
Abstract
Most prior climate change assessments for U.S. agriculture have focused on major world food crops such as wheat and maize. While useful from a national and global perspective, these results are not particularly relevant to the Northeastern U.S. agriculture economy, which is dominated by dairy milk production, and high-value horticultural crops such as apples (Malus domestica), grapes (Vitis vinifera), sweet corn (Zea mays var. rugosa), cabbage (Brassica oleracea var. capitata), and maple syrup (sugar maple, Acer saccharum). We used statistically downscaled climate projections generated by the HadCM3 atmosphere–ocean general circulation model, run with Intergovernmental Panel on Climate Change future emissions scenarios A1fi (higher) and B1 (lower), to evaluate several climate thresholds of direct relevance to agriculture in the region. A longer (frost-free) growing season could create new opportunities for farmers with enough capital to take risks on new crops (assuming a market for new crops can be developed). However, our results indicate that many crops will have yield losses associated with increased frequency of high temperature stress, inadequate winter chill period for optimum fruiting in spring, increased pressure from marginally over-wintering and/or invasive weeds, insects, or disease, or other factors. Weeds are likely to benefit more than cash crops from increasing atmospheric carbon dioxide. Projections of thermal heat index values for dairy cows indicate a substantial potential negative impact on milk production. At the higher compared to lower emissions scenario, negative climate change effects will occur sooner, and impact a larger geographic area within the region. Farmer adaptations to climate change will not be cost- or risk-free, and the impact on individual farm families and rural communities will depend on commodity produced, available capital, and timely, accurate climate projections.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
Agriculture is a major component of the Northeastern U.S. (NE) economy, with a value exceeding $7.5 billion per year http://www.nass.usda.gov/Census_of_Agriculture/index.asp). New York typically ranks within the top three states in the nation for production of apples (Malus domestica), grapes (Vitis vinifera and V. labruscana), fresh market sweet corn (Zea mays var. rugosa), snap beans (Phaseolus vulgaris), cabbage (Brassica oleracea var. capitata), milk, cottage cheese, and several other commodities. Vermont is famous for its maple syrup (sugar maple, Acer saccharum) and is the nation’s top producer of this specialty product. Throughout the NE, small family farms are vital to the economy of rural areas, and they fill an important market niche for fresh, high quality, affordable local produce.
Most analyses of climate change impact on U.S. agriculture are based on crop model simulations for one or more of the major world food crops such as wheat (Triticum aestivum) (e.g., Adams et al. 1995), field corn (maize, Z. mays var. indenta), soybean (Glycine max), and in some cases, potato (Solanum tuberosum) (e.g., Reilly 2002). The problem with this approach for the NE is that, with the exception of potato, these crops are relatively unimportant economically to the region compared to high value horticultural crops such as apples, grapes, cabbage, and sweet corn. The dairy industry, which dominates the NE agriculture economy ($1.5 billion per year for New York state alone), has not been well-addressed in prior analyses, although Klinedinst et al. (1993) conducted an evaluation for the entire U.S. using climate projections available at that time, and a more recent analysis for California (Hayhoe et al. 2004) addressed the dairy issue for that state. Another short-coming of many prior analyses is that potential effects of climate change on weeds, insects, and disease pests are not considered, as it is assumed that farmers will adapt with appropriate changes in control strategies as needed.
Well-tested simulation models are not available for most of the economically important crops and weed species in the NE. We can, however, examine climate projections for effects on the timing and frequency of threshold events that we know from the scientific literature will positively or negatively affect crops and livestock of economic importance to the region. Some crops and weeds will benefit by changing climate and increasing atmospheric carbon dioxide (CO2), at the expense of other species that do poorly and become less important in the region. A similar diversity of response to climate change will occur for insect and disease pests, and for their natural enemies. Climate change and CO2 will favor some invasive species, and will alter important interactions between plants and pollinators, insect pests, diseases, and weeds.
In this paper we review relevant past research, and describe results of a new analysis utilizing statistically downscaled climate projections generated by the UKMO HadCM3 model (Pope et al. 2000), run with Intergovernmental Panel on Climate Change (IPCC) emissions scenarios A1fi (higher) and B1 (lower) to assess several climate thresholds of direct relevance to agriculture in the region. (A similar analysis was conducted based on output from the DOE/NCAR PCM model (Washington et al. 2000), with results available at: http://www.northeastclimateimpacts.org). Climate thresholds we consider include: the frost-free period; the frequency of high temperature stress thresholds affecting crops and livestock; winter temperature effects on winter chill requirement for maximum yield of fruit and other crops minimum winter temperature effects on potential northward migration of invasive weeds and insect pests, and frequency of summer droughts. We conclude with a discussion of farmer adaptation options in relation to potential costs, risks, and vulnerabilities.
2 Methods
2.1 Climate modeling
The climate projections used in this assessment were based on simulations from the UKMO HadCM3 (Pope et al. 2000) atmosphere–ocean general circulation model (AOGCM). This is among the latest generation of numerical models that couple atmospheric, ocean, sea-ice, and land-surface components to represent historical climate variability and estimate projected long-term increases in global temperatures due to human emissions. Atmospheric processes are simulated at a grid resolution of 3.75 by 2.5°. The ability of the model to simulate regional atmospheric dynamics and surface climate patterns over the NE has been evaluated, with projections for NE temperature and precipitation presented in Hayhoe et al. (2007). We selected results from HadCM3 simulations, among several AOGCM options, for presentation here because its climate sensitivity – a metric that captures the magnitude of the model-simulated increase in global temperature in response to a doubling of atmospheric CO2 concentration – is 3.3°C, encompassing the middle part of the IPCC range of 1.5–4.5°C.
In addition to the HadCM3 results reported here, we also conducted future simulations with the DOE/NCAR PCM model (Washington et al. 2000), which has a grid resolution of approximately 2.8 by 2.8 degrees, and climate sensitivity to a doubling of atmospheric CO2 of 1.3°C, slightly below the IPCC range of 1.5–4.5°C. Those interested in comparing PCM with HadCM3 results are referred to: http://www.northeastclimateimpacts.org.
Future HacCM3 simulations presented here were forced by the A1fi and B1 scenarios as characterized in the IPCC Special Report on Emission Scenarios (SRES, Nakićenović et al. 2000). In the A1fi scenario, CO2 emissions climb throughout the century, reaching almost 30 Gt/year or 6 times 1990 levels by 2100. Emissions under the B1 scenario are lower, based on a world that transitions relatively rapidly to service and information economies. Carbon dioxide emissions in the B1 scenario peak at just below 10 Gt/year – around two times 1990 levels – at mid-century and decline slowly to below current-day levels. Together, these scenarios represent the range of IPCC non-intervention emissions futures with atmospheric CO2 concentrations reaching approximately double and triple pre-industrial levels, at 550 ppm (B1) and 970 ppm (A1fi), by 2100.
Monthly AOGCM temperature and precipitation fields were statistically downscaled to daily values for regions with a resolution of 1/8°, using an empirical statistical technique that maps the probability density functions for modeled monthly and daily precipitation and temperature for the climatological period (1961–1990) onto those of gridded historical observed data, so the mean and variability of both monthly and daily observations are reproduced by the climate model data. The bias correction and spatial disaggregation technique is one originally developed for adjusting AOGCM output for long-range streamflow projections, and compares favorably to different statistical and dynamic downscaling techniques (Wood et al. 2004).
