
Abstract
Context
The soil-atmosphere carbon exchange is an important component of the carbon cycle; however, dynamics of CO2 fluxes from urban landscapes are particular complicated and poorly understood due their heterogeneity.
Objectives
The objectives of this study were to examine the total and temporal variation in CO2 flux from wooded and turfgrass areas of local parks in different part of town and identify the biophysical characteristics and landscape level factors that contribute to CO2 flux spatiotemporal variance in urban greenspaces.
Methods
We characterized the soil CO2 fluxes, temporal variation, and response to soil temperature from five parks under uniform management of a medium-sized town in southwestern Virginia, USA. We measured site scale characteristics (soil properties, tree cover) as well as the urbanicity of the surrounding land (land cover composition, population).
Results
Soil total nitrogen, soil temperature, and bulk density explained approximately 70% variation in the annual CO2 flux across the five parks. Diurnal, weekly, and seasonal CO2 fluxes were primarily related to changing soil temperature and differed between the turfgrass and wooded areas. Contrary to predictions that increased urbanicity around parks would increase soil temperature and CO2 fluxes, both CO2 and soil temperature of turfgrass were higher at parks located on town edges compared to parks in the center.
Conclusions
In sum, this study indicates that soil nitrogen and compaction, urbanicity, and the resulting site-scale structure of vegetation have a strong influence on temperature dependent biogeochemical processes like CO2 efflux.
Similar content being viewed by others

Explore related subjects
Find the latest articles, discoveries, and news in related topics.Avoid common mistakes on your manuscript.
Introduction
Cities are expanding globally and by 2030 urban land could increase by 1.2 million km2, tripling the urban land area of 2000 (Seto et al. 2012). As cities expand, so do urban greenspaces, such as urban parks, forests, golf courses and lawns, which play a critical role in providing urban ecosystem functions and services (Lubowski et al. 2006). Even though urban greenspaces make up a very small area of the national landscape, locally they play a very important role in urban ecosystems (Belmeziti et al. 2018). Intensive management and disturbance can cause urban greenspaces to be sources of greenhouse gases (Chen et al. 2014a; Ng et al. 2015). Decina et al. (2016) found that during the growing season, soil respiration emitted from urban greenspaces was as much as 72% of fossil fuel emitted CO2. Numerous studies focus on carbon flux dynamics in undeveloped and agricultural ecosystems; however, fewer investigate urban soil carbon dynamics and CO2 flux (Chen et al. 2014b; Kaye et al. 2005). These studies compare carbon dynamics across the urban to rural gradient (urban, agriculture, forested), rather than variation within urban areas and temporal variation. In southwestern part of Seoul, a high-density urban area, the annual mean soil respiration rate within an urban residential area (0.45mg m−2 s−1) was approximately three times larger than that recorded in a nearby urban forest (0.14mg m−2 s−1) (Park et al. 2014). This heightened soil respiration in urban areas are attributed to a combination of municipal and individual landowner management practices. These practices involve the addition of compost or organic fertilizers, which introduce additional carbon into soil and stimulate primary productivity, enhancing soil respiration in urban landscapes (Pouyat et al. 2002; Kaye et al. 2005; Decina et al. 2016). At the local scale, finer-scale heterogeneity in soil CO2 fluxes independent of management differences within the same landscape has not been determined.
Urban landscapes and soils, while more similar to each other than their native ecosystems (Groffman et al. 2014; Trammell et al. 2020), are dynamic, heterogeneous systems that present as a patch-like network of commercial, residential, and undeveloped spaces. In urban settings, the proportion of undeveloped and forested space can vary significantly, encompassing anywhere from < 10% to > 40% of the total land cover (Steele and Wolz 2019). Open spaces covered with turfgrass typically occupying approximately 25% of the land area in most cities (Steele and Wolz 2019). Urban greenspaces in humid climates, such as parks, typically include areas of managed turfgrass, as well as managed and unmanaged trees and shrubs. These spaces range in size from small neighborhood parks space mostly located closer to urban centers, to square kilometers often located on the periphery of towns and cities (Oh and Jeong 2007; Talen 2010). These major structural characteristics of green spaces likely play an important role in the storage and loss of carbon as they experience difference in disturbance, microclimate, and nitrogen deposition depending on where in the urban area they are located and the intensity of use (Pouyat et al. 2010).
