
Abstract
Aminopeptidase N (APN)/CD13 is a widely expressed transmembrane ectoenzyme and has been implicated in a myriad of physiological processes that are specific to cell type and tissue origin, including cancer cell metastasis, angiogenesis, cholesterol uptake, apoptosis, and cell migration. Skin cells, in particular fibroblasts have a relatively high level of APN/CD13 expression. The migratory capacity of skin cells is critical for the outcome of wound repair, as successful wound healing requires timely re-epithelialization which involves reformation of epithelium over wound surface by migrating keratinocytes. While failure of keratinocytes to undergo proper migration leads to chronic non-healing wounds, the presence of excess fibroblasts may contribute to formation of hypertrophic scars and keloids. The aim of this study was to investigate the role of APN/CD13 in skin cell migration and explore its potential as a therapeutic target in wound healing. Our results show an elevated expression of APN/CD13 in fibroblasts on the edge of the wound compared to unwounded cells. The presence of anti-APN/CD13 antibodies WM15, 3D8, and H300 reduces the migratory activity of human dermal fibroblasts in a dose-dependent manner by 42, 21, and 28%, respectively. However, the antibodies have no effect on keratinocyte migration. Further, none of the anti-APN/CD13 antibodies used in this study has any antiproliferative and cytotoxic effect on primary human keratinocytes or fibroblasts when used at 10 μg/ml in vitro. The differential inhibition on the migratory capacity of fibroblasts and keratinocytes presents an opportunity for anti-APN/CD13 antibodies to be used as a therapeutic agent for high fibroblast cellularity seen in fibroproliferative disorders.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Wound closure requires concerted efforts of fibroblasts and keratinocytes. Following the inflammatory phase, basal keratinocytes at the wound edge begin migrating as a sheet to cover the wound surface - a process referred to as “re-epithelialization” [1]. While keratinocytes migrate and divide to generate a protective layer for the dermis, resident dermal fibroblasts from the deep layers of the reticular dermis and the hypodermis, as well as fibroblasts differentiating from bone marrow-derived mesenchymal stem cells in the circulation enter the wound, and synthesize extracellular matrix components needed for structural repair [2, 3]. Clinically, defects in keratinocyte migration are associated with chronic non-healing wounds, while the presence of excess fibroblasts in the wound seems to contribute to hypertrophic scar formation [4, 5].
Aminopeptidase N (APN, also known as APN/CD13; EC.3.4.11.2) is a heavily glycosylated transmembrane peptidase [6] that also serves as a receptor for coronaviruses such as the transmissible gastroenteritis virus and human coronavirus 229E [7, 8]. It is ubiquitously expressed in endothelial cells, monocytes, macrophages, dendritic cells, neutrophils, fibroblasts, fibroblast-like cells such as synoviocytes, epithelial cells, and synaptic membranes and pericytes [9–14]. With dual roles as a peptide-cleaving enzyme and a cell surface receptor, APN/CD13 performs a myriad of functions in tumor cell invasion, proliferation/apoptosis, differentiation, antigen presentation, and viral infection [15]. However, due to the short intracellular domain lacking obvious signaling motifs [16], APN/CD13 was not regarded as a signal mediator until recently [17]. Since then, the functionality of APN/CD13 has expanded to adhesion, angiogenesis, and monocyte signaling [18–20]. For instance, Wulfaenger et al. associated the expression of enzymatically active wild-type APN/CD13 with diminished cell proliferation, migration and invasion in embryonic kidney cells [21]. Meanwhile, Pasqualini et al. demonstrated that APN/CD13 peptidase activity is necessary for endothelial invasion [17]. The broad functionality and wide-ranging expression pattern suggest that the function of this transmembrane peptidase may very well depend on the cell type and tissue origin.
A recent study showed that the APN/CD13 protein expression in fibroblasts is three times as high as that of keratinocytes [22]. Although the exact role of APN/CD13 in skin disease remains to be elucidated, its preferential expression in fibroblasts and its implication in tumor cell invasion make targeting APN/CD13 of potential value [23–26]. Considering the fact that cell migration is critical for the wound healing process, the present study investigated whether APN/CD13 plays a role in the cell migration of fibroblasts and keratinocytes, and examined the potential of APN/CD13-specific antibodies as a therapeutic agent for high cellularity seen in fibroproliferative disorders such as hypertrophic scarring.
