
Abstract
Context
Many carnivores are attracted to rugged terrain, rocky areas, and conspicuous relief features. However, most of the previous research is limited to general topographical habitat characteristics and rarely consider the effects of microhabitat characteristics.
Objectives
We used the Eurasian lynx (Lynx lynx) as a model species to investigate the effects of microhabitat characteristics and human infrastructure on habitat selection. We also tested whether there is evidence for a functional response in habitat selection across a large gradient of habitat availability.
Methods
We developed a new approach for detecting rocky outcrops from airborne LiDAR data. In combination with other remote sensing techniques and GPS-telemetry data, we assessed lynx habitat selection and functional responses across two geologically contrasting areas in Europe.
Results
We detected > 1 million rocky outcrops and confirmed their strong selection by lynx. Lynx also selected steep, rugged, and rocky areas, especially for day-resting sites. Furthermore, lynx avoided paths during the day but selected them and other linear anthropogenic infrastructure during the night, indicating the behaviour-specific impact of human infrastructure. We also observed a functional response in the selection of rocky and rugged areas, as lynx’ selection of such habitats increased with their lower availability. This highlights the importance of preserving such terrains, especially when they are rare in a landscape.
Conclusions
Our results highlight the importance of incorporating remote sensing techniques and data on microhabitat features in animal habitat selection research. We also recommend caution when developing new infrastructure for human recreation or promoting its use near geomorphological features and in rugged terrain.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Animal communities and their habitats are shaped by a wide spectrum of biotic and abiotic factors and processes (Hugget 2004; Gordon et al. 2022). Habitats vary in shape and size, ranging from microhabitats to very large megahabitats, in which biota, landscape elements, and geomorphic processes interact and drive animal behaviours at various spatial scales (Johnson 1980; Huggett 2004; Gordon et al. 2022). Besides providing habitat and refugia for plants and animals, microtopography and specific relief features (i.e., geodiversity) may influence the ecology and various behaviours of wild animals, such as their foraging, movement, dispersal, breeding, and resting, which are essential for the survival and persistence of populations (Podgórski et al. 2008; Bouchet et al. 2015; Fitzsimons and Michael 2017; Bailey et al. 2018; Hočevar et al. 2021; Čonč et al. 2022; Smith and Mullins 2022). Large-bodied apex predators require extensive home ranges with good habitat connectivity, which is also important for dispersal that allows gene flow within and among different populations (Premier et al. 2020). Consequently, most studies relating to habitat use, habitat-suitability modelling or habitat connectivity focus on spatial analysis at a large scale: large contiguous natural areas, entire countries or even continents (e.g., Bouchet et al. 2015; Muhly et al. 2019; Ripari et al. 2022; Oeser et al. 2023; Tucker et al. 2023).
Previous research has shown that felids, such as cougars (Puma concolor), lions (Panthera leo), leopards (Panthera pardus), snow leopards (Panthera uncia), bobcats (Lynx rufus), and Eurasian lynx (Lynx lynx) are attracted to rugged and inaccessible terrain, rocky areas, mountain ridges, and conspicuous relief features (Jackson 1996; Dickson and Beier 2007; Bouchet et al. 2015; Abouelezz et al. 2018; Ironside et al. 2018; Farhadinia et al. 2020; Hočevar et al. 2021; Schmidt et al. 2023). However, these studies were usually restricted to single study areas. Consequently, our capacity of understanding the functional responses in habitat selection (i.e., how an individual adjusts their use of a particular habitat characteristic with changes in its availability) is limited (Mysterud and Ims 1998; Holbrook et al. 2019; Oeser et al. 2023). Furthermore, topographical predictor covariates are often limited to general terrain characteristics, such as elevation, ruggedness, and slope. Thus, large-scale studies typically do not consider the impact of microhabitat characteristics, like various relief features (Bailey et al. 2018; Ironside et al. 2018; Čonč et al. 2022). This is mainly due to the lack of detailed GIS layers containing information on such features across larger areas. Until the development of advanced remote sensing techniques, tasks, such as field mapping, digitising relief features, and the quantification of vegetation availability, were time consuming and costly, which resulted in low accuracy and poor data quality (Čonč et al. 2022; Ciglič et al. 2022; Sergeyev et al. 2023). High-resolution data, such as satellite images or LiDAR (Light Detection and Ranging)-based digital terrain models (hereafter DTM), combined with various (semi-)automatic methods for detecting and delineating relief and vegetation features, now enable no-contact, low-cost, and accurate mapping of large, remote, and densely forested areas (Bailey et al. 2018; Oeser et al. 2020; Ciglič et al. 2022; Čonč et al. 2022).
Besides the terrain characteristics, anthropogenic impacts can also have an important influence on animal behaviour and habitat selection, especially in human-dominated landscapes (Filla et al. 2017; Muhly et al. 2019; Ripari et al. 2022; Tucker et al. 2023). Although many large carnivores inhabit remote and/or protected natural areas, human recreational activities and infrastructure development have also increased in such areas during the past decades (Belotti et al. 2012, 2018; Muhly et al. 2019). Human infrastructure can have a positive or negative impact on animals. Specifically, during the day, when human disturbance is generally higher, animals tend to avoid human infrastructure. However, at night, when the disturbance is reduced, carnivores may be attracted to anthropogenic features, for example for scent marking, looking for prey or moving through the territories (Filla et al. 2017; Krofel et al. 2017; Muhly et al. 2019; Tucker et al. 2023). Additionally, animals often use abandoned human-modified areas, for example old quarries or artificial tunnels (Davis 1979; Marchewka and Postawa 2022); thus, human infrastructure could also be understood or interpreted as anthropogenic landforms (Li et al. 2017).
In the past, conservation and wildlife research focused primarily on biotic elements or general topographical characteristics, which limited our understanding of the importance of fine scale geodiversity (Albano 2015; Bailey et al. 2018; Gordon et al. 2022). However, microhabitats can facilitate carnivore adaptation to human pressure, providing cover for resting, and hosting prey species (Podgórski et al. 2008; Oeser et al. 2023). Understanding the relationship between spatial patterns, landscape heterogeneity, and ecological processes is crucial (Turner and Gardner 2015; Hersperger et al. 2021). Therefore, microhabitat characteristics, as well as specific relief features, should be incorporated in studies focusing on habitat selection, habitat connectivity and habitat suitability modelling, by including microhabitat data at finer scales.
To investigate the impact of rocky relief features (i.e., cliffs, rocky blocks, rocky ridges, karrens, and similar rocky features; hereafter ‘rocky outcrops’), as well as other abiotic microhabitat characteristics and human infrastructure on the Eurasian lynx (hereafter lynx) habitat selection, we selected two geologically and (bio-)geographically contrasting areas in Europe: the Slovenian Dinaric Karst (carbonate karst) and the Bohemian Forest Ecosystem (fluvial, glacial, and periglacial relief). We used the lynx as a model species due to their widespread distribution, general attraction to rugged and rocky terrain, and the importance of several relief features for their behavioural ecology (Krofel et al. 2007, 2017; Allen et al. 2017; Signer et al. 2019; Hočevar et al. 2021; Čonč et al. 2022; Schmidt et al. 2023). Similar to research on most other large carnivores, the majority of these studies on lynx habitat use have relied almost exclusively on a limited amount of data on microhabitat characteristics obtained in the field or by digitising relief features. Our main goals were to explore lynx selection of rocky outcrops as well as rugged and rocky terrain and to test for evidence of a functional response in lynx selection across a large gradient of availability of such terrain. To our knowledge, our study is one of the first that primarily relies on remote sensing data to identify detailed and accurate information on the microhabitat characteristics and studied relief features over a large area.
To examine how lynx select rocky outcrops and other abiotic microhabitat characteristics, as well as human infrastructure within their home ranges, we (1) developed a semi-automatic approach for identifying and delineating rocky outcrops from high-resolution LiDAR data and (2) evaluated the selection of these features by lynx in respect to their availability. To account for different behavioural states of lynx activity, we evaluated the selection under a used-available approach for two different behavioural states: day-resting sites, when lynx are typically resting on daybeds, and night locations, when lynx are engaged in various active behaviours (e.g., movement, foraging, territorial marking; Heurich et al. 2014; Hočevar et al. 2021). We tested the following hypotheses:
H1
Lynx select rocky outcrops, rugged terrain, and rocky surfaces during the day and at night.
H2
Lynx avoid the vicinity of human infrastructure (e.g., forest roads, paths, scattered buildings, and roads) for day-resting sites but select it during the nighttime.
H3
The selection of topographic features and avoidance of anthropogenic features is more pronounced during the day than at night.
H4
Lynx exhibit a functional response in selecting rocky outcrops, rugged terrain, and surface rockiness, with a stronger selection when availability is lower.