Downscaled temperature and precipitation data were used as input to the Variable Infiltration Capacity model (Liang et al. 1994; Cherkauer et al. 2002), which simulates the full water and energy balance at the earth’s surface by modeling such processes as canopy interception, evapotranspiration (ET), runoff generation, infiltration, soil water drainage and snow pack accumulation and melt. The model is applied on a spatial grid with forcings (precipitation, temperature, radiation, etc.), soil properties (porosity, saturated hydraulic conductivity, etc.) and vegetation parameters (leaf area index, stomatal and architectural resistances, etc.) specified at each grid cell.
2.2 Insect severity index model
Climate projections from the HadCM3 model of monthly winter average temperatures at A1 (higher) and B1 (lower) IPCC emissions scenarios for early- (2010–2039), mid- (2040–2069) and late (2070–2099) 21st century were used as input to a flea beetle (Chaetocnema pulicaria) “severity index” model (Castor et al. 1975). This simple model is commonly used by Integrated Pest Management (IPM) programs to predict spring outbreak severity of this marginally over-wintering insect, which not only does direct crop damage, but is a vector for Stewart’s Wilt (Erwinia sterwartii), an economically important bacterial disease of sweet corn and field corn (corn grown for grain or silage). The severity index is calculated by adding together the average monthly air temperature for December + January + February. (For example, if the monthly average temperature was −1.5, −2.0, and −2.0°C for December, January, February, respectively, the severity index would be −5.5.) The index values are categorized as follows:
Range of D + J + F Avg. Temp | Category |
−3.3 to 2.2°C (i.e., warm winter) | “severe” beetle/disease pressure |
−6.2 to −3.3°C | “moderate” |
−8.8 to −6.2°C | “light” |
below −8.8°C (i.e., cold winter) | “trace” to no beetle/disease pressure. |
2.3 Thermal heat stress model for livestock
Heat stress for dairy cows (Bos taurus) and other livestock has been modeled utilizing a simple Thermal Heat Index (THI) equation (Klinedinst et al. 1993):
We used temperature and humidity output generated from the HadCM3 model at A1 (higher) and B1 (lower) IPCC emissions scenarios to project THI values at early- (2010–2039), mid- (2040–2069) and late (2070–2099) 21st century for the NE.
Threshold THI values for heat stress are approximately 70–72 for dairy cows, 72–75 for beef cows (B. taurus), 72–74 for pigs (Sus domestica), and 70–78 for chickens (Gallus domestica) (St. Pierre et al. 2003).
For dairy, a simple model predicting milk production decline (MPD) based on THI values has been developed:
where NL is “normal” milk production rate for the region in kilograms per cow per day, and MPD is also in the same absolute units.
For example, if NL is assumed to be 32 kg cow−1 day−1, then a THI of 72 would result in a very minor MPD of 0.38 kg cow−1 day−1; and THI of 80 would result in a MPD of 6.71 kg cow−1 day−1 (or 21% reduction).
3 Crop responses
3.1 Temperature
Recent warming (Wake 2005) and projected continued warming (Hayhoe et al. 2007) will have a significant impact on plants in natural and managed ecosystems. An advance in spring bloom date for several plant species in the NE has already been documented (Wolfe et al. 2005; Primack et al. 2004), and projections of spring bloom utilizing a lilac (Syringa spp.)/honeysuckle (Lonicera spp.) Spring Index model indicate the advance in spring bloom dates is likely to continue during the coming decades (Hayhoe et al. 2007).
Both positive and negative climate change effects on crop productivity may already be occurring in the region. For example, the rapid expansion and success of the European wine grape (V. vinifera) industry in upstate New York during the past 20 years may in part be attributed to less severe winters (reduced frequency of temperatures below −24°C) and reduced risk of vine and root damage (A. Lakso, Cornell University, personal communication). In contrast, an analysis of apple yields for Western New York (1971–1982) found that yields were lower in years when winters were warmer than average (based on accumulated degree days >5°C from January 1 to budbreak), possibly related to more variable fruit set following warmer winters (A. Lakso, personal communication).
An extended frost-free period as projected for the NE (Hayhoe et al. 2007; Frumhoff et al. 2006) will tend to benefit those attempting to produce crops requiring a relatively long growing season such as watermelon (Citrullus lanatus), tomatoes, peppers (Capsicum annuum), peaches (Prunus persica), and European red wine grape (V. vinifera) varieties. However, as discussed below, climate projections for the region also indicate an increase in summer heat stress, drought, and weed and pest pressure, which can have negative consequences for warm temperature-adapted crops as well as crops adapted to the historically cool climate of the region.
Many important grain crops, such as field corn, wheat, and oats tend to have lower yields when summer temperatures increase because the plant developmental cycle is speeded up and the duration of the grain-filling period is reduced (Rosenzweig and Hillel 1998; Mitchell et al. 1993). In addition, an increase in the frequency of day and/or night temperatures exceeding a high temperature threshold (varies with species, but typically between 27–35°C, Peet and Wolfe 2000) will negatively affect flowering, fruit set, and/or seed production of many crop species. Farmers in the NE may be able to adapt to these new climate constraints by switching to longer growing season or more heat tolerant varieties. But suitable new varieties may not always be available, or the market may not accept quality features of the new varieties (see Section 7 regarding farmer adaptation).
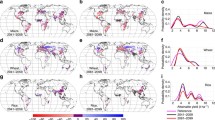
We compared projections of summer heat stress frequency (increase in number of days with maximum temperature exceeding 32°C) generated with IPCC future emissions scenarios A1fi (relatively higher) and B1 (lower). Figure 1 illustrates a subset of our results—the increase in number of heat stress days in the month of July at early-, mid-, and late-21st century based on the HadCM3 projections. At the higher emissions scenario, within just the next few decades (2010–2039), a 5–10 day increase in the number of July heat stress days is projected for the southern half of the region [i.e., much of Pennsylvania (PA), New Jersey (NJ), Delaware (DE), Connecticut (CT), and southern New York (NY)]. With a lower emissions scenario, the climate change impact does not become substantial until mid-century (2040–2069). By the end of century (2070–2099), with higher emissions, most days in July are projected to exceed the 32°C heat stress threshold for most of the NE. Even assuming relatively lower emissions, much of the NE is projected to have 10–15 more days of heat stress in July by end of century, except for some northern areas [e.g., northern Maine (ME) and Vermont (VT)], where the increase is in the range of 5–15 days.

Projected increase in number of crop heat stress days in July (where daytime temperatures exceed 32°C) for higher emissions (A1fi, upper panel) and lower emissions (B1, lower panel) scenarios compared to the 1961–1990 average
The projected increase in summer heat stress will be particularly detrimental to many cool temperature-adapted crops (e.g., cabbage, potato, apples) that currently dominate the NE agricultural economy. For example, the sensitivity of potato to climate change was illustrated by Rosenzweig et al. (1996), who used a physiologically-based crop simulation model and predicted −12, −22, and −49% yield reductions for Buffalo, NY with an average warming of +1.5, +2.5, and + 5.0°C, respectively, due primarily to negative warm temperature effects on tuberization in late summer and fall. Some crops will be particularly sensitive to night temperature as opposed to daytime maximum temperature, such as common snap bean, which shows substantial yield reductions when night temperatures exceed 27°C (Rainey and Griffiths 2005). Even crop species generally considered well-adapted to warm temperatures and a long growing season, such as tomato, can have reduced yield and/or fruit quality when daytime maximum temperatures exceed 32°C for short periods during critical reproductive stages (Sato et al. 2001). For many high value horticultural crops, very short-term (hours or a few days) of moderate heat stress at critical growth stages can reduce grower profits by negatively affecting visual or flavor quality even when total tonnage is not reduced (Peet and Wolfe 2000).