Landscape location influences factors like climate and land use distribution, which are important determinants of soil respiratory fluxes. Local temperatures, humidity, substrate types, and soil properties generate variations in the response of soil respiratory fluxes to biological drivers, water inputs, and substrate availability (Davidson et al. 2002; Crum et al. 2016). Local effects are affected by climate, topography, land-use structure, and soil characteristics, collectively shaping fluctuations in soil respiration fluxes (Zhang et al. 2014; Crum et al. 2016). For instance, within Southern California, USA, soil respiration decreased as one transitioned from lawns to agriculture and wildland land use types, spanning coastal, inland, and desert subregions (Crum et al. 2016). This phenomenon was attributed to the interplay of landscape-induced biological drivers and responsiveness of soil respiration to water and carbon substrate availability (Crum et al. 2016). Understanding the influence of landscape heterogeneity on soil CO2 fluxes due to changing landscape position also necessitates understanding ecosystem dynamics, biophysical processes, and landscape structure (Riveros-Iregui and McGlynn 2009). This interdisciplinary approach serves as the cornerstone for investigations into the spatial and temporal variations in soil CO2 fluxes.
Carbon dynamics are influenced by urban soil physical and chemical properties, as well as the carbon pools. These properties can be greatly altered by human management in urban ecosystems (Lorenz and Lal 2015; Herrmann et al. 2018; Trammell et al. 2020). Urban soils, in general, have been shown to have deeper A horizons indicating a sequestration of carbon (Herrmann et al. 2018). Well-managed urban greenspaces can sequester carbon, helping to balance local carbon budgets and mitigate global climate change (Chen et al. 2013a). However, urbanization is also often associated with disturbances such as vegetation clearing, impervious surfaces, and compaction that could reduce ecosystem productivity and soil CO2 flux emission (Seto et al. 2012). Compaction may cause oxygen deficiency belowground and change urban soil aggregate size distribution, thus altering carbon decomposition rates in urban soils (Drew 1983). In addition, urbanization can decrease soil hydraulic conductivity, leading to reduced soil moisture and thus decrease urban soil CO2 efflux (Chen et al. 2014b). On the other hand, soils previously subjected to grading and compaction through land development practices may have a higher potential to enhance their carbon storage through post-development soil management strategies (Chen et al. 2013b).
Urban disturbance can easily disrupt the delicate balance between nitrogen and carbon, consequently having detrimental impacts on the ecosystem. Urbanization affects CO2 fluxes through additions of nitrogen (Hu et al. 2010; Li et al. 2010; Zhang et al. 2014) and introduction of microbial, underground small animals, and plant species (McKinney 2008). Within urban systems, significant quantities of nitrogen are released into the ecosystem through industrial emissions, fuel combustion, and vehicle exhaust and leads to increased nitrogen deposition and substantial anthropogenic nitrogen inputs from activities like fertilizer applications (Hall et al. 2016a; Decina et al. 2020). Soil TN comprises various forms of nitrogen within the soil, and it typically acts as a limiting nutrient for plants, especially plants in urban landscapes (Spargo et al. 2009). Enhanced CO2 flux in response to N may be attributed to the promotion of plant root biomass and respiration (Cleveland and Townsend 2006; Tateno et al. 2004) by stimulating microbial population, activity, microbial biomass nitrogen, and other related characteristics (Tu et al., 2011). While forest management and turfgrass mowing resulting in nitrogen export (Kaye et al. 2005). Excess nitrogen can significantly affect the carbon cycle, leading to increased organic carbon storage within the ecosystem, due to nitrogen deposition (Schindler and Bayley, 1993). Thus disturbance, landcover change, and changes in nitrogen are all important features of the urban environment that influence soil carbon dynamics.