Study area
Dinaric Karst (Slovenia)
The Slovenian Dinaric Karst (hereafter SDK) is a karst landscape in the west and south of Slovenia (Fig. 1) formed on Mesozoic and Cenozoic rocks, mainly limestone and dolomite. Geographically and geologically, it is part of the Dinaric Mountains (Mihevc et al. 2010). The SDK is characterised by high and low karst plateaus, corrosion plains, and karst poljes running in a distinctive Dinaric direction (NW–SE), as well as by diverse surface and subsurface features (Mihevc et al. 2010; Zorn et al. 2020). In terms of lithology, karst types are divided into limestone karst and dolomite karst. Limestone karst exhibits a high density of caves, conical hills, karrens, and karst depressions (e.g., solution and collapse dolines). Dolomite karst, or fluviokarst, is characterised by dry shallow valleys, or dells, while other typical (sub)surface karst features are less common than in limestone karst (Zorn et al. 2020). The elevation in the study area ranges from about 200 to 1800 m.

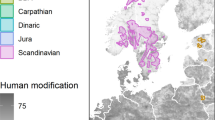
Map of the lynx home ranges in two selected study areas: the Slovenian Dinaric Karst (SDK) and Bohemian Forest Ecosystem (BFE) with lynx on rocky outcrops. Because some of the home ranges of lynx in the SDK included areas that were outside the study area, where LiDAR data were not available (i.e., Croatia), we included only the locations within the study area (Slovenia) in the analysis. Photos by ŠČ and MK
The SDK is known for dense forests, mainly Dinaric fir-beech associations (Omphalodo-Fagetum), extensive livestock farming, logging, hunting, and scattered small human settlements usually located in larger levelled areas, such as karst poljes or low karst plateaus (Mihevc et al. 2010). Around 20% of all hiking paths in Slovenia are located in the SDK region (Cigale and Gosar 2018), with the area around Snežnik being particularly popular. An average of 10 crossings/day (peaking at 161 crossings/day) were observed on the educational path, with the highest intensity on weekends and during the summer (Koželj et al. 2021). The area also has a dense network of forest roads (approximately 2 km of forest roads/km2) that were primarily established for logging activities (Krofel et al. 2017), but this infrastructure is frequently used for tourism and recreation as well (Potočnik 1996). In the Snežnik area, an average of 39 vehicles/day crossed the main roads, while the crossing on the forest roads was lower with an average of 10 vehicles/day. The intensity of traffic is highest on weekends and in the summer months (Koželj et al. 2021). Other human recreation activities are concentrated in and around famous tourist areas and natural landmarks (Cigale and Gosar 2018), mostly located on karst poljes.
Lynx were exterminated at the beginning of the twentieth century, but were reintroduced in 1973 and their current density is estimated to be 0.51 lynx/100 km2 (Fležar et al. 2023a). The main prey of the lynx are wild ungulates, which account for 88% of the biomass consumed by lynx, with roe deer (Capreolus capreolus) being the main prey species (Krofel et al. 2011).
Bohemian forest ecosystem (Germany and Czech Republic)
The Bohemian Forest Ecosystem (hereafter BFE) encompasses the Bavarian Forest National Park (240 km2) and a surrounding Bavarian Forest Natural Park (3007 km2) in Germany, and the Šumava National Park (690 km2) and the Bohemian Forest Protected Landscape Area (~ 1000 km2) in the Czech Republic (Fig. 1). It is one of the largest strictly protected forest areas in Central Europe (Navrátil et al. 2014; Heurich et al. 2015).
The BFE encompasses a fault-block mountain range on the border between Germany and the Czech Republic. Geographically and geologically, the BFE is part of the Bohemian Massif, which consists of Variscan igneous and metamorphic rocks, mainly gneiss and granite (Babůrek et al. 2006; Krause and Margold 2019). The BFE mountains were glaciated during the Pleistocene cold periods, resulting in the development of glacial features, such as cirques, cliffs, short glacial valleys, moraines, erratic blocks, and glacial lakes (Hauner et al. 2019; Krause and Margold 2019). It is also characterised by a fluvial relief, resulting in features such as gorges, river valleys, ridges, alluvial plains, cascades, and different sizes and shapes of rocky features. The elevation in the study area ranges from 600 to 1450 m.
The most common tree species at higher elevations is Norway spruce (Picea abies), complemented by mountain ash (Sorbus aucuparia). At lower elevations Norway spruce, European beech (Fagus sylvatica), and silver fir (Abies alba) are most common (van der Knaap et al. 2020).
The area is characterised by extensive land use and a low population density with < 2 people per km2 in the core area and 30–70 people per km2 in the peripheral areas (Heurich et al. 2015). In the national parks, the main human activities are related to recreation (e.g., hiking, cycling, horse-back riding, mushroom picking). In their core zones, such activities are only permitted on marked paths (Belotti et al. 2018). Recreation intensity is highest at weekends, peaks in July–August, and is concentrated during light hours. A maximum of around 5300 visitors/day and a total of around 1,840,000 people/year were counted at the main recreation sites of the national parks. The number of daily visits varies considerably between sites (Seibold and Shao 2014; Porst et al. 2020). Recreation in the peripheral areas is less intensive and human activities are more strongly associated with transportation, forestry and agriculture (Seibold and Shao 2014; Belotti et al. 2018).
In 2018, the density of the lynx population in BFE was estimated to be 1.33 lynx/100 km2 (Palmero et al. 2021). Roe deer accounts for up to 80% of the animals killed by lynx (Belotti et al. 2015).
Materials and methods
Rocky outcrops detection
Geological features that protrude above the surface of the surrounding land are defined as rocky outcrops (Twidale 2000). They are characterised by higher steepness, spatial isolation, and in many cases represent relatively undisturbed natural habitats. They occur in different bedrock and landscape types and form landscape features that vary in size and shape (examples shown in Fig. S1; Fitzsimons and Michael 2017; Smith and Mullins 2022).
Existing methods for the automatic detection of rocky outcrops are mostly based on satellite images (Kwok et al. 2018), which can cause rocky outcrops under forest canopy to be overlooked, or on the delineation from DTM based only on slope values (Smith and Mullins 2021), which can miss small outcrops on flat areas or misinterpret rocky outcrops as steep (soil-)covered slopes. Therefore, we developed a new two-step approach for the quantitative detection of rocky outcrops (Fig. 2) based on the spatial analysis of a high-resolution DTM (1 m × 1 m) and field measurements of the morphographic and morphometric characteristics of rocky outcrops. To create a DTM, we used LiDAR data from aerial laser scanning that was available in the form of a cloud of ground points (LAS files) provided by the Slovenian Environment Agency, the Bavarian Land Survey, and the administrations of Šumava National Park and Bavarian Forest National Park. The estimated density of the ground points in BFE is 55 points/m2, while the density in the SDK is considerably lower with an estimation of 0.5 per m2 in forested areas, respectively (Triglav Čekada and Bric 2015; Krzystek et al. 2020). We used ESRI software ArcGIS Pro 3.0.3 (ESRI, Redlands, CA, USA) to detect the rocky outcrops.

Flowchart of rocky outcrop detection. The grey boxes represent tools for raster analyses and raster layers, and the blue ovals represent vector layers
The Topographic Position Index (TPI; Weiss 2001) can be used to determine flat, concave (valleys), and convex (ridges) areas. Since rocky outcrops protrude above the surface of the surrounding land and are convex in shape, we identified all the convex-shaped areas in the first step. To calculate the TPI, we used the elevation for each cell (input DTM) and subtracted it by the mean elevation of the neighbouring cells (calculated by the Focal Statistics tool; Weiss 2001; Fig. 2). We tested different thresholds (5 m, 10 m, 15 m, 20 m, and 25 m) and found the 10 m search radius to be the most suitable for detecting convex features of different sizes (Čonč and Breg Valjavec 2023). Bigger ridges tend to be missed at lower thresholds, while smaller ridges are missed at higher thresholds (Weiss 2001). In the next step, we used the slope values of the rocky outcrops measured in the field as an additional criterion for rocky outcrop detection. Specifically, we measured the slope values between the lower and upper part of the rocky outcrops to determine the average slope values at which bedrock protrudes from the surface. In the SDK, the bedrock in areas with limestone outcropped at a lower surface slope (30°) than in areas with dolomite (50°) or other bedrock types (54°). In BFE (gneiss and granite), the bedrock outcropped at a surface slope of 40°. Based on the DTM (1 m × 1 m), we calculated Slope and assigned it to the slope categories determined in the field. We used the Select by Location tool to delineate the ridges (i.e., convex relief features) in each geological unit (limestone, dolomite, other, gneiss and granite) that overlapped with the determined slope values as rocky outcrops. In the final step, we calculated the heights of the rocky outcrops.