Methods and materials
Cell culture
Skin punch biopsies were obtained with informed consent from patients undergoing elective surgery under local anesthesia. Written informed consent was obtained from each participant, and the study was approved by the University of British Columbia Hospital Human Ethics Committee and conducted according to the Declaration of Helsinki Principles. The detailed protocol of harvesting fibroblasts and keratinocytes from skin is described previously in [25]. Fibroblasts were grown in DMEM with 10% FBS and keratinocytes in KSFM (Invitrogen Life Technologies, Carlsbad, CA) supplemented with bovine pituitary extract (50 μg/ml) and epidermal growth factor (0.2 μg/ml).
In vitro wound scratch assay
For wound scratch assay, primary human fibroblasts or keratinocytes of passages 3–5 were seeded on 12-well plates in 10% FBS/DMEM or KSFM supplemented with bovine pituitary extract (50 μg/ml) and epidermal growth factor (0.2 μg/ml), respectively. At 95% confluency, the cells were washed with PBS twice and a scratch wound was made across each well with a 200-μl pipette tip. The cells were treated with anti-APN/CD13 antibodies, WM15, 3D8, and H300 (10 μg/ml each; Santa Cruz Biotechnology, Santa Cruz, CA) in serum-free DMEM and incubated at 37°C. Photographs were taken immediately, 12, and 24 h after treatment. Similarly, keratinocytes were treated with the respective antibodies in KSFM, and photographs were taken immediately and 48 h after treatment. Photographs were taken using a Zeiss HB-100 fluorescence microscope and analyzed using Northern Eclipse image analysis software (Carl Zeiss Canada, Toronto, ON).
Enzymatic assay
The quantitative assay for APN/CD13 on cultured human dermal fibroblasts was performed according to the previously published method [27]. In brief, the assay was performed in triplicate using Ala-4 nitroanilide (Ala-pNA, Sigma-Aldrich, Oakville, ON, Canada) in phosphate buffer pH 7.4 as chromogenic substrate. Cells were incubated with equal amounts of various CD13-specific antibodies 30 min prior to the assay. Ala-pNA (100 μl) was added in a concentration of 5 mM to 1 × 105 cells. Reactions were stopped by adding 900 μl acetate buffer (1.0 M, pH 4.4) after 60 min of incubation at 37°C. After centrifugation (2 min at 10,000×g), the absorbance of the supernatant was detected spectrophotometrically at 405 nm.
Cell proliferation assay
Cells were seeded on clear 96-well plates at a density of 1 × 104 cells/well. After 24 h, the culture medium was replaced by fresh medium (serum-free DMEM for fibroblasts and KSFM for keratinocytes). Antibodies 3D8, WM15, and H300 were added to the cells individually (10 μg/ml each), and the cells were incubated at 37°C for 2 days (fibroblasts) or 5 days (keratinocytes) in a 5% CO2 atmosphere. In untreated control cells, no antibody was added. Cell proliferation was evaluated using the MTT cell proliferation assay. Briefly, cells were rinsed twice with PBS and 100 μl of 4 mg/ml thiazolyl blue terzolium bromide (Sigma-Aldrich, Oakville, Ontario, Canada) was added to each well. After 5 h incubation, the blue crystal was solubilized by adding 200 μl of DMSO, and the absorbance was measured at 570 nm on a BioTek Microplate Spectrophotometer (BioTek Instruments, Inc., Winooski, VT).
Live/dead viability/cytotoxicity assay
The cell viability of dermal fibroblasts and keratinocytes after antibody treatment was examined using the live/dead viability/cytotoxicity kit for mammalian cells according to the manufacturer’s instructions (Invitrogen Life Technologies, Carlsbad, CA). Live cells appear green as a result of the intracellular hydrolysis that converts the non-fluorescent calcein AM (acetoxymethyl ester) to green fluorescent calcein. Dead cells yield red fluorescence as a result of the permeation of ethidium homodimer-1 into the compromised membranes of dead cells.