The urban heat island, a prominent characteristic of the urban local and regional climates, may also contribute to increases in CO2 fluxes. Heat affects the plant physiology as well as the respiration of carbon from soil and plants (Hansen et al. 2010). Both modelled results and field measurements predict that autotrophic respiration will increase as temperatures increase (Cramer et al. 2001; Dorrepaal et al. 2009; Frey et al. 2013; Karhu et al. 2014; Davidson 2016). In warm regions, the city might be cooler and more humid than the surrounding desert, CO2 fluxes remain high (Hall et al. 2016b). In the Phoenix, Arizona metropolitan region, Koerner and Klopatek (2002) compared soil CO2 efflux from native desert with human-maintained lands including golf courses, landfills, mesic and xeric landscapes, and agricultural sites. Not surprisingly, their results showed that the desert canopy and interspace showed the lowest rate of CO2 evolution, while mesic landscaping (grass lawns), golf courses and all agricultural land uses resulted in higher rates of CO2 evolution, and landfills exhibited the highest rates of CO2 evolution (Koerner and Klopatek 2002). These results underscore the impact of various land use types on soil CO2 flux, suggesting that urban greenspaces, which contribute to cooling the urban heat island effect, may also yield positive benefits for carbon storage.
Soil CO2 flux is the consequence of interaction between environmental conditions and human activities, making the spatial and temporal variation of urban soil CO2 flux complex in urban landscapes. While the differences between urban and other land uses land have been previously explored (Pouyat et al. 1994), the variability of CO2 fluxes from urban greenspaces remain poorly understood and our ability to understand the carbon dynamics of soil carbon in urban ecosystems is limited. Our goal was to understand heterogeneity attributed to factors such as park size, urbanicity (the degree of surrounding urban development), and soil physical properties, rather effects of management regimes that are addressed elsewhere (Lorenz and Lal 2015; Trammell et al. 2020). For that reason, we chose to study parks that are all under the same management located in different parts of Blacksburg, a medium-sized town. Blacksburg is smaller than most study cities and likely does not experience the same intensity of heat and air pollution of larger cities (Oke 1973; Zhou et al. 2017). However, the ecosystems of smaller cities and towns tend to be understudied compared to larger cities (Forman 2019), thus an important gap in the understanding of urban ecosystems. The objectives of this study were to: (1) examine the CO2 fluxes and temporal variation from wooded and turfgrass areas of local parks within and on the edge of town, and (2) identify the site characteristics and landscape level factors that contribute to CO2 flux spatiotemporal variance in urban greenspaces. We hypothesized that parks within the city would have higher CO2 fluxes due to increased temperatures, but that effect would be moderated by poorer soil conditions, such as compaction. Determining the variability of CO2 fluxes from urban green spaces helps in predicting biogeochemical cycling in urban greenspace.
Methods
Site information
For this study we selected parks within the town limits of Blacksburg, Virginia, USA. Blacksburg is a town of − 44,000 people, 50 km2, and population density of 880/km2 as of 2020 located in the Appalachian Mountain of Southwest Virginia. The land cover composition is dominated by low intensity developed and agricultural land cover (Steele and Wolz 2019). Compared to other cities, it would have an average compactness compared to other US cities based on the assessment by Steele and Wolz (2019). The vegetation is dominated by temperate forests and topography is variable with rolling hills. The climate of the region is temperate humid. On average, there are 214 sunny days per year in Montgomery County. The average air temperature is 11.78 °C, the highest air temperature appears in July (28.33 °C), and the lowest air temperature appears in January (− 5.00 °C). The annual average rainfall is 1034 mm, primarily occurring between March and November, and annual average snowfall is 558.8 mm, with the majority of snowfall taking place between November and March.