To assess the accuracy of the developed approach for rocky outcrop detection, we compared the results of the semi-automatically detected rocky outcrops with the Sky-View Factor layer for surface visualisation, created from a DTM (Zakšek et al. 2011), topographic maps, and field-mapped rocky outcrops (164 mapped in the SDK and 133 in BFE).
Lynx habitat selection in respect to rocky outcrops
Telemetry data
We used GPS-telemetry data from a total of 22 lynx (14 males and eight females; Table S1), with 12 lynx (eight males and four females) captured and collared between 2006 and 2023 in the SDK and 10 lynx (six males and four females) from the BFE captured between 2005 and 2012 (Table S1). Standardized protocols were used to capture, collar, and track the lynx (Heurich 2011; Krofel et al. 2013; Belotti et al. 2015). In both study areas, the animals were equipped with GPS-GSM collars (Vectronic Aerospace GmbH, Germany and TVP Positioning AB, Sweden; for details, see Heurich 2011; Krofel et al. 2013 and Topličanec et al. 2022). The GPS collars had an accuracy of 4–16 m, depending on habitat type (Stache et al. 2012). Some individuals were captured multiple times and recollared to prolong the tracking period.
In the SDK, nine residential lynx were captured in the study area and three lynx were released as part of a reinforcement project (Topličanec et al. 2022; Fležar et al. 2023b). The collars transmitted the lynx’ locations over a period of 1.9 to 38.1 months (Table S1) and were scheduled to obtain 3–8 GPS locations per day (variable among individuals and time periods) (Krofel et al. 2013; Topličanec et al. 2022).
The collars in the Bohemian Forest Ecosystem transmitted locations over a period of 3.5–18 months (Table S1) and were scheduled to store two locations daily. In addition, the collars recorded one location per hour every second week from 15:00 to 7:00 the following day, and two additional locations per day were recorded at dusk and dawn for a period of one month in each season of the year (Belotti et al. 2015; Filla et al. 2017). Prior to the analyses, we performed quality controls and scanned all the GPS data. Specifically, we detected and removed locations that indicated the collar malfunctioning, such as duplicated locations, locations falling in impossible places (e.g., large water bodies), or locations with a low degree of reliability (e.g., a very large distance between two consecutive points in a short amount of time).
Detection of behavioural states
The dataset contained 19,230 GPS locations, with each individual contributing between 61 and 2583 locations (Table S1). To differentiate between the behavioural states (e.g., resting, feeding, moving, searching for food) and phases (night and day) of lynx activity, we divided the data into two different categories: day-resting sites and night locations. A two-step approach was used to define the day-resting sites. Specifically, since lynx are primarily active during the night and twilight and generally rest in the middle of the day (Podolski et al. 2013; Heurich et al. 2014; Filla et al. 2017), we defined the daytime locations (9:00 a.m. to 3:00 p.m.) as potential day-resting sites. To confirm inactivity, we used activity data from built-in accelerometers for 14 lynx as a second criterion (Signer et al. 2019; Hočevar et al. 2021). Activity data with values of 0–11% of maximum activity were considered as corresponding to the resting behaviour (Heurich et al. 2014; Hočevar et al. 2021). In cases where multiple locations were recorded for the same individual during the daytime period on the same day, the location with the lowest activity value was included in the analysis, and in cases where no activity data was available, we considered the GPS location with the closest time to noon as a day-resting site. We classified GPS locations during the high-activity phase as night locations (from 7:00 p.m. to 7:00 a.m.) (Heurich et al. 2014).
To account for lynx selection of familiar habitat (i.e., within their home ranges – 95% MCP), we removed the periods of long-distance mating excursions (i.e., the period when they temporarily left their territory to find additional mates; Oliveira et al. 2023) and periods for females during the denning period (birth date and the following 60 days; Mattisson et al. 2022; Dalpiaz 2023). Furthermore, we removed the periods of the initial exploratory excursions for translocated lynx (i.e., the period before they established their territories; Topličanec et al. 2022).
Predictor covariates
We used 11 predictor covariates to test for the selection or avoidance of habitat features by lynx and categorised them into two groups: topographical and human infrastructure (Table 1). All the covariates were mapped and processed in ArcGIS Pro. The data for calculating the predictor covariates were provided by the Slovenian Environment Agency (ARSO-Agencija RS za okolje), the administrations of Šumava National Park and Bavarian Forest National Park, the Bavarian Land Survey (Bayerisches Vermessungsamt), OpenStreetMap (https://www.openstreetmap.org), the State Administration of Land Surveying and Cadastre (ZABAGED, ČÚZK-Český úřad zeměměřický a katastrální), and the Surveying and Mapping Authority of the Republic of Slovenia (GURS-Geodetska uprava RS; Table 1).
Within the topographical features, we obtained elevation data with a cell resolution of 1 × 1 m in the form of a LiDAR-DTM. Based on the DTM, we further calculated the aspect, rocky outcrops, slope, Topographic Position Index (TPI; Weiss 2001), and Vector Ruggedness Measure (VRM; Sappington et al. 2007). For the aspect, slope, and VRM, we calculated the mean values of the raster layers within a 25 m buffer of each point. We used a 25 m buffer to account for the GPS measurement error (Stache et al. 2012) and to consider the characteristics of the close proximity of lynx locations. We considered rocky outcrops as two different predictor covariates: the distance to the closest rocky outcrop (with a height ≥ 3 m; Signer et al. 2019) and surface rockiness based on the calculation of the share of area of (all) rocky outcrops within a 25 m buffer around each location.
Since lynx habitat selection is known to be influenced by the presence of human activities (Filla et al. 2017; Ripari et al. 2022; Oeser et al. 2023), we also included covariates describing human infrastructure in the analysis. We calculated the distances from scattered buildings and transportation infrastructure. Scattered buildings (e.g., forest cabins, which are regularly used for scent-marking by lynx; Allen et al. 2017) included buildings within categories 3–5 of the Corine Land Cover (CLC) data provided by the European Environment Agency that are at the same time at least 200 m away from the 1st and 2nd category of CLC. We divided transportation infrastructure into three categories: forest roads, paths, and transport infrastructure (Table 1). We grouped paths used for hiking, cycling, horse riding, and similar as paths, while the transport infrastructure layer included all other major roads and railways. The mean values with a standard deviation and the median values with interquartile ranges of covariates for used and available locations by both study areas are shown in Table S2.
Data analyses
We analysed lynx selection of topographical features and human infrastructure at the 3rd order of habitat selection, i.e., the home-range level (Johnson 1980). Specifically, we used a use-availability approach to compare a used sample (i.e., day -resting sites and night locations) with the available habitat (sample of random locations, see below for details) within 95% minimum convex polygon (MCP) home ranges for each individual lynx. As the home ranges of some lynx in the SDK included areas outside the study area (their home ranges extend across the Slovenian-Croatian border), we considered only the locations within the study area for which LiDAR-based topographical data were available.
We estimated the availability using a systematic approach (Benson 2013), by sampling points at a 100 × 100 m resolution, created with ArcGIS Pro 3.0.3. We set a value of 100 m based on the GPS locations resolution (see Sect. "Telemetry data" and "Detection of behavioural states") and a high density of rocky outcrops, especially in the SDK study area (Benson 2013).
To test the first three hypotheses (H1–3), we built generalized linear mixed-effect models (GLMMs) implemented in the R (R Core Team 2023) package “lme4” v.1.1–30 (Bates et al. 2015) with a binary (0 = available, 1 = used) response variable (Table 2). Lynx IDs were included as random effects to account for interindividual variation and the uneven sample size (Gillies et al. 2006). We built models with different groups of covariates considering their potential influence on the selection of topographical features and human infrastructure by lynx for both behavioural states (Table 2). We ranked the models based on the Akaike information criteria (AIC) and calculated the ∆AIC per model. We considered the models with ∆AIC values < 2 as the best models (Burnham and Anderson 2002). We also included a null model to test the strength of our hypothesis and the importance of the chosen covariates. We used k-fold cross-validation to assess model fit (Boyce et al. 2002) and randomly split the data into 80%/20% for modelling and testing, respectively. We repeated this procedure five times (k = 5) and then used the Spearman rank correlation to evaluate the relationships between the model ranks and the withheld subsample of the data. A well-performing model is expected to demonstrate a robust correlation (ρ > 0.80; Boyce et al. 2002). Prior to modelling, we checked for collinearity between covariates using a variance inflation factor (VIF), and we kept all predictor covariates since the VIF value was < 3 (Zuur et al. 2010). We rescaled the continuous covariates to z-scores to improve model convergence by subtracting their mean and dividing by 2 standard deviations (Gelman 2008).