Immunohistochemistry
Following a scratch assay described above, cultured fibroblasts were fixed with 4% paraformaldehyde (PFA) for 30 min on ice, and blocked with TBS containing 5% goat serum and 1% goat albumin (Sigma-Aldrich, Oakville, Ontario, Canada). APN and vinculin were detected using 1:250 rabbit anti-APN (Santa Cruz Biotechnology, Santa Cruz, CA) and 1:100 mouse anti-vinculin antibodies (Sigma-Aldrich, Oakville, Ontario, Canada) followed by 1:4,000 Rhodamine-conjugated goat anti-rabbit (Millipore, Billerica, MA) and 1:4,000 Alexa 488-conjugated goat anti-mouse IgG (Invitrogen, Eugene, OR), respectively. Samples were mounted in ProLong Gold antiface reagent with DAPI (Invitrogen, Eugene, OR) and examined by a Zeiss Axioplan-2 fluorescent microscope and Northern Eclipse image analysis software (Carl Zeiss Canada, Toronto, ON).
Statistical analysis
Data were expressed as mean ± SD and the t -test was used to compare the mean values between different treatments. P values <0.02 were considered statistically significant in this study.
Results
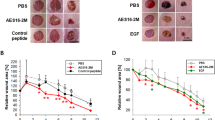
To examine the role of APN/CD13 in fibroblast migration, primary human fibroblasts were treated with anti-APN/CD13 antibodies with differential enzyme-inhibiting activities [28] in an in vitro wound healing assay. Images of cells were taken at 0, 12, and 24 h post-treatment (Fig. 1a). The total number of cells in the untreated (control) and antibody-treated wells that have migrated into the open wound area by the end of the 12- and 24-h incubation was recorded. Cell migration was evaluated as the percentage of cells present in the open wound area of the treated wells relative to the control well at the end of the 24-h incubation. There was no significant difference between treatment and control at 12 h post-scratch. However, at the end of the 24-h incubation when the control wound has closed, the WM15-treated cells showed a 42% reduction in migratory activity relative to the control, whereas the 3D8 and H300 antibodies slowed down cell migration by 21 and 28%, respectively (Fig. 1b). To examine whether the differential effect of the antibodies on cell migration inhibition is related to their ability to inhibit the enzymatic activity of endogenous APN/CD13, human dermal fibroblasts treated with the same concentrations of antibodies used in the scratch assay, and the enzymatic activity was measured using a chromogenic substrate, Ala-4 nitroanilide. As shown in Fig. 1c, 3D8, WM15, and H300 reduced the endogenous APN/CD13 enzymatic activity by 36, 76, and 9%, respectively.

Evaluation of the effect of anti-APN/CD13 antibodies on fibroblast migration. a Images of human fibroblasts taken immediately, 12 and 24 h after addition of anti-APN/CD13 antibodies 3D8, WM15 and H300 (10 μg/ml) in an in vitro wound scratch assay. Scale bar: 10 μm, b Reduction of fibroblast migration in the presence of the anti-APN/CD13 antibodies. Fibroblast migration was evaluated by counting the number of cells migrated from the edges of the wound into the denuded area after the 12- and 24-h treatment, and expressed as the percentage of cells present in the open wound area relative to that of the control well at the end of the 24-h incubation (n = 6, * P < 0.01), c Enzymatic assay of endogenous APN/CD13 in human fibroblasts after treatment with the various APN/CD13-specific antibodies (10 μg/ml; n = 6, * P < 0.02)
The inhibitory effect of WM15 was further examined with different doses of the antibody. As shown in Fig. 2, WM15 inhibits fibroblast migration into the wound in a dose-dependent manner. Specifically, the percent inhibition of fibroblast migration as a result of adding 1.0, 5.0, and 10.0 μg/ml of WM15 was 18, 34, and 48%, respectively.

Dose-dependent inhibitory effect of the WM15 antibody on fibroblast migration. a Photos of human fibroblasts were taken immediately, 12 and 24 h after addition of different doses of the WM15 antibody. The in vitro wound scratch assay was performed as described in Methods and materials. Scale bar: 10 μm, b Dose-dependent inhibitory effect of WM15 on fibroblast migration. Fibroblast migration was evaluated by counting the number of cells migrated from the edges of the wound into the denuded area after the 12- and 24-h treatment, and expressed as the percentage of cells present in the open wound area relative to that of the control well at the end of the 24-h incubation (n = 5, * P < 0.01)
Although effective inhibition of fibroblast migration is a critical aspect in preventing hypertrophic scar formation, a similar effect in keratinocytes may lead to failure of wound closure. For this reason, the influence of anti-APN/CD13 antibodies on keratinocyte migration was also examined. A monolayer of confluent human keratinocytes was similarly scratched as described above, and cell migratory activity was observed up to 48 h at which time the control scratch closed. As shown in Fig. 3a, keratinocyte migration appeared to be little affected by the antibodies even after 48 h. Few keratinocytes have moved by the 24-h time point (data not shown) as there is generally a time lag in keratinocyte migration after wounding, which correlates with the reported optimal time (24–48 h) for keratinocyte movement [29]. Figure 3b shows that the percent inhibition of keratinocyte migration by 3D8, WM15, and H300 was 4, 1, and 11%, respectively, which are not significantly different from that of the control.