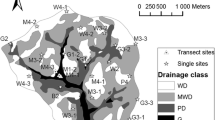
Criteria for selecting parks were: (1) each park should have both managed turfgrass and wooded areas, each with no less than 10 × 10 m of area; (2) the selected parks are distributed across a range of urbanicity as measured by land cover and population to understand how urbanization impact soil CO2 efflux; (3) the selected parks vary in size to capture potential differences in soil CO2 efflux associated with park size; (4) permission for research activities and access to the selected parks; (5) distributed across Blacksburg to minimize spatial bias. With above criteria five parks were selected (37.2106°N to 37.2530°N, 80.4594°W to 80.3925°W, Fig. 1). Within each park, we selected a 10 × 10 m area for both turfgrass and wooded area for measuring CO2 fluxes. The criteria for selecting the 10 × 10 m areas were as follows: (1) included the dominant tree and grass species but representative of vegetation diversity, (2) avoiding buildings, park infrastructure, footpaths, (3) the selected area was located in the center of turfgrass and wooded areas the park to avoid edge effect.
Three of the parks were located closer to the center Blacksburg and surrounded by urban development, primarily residential neighborhoods. These parks included Owen Street Park; Dehart Road Park; Nellies Cave Park (hereafter named U-1, U-2, and U-3). Two parks were located on the edge of town and surrounded by a combination of very low-density residential neighborhoods, forest, and agriculture. These included Tom’s Creek Park and Heritage Community Park (hear after named R-1 and R-2). The age of the parks varied, where U-1, U-2, and R-1 were built during the 1970s, U-3 was developed in the 1980s, while R-2 was developed most recently, in the late 1990s (Table 1). All parks were managed by the local municipality and received the same management (verified by interviews with park managers). All five parks were not irrigated or fertilized, but the turfgrass was regularly mowed once per week during the spring, summer, and fall (Table S1).

Location of selected parks. Three parks were located within the urban land cover (U-1, U-2, U-3) and two were located on the edge of town near agricultural and ex-urban land cover (R-1 and R-2). All parks contained areas of managed turfgrass and remnants of urban woodland (Pictures on Left)
Soil taxonomic information was obtained from the UC Davis “SoilWeb” (https://casoilresource.lawr.ucdavis.edu/gmap/ ). Park soils are representative of the major soil series that dominate the developed and undeveloped landscape (Table 1). The turfgrass vegetation was similar among the five parks, dominated by white clover (Trifolium repens L.), Narrow leaf plantain (Plantago lanceolata L.), Dandelion [Taraxacum officinale (L.)], Bermudas grass (Cynodon spp.), broadleaf plantain (Plantago major L.), perennial ryegrass (Lolium perenne L.), and fescue (Festuca spp.); however, tree species differed from park to park (Table 1).
The surrounding urbanicity of the park was characterized using land cover composition and population. Land cover characteristics of 500 m buffers around each park were calculated using the 2011 National Land Cover Database (NLCD) (Homer et al. 2015). NLCD has a resolution of 30 × 30 m and defines four developed land cover classes by use and the percentage of impervious surface: developed open-areas (NLCD = 21), low-intensity (NLCD = 22), medium-intensity (NLCD = 23), and high-intensity (NLCD = 24) developed. We reclassified these into two developed categories: total urban development (NLCD = 21, 22, 23, 24) and medium and high intensity development (NLCD = 23, 24). The forest land cover categories in the NLCD were aggregated into a single forested category (NLCD = 41, 42, 43). The percent surrounding land cover of the park was calculated by dividing the area of each land cover category by the total area of the buffers. The surrounding population was calculated by summing the population of the 2010 Census Blocks that intersected the 500 m buffer. In addition to characterizing the area around the park, we also calculated the tree canopy cover around each plot within the parks. We identified a 20 m buffer around each plot and then visually identified the tree cover from aerial imagery within each buffer. All landscape characterization was done in ArcGIS Pro 2.3.0 (ESRI, Inc, Redlands, CA, USA).