To test for a functional response in habitat selection according to the availability of rocky outcrops, rugged, and rocky terrain within home ranges (H4), we created three different generalised additive models (GAMs), i.e., one per covariate. We first included random slopes for these three covariates in the two best ranked GLMM models for day-resting sites and night locations (Table 2), and then we extracted individual-level coefficients. These coefficients were then included as a response variable in the GAMs. We included the mean distance (rocky outcrops) or value (surface rockiness and VRM) of each covariate across the home range of each lynx as the predictor variable. These means represented the availability of the habitat variable to the individual lynx in question. We specified predictor variables as non-parametric smooth functions in the GAMs to allow non-linear relationships (Wood 2017). We used the R package “mgcv” v.1.8–33 (Wood 2015) to build GAMs.
Results
Rocky outcrops detection
We identified 984,833 rocky outcrops within the lynx home ranges in the SDK study area (area size: 1184.1 km2) with an average density of 831.7 rocky outcrops/km2. The highest number (n = 881,895) and average density (1115.2 rocky outcrops/km2) was recorded in the limestone area (790.8 km2, 66.8% of the study area), compared to the dolomite area (308.1 km2, 26.0% of the study area) where the number (n = 89,436) and average density (290.3 rocky outcrops/km2) of rocky outcrops was considerably lower. Only 14,546 rocky outcrops were detected in parts with other bedrock types (85.3 km2, 7.2% of the study area), with an average density of 17.1 rocky outcrops/km2.
In the BFE study area (area size: 1914.7 km2), we identified 80,590 rocky outcrops within lynx home ranges with an average density of 42.1 rocky outcrops/km2. The highest number (n = 63,427) of rocky outcrops with an average density of 45.4 rocky outcrops/km2 was recorded in the gneiss area (1396.5 km2, 72.9% of the study area), followed by a considerably lower number (n = 10,658) and average density (28.3 rocky outcrops/km2) of detected rocky outcrops in areas with granite (376.5 km2, 19.7% of the study area). Even though the areas with other bedrock types encompass the smallest area and proportion of the BFE (141.9 km2, 7.4% of the study area) and that we detected the lowest number of rocky outcrops (n = 6790) there, the average density was the highest here with 47.9 rocky outcrops/km2.
Additionally, we compared the results of the detected rocky outcrops with layers for surface visualisation (Hillshade and Sky View Factor created from a DTM), topographic maps, and field-mapped rocky outcrops (164 mapped in the SDK and 133 in BFE). A total of 153 (93.3%) in the SDK and 111 (83.5%) in BFE of field-mapped rocky outcrops matched with the automatically detected ones. By comparing automatically detected rocky outcrops with digitised ones (n = 627) from topographic maps in BFE that were used by Signer et al. (2019), a total of 620 (98.9%) rocky outcrops matched.
Lynx habitat selection
Day-resting sites
We used 2841 day-resting sites from 18 lynx (eight from the SDK and 10 from BFE; GPS collars of four lynx from the SDK were programmed to collect only night locations; Table S1) to assess the selection of topographic characteristics and human infrastructure by comparing them to available locations (n = 226,117). According to the AIC values, the model that best explained lynx day-resting site selection included both covariate groups: topographical and human infrastructure (Table 2). The k-fold cross-validation revealed a very high model fit (p = 0.97; Table 2). All the topographical covariates except aspect were statistically significant (Fig. 3). Specifically, we observed that for day-resting sites, lynx selected the vicinity of rocky outcrops, steeper slopes, areas with higher ruggedness, higher rockiness, and higher altitudes than available. The TPI showed that they selected areas where the index had higher values than what was available. The strongest selection was observed for the proximity to rocky outcrops and surface rockiness (Fig. 3). In the SDK, 73.2% of lynx resting sites and 51.2% of available locations were within a 50 m buffer of rocky outcrops, while in the BFE, the values were 31.1% for resting sites and 8.5% for available locations.

Estimated coefficients with 95% confidence intervals for the models with the best fit for lynx night locations (green) and day-resting sites (brown). The coefficients indicate the selection/avoidance of the target covariates and intervals that do not overlap with zero, indicating significant effects. Rocky outcrops, forest roads, paths, scattered buildings, roads, and railways are distance-based covariates (negative values indicate selection and positive avoidance)
Only distance to paths had a statistically significant effect among human infrastructure, indicating that lynx avoided paths when resting. Distance to forest roads, scattered buildings, and major transport infrastructure did not influence day-resting site selection (Fig. 3).
Night locations
We used GPS locations from 22 lynx (12 from the SDK and 10 from BFE; Table S1) to assess their habitat selection during the night by comparing used night locations (n = 16,389) to available locations (n = 226,117). According to the AIC values, the model that best explained habitat selection for night locations also included both covariate groups—topographical and human infrastructure (Table 2). The k-fold cross-validation revealed a moderate model fit for the best model (p = 0.73; Table 2). All the topographical covariates were statistically significant. We observed that during the night, lynx selected the vicinity of rocky outcrops, steeper slopes, more rugged and rocky surfaces, steeper slopes and higher altitudes, as well as areas with a higher TPI than available, but the values of the coefficients for continuous variables were lower than for day-resting site selection. Compared to southern exposures, they avoided other orientations (Fig. 3). In contrast to day-resting sites, the proportion of used locations within a 50 m buffer of rocky outcrops was lower than available in both study areas (43.9% of used vs. 51.2% of available locations in SDK and 0.6% of used vs. 8.5% of available locations in BFE).
All the covariates of the human infrastructure group were statistically significant. During the night, lynx avoided scattered buildings, but selected the vicinity of forest roads, paths, and major transport infrastructure. The strongest selection was observed for the proximity to forest roads, followed by the proximity to rocky outcrops and altitude (Fig. 3).
Functional response
We observed a functional response in the selection of rocky and rugged surface for day-resting sites and night locations. Specifically, lynx exhibited a stronger selection for rocky and rugged surfaces where such habitats were less available (particularly in the Bohemian Forest Ecosystem; Fig. 4). As the availability increased, we observed a general trend of avoidance of rocky and rugged surface. The main exception was at higher availability of rocky surfaces, where the lynx did not exhibit strong selection/avoidance patterns (Fig. 4). On the other hand, we did not detect a functional response in the selection of rocky outcrops at night or daytime periods, as their selection did not change with increased availability (Fig. 4). The AIC values, ΔAIC, and degrees of freedom of functional response models are reported in Table S3.

Selection of rocky outcrops, surface rockiness, and ruggedness by individual lynx as a function of the availability of the same covariates as predicted by generalized additive models. Each point indicates the value per individual lynx coloured by study areas (Slovenian Dinaric Mountains (SDK)—red and Bohemian Forest Ecosystem (BFE)—blue). Note that for the distance-based parameter (i.e., distance to rocky outcrops), the negative values represent selection and the positive values avoidance
Discussion
Rocky outcrops detection and high-resolution LiDAR-DTM
The use of a semi-automatic approach to analyse high-resolution DTM (LiDAR) enabled us to efficiently detect and measure a very large number (more than 1 million) of studied relief features (i.e., rocky outcrops) in two densely forested, large (> 1000 km2), and remote landscapes of the Slovenian Dinaric Mountains (SDK) and the Bohemian Forest Ecosystem (BFE). Such a task would be nearly impossible or would require enormous effort if done by traditional methods, such as field mapping or a visual digitisation of features based on different layers (e.g., topography maps with a much lower resolution or Hillshade derived from the DTM).
In the BFE, Signer et al. (2019) used a layer of digitalised rocky outcrops from topographic maps (1:25,000) that contained merely 627 polygons of rocky outcrops, while we identified 80,590 rocky outcrops in the same study area. In addition to fewer detected features, the previously-used digitised layer was spatially inaccurate as some polygons were mapped at the wrong locations, many outcrops were missing, and the shapes of polygons did not match with feature shapes (examples shown in Figs. S2 and S3). This highlights the potential of LiDAR-DTMs for large-scale ecological research, as they can be a useful and more reliable source of information for efficient detecting and studying the spatial patterns and morphometric settings of rocky outcrops, other landforms, or vegetation characteristics (Čonč et al. 2022; Smith and Mullins 2022; Sergeyev et al. 2023). Our study also showed that the abundance and spatial distribution of the rocky outcrops is closely related to the geology of the study areas. Rocky outcrops are much denser and more numerous on limestones than on dolomites, gneiss, and granites. We also found that they often occur on certain relief features, e.g., on the slopes of dolines and other karst depressions, valleys, cirques, or on ridges.
Nevertheless, the method had some drawbacks. When compared to field-mapped rocky outcrops, 6.7% of the rocky outcrops in the SDK and 16.5% in the BFE were not detected by the automatic method. The proportion of missed rocky outcrops could be caused by manual errors when placing the outcrops on the map or because some of the mapped rocky outcrops were too small (personal observations from the field) and were consequently not detected by the method (the search radius of the method was 10 m). By comparing automatically detected rocky outcrops with digitised ones from topographic maps in BFE, 98.9% automatically detected rocky outcrops matched with digitised ones. Those that did not match were mapped on the topographic maps on the wrong locations (i.e., there are no rocky outcrops there), which could be connected to the mapping of rocky outcrops on a small scale (example shown in Figs. S2 and S3).