Evaluation of the effect of anti-APN/CD13 antibodies on keratinocyte migration. a Images of human keratinocytes were taken immediately and 48 h after addition of the 3D8, WM15 and H300 antibodies (10 μg/ml each) in an in vitro wound scratch assay. Scale bar: 10 μm, b Keratinocyte migration was not affected by antibody addition. Keratinocyte migration was evaluated by counting the number of cells migrated from the edges of the wound into the denuded area after the 48-h treatment, and expressed as the percentage of cells present in the open wound area relative to that of the control well at the end of the 48-h incubation (n = 5, * P < 0.01)
To ensure that the inhibitory effect on fibroblast migration was not due to cell growth retardation, an MTT assay was performed to study whether the antibodies have any effect on cell proliferation. After exposing confluent cell cultures of fibroblasts to individual antibodies for a period of 48 h, cell proliferation was measured in terms of the fluorescence intensity in each sample. The result showed that the antibodies had no anti-proliferative effect on fibroblasts, as the proliferation relative to control cells was 99, 99, and 98% in the presence of 3D8, WM15, and H300, respectively (Fig. 4a, left).

Evaluation of the effect of anti-APN/CD13 antibodies 3D8, WM15 and H300 on the cell proliferation and viability of fibroblasts and keratinocytes after 2 and 5 days of antibody treatment (10 μg/ml), respectively. a Determination of cell proliferation by MTT assay, b Assessment of cell viability by live/dead viability/cytotoxicity assay. Fluorescent images of fibroblasts (upper panels) and keratinocytes (lower panels) were taken at the end of the indicated incubation periods. Scale bar: 10 μm Higher magnification images are shown in the square boxes (×40). The viable and non-viable cells are shown in green and red, respectively, with the nuclei of the cells stained with DAPI (blue), c Cell viability is represented by the percentage of viable cells in the total cell population
As any negative effect on cell proliferation may impair the outcome of wound healing, an MTT assay was similarly performed on keratinocytes to examine the effect on keratinocyte proliferation. In consideration of the fact that keratinocytes require more time to cover the wound surface in the in vitro scratch assay, we have extended the period of antibody exposure to 5 days in keratinocyte MTT assay. The cell proliferation was 95, 95, 94% in the presence of 3D8, WM15, and H300, respectively, indicating a subtle effect on keratinocyte proliferation (Fig. 4a, right). However, this reduction was not significantly different from that of the control.
To ensure the safety of these antibodies as a therapeutic agent, a cytotoxicity/cell viability assay was performed. The assay utilizes two reagents, calcein AM and ethidium homdimer-1, for labeling live and dead cells, respectively. Calcein AM is an esterase substrate hydrolyzed intracellularly to calcein, showing live cells in green. Ethidium homdimer-1 is a red fluorescent nucleic acid stain capable of permeating only the compromised membranes of dead cells which are shown in red. The photos presented in Fig. 4b are superimposed images of the live and dead cells in fibroblast (upper panels) and keratinocyte cultures (lower panels). Fibroblasts were incubated with the antibodies for 2 days prior to cytotoxicity assay, while keratinocytes were exposed to the antibodies for a longer period (5 days). Live and dead cells were counted visually, and the percentage of live cells was calculated and converted into percent cell viability by taking the percentage of live cells in the control sample as 100%. The antibodies produced a negligible cytotoxic effect on fibroblasts, with less than 1% reduction in cell number for all three antibodies (Fig. 4c, left). Although the difference was not significant, the 3D8 antibody seemed to have a more pronounced cytotoxic effect on keratinocytes, causing 10% reduction in cell viability compared to the control, while the effect was only slightly stronger with WM15 and H300, reducing the cell number by 2 and 3%, respectively (Fig. 4c, right).