CO2 flux measurements
The Closed Dynamic Chamber technique (LI-COR 8100, Nebraska, USA) and a flux chamber consisting of an opaque polyvinyl chloride collar (20 cm diameter × 13 cm height, Fig. 1) were used to measure CO2 flux. The Closed Dynamic Chamber technique uses a closed chamber to cover a ground surface area (20 cm diameter circle in this study) for a period (usually between 1 and 3 min, 2 min used in this study). We did not remove near-surface plants before or during measurements, thus the CO2 flux measured in this study included autotrophic respiration from near-surface plants, heterotrophic respiration from soil, and root respiration. The LI-COR was calibrated regularly in the laboratory using calibration tanks with a standard of CO2 concentration (0, 200, and 2000 ppm) at controlled temperature.
The depth of measurement in the soil, determined by depth collars are inserted, is an important factor that affects CO2 flux measurement accuracy. A collar inserted too deep likely severs surface roots, thus weakening autotrophic respiration. On the other hand, a collar insert too shallow may lead to gas leakage problems (Heinemeyer et al. 2011). In this study, collars were inserted into soil (including litter and organic layers) from 2 to 10 cm, with a mean of 5.05 cm in turfgrass (median = 5.00, Figure S1a), with a mean of 5.77 cm in wooded areas (median = 5.70, Figure S1b), very close to the mean of a synthesized studies in forest ecosystem (4.6 cm), but deeper than the mean of nine studies in turfgrass (2.7 cm) (Heinemeyer et al. 2011). The collar insertion depth in this study was within a reasonable range, thus unlikely contributing bias to measurement accuracy.
PVC collars were set up at each park in both urban turfgrass and wooded plots 24 h before CO2 flux sampling. To avoid PVC collars being broken by mowing or tampering, we removed all collars after sampling. Setting up PVC collars disturbs soil structure, and thus influences CO2 flux. To determine how many days the collars should be left to equilibrate and avoid the pulse of CO2 flux associated with severed roots caused by installation, we measured the CO2 flux rates once a day for seven days after PVC collars were inserted in the soil in the Nellies Cave Park (U-3) between August 12th and August 18th 2016. The results indicated that the daily CO2 flux rate did not show a clear trend within one week, but was affected by soil temperature (Ts) and soil moisture (Figure S2). Based on this, we concluded that 24 h was sufficient for CO2 flux equilibration and PVC collars were set up at least 24 h before CO2 flux measurements, same as the minimum waiting time suggested by previous studies (Luo and Zhou 2006).
CO2 flux was measured every two weeks from June 2016 to July 2017. Diurnal variability in CO2 flux is another source of uncertainty that affects measurement accuracy. Diurnal variability affects the ability to capture the daily mean CO2 flux rate, making it difficult to scale up to annual CO2 flux rate based on daily measured CO2 flux rate (Luo and Zhou 2006). CO2 fluxes were measured during a consistent time window (8:00 to 12:00) to limit the effect of diurnal CO2 fluxes variations (Jian et al. 2018). Every week, we employed a randomized sequence for the sites and sub-plots (turfgrass vs. wooded area) we measured, but all the measurements finished during the sampling window. In addition to bi-weekly measurement, we measured CO2 flux once per hour (but once per three hours from 0:00 to 5:00) on August 16th, 2016 (representing summer), October 23rd, 2016 (representing autumn), and March 6th, 2017 (representing spring) to characterize seasonal variation in the diurnal variation and test whether CO2 flux measured from 8:00 to 12:00 accurately captured the daily mean CO2 flux rate for our sites. Note, we did not measure diurnal variation in winter because winter CO2 flux diurnal variation is negligible (Jian et al. 2018). Near the collar we also measured soil water content (SWC) and soil temperature (Ts) at 10 cm depth using Portable Soil Moisture Meter (Model No: MO750, Manufacturer: Extech, Melrose, MA, USA) and Digital Pocket Thermometer (Model No: PDT550, Manufacturer: UEI, Melrose, MA, USA) respectively.
