (EC 2.8.1.2): a simple assay adapted to human blood cells. Clin Chim Acta 14:05–210
Wróbel M, Jurkowska H, Śliwa L, Srebro Z (2004) Sulfurtransferases and cyanide detoxification in mouse liver, kidney, and brain. Toxicol Mech Methods 14:331–337
Bronowicka-Adamska P, Wróbel M, Magierowski M, Magierowska K, Kwiecień S, Brzozowski T (2017) Hydrogen sulphide production in healthy and ulcerated gastric mucosa of rats. Molecules 22:1–13
Matsuo Y, Greenberg DM (1958) A crystalline enzyme that cleaves homoserine and cystathionine. J Biol Chem 230:545–560
Czubak J, Wróbel M, Jurkowska H (2002) Cystathionine γ-lyase (EC 4.4.1.1): an enzymatic assay of α-ketobutyrate using lactate dehydrogenase. Acta Biol Cracov Ser Zool 44:113–117
Bronowicka-Adamska P, Wróbel M, Zagajewski J (2015) An application of RP-HPLC for P determination of the activity of cystathionine beta-synthase and gamma-cystathionase in tissue homogenates. Nitric Oxide 46:186–191
Bronowicka-Adamska P, Wróbel M, Zagajewski J (2011) RP-HPLC method for quantitative determination of cystathionine, cysteine and glutathione: an application for the study of the metabolism of cysteine in human brain. J Chromatogr B 879:2005–2009
Wood L (1987) Sulfane sulfur. Methods Enzymol 143:25–29
Lowry OH, Rosenbrough NJ, Farr AL, Randall RI (1951) Protein measurement with the Folin phenol reagent. J Biol Chem 193:265–275
Dominik PK, Cassidy PB, Roberts JC (2001) A new and versatile method for determination of thiolamines of biological importance. J Chromatogr B 761:1–12
Olson KR (2012) A practical look at the chemistry and biology of hydrogen sulfide. Antioxid Redox Signal 17:32–44
Prathapasinghe GA, Siow YL, Xu ZOK (2008) Inhibition of cystathionine-synthase activity during renal ischemia–reperfusion: role of pH and nitric oxide. Am J Physiol Ren Physiol 295:F912–F922
Nagahara N (2013) Regulation of mercaptopyruvate sulfurtransferase activity via intrasubunit and intersubunit redox-sensing switches. Antioxid Redox Signal 19:1792–1802
Zheng Y, Luo YN, Mu D, Jiang P, Liu R, Sun H, Xiong S, Liu X, Wang L, Chu Y (2013) Lipopolysaccharide regulates biosynthesis of cystathionine γ-lyase and hydrogen sulfide through toll-like receptor-4/p38 and toll-like receptor-4/NF-κB pathways in macrophages. In Vitro Cell Dev Biol Anim 49:679–688
Miljkovic D, Spasojevic I (2013) Multiple sclerosis: molecular mechanisms and therapeutic opportunities. Antioxid Redox Signal 19:2286–2334
Mijušković A, Oreščanin-Dušić Z, Nikolić-Kokić A, Slavić M, Spasić MB, Spasojević I, Blagojević D (2014) Comparison of the effects of methanethiol and sodium sulphide on uterine contractile activity. Pharmacol Rep 66:373–379
Bolić B, Mijušković A, Popović-Bijelić A, Nikolić-Kokić A, Spasić S, Blagojević D, Spasić MB, Spasojević I (2015) Reactions of superoxide dismutases with HS(−)/H2S and superoxide radical anion: an in vitro EPR study. Nitric Oxide 51:19–23
Searcy DG, Whitehead JP, Maroney MJ (1995) Interaction of Cu, Zn superoxide dismutase with hydrogen sulfide. Arch Biochem Biophys 318:251–263
Pálinkás Z, Furtmüller PG, Nagy A, Jakopitsch C, Pirker KF, Magierowski M, Jasnos K, Wallace JL, Obinger C, Nagy P (2015) Interactions of hydrogen sulfide with myeloperoxidase. Br J Pharmacol 172:1516–1532
Dai J, Wang X, Feng J, Kong W, Xu Q, Shen X, Wang X (2008) Regulatory role of thioredoxin in homocysteine-induced monocyte chemoattractant protein-1 secretion in monocytes/macrophages. FEBS Lett 582:3893–3898
Nagpure BV, Bian JS (2016) Interaction of hydrogen sulfide with nitric oxide in the cardiovascular system. Oxidative Med Cell Longev 2016:1–16
Libiad M, Yadav PK, Vitvitsky V, Martinov M, Banerjee R (2014) Organization of the human mitochondrial H2S oxidation pathway. J Biol Chem 289:30901–30910
Acknowledgements
This work was supported by a Grant from the National Committee for Scientific Research K/DSC/003590 and K/ZDS/007220.
Author information
Authors and Affiliations
Contributions
P.B.-A. provided the experimental data, analyzed the data, wrote the manuscript in consultation with M.W., P.S., A.G. provided the experimental data. H.J. performed and developed the RT-PCR analysis, contributed to the interpretation of the results. K.N. supervised and performed cell cultures. J. M. and M.W. discussed the results and contributed to the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no conflict of interest.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Bronowicka-Adamska, P., Jurkowska, H., Gawda, A. et al. Expression and activity of hydrogen sulfide generating enzymes in murine macrophages stimulated with lipopolysaccharide and interferon-γ. Mol Biol Rep 46, 2791–2798 (2019). https://doi.org/10.1007/s11033-019-04725-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11033-019-04725-7