In addition to the remote sensing of specific relief features, high-resolution DTMs can provide more realistic and accurate information about abiotic microhabitat characteristics for covariates derived from DTMs. While high-resolution LiDAR data is not available everywhere, our approach can also be applied to lower resolution data, such as TanDEM-X (12 × 12 m) provided by German Aerospace Center (DLR; https://tandemx-science.dlr.de), available worldwide; this would allow extending the analysis to more countries or even continental scales and enable faster processing. On the other hand, low resolution DTMs pose challenges, risking the loss of microhabitat details. The evaluation of the differences between input DTMs warrants further research.
Certain challenges are also posed by the GPS-telemetry data inaccuracies (estimated errors are 4–16 m, depending on habitat type; Stache et al. 2012). To account for GPS error, we would usually use buffers around the GPS locations and calculate the average values of covariates within the buffers (e.g., average slope). However, such a data generalisation can misrepresent the actual situation. For example, if a day-resting site is located in a flat area between rocky outcrops with a TPI value indicating “flat”, the calculation of the average value in a 50 m buffer may incorrectly result in the “upper slope” category, which does not correspond to reality. At the same time, these values can be misleading because the day-resting site might be on a flat area (e.g., rocky shelf) between or under a rocky outcrop (example shown in Fig. S4) and not necessarily on an (open) flat area. Therefore, the appropriate approach is to use distance-based covariates or the proportion of studied features in a given search area, as done in this study.
Lynx habitat selection and functional responses
Our results showed that lynx generally selected the vicinity of rocky outcrops, steep slopes, and areas with high ruggedness, rockiness, topographic position index, and altitudes for day resting in both study areas (H1 supported). These results are in line with the findings of previous studies. Specifically, in the Slovenian Dinaric Mountains, Hočevar et al. (2021) revealed a preference for lynx to rest on the top of mountain ridges or hilltops, as well as on steep slopes and microhabitats with good cover. These areas also provide concealment for the lynx, as well as a good view of the surrounding area to detect potential incoming danger or prey. Similarly, Signer et al. (2019) observed that lynx tend to select rocky outcrops when resting, 12.3% of day-resting sites were located within a 50 m buffer around these features. Our study improved the detection of rocky outcrops (i.e., using LiDAR DTM) and day-resting sites (i.e., incorporating accelerometer data), allowing us to observe that 31.1% of day-resting sites were located within a 50 m buffer around rocky outcrops in the same study area as in Signer et al. (2019). This comparison suggests that more detailed and accurate information on the studied features and lynx behaviour may have important implications for understanding lynx ecology.
During the night, when lynx are generally active (Heurich et al. 2014), we observed a similar selection of topographical covariates, which supports our first hypothesis. However, in line with our third hypothesis, the values of the coefficients were lower compared to the day-resting sites, indicating a weaker selection during nighttime (Table S2, Fig. 3). The selection of the vicinity of rocky outcrops, as well as rugged, steep and rocky terrain during the night may indicate that lynx select such terrain for scent-marking (Allen et al. 2017) or hunting (Krofel et al. 2007; Čonč et al. 2022). Felids often seek out features that serve as communication hubs (Melzheimer et al. 2020). Previous studies (Vogt et al. 2016; Allen et al. 2017; Mohorović and Krofel 2020) showed that lynx most often scent mark rocks and juvenile conifers, and among rocks, they often select those that are covered with moss (better absorption of urine and longer persistence) and shaped as overhangs (protection from the weather). As lynx are a stalking and ambush predator, their hunting may be more successful in rugged, rocky terrain and around rocky outcrops, which are often covered with moss. This enables a quieter approach and provides cover for the lynx, as well as escape impediments to ungulates (Krofel et al. 2007; Podgórski et al. 2008; Belotti et al. 2013; Čonč et al. 2022; Schmidt et al. 2023).
Beside the topographical characteristics, lynx were also influenced by anthropogenic factors. Our results indicated that, during the daytime, lynx avoided paths used for human recreation. This was expected, given that people mainly use these paths during the day, which is also when lynx are most vulnerable due to their lower alertness during sleeping periods. Therefore, they require safer (i.e., less accessible and less disturbed) locations (Belotti et al. 2012, 2018; Filla et al. 2017; Hočevar et al. 2021) while resting. Although metric data on the intensity of the recreational use of paths were not available for our study areas, further research incorporating such data would be important to understand the thresholds at which lynx start avoiding such infrastructure. In contrast to the day-resting sites, we observed that lynx selectively use linear human infrastructures (roads, paths) during the nighttime, when human use is reduced and lynx are more active. These findings support our second and third hypothesis. By using linear features, lynx may benefit from easier movement and the increased efficiency of scent-marking (Krofel et al. 2017). We observed that lynx avoid scattered buildings during the night, even though they regularly use them for scent-marking, especially abandoned or less used cabins (Allen et al. 2017; Krofel et al. 2017).
Finally, by considering a large gradient of availability of the microhabitats used by the lynx, we were able to test whether lynx exhibit functional responses within two regions in Europe, as hypothesised in previous local studies (Hočevar et al. 2021). Our analyses confirmed the functional response in the selection of rugged and rocky terrain during two behavioural states (day-resting sites and night locations). Indeed, lynx exhibited a stronger selection towards rugged and rocky terrain when the availability was lower. As the availability of such features increases, lynx might not show a particular selection pattern (no selection/avoidance), although they would still regularly use them. Our findings partially confirm our fourth hypothesis, as we did not detect a functional response in the selection of rocky outcrops. Despite the unknown ecological importance of these features, it is important to have in mind that we only considered rocky outcrops higher than 3 m. Therefore, our results do not reflect the selection towards smaller rocky outcrops, which could be equally important for different lynx behaviours. Furthermore, we observed a high individual variability, especially in the selection of rocky outcrops and rocky surface in both study areas, indicating that these features might be particularly relevant for some individuals. Such variability may rise from different reasons, such as intrinsic (sex, age) or external factors (such as seasonality, presence of conspecifics, prey type and availability). Moreover, the general increase of human recreational activities (e.g., hiking, geocaching, mushroom picking, and climbing, among others) in natural areas during the past decades might negatively affect lynx, especially during the daytime (Belotti et al. 2012, 2018). Thus, it would not be surprising if lynx increasingly select habitats that are less accessible to people, especially in areas where the availability of such terrain is limited. On the other hand, in areas with abundant rugged and rocky surfaces, their selection might not be so obvious or lynx might even avoid them in some behaviours (e.g., when moving through the landscape for the ease of movement).
Conclusions and management implications
The use of remote sensing methods and lynx GPS-telemetry data from two geographically and geologically contrasting study areas enabled us to show that lynx select the vicinity of rocky outcrops, as well as rugged and rocky terrain. This pattern was particularly pronounced when selecting day-resting sites and in areas with a lower availability of the preferred microhabitats. We also showed that lynx avoid paths during the day (i.e., while resting), while they are attracted to linear human infrastructure at night. This highlights the importance of evaluating habitat selection among different behavioural states, as the selection patterns might change considerably among activity phases.
Our results indicate that specific relief features may increase the suitability of a given area for lynx and possibly other felids. Furthermore, this study emphasises the importance of including such microhabitat characteristics in habitat selection analyses and landscape ecology in general. The results provided by such research can also be used for guiding conservation efforts and decision-making in spatial planning (Dickson and Beier 2007; Podgórski et al. 2008; Abouelezz et al. 2018; Bailey et al. 2018; Signer et al. 2019; Farhadinia et al. 2020; Čonč et al. 2022). For example, as human populations grow, disturbance of natural habitats and habitat fragmentation increases, with transportation infrastructure being among the main problems (Meijer et al. 2018). The approach used in our study to detect microhabitat characteristics could contribute to identifying the most suitable micro-sites for the placement (Huck et al. 2010; Potočnik et al. 2020), structure, and design of wildlife crossings over fragmentation barriers (Brennan et al. 2022). In addition, the results could also be useful for the selection of micro-sites for the deployment of camera-traps and box-traps to increase the efficiency of population monitoring and animal capture for research purposes (Fležar et al. 2023). Furthermore, our results indicate that rocky and rugged areas should be avoided whenever possible when planning new recreation infrastructure in lynx habitat (e.g., hiking paths, geocaching locations, and climbing routes). This appears to be especially important where such areas are rare in the landscape.
We draw special attention to the construction of new paths (e.g., for hiking, cycling, and horse-riding), towards which lynx seem to be particularly sensitive during the daytime. Lynx’ avoidance of paths appears to be stronger compared to forestry roads, indicating the importance of separating these types of linear human infrastructure in wildlife ecology and behavioural studies. However, it should first be mentioned that regulations in the use of forestry roads can differ among different regions, and second, the use of forestry roads for recreational purposes is generally increasing, especially with the recent availability of map applications.