Spatial heterogeneity of CO2 flux can manifest even in landscapes that outwardly appear largely homogeneous (Davidson et al. 2002). Therefore, an important consideration is how many collars are required to capture the CO2 flux spatial variability within a certain accuracy. We used three PVC collars (three subsamples) within the selected 10 m × 10 m study area per experimental unit when we started the experiment at June 18th, 2016. To determine the accuracy of using three collars, we set up 7 collars in the turfgrass, and 8 collars in the wooded area in the U-3 between August 12th and August 18th, 2016. CO2 flux rates were measured between 8:00 and 9:00 am for one week. Based on those measurements, we adapted the method developed by Davidson et al. (2002) to determine the measurement accuracy of three collars according to Eqs. (1),
where n is the number of collars, t is the two-way t statistical value for a given confidence level (95%, 90%, and 80%, respectively) and degrees of freedom (43 measurements for the turfgrass and 47 measurements for the wooded area), s is the standard deviation of the all measurements of turfgrass (all 43 measurements) and wooded area (all 47 measurements), and range is the width of the overall mean ×10%, 20%, or 30%. We found that with three collars in each experimental unit, we have a 95% confidence interval that the measured CO2 flux is within ± 30% of the overall mean, or an 80% confidence interval that the measured CO2 flux follows ± 20% of the overall mean (Table S2), which would reasonably capture the spatial CO2 flux variation in both turfgrass and wooded area.
Soil and plant sampling
Soil samples were taken on 22-March-2017 to measure the soil texture, pH, soil bulk density (BD), soil porosity, soil total nitrogen (TN), total carbon (TC), and carbon to nitrogen ratio (C/N). Soil samples were collected from both turfgrass and wooded areas within each park, with three replicates for each type of area. Sampling was conducted using a 2 cm diameter soil drill, extracting soil samples from a depth of 0–10 cm. Within each 10 m×10 m sampling area, five soil cores were randomly extracted, and these five soil cores were combined to create a composite soil sample. For measuring soil BD, soil augers were used to collect soil samples (n = 3) from both turfgrass and wooded areas within each park. Soil samples from each park were air dried, and then sieved to pass a 2 mm sieve. A 1 g subsample was used for TC, TN, and C/N test using a High Temperature Combustion Analyzer (Elementar, VarioMax). Another 1 g subsample was used to measure the soil texture by the Particle Size Analyzer (Cilas Particle Size 1190, Madison, Wisconsin). Soil BD was measured by drying known volumes of samples to a constant weight at 103 °C. Plant species for turfgrass and wooded area of each park were identified by a local expert. Mowing frequency and grass cut length in each park was identified by interviewing park mowing workers, the park manager, and on-site measurements.
Statistics analysis
A two-way ANOVA was used to test whether CO2 flux from turfgrass or wooded areas were significantly differing from park to park. Simple linear regression (SLR) and non-linear regression (exponential, Eq. 2) were used to test the response of CO2 flux to temperature. SLR was used to test the response of CO2 flux soil moisture. Based on the parameters in Eq. 2, temperature sensitivity of CO2 flux was evaluated by the Q10 function (Eq. 3) in turfgrass and wooded area. In addition, the paired-t test was used to test for significant differences between simulated and measured CO2 flux rates. The relationship between the annual mean CO2 flux from different parks and environmental factors [e.g., soil texture, TC, TN, total urban land cover, high + medium intensity land cover (HMILC)] was fitted by SLR and stepwise approach, p value, R2, AIC and BIC values were selected to evaluate model performance. All statistical analyses were performed in R (version 3.2), and all significant differences were based on p < 0.05.
Where F is a constant value, a indicates the increase rate of soil CO2 flux with soil temperature, and Ts stands for soil temperature at 10 cm.
Results
CO2 flux among parks and vegetation type
We detected significant differences in CO2 fluxes from both turfgrass and wooded areas among the five parks, annual mean CO2 fluxes ranged from 4.89 to 7.73 g C m−2 day−1 in turfgrass, and 4.07 to 5.09 g C m−2 day−1 in wooded areas (Fig. 2). CO2 fluxes from turfgrass were more variable among the parks than fluxes from the wooded areas as quantified by the standard error of mean (Fig. 2) and the number of collars required for soil CO2 flux measurement precision (Table S1). In the turfgrass, CO2 flux from parks closest to the center of town (U-1, U-2, and U-3) were significantly lower than parks (R-1 and R-2) on the edge of town (Fig. 2a). In the wooded areas, CO2 flux was more consistent across parks, where only one (U-3) was significantly lower than the others (Fig. 2b). The annual mean Ts was consistent across parks in both turfgrass and wooded area, with no significant difference was detected among parks.