An increase in winter temperatures will also have a profound effect on the region’s plant life and agriculture. Mid-winter warming can lead to bud-burst or bloom of some perennial plants, resulting in frost damage when cold winter temperatures return. For crops requiring a prolonged “winter chilling” period to flower, yields will be negatively affected if the chilling requirement is not completely satisfied, even if spring and summer temperatures are optimum for growth. Many varieties of agricultural shrubs (e.g., blueberry, Vaccinum corymbosum), fruit trees (e.g., apples, grapes), and winter cereal grains (e.g., winter wheat, T. aestivum), have a winter chilling requirement of 200 to 2000 cumulative hours within a very narrow temperature range (typically 0–10°C with optimum chill-hour accumulation at 7.2°C, Westwood 1993). Temperatures below or above this range are usually ineffective in meeting the chill requirement, and in some cases warm temperatures (e.g., >15°C) can negate previously accumulated chill hours (Michaels and Amasino 2000).
We used the HadCM3 AOGCM and higher (A1fi) and lower (B1) emission scenarios to forecast the percentage of years within a 30-year period at early- (2010–2039), mid- (2040–2069) and late (2070–2099) when winter temperatures satisfied (stayed below) a chilling threshold of 7.2°C for 400, 1000, and 1800 cumulative hours. Currently, most of the NE satisfies even the highest chilling requirement (1800 h) in most years (based on 1961–1990 simulations, data not shown). Projection results for the medium chilling requirement (1000 h) are shown in Fig. 2. Only a slight diversion between emission scenarios is observed at mid-century. By late century, with higher emissions much of southern NE (all of PA, NJ, DE, CT, and most of MA and NY) has less than 50% of years meeting the 1000 h chilling requirement, while with lower emissions (B1) most of NY and MA still meet the chilling requirement in the majority of years. Projections for the high (1800 h) and low (400 h) chilling requirement (data not shown here, but available at http://www.northeastclimateimpacts.org) essentially bracket the results for the 1000 h requirement presented in Fig. 2. Compared to results for the 1000 h requirement, climate change impact occurs more quickly for the 1800 h requirement, where by mid-century significant diversion between emission scenarios is observed and much of southern NE has less than 50% of years meeting the requirement. The low chilling requirement threshold (400 h), in contrast, is projected to continue to be met for most of the region in most years through mid-century regardless of emissions scenario.

Projected percentage of years within 30-year periods during the 21st century when a 1000 h winter chill requirement (1000 h below 7.2°C vernalization threshold) will be met for the higher emissions (A1fi, upper panel) and lower emissions (B1, lower panel) scenarios
To summarize, our analyses (based on a simple <7.2°C threshold) indicate that a 400 h chilling requirement will continue to be met for most of the NE during this century regardless of emissions scenario. However, crops with prolonged cold requirements (1000 or more hours) could be negatively affected, particularly in southern sections of the NE and at the higher emissions scenario, where less than 50% of years satisfy the chill requirement by mid 21st century. The impact on crops will vary with species and variety. For example, native American grapes (V. labruscana) have a much longer chilling requirement than V. vinifera varieties (Westwood 1993). Chill requirements for apple range from 400–1800 h, with varieties Gala and Red Delicious at the low end of the scale, and MacIntosh and Empire at the high end of the scale. An important next step will be more detailed studies with crop-specific winter chilling models.
3.2 Rainfall and drought
Historical data for the NE reveal a trend for increased frequency of high-precipitation events (>5 cm in 48 h) (Wake 2005). This trend is expected to continue, with projections of a further increase in number of high precipitation events of 8% by mid-century and 12–13% by the end of the century (Frumhoff et al. 2006).
One economic consequence of excessive rainfall is delayed spring planting, which jeopardizes profits for farmers paid a premium for early season production of high value horticultural crops such as melon (Cucumis melo), sweet corn, and tomatoes (Lycopersicon esculentum). Field flooding during the growing season causes crop losses associated with anoxia, increases susceptibility to root diseases, increases soil compaction (due to use of heavy farm equipment on wet soils), and causes more runoff and leaching of nutrients and agricultural chemicals into ground- and surface-waters.
More rainfall concentrated into high precipitation events, combined with stable to modest reductions in summer and fall rainfall and increased temperatures leads to a projection for more short- (1–3 month) and medium-term (3–6 month) droughts for the region, particularly in the north and eastern parts of the NE (Hayhoe et al. 2007; Frumhoff et al. 2006). Drought frequency is projected to be much greater at the higher (A1fi) compared to lower (B1) emissions scenario. By the end of century (2070–2099) and with higher emissions, short-term droughts are projected to occur as frequently as once per year for much of the NE, and occasional long-term droughts (>6 month) are projected for western upstate NY—a region where many high value fruit and vegetable crops of considerable economic importance are grown.
Increased drought will be occurring at a time when crop water requirements are also increasing due to warmer temperatures. All crops are negatively affected by water deficits, but the relative impact on various agriculture sectors of the NE economy will depend on existing capacity for irrigation. Grain and silage crops do not bring sufficient profit to warrant investment in irrigation equipment and thus are typically “rainfed” in the NE. These crops, and the farmers growing them, would be particularly vulnerable to an increase in drought frequency. While many producers of high value horticultural crops in the NE have some irrigation equipment, most have not invested in enough equipment to optimize irrigation scheduling and fully meet ET requirements of all of their acreage in below “normal” years (Wilks and Wolfe 1998). Many horticultural crops are susceptible to quality defects (e.g., blossom end rot in tomato; tip burn in cabbage) caused by very short term (hours to days) fluctuations in water availability. Thus, the entire agricultural industry of the NE is vulnerable to drought, although farmers with sufficient capital may adapt by investment in expanded and more sophisticated irrigation systems (see Section 7 on farmer adaptation).
3.3 Direct CO2 fertilization effects
Carbon dioxide, in addition to being a greenhouse gas, can also directly affect Earth’s plant life because plants take up CO2 during photosynthesis to produce sugars for growth. Atmospheric CO 2 is projected to increase from current levels of approximately 380 ppm to 550 and 970 ppm by 2100 with the B1 and A1fi scenarios, respectively (Nakićenović et al. 2000). The plant growth stimulation from increasing CO2 generally follows an asymptotic curve that begins to saturate above about 600–800 ppm for most species (Wolfe 1994), so the potential “CO2 fertilization effect” will diminish over time under both emission scenarios.
Early studies, conducted primarily in growth chambers and greenhouses, found that plants with the so-called “C3” photosynthetic pathway, which includes most NE crop species (with the notable exception of corn) and many weed species, can show productivity increases of 20 to 30% or more when grown at twice current CO2 levels and at optimal conditions (Cure and Acock 1986). However, a recent review (Long et al. 2006) of more modern open-air field studies using the “Free-Air CO2 Enrichment” technology (no chamber effect), suggests that yield benefits for soybean, wheat and other C3 crops may be only about half of what was reported in the earlier enclosure studies.
Maximum CO2 beneficial effects on crop yields will likely require more fertilizer (to support bigger plants), optimum temperatures, unrestricted root growth, and excellent control of weeds, insects, and disease (Wolfe 1994). Weeds will be more aggressive and difficult to control as CO2 continues to rise (see Section 4.2, below).