The APN/CD13 protein expression during fibroblast cell migration was examined using independent in vitro models. As shown in Fig. 5a, fibroblasts were seeded at different cell densities ranging from 5.0 × 104 to 4.0 × 105 cells per cm2, and cultured under normal conditions for 24 h prior to lysis. A density-dependent protein expression was observed following immunoblotting analysis. The APN/CD13 expression peaked at 1.0 × 105 cell/cm2 where cells are most likely in the proliferative and migratory phase of growth. This expression quickly diminished as fibroblasts reached confluency at higher cell densities. In addition, we analyzed the APN/CD13 expression in a fibroblast migration assay (scratch assay), where a confluent monolayer at its maximal density was “wounded” by removing several wide bands of cells in both X and Y dimensions. Migration of fibroblasts into the open wound area occurred within 24 h before the cells were lysed for SDS-PAGE and Western blot analysis. When compared to uninjured cells, cell populations of migratory fibroblasts displayed a 20% higher expression of the APN/CD13 protein (Fig. 5b). Further, expression of the APN/CD13 protein on individual migratory cells was examined. A scratch wound was made across a confluent monolayer of dermal fibroblasts, and vinculin expression was used as an indicator of cells in motion. As shown in the upper panels of Fig. 5c, APN/CD13 expression was highest in the actively migrating cells within the denuded wound area, and lowest in the stationary cells distant from the wound scratch. As shown in the merged image, staining the same cell population with an anti-vinculin antibody confirmed significant overlap in the expression of APN/CD13 and vinculin. Higher magnification images are shown in the lower panels of Fig. 5c.

Fibroblast cell density and APN/CD13 expression. a Cells seeded overnight at increasing density (50, 100, 200, 400 × 103 cells) were analyzed for APN/CD13 protein expression by Western blotting, b An equal number of scratch wounds were made in X and Y directions on a confluent monolayer of dermal fibroblasts. “Unwounded” confluent cells were used as control. Cell lysate was obtained following overnight incubation and analyzed for APN/CD13 protein expression by Western blotting. β-actin expression was used as a loading control in both panels, c A scratch wound was made across a confluent monolayer of primary human fibroblasts, and the cells were stained with an anti-vinculin antibody and an anti-CD13 antibody, followed by Alexa 488 and Rhodamine secondary antibodies, respectively. The dotted line in the upper panels (×10; Scale bar: 10 μm) shows the border of the scratch. Higher magnification (×40; Scale bar: 80 μm) images are shown in the lower panels
Discussion
The concept of inhibiting cell migration by reducing APN/CD13 expression or activity through use of siRNA, chemical inhibitors or neutralizing antibodies has proven to be successful in tumor cells such as ovarian carcinoma cells [30], and in lymphocytes and endothelial cells [31]. For instance, Mina-Osorio et al. utilized anti-APN/CD13 antibodies to induce adhesion of monocytes and endothelial cells [20]. In this study, we took advantage of the preferential expression of APN/CD13 in fibroblasts, and proved for the first time that the APN/CD13-neutralizing antibody WM15 can be used to effectively reduce the migration of fibroblasts into an open scratch wound without slowing down keratinocyte movement. Similarly to the previous finding using human lung cancer cells [24], the inhibitory effect of the antibodies on cell migration is only partly due to the suppression of APN/CD13 enzymatic activity as shown by the enzymatic assay results. The most potent cell migration-inhibiting WM15 antibody is also the most effective enzymatic inhibitor. The other two antibodies 3D8 and H300 have similar effects on cell migration inhibition (21 vs. 28%) but suppress the enzymatic activity differently (35 vs. 15%), indicating that the observed cell migration-inhibitory effect is not entirely dependent on the enzymatic activity of APN/CD13. One possible reason is the differential binding avidity of 3D8 and H300 (monoclonal vs. polyclonal). Importantly, fibroblasts and keratinocytes were both able to maintain a healthy state even after days of exposure to WM15, preparing the antibody in a better position for in vivo clinical use which may require longer exposure to be effective. With the use of activity inhibitors, actinonin, bestatin, and PAQ-22, Thielitz et al. reported a direct correlation between enzyme activity and cell proliferation in HaCaT cells, but did not observe any antiproliferative effect by WM15 [32]. This reconfirmed our finding that neither fibroblast nor keratinocyte proliferation was affected by the presence of WM15. Thus, WM15 which has an effective dosage of as low as 1.0 μg/ml in vitro is unlikely to reduce the viability of fibroblasts and keratinocytes or impair normal wound healing.