In conclusion, our study provides further evidence for the importance of protecting geomorphological features (geodiversity) for biodiversity conservation, as rocky outcrops and other relief features provide an important habitat for lynx, as well as for numerous other endangered animals, plants, and fungi (Fitzsimons and Michael 2017; Bailey et al. 2018; Signer et al. 2019; Čonč 2020; Čarni et al. 2022; Čonč et al. 2022; Gordon et al. 2022; Smith and Mullins 2022).
Data availability
Data supporting the findings of this study are not included due to conservation concerns. Data may, however, be available from the authors upon reasonable request and with permission of Administrations of NP Bayerischer Wald, NP Šumava, BF UL and SFS.
References
Abouelezz HG, Donovan TM, Mickey RM, James DM, Freeman M, Royar K (2018) Landscape composition mediates movement and habitat selection in bobcats (Lynx rufus): implications for conservation planning. Landsc Ecol 33:1301–1318. https://doi.org/10.1007/s10980-018-0654-8
Albano CM (2015) Identification of geophysically diverse locations that may facilitate species’ persistence and adaptation to climate change in the southwestern United States. Landsc Ecol 30:1023–1037. https://doi.org/10.1007/s10980-015-0167-7
Allen ML, Hočevar L, de Groot M, Krofel M (2017) Where to leave a message? The selection and adaptive significance of scent-marking sites for Eurasian lynx. Behav Ecol Sociobiol 71:136. https://doi.org/10.1007/s00265-017-2366-5
Babůrek J, Pertoldová J, Verner K, Jiřička J (2006) Guide to the geology of Šumava Mountains. Czech Geological Survey, Prague
Bailey JJ, Boyd DS, Field R (2018) Models of upland species’ distributions are improved by accounting for geodiversity. Landsc Ecol 33:2071–2087. https://doi.org/10.1007/s10980-018-0723-z
Bates D, Mächler M, Bolker B, Walker S (2015) Fitting linear mixed-effects models using lme4. arXiv:1406.5823. https://doi.org/10.48550/arXiv.1406.5823
Belotti E, Heurich M, Kreisinger J, Šustr P, Bufka L (2012) Influence of tourism and traffic on the Eurasian lynx hunting activity and daily movements. Anim Biodivers Conserv 35:235–246. https://doi.org/10.32800/abc.2012.35.0235
Belotti E, Červený J, Šustr P, Kreisinger J, Gaibani G, Bufka L (2013) Foraging sites of Eurasian lynx Lynx Lynx: relative importance of microhabitat and prey occurrence. Wildlife Biol 19:188–201. https://doi.org/10.2981/12-077
Belotti E, Weder N, Bufka L, Kaldhusdal A, Küchenhoff H, Seibold H et al (2015) Patterns of Lynx predation at the interface between protected areas and multi-use landscapes in Central Europe. PLoS ONE 10:e0138139. https://doi.org/10.1371/journal.pone.0138139
Belotti E, Mayer K, Kreisinger J, Heurich M, Bufka L (2018) Recreational activities affect resting site selection and foraging time of Eurasian lynx (Lynx lynx) Hystrix. Italian J Mammal 29:181–189. https://doi.org/10.4404/hystrix-00053-2018
Benson JF (2013) Improving rigour and efficiency of use-availability habitat selection analyses with systematic estimation of availability. Methods Ecol Evol 4:244–251. https://doi.org/10.1111/2041-210x.12006
Bouchet PJ, Meeuwig JJ, Salgado Kent CP, Letessier TB, Jenner CK (2015) Topographic determinants of mobile vertebrate predator hotspots: current knowledge and future directions. Biol Rev 90:699–728. https://doi.org/10.1111/brv.12130
Boyce MS, Vernier PR, Nielsen SE, Schmiegelow FKA (2002) Evaluating resource selection functions. Ecol Modell 157:281–300. https://doi.org/10.1016/S0304-3800(02)00200-4
Brennan L, Chow E, Lamb C (2022) Wildlife overpass structure size, distribution, effectiveness, and adherence to expert design recommendations. PeerJ 12(10):e14371. https://doi.org/10.7717/peerj.14371
Burnham KP, Anderson DR (2002) Basic use of the information-theoretic approach. In: Burnham KP, Anderson DR (eds) Model selection and multimodel inference. Springer, New York, pp 98–148. https://doi.org/10.1007/978-0-387-22456-5_3
Čarni A, Čonč Š, Breg Valjavec M (2022) Landform-vegetation units in karstic depressions (dolines) evaluated by indicator plant species and Ellenberg indicator values. Ecol Indic 135:108572. https://doi.org/10.1016/j.ecolind.2022.108572
Cigale D, Gosar A (2018) Značilnosti in težnje razvoja turizma v Sloveniji. In: Cigale D, Lampič B, Potočnik Slavič I, Repe B (eds) Geografsko raziskovanje turizma in rekreacije v Sloveniji. Znanstvena založba Filozofske fakultete, Ljubljana, pp 13–46. https://doi.org/10.4312/9789610600183
Ciglič R, Čonč Š, Breg Valjavec M (2022) The impact of digital elevation model preprocessing and detection methods on Karst depression mapping in densely forested dinaric mountains. Remote Sens 14:2416. https://doi.org/10.3390/rs14102416
Čonč Š (2020) Pomen varovanja vročih točk geodiverzitete za ohranitev biotske pestrosti na območju Nature 2000 v dolini reke Dragonje. Geogr Vestn 92:9–28. https://doi.org/10.3986/GV92201
Čonč Š, Oliveira T, Portas R, Černe R, Breg Valjavec M, Krofel M (2022) Dolines and cats: remote detection of karst depressions and their application to study wild felid ecology. Remote Sens 14:656. https://doi.org/10.3390/rs14030656
Čonč Š, Breg Valjavec M (2023) Detection of rocky outcrops from LiDAR-derived DEM in Dinaric Karst. In: EGU General Assembly 2023. Vienna, Austria, 24–28 Apr 2023, EGU23-13606. https://doi.org/10.5194/egusphere-egu23-13606
Dalpiaz N (2023) Movement patterns of female Eurasian lynx (Lynx lynx) during the denning season across Europe. MSc thesis. Padova
Davis BNK (1979) Chalk and limestone quarries as wildlife habitats. Min Environ 1:48–56. https://doi.org/10.1007/BF02010717
Dickson BG, Beier P (2007) Quantifying the influence of topographic position on cougar (Puma concolor) movement in southern California, USA. J Zool 271:270–277. https://doi.org/10.1111/j.1469-7998.2006.00215.x
Farhadinia MS, Heit DR, Montgomery RA, Johnson PJ, Hobeali K, Hunter LTB, Macdonald DW (2020) Vertical relief facilitates spatial segregation of a high density large carnivore population. Oikos 129:346–355. https://doi.org/10.1111/oik.06724
Filla M, Premier J, Magg N, Dupke C, Khorozyan I, Waltert M, Bufka L, Heurich M (2017) Habitat selection by Eurasian lynx (Lynx lynx) is primarily driven by avoidance of human activity during day and prey availability during night. Ecol Evol 7:6367–6381. https://doi.org/10.1002/ece3.3204
Fitzsimons JA, Michael DR (2017) Rocky outcrops: a hard road in the conservation of critical habitats. Biol Conserv 211:36–44. https://doi.org/10.1016/j.biocon.2016.11.019
Fležar U, Aronsson M, Černe R, Pičulin A, Bartol M, Stergar M et al (2023) Using heterogeneous camera-trapping sites to obtain the first density estimates for the transboundary Eurasian lynx (Lynx lynx) population in the Dinaric Mountains. Biodivers Conserv 32:3199–3216. https://doi.org/10.1007/s10531-023-02646-3
Fležar U, Hočevar L, Sindičić M, Gomerčić T, Konec M, Slijepčević V et al. (2023b) Surveillance of the reinforcement process of the Dinaric—SE Alpine lynx population in the lynx-monitoring year 2021–2022. Technical report. Ljubljana
Gelman A (2008) Scaling regression inputs by dividing by two standard deviations. Stat Med 27:2865–2873. https://doi.org/10.1002/sim.3107
Gillies CS, Hebblewhite M, Nielsen SE, Krawchuk MA, Aldridge CL, Frair JL et al (2006) Application of random effects to the study of resource selection by animals. J Anim Ecol 75:887–898. https://doi.org/10.1111/j.1365-2656.2006.01106.x
Gordon JE, Bailey JJ, Larwood JG (2022) Conserving nature’s stage provides a foundation for safeguarding both geodiversity and biodiversity in protected and conserved areas. Parks Stewardship Forum 38. https://doi.org/10.5070/P538156118