High CO2 can have another direct effect on plants—reducing leaf conductance to transpirational water loss by causing partial closure of the stomates (the small openings on the leaf surface). However, this water conserving response to high CO2 at the leaf scale is modulated by processes at the whole-plant and/or ecosystem scales [e.g., high CO2 can cause an increase in total leaf (transpirational surface) area]. As a result, ET and soil water use are much less affected by high CO2 than is conductance at the leaf scale (Field et al. 1995), and an increase in ET at elevated compared to current ambient CO2 is sometimes observed (e.g., Hui et al. 2001).
Studies with bean (Jifon and Wolfe 2005), potato (see Peet and Wolfe 2000), and winter wheat (Mitchell et al. 1993) have shown that increasing CO2 cannot compensate for yield losses associated with negative heat stress effects on flower, fruit, or seed development. Thus, for heat-sensitive crops, much of the potential CO2 beneficial effect on crop growth will not be realized if CO2 increase is concomitant with an increase in frequency of high temperature stress as projected for the NE (Fig. 1).
4 Weed responses
4.1 Climate effects on ecology and distribution
Temperature and precipitation are key abiotic variables that control vegetative distribution (Woodward and Williams 1987), and as such will impact the geographical distribution of weeds with subsequent effects on their growth, reproduction and competitive abilities. Currently, estimated crop losses due to weeds without the use of herbicides are substantially larger in the south than in the north in both field corn (22 vs. 35%) and soybeans (22 vs. 64%) (Bridges 1992). This may be associated with the occurrence in the south of some very aggressive weeds such as kudzu (Pueraria lobata var. montana), whose presence is limited in the northern states by winter low temperatures (Bunce and Ziska 2000). Alternatively, greater increases in summer night-time relative to day-time temperatures projected with global warming (McCarthy et al. 2001) could decrease seed production to a greater extent in crop relative to weed species (e.g., cowpea, Vigna unguiculata, Ahmed et al. 1993) with subsequent competitive effects. Differential responses of seed emergence to temperature could also influence species establishment and subsequent weed/crop competition (e.g., Houghton and Thomas 1996).
Increased frequency of droughts as projected for the NE (Hayhoe et al. 2007; Frumhoff et al. 2006) may favor the crop by reducing the competitive impact of the weed (Patterson 1995). That is, when potential yield is already limited by water, weed competition for other resources has less impact. Water availability may also affect the duration of weed-free periods during crop development. Coble et al. (1981) demonstrated that in competition with common ragweed, a critical period to avoid competitive effects was 2 weeks in a dry year and 4 weeks in a wet year. However, the duration of the critical period varied by weed and crop (Harrison et al. 1985; Jackson et al. 1985).
Less is known regarding how alterations in temperature or moisture might affect the spread or success of invasive weeds. Detailed studies of invasive species such as itchgrass (Rottboelliia cochinchinensis) indicated that a warming of 3°C (day night temperature increase from 26/20 to 29/23°C) increased biomass and leaf area by 88 and 68%, respectively (Patterson et al. 1979). Empirically, based on its temperature response, itchgrass could effectively increase its percent of maximum growth from 50–75% in the Middle Atlantic States to 75–100% (Patterson et al. 1999). Northward expansion of other invasive weeds such as cogongrass (Imperata cylindrica) and witchweed (Striga asiatica) is also anticipated (Patterson 1995), although warming may restrict the southern expansion of some exotic weeds such as wild proso millet (Panicum miliaceum) due to increased competition (Patterson et al. 1986).
One potential threat of concern is northward expansion of kudzu, which currently infests over one million hectares in the southeastern U.S. Sasek and Strain (1990) used climate projections to illustrate how the latitudinal distribution of kudzu, limited by low winter temperatures (−15 to −20°C), could shift northward with climate change. Climate projections for the NE suggest a northward expansion of the suitable habitat for kudzu into the region during this century as the number of days with temperatures below −20°C decrease (Fig. 3). Emission scenarios begin to diverge at mid-century, with a larger fraction of the NE suitable for kudzu at the higher emissions scenario. While temperature is not the only factor that could constrain the spread of kudzu into the region, given the aggressive nature of this species, regenerative abilities, and recent migration pattern, continued northward expansion seems almost inevitable.

Projected number of days per year with minimum temperatures below −20°C at higher emissions (A1fi, upper panel) and lower emissions (B1, lower panel) scenarios. Spread of dark orange area indicates potential expansion of suitable habitat for Kudzu and other weed species constrained by cold winter temperatures
4.2 Weed responses to CO2
There are two lines of evidence suggesting that weeds will respond to a greater extent than crops to increasing atmospheric CO2, projected to reach 550 and 970 ppm by 2100 with the B1 and A1fi scenarios, respectively (Nakićenović et al. 2000). First, weed species have a greater genetic diversity than most crops. Consequently, if a resource (e.g., light, water, nutrients or CO2) changes within the environment, it is more likely that weeds will have a greater capacity for growth and reproductive response than crops (see Vengris et al. 1955 for nitrogen).
The worst weeds for a given crop are often similar in growth habit or photosynthetic pathway; indeed, they are often the same uncultivated or “wild” species, e.g. oat (Avena sativa) and wild oat (Avena fatua), sorghum (Sorghum vulgaris) and shattercane (Sorghum bicolor), rice (Oryza sativa) and red rice (also O. sativa). To date, for all weed/crop competition studies where the photosynthetic pathway is the same, weed growth is favored as CO2 is increased (Ziska and Runion 2006b). Secondly, in most agronomic situations there are more weed species than crop species. Although many weed species have the C4 photosynthetic pathway (and therefore may show a smaller response to atmospheric CO2 relative to C3 crops) no crop competes solely with all C3 or all C4 weeds. For fresh sweet corn in Pennsylvania for example, eight different weed species (both C3 and C4) are reported based on grower surveys (Bridges 1992). Therefore it seems likely that any resource increase is likely to stimulate the growth of one or more weed species in a crop–weed mix than the crop. Recent data for a soybean/weed system in fact, indicate C3 annuals (e.g., lambsquarters, Chenopodium album) are favored by natural selection with prolonged exposure to increased CO2, leading to increased competition with soybean (Ziska and Goins 2006a).
4.3 Implications for control and management
For the United States and many developed countries, chemical methodologies allow for cheap, effective weed control. Actually, a single herbicide, glyphosate (commercially sold as “Round-Up”), is so effective in controlling weeds that more than three-quarters of the U.S. soybean crop, and over a third of the U.S. corn crop have been genetically modified to be glyphosate resistant (e.g., Gaskell et al. 1999).
Unfortunately, there are also an increasing number of studies (Ziska et al. 1999, 2004; Ziska and Teasdale 2000; Ziska and Goins 2006a) that demonstrate a decline in chemical efficacy with rising CO2 per se. Given the evidence from field and laboratory trials, it appears that declining efficacy of herbicides with increasing CO2 is very likely to affect the environmental and economic cost of pesticide usage (Ziska et al. 1999; Ziska and Teasdale 2000).