While keratinocyte migration is known to involve formation of lamellipodia, loss of contact with hemidesmosomes and desmosomes, changes in keratin phenotype and redistribution of the actin cytoskeleton into lamellipodia [33, 34], the molecular mechanism and cellular events undertaken by migrating fibroblasts are poorly understood. Nevertheless, fibroblasts and keratinocytes are most likely to adopt distinctive regulatory mechanisms in cell migration due to differences in their external environments and surrounding cells. Thus, in order to understand why the APN/CD13-neutralizing antibody WM15 affects the migratory capacity of fibroblasts but not that of keratinocytes, more work on the mechanistic details involved in fibroblast migration will be needed. Meanwhile, a plausible rationale suggests that the differential effect might be due to enhanced antibody responsiveness in fibroblasts which have higher levels of APN/CD13 expression and activity than keratinocytes.
The density-dependent expression pattern of APN/CD13 indicated the highest protein expression at 1.0 × 105 cell/cm2, which allows fibroblasts to achieve an optimal cell–cell contact for migration, thus requiring higher expression of APN/CD13. Kehlen and colleagues [35, 36] have shown that direct cell–cell contact of lymphocytes with fibroblast-like synoviocytes resulted in an increase in APN/CD13 mRNA and cell surface protein in the lymphocytes. In that regard, fibroblasts seeded at a lower cell density (5.0 × 104 cell/cm2) expressed less APN/CD13 possibly due to lack of cell–cell contact, while the limited space at higher cell densities (5.0 × 105 and 1.0 × 106 cell/cm2) prevented cell movement by contact inhibition. Further, immunohistochemical staining of cells migrating into an artificially created scratch wound showed that compared to stationary cells, actively migrating cells as indicated by vinculin expression [37, 38] were expressing significantly more APN/CD13. This observation further supports our findings on the significant role of APN/CD13 in fibroblast motility.
In general, pathological skin conditions such as hypertrophic scars or keloids have high cellularity and vessel density [39], and remain in that condition well into the remodeling phase. In normal wound healing, timely apoptosis significantly reduced tissue cellularity and blood supply in early remodeling phase. We believe anti-CD13 antibodies can play a key therapeutic role in reducing wound cellularity by inhibiting fibroblast migration into the wound as well as in reversing high vascularity by blocking migration of endothelial cells [40]. In conclusion, APN/CD13-specific antibodies may be used as a therapeutic agent for normalizing fibroblast cellularity and vessel density which are elevated in many fibroproliferative disorders.
References
Hell E, Lawrence JC (1979) The initiation of epidermal wound healing in cuts and burns. Br J Exp Pathol 60:171–179
Fathke C, Wilson L, Hutter J, Kapoor V, Smith A, Hocking A et al (2004) Contribution of bone marrow-derived cells to skin: collagen deposition and wound repair. Stem cells 22:812–822. doi:10.1634/stemcells.22-5-812
Ham AW (1979) The integumentary system. In: Cormack DH (ed) Histology, 8th edn. JB Lippincott, Philadelphia and Toronto, pp 614–644
Raja, Sivamani K, Garcia MS, Isseroff RR (2007) Wound re-epithelialization: modulating keratinocyte migration in wound healing. Front Biosci 12:2849–2868. doi:10.2741/2277
Nakaoka H, Miyauchi S, Miki Y (1995) Proliferating activity of dermal fibroblasts in keloids and hypertrophic scars. Acta Dermatol Venereol 75:102–104
Danielsen EM, Cowell GM, Norén O, Sjöström H, Dorling PR (1983) Biosynthesis of intestinal microvillar proteins. The effect of swainsonine on post-translational processing of aminopeptidase N. Biochem J 216(2):325–331