Hauner U, Lehrberger G, Brugger M (2019) Der Naturraum Bayerischer Wald—Šumava in den Eiszeiten. Nationalparkverwaltung Bayerischer Wald, Grafenau
Hersperger AM, Grădinaru SR, Pierri Daunt AB, Imhof CS, Fan P (2021) Landscape ecological concepts in planning: review of recent developments. Landsc Ecol 36:2329–2345. https://doi.org/10.1007/s10980-021-01193-y
Heurich M (2011) Berücksichtigung von Tierschutzaspekten beim Fang und der Markierung von Wildtieren. In: 12 Internationale Fachtagung zu Fragen von Verhaltenskunde, Tierhaltung und Tierschutz. München, pp 142–158
Heurich M, Hilger A, Küchenhoff H, Andrén H, Bufka L, Krofel M et al (2014) Activity patterns of Eurasian lynx are modulated by light regime and individual traits over a wide latitudinal range. PLoS ONE 9:e114143. https://doi.org/10.1371/journal.pone.0114143
Heurich M, Brand TT, Kaandorp MY, Šustr P, Müller J, Reineking B (2015) Country, cover or protection: what shapes the distribution of red deer and roe deer in the Bohemian Forest Ecosystem? PLoS ONE 10:e0120960. https://doi.org/10.1371/journal.pone.0120960
Hočevar L, Oliveira T, Krofel M (2021) Felid bedrooms with a panoramic view: selection of resting sites by Eurasian lynx (Lynx lynx) in a karstic landscape. Behav Ecol Sociobiol 75:34. https://doi.org/10.1007/s00265-021-02977-7
Holbrook JD, Olson LE, DeCesare NJ, Hebblewhite M, Squires JR, Seenweg R (2019) Functional responses in habitat selection: clarifying hypotheses and interpretations. Ecol Appl 29:e01852. https://doi.org/10.1002/eap.1852
Huck M, Jędrzejewski W, Borowik T, Miłosz-Cielma M, Schmidt K, Jędrzejewska B, Nowak S, Mysłajek RW (2010) Habitat suitability, corridors and dispersal barriers for large carnivores in Poland. Acta Theriol 55:177–192. https://doi.org/10.4098/j.at.0001-7051.114.2009
Huggett RJ (2004) Fundamentals of biogeography. Routledge, London. https://doi.org/10.4324/9780203356586
Ironside KE, Mattson DJ, Arundel T, Theimer T, Holton B, Peters MC et al (2018) Geomorphometry in landscape ecology: issues of scale, physiography, and application. Environ Ecol Res 6:397–412. https://doi.org/10.13189/eer.2018.060501
Jackson MR (1996) Home range, movements and habitat use of snow leopard (Uncia uncia) in Nepal. Dissertation, University of London
Johnson DH (1980) The comparison of usage and availability measurements for evaluating resource preference. Ecology 61:65–71. https://doi.org/10.2307/1937156
Koželj G, Javornik J, Koblar Habič Š (2021) Končno poročilo o izvedbi delovnih sklopov DS T3.3.1 in T3.3.4: Usmerjanje obiska na območju Snežnik/Mašun. Technical report. Ljubljana
Krause D, Margold M (2019) Glacial geomorphology of the Šumava/Bayerischer Wald mountains, Central Europe. J Maps 15:719–725. https://doi.org/10.1080/17445647.2019.1661881
Krofel M, Potočnik H, Kos I (2007) Topographical and vegetational characteristics of lynx kill sites in Slovenian Dinaric mountains. Natura Sloveniae 9:25–36. https://doi.org/10.14720/ns.9.1.25-36
Krofel M, Huber D, Kos I (2011) Diet of Eurasian lynx Lynx lynx in the northern Dinaric Mountains (Slovenia and Croatia): importance of edible dormouse Glis glis as alternative prey. Acta Theriol 56:315–322. https://doi.org/10.1007/s13364-011-0032-2
Krofel M, Skrbinšek T, Kos I (2013) Use of GPS location clusters analysis to study predation, feeding, and maternal behavior of the Eurasian lynx. Ecol Res 28:103–116. https://doi.org/10.1007/s11284-012-1005-x
Krofel M, Hočevar L, Allen ML (2017) Does human infrastructure shape scent marking in a solitary felid? Mamm Biol 87:36–39. https://doi.org/10.1016/j.mambio.2017.05.003
Krzystek P, Serebryanyk A, Schnörr C, Červenka J, Heurich M (2020) Large-scale mapping of tree species and dead trees in Šumava National Park and Bavarian Forest National Park using Lidar and multispectral imagery. Remote Sens 12:661. https://doi.org/10.3390/rs12040661
Kwok CYT, Sing Wong M, Ho HC, Lo FLC, Ko FWY (2018) Deep learning approach for rock outcrops identification. In: 2018 Fifth International Workshop on Earth Observation and Remote Sensing Applications (EORSA), pp 1–6. https://doi.org/10.1109/EORSA.2018.8598577
Li J, Yang L, Pu R, Liu Y (2017) A review on anthropogenic geomorphology. J Geogr Sci 27:109–128. https://doi.org/10.1007/s11442-017-1367-7
Marchewka A, Postawa T (2022) Characteristics of underground shelters used by the Edible Dormouse (Glis glis) in its European range. In: ARPHA Conference Abstracts 5:e84663. https://doi.org/10.3897/aca.5.e84663
Mattisson J, Linnell JDC, Anders O, Belotti E, Breitenmoser-Würsten C, Bufka L et al (2022) Timing and synchrony of birth in Eurasian lynx across Europe. Ecol Evol 12:e9147. https://doi.org/10.1002/ece3.9147
Meijer JR, Huijbregts MAJ, Schotten KCGJ, Schipper AM (2018) Global patterns of current and future road infrastructure. Environ Res Lett 13:064006. https://doi.org/10.1088/1748-9326/aabd42
Melzheimer J, Heinrich SK, Wasiolka B, Mueller R, Thalwitzer S, Palmegiani I et al (2020) Communication hubs of an asocial cat are the source of a human–carnivore conflict and key to its solution. PNAS 117:33325–33333. https://doi.org/10.1073/pnas.2002487117
Mihevc A, Prelovšek M, Zupan Hajna N (2010) Introduction to the Dinaric Karst. Založba ZRC, Postojna. https://doi.org/10.3986/9789612541989
Mohorović M, Krofel M (2020) The scent world of cats: where to place a urine scent mark to increase signal persistence? Anim Biol 71:151–168. https://doi.org/10.1163/15707563-bja10018
Muhly TB, Johnson CA, Hebblewhite M, Neilson EW, Fortin D, Fryxell JM et al (2019) Functional response of wolves to human development across boreal North America. Ecol Evol 9:10801–10815. https://doi.org/10.1002/ece3.5600
Mysterud A, Ims RA (1998) Functional responses in habitat use: availability influences relative use in trade-off situations. Ecol 79:1435–1441. https://doi.org/10.1890/0012-9658(1998)079[1435:FRIHUA]2.0.CO;2
Navrátil J, Lesjak M, Pícha K, Martinát S, Navratilová J, White Baravalle Gilliam VL et al (2014) The importance of vulnerable areas with potential tourism development: a case study of the Bohemian forest and South Bohemia tourism regions. Acta Geogr Slov 54:115–130. https://doi.org/10.3986/AGS54108
Oeser J, Heurich M, Senf C, Pflugmacher D, Belotti E, Kuemmerle T (2020) Habitat metrics based on multi-temporal Landsat imagery for mapping large mammal habitat. Remote Sens Ecol Conserv 6:52–69. https://doi.org/10.1002/rse2.122
Oeser J, Heurich M, Kramer-Schadt S, Andrén H, Bagrade G, Belotti E et al (2023) Prerequisites for coexistence: human pressure and refuge habitat availability shape continental-scale habitat use patterns of a large carnivore. Landsc Ecol 38:1713–1728. https://doi.org/10.1007/s10980-023-01645-7
Oliveira T, Rodríguez-Recio M, Černe R, Krofel M (2023) High risk, high reward? Influence of experience level in the selection or avoidance of artificial feeding sites by Eurasian lynx. Glob Ecol Conserv 45:e02529. https://doi.org/10.1016/j.gecco.2023.e02529
Palmero S, Belotti E, Bufka L, Gahbauer M, Heibl C, Premier J et al (2021) Demography of a Eurasian lynx (Lynx lynx) population within a strictly protected area in Central Europe. Sci Rep 11:19868. https://doi.org/10.1038/s41598-021-99337-2
Podgórski T, Schmidt K, Kowalczyk R, Gulczyńska A (2008) Microhabitat selection by Eurasian lynx and its implications for species conservation. Acta Theriol 53:97–110. https://doi.org/10.1007/BF03194243
Podolski I, Belotti E, Bufka L, Reulen H, Heurich M (2013) Seasonal and daily activity patterns of free-living Eurasian lynx Lynx lynx in relation to availability of kills. Wildlife Biol 19:69–77. https://doi.org/10.2981/12-049