What about other means of control? In the developing world, mechanical control is still the predominant means to prevent weed induced crop losses. Tillage, a common form of weed control, cuts and discs roots. But one response to rising CO2 may be an increase in below-ground root growth relative to above-ground shoot growth (Ziska 2003). For some invasive weeds, asexual propagation from small root segments is commonly observed. For Canada thistle (Cnicus benedictus), an invasive North American weed, rising CO2 can double root growth relative to shoot growth in the field (Ziska et al. 2004). As a consequence, increasing tillage as a control measure would lead to additional plant propagation for this species in a higher CO2 environment.
5 Insects and plant pathogens
5.1 Climate effects on ecology and distribution
Farms are complex ecosystems composed of a multitude of above- and below-ground animal, insect, and microbial species, in addition to plant species. Each of these species will be responding to the changing climate in unique ways. Beneficial soil microbial populations important for nutrient re-cycling, suppression of soil-born diseases, and other processes are likely to be highly sensitive to climate change and CO2-mediated effects on chemical composition of vegetation (e.g., carbon:nitrogen ratio), but we still no very little about potential impacts on the vast soil biota (Wolfe 2001). A major concern is how climate change might affect the synchrony among species that benefits crop plants, such as timing of spring bloom and arrival of insect pollinators, and presence and activity of “natural enemies” of crop insect or disease pests.
Studies conducted in Western Europe and other regions have already documented changes in spring arrival and/or geographic range of insect species due to climate change (Montaigne 2004; Goho 2004; Walther 2002). Temperature is the single most important factor affecting insect ecology, epidemiology, and distribution, while plant pathogens will be highly responsive to humidity and rainfall, as well as temperature. Wetter summers would tend to favor many foliar pathogens (Coakley et al. 1999). The increase in short- to medium-term drought projected for the NE (Hayhoe et al. 2007) will tend to decrease the duration of leaf wetness and reduce some forms of pathogen attack on leaves. An overall increase in humidity and frequency of heavy rainfall events projected for the NE (Frumhoff et al. 2006) will tend to favor some leaf and root pathogens (Coakley et al. 1999). However, the region is also expected to have more frequent drought periods (Hayhoe et al. 2007), and when those occur, pressure and spread of many potential pathogens could be reduced.
Warmer winters will likely increase the populations of marginally over-wintering insect species, such as flea beetles [C. pulicaria, a vector for bacterial Stewart’s Wilt (E. sterwartii) in corn] in the NE (Harrington et al. 2001). Figure 4 shows projected changes in the flea beetle/Stewart’s Wilt index (see Section 2) using the HadCM3 model output at A1fi (higher) and B1 (lower) emission scenarios.

Average index of flea beetle/Stewart’s Wilt severity (based on winter degree day accumulation model) at higher emissions (A1fi, upper panel) and lower emissions (B1, lower panel) scenarios. Expansion of yellow and orange region indicates expansion of region with frequent over-wintering and moderate to severe flea beetle pressure
Currently, only the southern-most regions of PA, the southern half of NJ, and MD have a moderate to severe flea beetle pressure in a typical year (based on 1961–1990 simulations, data not shown, and verified by personal communication, C. Petzoldt, Cornell University). Figure 4 illustrates that by mid-century for both scenarios there is a significant northward migration of habitable zone for flea beetle, and emission scenarios are beginning to diverge. By the end of century, “trace” levels are projected for only a relatively small acreage in northern ME at higher emissions, while at lower emissions, much of the northern half of ME and VT remain at this level. Overall, these results indicate the potential for a significant northern migration of the over-wintering range of flea beetle due to climate change during this century. It is reasonable to assume a similar expansion of over-wintering range would occur for other insect species, some of which may be crop pests, and others may be beneficial.
5.2 Indirect CO2 effects
The frequently observed higher carbon:nitrogen ratio of leaves of plants grown at high CO2 (Wolfe 1994) can require increased insect feeding to meet nitrogen (protein) requirements (Coviella and Trumble 1999). However, slowed insect development on high CO2-grown plants can lengthen the insect life stages vulnerable to attack by parasitoids (Coviella and Trumble 1999). Recently, Hamilton et al. (2005), in a field free-air CO2 enrichment study, found that early season soybeans grown at elevated CO2 atmosphere had 57% more damage from insects, presumably due in this case to measured increases in simple sugars in leaves of high CO2-grown plants. The “elevated” CO2 level in this study was about 550 ppm, approximately the level projected to occur by mid to late century with A1fi and B1 scenarios, respectively (Nakićenović et al. 2000).
5.3 Implications for control and management
For plant pathogens, the more frequent rainfall events projected by climate change models for the region (Frumhoff et al. 2006) could result in farmers finding it difficult to keep residues of contact fungicides on plants, triggering more frequent applications. Systemic fungicides could be affected negatively by plant morphological responses to CO2 and temperature that slow uptake rates, such as smaller stomatal opening or thicker epicuticular waxes on the leaves. Counteracting this effect, increased plant metabolic rates at warmer temperatures could increase fungicide uptake. It is not well understood how naturally-occurring biological control of pathogens by other microbial organisms could change as populations of microorganisms shift under changed temperature and moisture regimes—in some cases antagonistic organisms may out-compete pathogens while in others pathogens may be favored. Exclusion of pathogens and quarantines through regulatory means may become more difficult for authorities as unexpected pathogens might appear more frequently on imported crops.
For insect pests, a continued warming trend is likely to lead to increased pesticide use in the NE due to a number of factors, such as earlier arrival of migratory insects, more winter survival of insects that currently are only marginally adapted to the region, and more generations of insects within a single season. Additionally, some classes of pesticides (pyrethroids and spinosad) have been shown to be less effective in controlling insects at higher temperatures (Musser and Shelton 2005). There is currently a clear trend for increased pesticide use in warmer, more southern regions of the U.S. compared to the NE. For example, the frequency of pesticide sprays for control of lepidopteran insect pests in sweet corn currently ranges from 15–32 applications per year in Florida (Aerts et al. 1999), to 4–8 applications in Delaware (Whitney et al. 2000), and 0–5 applications per year in New York (Stivers 1999).
The IPM infrastructure for monitoring insect and disease populations already in place in much of the NE will be particularly valuable for helping farmers cope with climate change. The timing and placement of IPM monitoring equipment, as well as educational outreach efforts, will need to be consistently updated to keep pace with a rapidly changing climate. The preponderance of evidence indicates that there will be an overall increase in the number of outbreaks and northward migration of a wide variety of insects and pathogens. Minimizing negative environmental and economic impacts associated with increased use of fungicides and insecticides will require frequent review of IPM recommendations.
6 Livestock and dairy
6.1 Availability and price of feed
Climate change could indirectly affect livestock and dairy industries by affecting the availability and price of crops used for animal feed. Yields of grain corn and corn silage will be affected by multiple forces associated with climate change. Yields may increase and prices go down due to a longer growing season and increased CO2 (see Sections 3.1 and 3.3). On the other hand, yields may decline and prices increase due to crop losses associated with the increase in frequency of crop heat stress (Section 3.1, Fig. 1), drought (Section 3.2, Hayhoe et al. 2007), and heavy precipitation events (Section 3.2, Frumhoff et al. 2006) projected for the NE. Increased use of corn for ethanol production is already increasing feed prices in some areas.
6.2 Summer heat stress
Climate change will have more direct effects on these industries by affecting the intensity and frequency of animal summer heat stress. Heat stress in dairy cattle can have a long-term effect (weeks to months) on both milk production and birthing rates (Klinedinst et al. 1993). Dairy cows like it cool, with the temperature optimum for maximum milk production at temperatures between 4.5–23.8°C. Humidity also is an important factor. For example, heat stress can occur at temperatures as low as 23.8°C with a relative humidity >65%, or at temperatures of 26.7°C at relative humidity >30%.