Delmas B, Gelfi J, L’Haridon R, Vogel LK, Sjöström H, Norén O, Laude H (1992) Aminopeptidase N is a major receptor for the entero-pathogenic coronavirus TGEV. Nature 357(6377):417–420. doi:10.1038/357417a0
Yeager CL, Ashmun RA, Williams RK, Cardellichio CB, Shapiro LH, Look AT et al (1992) Human aminopeptidase N is a receptor for human coronavirus 229E. Nature 357(6377):420–422. doi:10.1038/357420a0
Turek JJ, Robinson JP (1994) Leucine aminopeptidase activity by flow cytometry. Methods Cell Biol 41:461–467. doi:10.1016/S0091-679X(08)61735-5
Shah T, Webster AD, Peters TJ (1983) Enzyme analysis and subcellular fractionation of human peripheral blood lymphocytes with special reference to the localization of putative plasma membrane enzymes. Cell Biochem Funct 1(2):117–124. doi:10.1002/cbf.290010214
Favaloro EJ (1991) CD13 (‘gp150’; aminopeptidase-N): co-expression on endothelial and haemopoietic cells with conservation of functional activity. Immunol Cell Biol 69(Pt 4):253–260. doi:10.1038/icb.1991.36
Harris CA, Hunte B, Krauss MR, Taylor A, Epstein LB (1992) Induction of leucine aminopeptidase by interferon-gamma. Identification by protein microsequencing after purification by preparative two-dimensional gel electrophoresis. J Biol Chem 267(10):6865–6869
Riemann D, Hansen GH, Niels-Christiansen L, Thorsen E, Immerdal L, Santos AN et al (2001) Caveolae/lipid rafts in fibroblast-like synoviocytes: ectopeptidase-rich membrane microdomains. Biochem J 354(Pt 1):47–55. doi:10.1042/0264-6021:3540047
Olsen J, Kokholm K, Norén O, Sjöström H (1997) Structure and expression of aminopeptidase N. Adv Exp Med Biol 421:47–57
Mina-Osorio P (2008) The moonlighting enzyme CD13: old and new functions to target. Trends Mol Med 14(8):361–371. doi:10.1016/j.molmed.2008.06.003
Olsen J, Cowell GM, Kønigshøfer E, Danielsen EM, Møller J, Laustsen L et al (1988) Complete amino acid sequence of human intestinal aminopeptidase N as deduced from cloned cDNA. FEBS Lett 238(2):307–314. doi:10.1016/0014-5793(88)80502-7
Pasqualini R, Koivunen E, Kain R, Lahdenranta J, Sakamoto M, Stryhn A et al (2000) Aminopeptidase N is a receptor for tumor-homing peptides and a target for inhibiting angiogenesis. Cancer Res 60(3):722–727
Mina-Osorio P, Ortega E (2004) Signal regulators in FcR-mediated activation of leukocytes? Trends Immunol 25(10):529–535. doi:10.1016/j.it.2004.08.008
Mina-Osorio P, Ortega E (2005) Aminopeptidase N (CD13) functionally interacts with FcgammaRs in human monocytes. J Leukoc Biol 77(6):1008–1017. doi:10.1189/jlb.1204714
Mina-Osorio P, Winnicka B, O’Conor C, Grant CL, Vogel LK, Rodriguez-Pinto D et al (2008) CD13 is a novel mediator of monocytic/endothelial cell adhesion. J Leukoc Biol 84(2):448–459. doi:10.1189/jlb.1107802
Wulfaenger J, Niedling S, Riemann D, Seliger B (2008) Aminopeptidase N (APN)/CD13-dependent CXCR4 downregulation is associated with diminished cell migration, proliferation and invasion. Mol Membr Biol 25(1):72–82. doi:10.1080/09687680701551855
Gabrilovac J, Cupić B, Breljak D, Zekusić M, Boranić M (2004) Expression of CD13/aminopeptidase N and CD10/neutral endopeptidase on cultured human keratinocytes. Immunol Lett 91(1):39–47. doi:10.1016/j.imlet.2003.10.006
Fukasawa K, Fujii H, Saitoh Y, Koizumi K, Aozuka Y, Sekine K et al (2006) AminopeptidaseN(APN/CD13) is selectively expressed in vascular endothelial cells and plays multiple roles in angiogenesis. Cancer Lett 243(1):135–143. doi:10.1016/j.canlet.2005.11.051
Chang YW, Chen SC, Cheng EC, Ko YP, Lin YC, Kao YR et al (2005) CD13 (aminopeptidase N) can associate with tumor-associated antigen L6 and enhance the motility of human lung cancer cells. Int J Cancer 116(2):243–252. doi:10.1002/ijc.21089
Hashida H, Takabayashi A, Kanai M, Adachi M, Kondo K, Kohno N et al (2002) Aminopeptidase N is involved in cell motility and angiogenesis: its clinical significance in human colon cancer. Gastroenterology 122(2):376–386. doi:10.1053/gast.2002.31095