Porst F, Bečka P, Kučeravá B, Štemberk J, Allex B, Arnberger A, Eder R, Preisel H (2020) Grenzüberschreitendes sozioökonomisches monitoring in den Nationalparken Bayerischer Wald und Šumava in den Jahren 2017–2019. Grafenau, Vimperk
Potočnik I (1996) The multiple use of forest roads and their classification. In: Proceedings of the Seminar on Environmentally Sound Forest Roads and Wood Transport. Sinaia, Romania
Potočnik H, Črtalič J, Kos I, Skrbinšek T (2020) Characteristics of spatial use and importance of landscape features for recovering populations of Eurasian lynx (Lynx lynx). Acta Biol Slovenica 63:65–88. https://doi.org/10.14720/abs.63.2.15938
Premier J, Fickel J, Heurich M, Kramer-Schadt S (2020) The boon and bane of boldness: movement syndrome as saviour and sink for population genetic diversity. Mov Ecol 8:1–17. https://doi.org/10.1186/s40462-020-00204-y
Ripari L, Premier J, Belotti E, Bluhm H, Breitenmoser-Würsten C, Bufka L et al (2022) Human disturbance is the most limiting factor driving habitat selection of a large carnivore throughout Continental Europe. Biol Conserv 266:109446. https://doi.org/10.1016/j.biocon.2021.109446
Sappington JM, Longshore KM, Thompson DB (2007) Quantifying landscape ruggedness for animal habitat analysis: a case study using bighorn sheep in the Mojave Desert. J Wildl Manage 71:1419–1426. https://doi.org/10.2193/2005-723
Schmidt K, Górny M, Jędrzejewski W (2023) Effect of microhabitat characteristics for predicting habitat suitability for a stalking large carnivore—the Eurasian lynx in middle Europe. Anim Conserv 26:851–864. https://doi.org/10.1111/acv.12873
Seibold H, Shao S (2014) Analysis of camera trap data to evaluate the association between activity patterns of park visitors and animals. Statistical Consulting Report. Munich
Sergeyev M, Crawford DA, Holbrook JD, Lombardi JV, Tewes ME, Campbell TA (2023) Selection in the third dimension: using LiDAR derived canopy metrics to assess individual and population-level habitat partitioning of ocelots, bobcats, and coyotes. Remote Sens Ecol Conserv. https://doi.org/10.1002/rse2.369
Signer J, Filla M, Schoneberg S, Kneib T, Bufka L, Belotti E, Heurich M (2019) Rocks rock: the importance of rock formations as resting sites of the Eurasian lynx Lynx lynx. Wildl Biol 2019:1–5. https://doi.org/10.2981/wlb.00489
Smith WH, Mullins CZ (2022) Light detection and ranging (LiDAR)-assisted detection of rock outcrops in Appalachian hardwood forests. J Fish Wildl Manag 13:250–261. https://doi.org/10.3996/JFWM-21-043
Stache A, Löttker P, Heurich M (2012) Red deer telemetry: dependency of the position acquisition rate and accuracy of GPS collars on the structure of a temperate forest dominated by European beech and Norway spruce. Silva Gabreta 18:35–48
Topličanec I, Gomerčić T, Černe R, Krofel M, Pop IM, Kubala J et al (2022) Early post-release behaviour of Eurasian lynx translocated to the transboundary region of the Dinaric Mountains. J Vertebr Biol 71(22064):1–16. https://doi.org/10.25225/jvb.22064
Triglav Čekada M, Bric V (2015) Končan je projekt laserskega skeniranja Slovenije. Geodetski Vestnik 59:586–592
Tucker MA, Schipper AM, Adams TSF, Attias N, Avgar T, Babic NL et al (2023) Behavioral responses of terrestrial mammals to COVID-19 lockdowns. Science 380:1059–1064. https://doi.org/10.1126/science.abo6499
Turner MG, Gardner RH (2015) Introduction to landscape ecology and scale. In: Landscape ecology in theory and practice. Springer, New York. https://doi.org/10.1007/978-1-4939-2794-4_1
Twidale CR (2000) Granite outcrops: their utilisation and conservation. J R Soc West Aust 83:115–122
van der Knaap WO, van Leeuwen JFN, Fahse L, Sönke S, Studer T, Baumann J et al (2020) Vegetation and disturbance history of the Bavarian Forest National Park, Germany. Veg Hist Archaeobot 29:277–295. https://doi.org/10.1007/s00334-019-00742-5
Vogt K, Hofer E, Ryser A, Kölliker M, Breitenmoser U (2016) Is there a trade-off between scent marking and hunting behaviour in a stalking predator, the Eurasian lynx, Lynx lynx? Anim Behav 117:59–68. https://doi.org/10.1016/j.anbehav.2016.04.004
Weiss A (2001) Topographic position and land-forms analysis. In: Poster presentation, ESRI user conference. San Diego, USA
Wood S (2015) Package 'mgcv'. R package version 1:729
Wood S (2017) Generalized additive models: an introduction with R. Chapman and Hall, Boca Raton. https://doi.org/10.1201/9781315370279
Zakšek K, Oštir K, Kokalj Ž (2011) Sky-view factor as a relief visualization technique. Remote Sens 3:398–415. https://doi.org/10.3390/rs3020398
Zorn M, Ferk M, Lipar M, Komac B, Tičar J, Hrvatin M (2020) Landforms of Slovenia. In: Perko D, Ciglič R, Zorn M (eds) The geography of Slovenia. Springer International Publishing, Cham, pp 35–57. https://doi.org/10.1007/978-3-030-14066-3_3
Zuur AF, Ieno EN, Elphick CS (2010) A protocol for data exploration to avoid common statistical problems. Methods Ecol Evol 1:3–14. https://doi.org/10.1111/j.2041-210X.2009.00001.x
Acknowledgements
We would like to thank the staff (veterinarians, hunters, foresters, technicians, students and volunteers) of the University of Ljubljana, the Slovenia Forest Service, Ljubljana ZOO (Slovenia), the Association for the Biological Diversity Conservation (Romania), the Technical University in Zvolen (Slovakia), National Park Bayerischer Wald (Germany), and National Park Šumava (Czech Republic) for their invaluable help with capturing and collaring lynx, with especial thanks to Urša Fležar, Andrea Gazzola, Lan Hočevar, Pavel Kvapil, Jakub Kubala, Ioan-Mihai Pop, Teodora Sin, Martin Gahbauer, Oldřich Vojtěch, Jan Mokrý. We are also grateful to Pavel Němčák (National Park Šumava), Tabea Olszewski and Simon König (National Park Bayerischer Wald) for providing the spatial data, Lenart Štaut (Anton Melik Geographical Institute, ZRC SAZU) and Aljaž Jakob (Jovan Hadži Institute of Biology, ZRC SAZU) for their help with data analysis and Živa Malovrh for (grammar) proofreading.
Funding
This study was financed by the Slovenian Research and Innovation Agency (projects InterMuc N1-0163, ExtremePredator J1-50013, Connectivity concept in karst J6-2592) and the European Commission (project LIFE Lynx LIFE16 NAT/SL/000634; EU Program Interreg IV Objective 3 Czech Republic—the Independent State of Bavaria [Grant No. 19]). Š.Č., M.B.V. and M.K. were supported by the Slovenian Research and Innovation Agency (grants no. P6-0101, MR-54767 and P4-0059). T.O. was supported by the Portuguese Foundation for Science and Technology (FCT, Grant No. SFRH/BD/144110/2019).
Author information
Authors and Affiliations
Contributions
Conceptualization, Š.Č., M.B.V. and M.K.; methodology, Š.Č. and T.O.; validation, Š.Č., T.O., E.B., L.B., R.Č., M.H., M.B.V., and M.K.; formal analysis, Š.Č. and T.O.; investigation, Š.Č. and T.O.; data curation, Š.Č., T.O., R.Č., L.B., E.B., M.H., and M.K.; writing—original draft preparation, Š.Č.; writing—review and editing, Š.Č., T.O., E.B., L.B., R.Č., M.H., M.B.V., and M.K.; visualization, Š.Č. and T.O.; supervision, M.B.V. and M.K.; project administration, R.Č., M.B.V. and M.K; funding acquisition, Š.Č., T.O., E.B., L.B., R.Č., M.H., M.B.V. and M.K. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors have no relevant financial or non-financial interests to disclose.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Čonč, Š., Oliveira, T., Belotti, E. et al. Revealing functional responses in habitat selection of rocky features and rugged terrain by Eurasian lynx (Lynx lynx) using LiDAR data. Landsc Ecol 39, 121 (2024). https://doi.org/10.1007/s10980-024-01923-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10980-024-01923-y