A recent analysis (St. Pierre et al. 2003) estimated historical average annual economic losses associated with heat stress for dairy and other livestock industries at $50.8, $24.9, and $5.4 million per year for PA, NY, and VT, respectively. This was based on weather station records for the past 68 to 129 years and United States Department of Agriculture and industry data. Most of these losses were associated specifically with the dairy industry (e.g., >95% of the $24.9 million annual loss in New York was dairy). In the unusually warm summer of 2005, many New York dairy herds reported decreases in milk production of 5 to 15 lb per cow per day (8 to 20% decrease of normal production; L. Chase, personal communication).
Livestock THI calculations (see Section 2) for the month of July from 1961–1990 indicated that the entire NE was in the non-stress category (THI <72). For comparison, THI projections for the month of July at early-, mid-, and late-21st century are shown in Fig. 5. The THI increases from 1961–1990 levels by early 21st century, but there is only slight diversion between emission scenarios, and THI remains below 72 for almost the entire region. By mid-century (2040–2069), at the higher emissions scenario, NJ, DE, and southern PA have reached moderate stress levels (THI from 72–76), but this does not occur at the lower emissions until late 21st century (2070–2099). The diversion between emission scenarios is pronounced by end of century, where the higher emissions pathway leads to a THI in the 75–80 range (equivalent to a 10–25% reduction in milk production, see Section 2) for much of the NE except for parts of northern ME and VT.

Average THI for dairy cattle for July at higher emissions (A1fi, upper panel) and lower emissions (B1, lower panel) scenarios. A THI >72 (orange shaded regions) indicates sufficient heat stress to reduce milk production below potential at optimum temperature
6.3 Implications for livestock management
Heat stress can be minimized by several approaches. Lower cost measures include: reduce overcrowding, minimize time in hot holding areas, and maximize shade. Moderate cost measures include improved ventilation and fan systems improved insulation; installation for misters or sprinklers for cooling. Higher cost measures include new building design and construction and installation of thermostat controlled air conditioning systems. Some of the increased costs for cooling in summer could be compensated for by reduced heating requirements in winter.
Modification of feeding schedules and feed rations can also have some impact on heat stress. Feeding during cooler parts of the day, and increasing the proportion of easily digestible feeds is recommended. Increasing fats, avoiding excess protein, replacing some forage with non-fiber feeds (beet pulp, soy hulls), and adding buffers, yeast, and minerals lost during heat stress can help. Water consumption of dairy cattle can increase by 20–50% on hot, humid days, so providing plenty of water is essential.
7 Impact on rural economies and farmer adaptation
Most assessments of climate change impacts on agriculture are optimistic about farmer adaptation. These adaptations are typically not assigned a cost, yet it is important to recognize that even an adaptation with economic benefits in the long term will not be cost- or risk-free in the short term. Farmer vulnerability and risks associated with adaptation will be greater if climate variability increases with climate change (Smit and Skinner 2002). Mendelsohn (2000) concluded that most adaptation will be reactive and not anticipatory because future risks and benefits of adaptation are uncertain, while current costs of adaptation measures are relatively clear.
Available capital will undoubtedly influence decisions regarding climate change adaptation. Small family farms with little capital to invest in adaptation strategies will be more constrained and therefore more vulnerable in an uncertain climate than large corporate farms with more capital and more capacity for spreading risk geographically and temporally. Also, as Mendelsohn (2000) pointed out, a landowner will make decisions about adaptation based primarily on his/her own costs and benefits, which could lead to wrong decisions and inefficient adaptation from a societal perspective (e.g., when benefits are valued by others but not the landowner, or when true costs of adaptation are not paid by the landowner).
Specific implications of climate change for management of weeds, insects and pathogens, and livestock were discussed in detail in prior sections. Below, these and other potential farmer adaptation measures are placed into a broader framework and briefly discussed.
7.1 Change planting date
Among farmer adaptation options, changing planting and/or harvest date can be an effective, low-cost option to take advantage of a longer growing season or to avoid crop exposure to adverse climate (e.g., high temperature stress, low rainfall). Effectiveness will depend on region and the magnitude of climate change, as illustrated by Rosenzweig et al. (1996) in a potato crop model simulation study. They found that delaying planting date by 2 weeks so that tuber development would occur at more favorable cooler temperatures was effective at mitigating yield reductions associated with 1.5 and 2.5°C of warming in some regions (e.g., Caribou, ME) but not others (e.g., Boise, ID, Indianapolis, IN). Regardless of location, delayed planting date could not overcome yield reductions with a 5°C warming scenario.
It is important to keep in mind that changing planting date to mitigate yield reductions may not be an adaptive strategy if it means the farmer will be going to market when the supply/demand balance drives prices down. Predicting the optimum planting date for maximum profits will be very challenging in a future with increased uncertainty regarding climate effects on not only local productivity, but also on supply from competing regions.
7.2 Change crop varieties
Varieties with improved tolerance to heat or drought, or adapted to take advantage of a longer growing season for increased yield, will be available for some crop species. Changing varieties, like changing planting date, is a “first line of defense” for farmers to consider. There are a number of situations in which this might not be an effective strategy, however. An obvious case is perennial crops, where changing varieties is extremely expensive and new plantings take several years to reach maximum productivity. Even for annual crops, changing varieties is not always a low-cost option. Seed for new stress-tolerant varieties is sometimes expensive, new varieties often require investments in new planting equipment, or require adjustment in a wide range of farming practices. In some cases, it may not be possible to identify an alternative variety that is adapted to the new climate, and is also adapted to local soils and farming practices, and meets local market demand regarding timing of harvest and quality features such as fruit size and color.
7.3 Change crop or livestock species
This is a much more extreme, high-risk, and in most cases, high-cost option than simply changing varieties. It could bring new and even increased profits in the long term, but will require the capital to essentially enter into a new business. Accurate predictions of climate trends, and development of the infrastructure and market for the new crops or livestock products will be essential for this to be an effective response.
7.4 Increase water, fertilizer, herbicide, pesticide use
Warmer temperatures, longer growing seasons, and increased drought will lead to increase agricultural water use. Obtaining the maximum “CO2 fertilization” benefit often requires increases in water and fertilizer to support bigger plants (Wolfe 1994). As discussed previously, farmers are likely to respond to more aggressive and invasive weeds, insects, and pathogens with increased use of herbicides, insecticides, and fungicides. Although in some cases changes in the crop–weed–pest complex may favor the crop and lead to reduced usage. Where increases in water and chemical inputs become necessary, this will not only increase costs for the farmer, but will have society-wide impacts by depleting water supply, increasing reactive nitrogen and pesticide loads to the environment, and increasing risks to food safety and human exposure to pesticides.
7.5 New capital investments
Climate change could require significant capital investment for survival of the business, or to take advantage of new opportunities. Examples would include new irrigation or drainage systems, improved cooling facilities for livestock, or new or expanded crop storage facilities. The challenge will be strategic investment in relation to the timing and magnitude of climate change.