Saitoh Y, Koizumi K, Minami T, Sekine K, Sakurai H, Saiki I (2006) A derivative of aminopeptidase inhibitor (BE15) has a dual inhibitory effect of invasion and motility on tumor and endothelial cells. Biol Pharm Bull 29(4):709–712. doi:10.1248/bpb.29.709
Raynaud F, Bauvois B, Gerbaud P, Evain-Brion D (1992) Characterization of specific proteases associated with the surface of human skin fibroblasts, and their modulation in pathology. J Cell Physiol 151(2):378–385. doi:10.1002/jcp.1041510219
Giugni TD, Soderberg C, Ham DJ, Bautista RM, Hedlund KO, Moller E, Zaia JA (1996) Neutralization of human cytomegalovirus by human CD13-specific antibodies. J Infect Dis 173:1062–1071
O’Toole EA (2001) Extracellular matrix and keratinocyte migration. Clin Exp Dermatol 26:525–530. doi:10.1111/j.1365-2230.2001.00891.x
Terauchi M, Kajiyama H, Shibata K, Ino K, Nawa A, Mizutani S et al (2007) Inhibition of APN/CD13 leads to suppressed progressive potential in ovarian carcinoma cells. BMC Cancer 7:140. doi:10.1186/1471-2407-7-140
Proost P, Mortier A, Loos T, Vandercappellen J, Gouwy M, Ronsse I et al (2007) Proteolytic processing of CXCL11 by CD13/aminopeptidase N impairs CXCR3 and CXCR7 binding and signaling and reduces lymphocyte and endothelial cell migration. Blood 110(1):37–44. doi:10.1182/blood-2006-10-049072
Thielitz A, Bukowska A, Wolke C, Vetter R, Lendeckel U, Wrenger S et al (2004) Identification of extra- and intracellular alanyl aminopeptidases as new targets to modulate keratinocyte growth and differentiation. Biochem Biophys Res Commun 321(4):795–801. doi:10.1016/j.bbrc.2004.07.029
Bereiter-Hahn J, Strohmeier R, Kunzenbacher I, Beck K, Vöth M (1981) Locomotion of Xenopus epidermis cells in primary culture. J Cell Sci 52:289–311
Stenn KS (1986) Keratinocyte adherence. J Invest Dermatol 86(2):214. doi:10.1111/1523-1747.ep12284305
Riemann D, Kehlen A, Thiele K, Löhn M, Langner J (1997) Induction of aminopeptidase N/CD13 on human lymphocytes after adhesion to fibroblast-like synoviocytes, endothelial cells, epithelial cells, and monocytes/macrophages. J Immunol 158(7):3425–3432
Kehlen A, Olsen J, Langner J, Riemann D (2000) Increased lymphocytic aminopeptidase N/CD13 promoter activity after cell-cell contact. J Cell Biochem 80(1):115–123. doi:10.1002/1097-4644(20010101)80:1<115:AID-JCB100>3.0.CO;2-R
Zieske JD, Bukusoglu G, Gipson IK (1989) Enhancement of vinculin synthesis by migrating stratified squamous epithelium. J Cell Biol 109(2):571–576. doi:10.1083/jcb.109.2.571
Beurden HE, Snoek P, Hoff J, Torensma R, Maltha J, Kuijpers-Jagtman A (2006) In vitro migration and adhesion of fibroblasts from different phases of palatal wound healing. Wound Repair Regen 14(1):66–71. doi:10.1111/j.1743-6109.2005.00090.x
Petrovic N, Schacke W, Gahagan JR, O’Conor CA, Winnicka B, Conway RE, Mina-Osorio P, Shapiro LH (2007) CD13/APN regulates endothelial invasion and filopodia formation. Blood 110(1):142–150. doi:10.1182/blood-2006-02-002931
Amadeu T, Braune A, Mandarim-de-Lacerda C, Porto LC, Desmoulière A, Costa A (2003) Vascularization pattern in hypertrophic scars and keloids: a stereological analysis. Pathol Res Pract 199(7):469–473. doi:10.1078/0344.0338.00447
Acknowledgments
We would like to thank Dr. Anthony Behrmann and his staff for their generous provision of skin tissue samples. This study was supported by the Canadian Institute of Health Research (CIHR-MOP-13387). Amy Lai holds a CIHR-SRTC award.
Conflict of interest statement
The authors state no conflict of interest.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Lai, A., Ghaffari, A. & Ghahary, A. Inhibitory effect of anti-aminopeptidase N/CD13 antibodies on fibroblast migration. Mol Cell Biochem 343, 191–199 (2010). https://doi.org/10.1007/s11010-010-0513-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11010-010-0513-7