7.6 “Win–win opportunities
Climate change may be an incentive for farmers to take advantage of some “win–win” opportunities that benefit both the farmer and the environment. These include:
-
Conserve energy and reduce greenhouse gas emissions (increase profit margin and minimize contribution to climate change)
-
Increase soil organic matter (this not only improves soil health and productivity, but organic matter is mostly carbon derived from CO2 in the atmosphere (via plant photosynthesis), so it reduces the amount of this greenhouse gas in the atmosphere)
-
Improve nitrogen (N) use efficiency (synthetic N fertilizers are energy-intensive to produce, transport and apply; and soil emissions of nitrous oxide (a greenhouse gas) increase with N fertilizer use)
-
Enter the expanding market for renewable energy (e.g., wind energy, biomass fuels) using marginal land.
7.7 Beyond farm-level adaptation
Most adaptations discussed above directly involve farmers and farming practices. Smit and Skinner (2002), in their “typology” of agricultural adaptations, include other adaptation categories, such as technological development (e.g., seed company development of new varieties), farm policy, and government programs and insurance (e.g., low cost loans for adaptation). Some adaptations bridge across categories, such as timely release of climate projections, which can be viewed as a technological development, may be part of a government-subsidized education program, and would presumably be used by farmers and other land owners in the decision-making process.
8 Concluding remarks
Some sectors of the NE agricultural economy will expand while others decline or disappear as climate change alters growing season length, seasonal high and low temperatures, and other factors affecting crops and livestock. The dominant crop–weed–pest complexes will be different than what we have today. In general, weed, insect and disease pressures are likely to increase due to greater success of invasives in a changing climate, substantial beneficial effects of increasing CO2 on many weed species, and increased winter survival of insect and disease pests. A consequence of this will be increased herbicide and pesticide loads to the environment.
The sustainability of agriculture in the region will require strategic adaptations by farmers, government agencies, and others that will not be cost- or risk-free. Adaptations will have to occur sooner, and will be more complex and costly, if we follow a higher emissions pathway.
Under a lower emissions pathway, potential beneficiaries of climate change may include farmers currently producing or shifting to crops that may benefit from climate change and high CO2, farmers who guess correctly about climate and market trends, and farmers and corporations with sufficient capital for timely implementation of adaptation measures, or with multi-regional production options.
Those most vulnerable to climate change in the NE include small family farms with little capital and few resources to invest in adaptation strategies, as well as those who make poor decisions regarding the type and/or timing of adaptation measures to take. Adaptation to climate change will put additional stresses on the fragile and economically important dairy industry in the region.
References
Adams RM, Fleming RA, Chang CC, McCarl BA, Rosenzweig C (1995) A reassessment of the economic effects of global climate change on U.S. agriculture. Clim Change 30:147–167
Aerts M, Cockrell P, Nuessly G, Raid R, Schueneman T, Seal D (1999) Crop profile for corn (sweet) in Florida. Organization site: http://www.ipmcenters.org/CropProfiles/docs/FLcorn-sweet.html. Cited 22 January 1999
Ahmed FE, Hall AE, Madore MA (1993) Interactive effect of high temperature and elevated carbon dioxide concentration on cowpea [Vigna unguiculata (L.) Walp.]. Plant Cell Environ 16:835–842
Bridges DC (1992) Crop losses due to weeds in the United States. Weed Science Society of America, Champaign
Bunce JA, Ziska LH (2000) Crop ecosystem responses to climatic change: crop/weed interactions. In: Reddy KR, Hodges HF (eds) Climate change and global crop productivity. CABI, New York, pp 333–348
Castor LL, Ayers JE, MacNab AA, Krause RA (1975) Computerized forecasting system for Stewart’s bacterial disease in corn. Plant Dis Rep 59:533–536
Cherkauer KA, Bowling LC, Lettenmaier DP (2002) Variable infiltration capacity (VIC) cold land process model updates. Glob Planet Change 38:151–159
Coakley SM, Scherm H, Chakraborty S (1999) Climate change and plant disease management. Annu Rev Phytopath 37:399–426
Coble HD, Williams FM, Ritter RL (1981) Common ragweed (Ambrosia artemisiifolia) interference in soybeans (Glycine max). Weed Sci 29:339–342
Coviella C, Trumble J (1999) Effects of elevated atmospheric carbon dioxide on insect–plant interactions. Conserv Biol 13:700–712
Cure JD, Acock B (1986) Crop responses to carbon dioxide doubling: a literature survey. Agric For Meteorol 38:127–145
Field CB, Jackson RB, Mooney HA (1995) Stomatal responses to increased CO2: implications from the plant to global scale. Plant Cell Environ 18:12214–1225
Frumhoff, P, McCarthy J, Mellilo J, Moser S, Wuebbles D (2006) Climate change in the U.S. Northeast. Union of Concerned Scientists Publications, Cambridge (also available at http://www.climatechoices.org)
Gaskell G, Bauer M, Durant J, Allum N (1999) Worlds apart? The reception of genetically modified foods in Europe and the U.S. Science 285:384–387
Goho A (2004) Gardeners anticipate climate change. Am Gard 83(4):36–41
Hamilton JG, Dermody O, Aldea M, Zangerl AR, Rogers A, Berenbaum MR, DeLucia EH (2005) Anthropogenic changes in tropospheric composition increase susceptibility of soybean to insect herbivory. Environ Entomol 34(2):455–479
Harrington R, Fleming R, Woiwood IP (2001) Climate change impacts on insect management and conservation in temperate regions: can they be predicted? Agric For Entomol 3:233–240
Harrison SK, Williams CS, Wax LD (1985) Interference and control of giant foxtail (Setaria faberi) in soybeans (Glycine max). Weed Sci 33:203–208
Hayhoe K, Cayan C, Field CB, Frumhoff PC, Maurer EP, Miller NL, Moser SC, Schneider SH, Cahill KN, Cleland EE, Dale L, Drapek R, Hanemann RM, Kalkstein LS, Lenihan J, Lunch CK, Neilson RP, Sheridan SC, Verville JH (2004) Emissions pathways, climate change, and impacts on California. Proc Nat Acad Sci USA 101(34):12422–12427
Hayhoe K, Wake C, Huntington T, Luo L, Schwartz M, Sheffield J, Wood E, Anderson B, Bradbury J, DeGaetano A, Troy T, Wolfe D (2007) Past and future changes in climate and hydrological indicators in the U.S. Northeast. Clim Dyn 28:381–407
Houghton SK, Thomas TH (1996) Effects of elevated carbon-dioxide concentration and temperature on the growth and competition between sugar beet (Beta vulgaris) and fat-hen (Chenopodium album). Asp Appl Biol 45:197–204
Hui D, Luo Y, Cheng W, Coleman JS, Johnson D, Sims DA (2001) Canopy radiation- and water-use efficiencies as affected by elevated CO2. Glob Chang Biol 7:75–91
Jackson LA, Kapusta G, Mason DJS (1985) Effect of duration and type of natural weed infestations on soybean yield. Agron J 77:725–729
Jifon J, Wolfe DW (2005) High temperature-induced sink limitation alters growth and photosynthetic acclimation response to elevated CO2 in beans. J Am Soc Hortic Sci 130(4):515–520
Klinedinst PL, Wilhite DA, Hahn GL, Hubbard KG (1993) The potential effects of climate change on summer season dairy cattle milk production and reproduction. Clim Change 23:21–36
Liang X, Lettenmaier DP, Wood EF, Burges SJ (1994) A simple hydrologically based model of land surface water and energy fluxes for GSMs. J Geophys Res 99(D7):14415–14428
Long SP, Aisnsworth EA, Leakey A, Nosberger J, Ort DR (2006) Food for thought: lower than expected crop yield stimulation with rising CO2 concentrations. Science 312:1918–1921
McCarthy JJ et al (2001) Climate change 2001: impacts, adaptation, and vulnerability. Cambridge University Press, Cambridge
Mendelsohn R (2000) Efficient adaptation to climate change. Clim Change 45:583–600
Michaels SD, Amasino RM (2000) Memories of winter: vernalization and the competence to flower. Plant Cell Environ 23:1145–153
Mitchell RAC et al (1993) Effects of increase CO2 and temperature on growth and yield of winter wheat at two levels of nitrogen application. Plant Cell Environ 16:521–529
Montaigne F (2004) The heat is on: eco-signs. Natl Geogr 206(3):34–55
Musser FP, Shelton AM (2005) The influence of post-exposure temperature on the toxicity of insecticides to Ostrinia nubilalis (Lepidoptera:Crambidae). Pest Manag Sci 61:508–510
Nakićenović N et al (2000) IPCC special report on emissions scenarios. Cambridge University Press, Cambridge
Patterson DT (1995) Weeds in a changing climate. Weed Sci 43:685–701
Patterson DT et al (1986) Effects of temperature and photoperiod on Texas panicum (Panicumtexanum) and wild proso millet (Panicum miliaceum). Weed Sci 34:876–882
Patterson DT, Meyer CR, Flint EP, Quimby PC Jr (1979) Temperature responses and potential distribution of itchgrass (Rottboellia exaltata) in the United States. Weed Sci 27:77–82
Patterson DT, Westbrook JK, Joyce RJC, Lingren PD, Rogasik J (1999) Weeds, insects and diseases. Clim Change 43:711–727
Peet MM, Wolfe DW (2000) Crop ecosystem responses to climate change—vegetable crops. In: Reddy KR, Hodges HF (eds) Climate change and global crop productivity. CABI, New York
Pope VD, Gallani ML, Rowntree PR, Stratton RA (2000) The impact of new physical parameterizations in the Hadley Centre climate model-HadCM3. Clim Dyn 16:123–146
Primack D, Imbres C, Primack RB, Miller-Rushing A, del Tredici P (2004) Herbarium specimens demonstrate earlier flowering times in response to warming in Boston. Am J Bot 91(8):1260–1264
Rainey KM, Griffiths PD (2005) Differential response of common bean genotypes to high temperature. J Am Soc Hort Sci 130(1):18–23
Reilly JM (ed) (2002) Agriculture: the potential consequences of climate variability and change for the United States. Cambridge University Press, Cambridge
Rosenzweig C, Hillel D (1998) Climate change and the global harvest. Oxford University Press, Oxford
Rosenzweig C, Phillips J, Goldberg R, Carroll J, Hodges T (1996) Potential impacts of climate change on citrus and potato production in the US. Agric Syst 52:455–479
Sasek TW, Strain BR (1990) Implications of atmospheric CO2 enrichment and climatic change for the geographical distribution of two introduced vines in the USA. Clim Change 16:31–51
Sato S, Peet MM, Gardener RG (2001) Formation of parthenocarpic fruit and aborted flowers in tomato under moderately elevated temperatures. Sci Hortic 90:243–254
St. Pierre NR, Cobanov B, Schnitkey G (2003) Economic losses from heat stress by U.S. livestock industries. J Dairy Sci 86(E Suppl):E52–E77
Smit B, Skinner MW (2002) Adaptation options in agriculture to climate change: a typology. Mitig Adapt Strategies Glob Chang 7:85–114
Stivers L (1999) Crop profile for corn (sweet) in New York. http://pestdata.ncsu.edu/cropprofiles/docs/nycorn-sweet.html. Cited 20 January 1999
Vengris J, Colby WG, Drake M (1955) Plant nutrient competition between weeds and corn. Agron J 47:213–216
Wake CP (2005) Indicators of climate change in the Northeast. Clean air-cool planet and the climate change research center, University of New Hampshire, Durham (http://cleanair-coolplanet.org/information/pdf/indicators.pdf)
Walther G-R (2002) Ecological responses to recent climate change. Nature 416:389–395
Washington WM et al (2000) Parallel climate model (PCM) control and transient simulations. Clim Dyn 16:755–774
Westwood MN (1993) Temperate zone pomology, 3rd edn. Timber, Portland
Wilks DS, Wolfe DW (1998) Optimal use and economic value of weather forecasts for lettuce irrigation in a humid climate. Agric For Meteorol 89:115–129
Whitney SJ, Whalen M, VanGessel B, Mulrooney M (2000) Crop profile for corn (sweet) in Delaware. http://www.ipmcenters.org/CropProfiles/docs/DEcorn-sweet.html. Cited 15 February 2000
Wolfe DW (1994) Physiological and growth responses to atmospheric CO2 concentration. In: Pessarakli M (ed) Handbook of plant and crop physiology. Marcel Dekker, New York
Wolfe DW (2001) Tales from the underground: a natural history of subterranean life. Perseus, Cambridge
Wolfe DW, Schwartz MD, Lakso AN, Otsuki Y, Pool RM, Shaulis NJ (2005) Climate change and shifts in spring phenology of three horticultural woody perennials in northeastern USA. Int J Biometeorol 49:303–309
Wood A, Leung LR, Sridhar V, Lettenmaier D (2004) Hydrologic implications of dynamical and statistical approaches to downscaling climate model surface temperature and precipitation fields. Clim Change 62:189–216
Woodward FI, Williams BG (1987) Climate and plant distribution at global and local scales. Vegetatio 69:189–197
Ziska LH (2003) Evaluation of the growth response of six invasive species to past, present and future carbon dioxide concentrations. J Exp Bot 54:395–404
Ziska LH, Goins EW (2006a) Elevated atmospheric carbon dioxide and weed populations in glyphosate treated soybean. Crop Sci 46:1354–1359
Ziska LH, Runion GB (2006b) Future weed, pest and disease problems for plants. In: Newton P, Carran A, Edwards G, Niklaus P (eds) Agroecosystems in a changing climate. CRC, New York
Ziska LH, Teasdale JR (2000) Sustained growth and increased tolerance to glyphosate observed in a C3 perennial weed (Elytrigia repens (L.) Nevski), grown at elevated CO2. Aust J Plant Physiol 27:159–164
Ziska LH, Teasdale JR, Bunce JA (1999) Future atmospheric carbon dioxide may increase tolerance to glyphosate. Weed Sci 47:608–615
Ziska LH, Faulkner SS, Lydon J (2004) Changes in biomass and root:shoot ratio of field-grown Canada thistle (Cirsium arvense), a noxious, invasive weed, with elevated CO2: implications for control with glyphosate. Weed Sci 52:584–588
Acknowledgements
This work was funded in part by a Northeast Sustainable Agriculture Research and Education grant (ENE05-091) from the United States Department of Agriculture. The authors also thank the Union of Concerned Scientists for partial support for climate modeling and research team communications associated with this project.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This is an open access article distributed under the terms of the Creative Commons Attribution Noncommercial License ( https://creativecommons.org/licenses/by-nc/2.0 ), which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
About this article
Cite this article
Wolfe, D.W., Ziska, L., Petzoldt, C. et al. Projected change in climate thresholds in the Northeastern U.S.: implications for crops, pests, livestock, and farmers. Mitig Adapt Strateg Glob Change 13, 555–575 (2008). https://doi.org/10.1007/s11027-007-9125-2
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11027-007-9125-2