Mean annual soil CO2 flux, soil temperature (Ts), and soil water content (SWC) of five parks for turfgrass and wooded area over the study period. Vertical bars represent standard error of the mean (n = 75 for turfgrass and n = 78 for wooded area). Different letters above the vertical bars represent significant difference by HSD test. The soil temperature was not differing among sites, thus HSD results for soil temperature were not labeled
We evaluated sixteen environmental factors to explore what causes the variation in soil CO2 flux among the five parks. These factors included soil characteristics [SWC, C/N, TC, TN, pH, porosity, soil texture (sand, silt, and clay), BD, and Ts], as well as landscape characteristics (forested land cover, high + medium intensity land cover, total urban land cover, and population within the 500 m buffer around the park, as well as the park’s area). For turfgrass, the results showed that the annual mean soil CO2 flux rate exhibited variability from one park to another (hereafter referred to as soil CO2 flux spatial variability). This variability can be partially attributed to the surrounding landcover (high + medium intensity land cover and total urban land cover explained 74% and 51% of variability among parks, respectively) and soil (TN, TC, silt, clay, BD, and C/N explained 54%, 47%, 28%, 41%, 38%, and 8% of variability among parks, respectively) properties. In the wooded areas, soil CO2 flux spatial variability can also be partly explained by landscape (forest land cover explained 85% variability among parks) and soil properties (TN, clay, BD, SWC, and TC explained 75%, 38%, 33%, 18%, and 26% variability among parks) (Fig. 3). The result from the stepwise analysis also showed that TN is the most important single parameter explain soil CO2 flux spatial variability among 5 parks, when the model includes TN, Ts, BD, and total urban land cover, the model can explain 91% of soil CO2 flux spatial variability (Table 2; Fig. 3).

The radar chart shows adjusted R2 value of simple linear model between annual soil CO2 flux from five parks and each environmental factor in the turfgrass (a) and wooded area (b). Note that purple petals mean significant relationship (p-value < 0.05), while yellow petals mean no significant relationship (p-value > 0.05). The values in the circles represent adjusted R2 values of linear regression; the sign of (−) and (+) means the variable showed a significant (p < 0.05) positive or negative correlation with annual soil CO2 flux, respectively
Inter-annual variation of CO2 flux
Seasonal variation of CO2 flux from all parks followed a clear pattern which paralleled changes in Ts (Fig. 4a–d). The CO2 flux peaked in summer (June, July, and August) and then gradually fell into its lowest values in winter (December and January). Both CO2 fluxes and Ts tended to be lower in the wooded areas than the turfgrass (Fig. 4a–d). This difference was greatest during the summer months. Surprisingly, Ts and CO2 fluxes tended to be lower in urban parks (i.e., U-1, U2, and U-3) for both wooded and grasses areas compared to Ts and CO2 fluxes from parks located in on the edge of town and surrounded at least in part by rural land uses (i.e., R-1 and R2, Fig. 4e). Temperature differences ranged from − 0.5 to 5.0 °C and tended to be greater for grassed areas than wooded areas (Fig. 4e). We found soil CO2 flux and TS were negatively correlated with tree cover of 20 m buffers around sampling plots (%, Figure S3). Parks within town (U1-U3) had a higher percentage of tree canopy coverage surrounding the plots than the parks on the edge (R1-R2) (Table 1).
Weekly CO2 flux was strongly correlated with weekly Ts for turfgrass and wooded areas of both urban and edge parks (Fig. 5a, b), making it the best single predictor for seasonal CO2 flux at individual sites. The relationships between CO2 flux and Ts were positively related in both turfgrass and wooded areas, characterized by exponential curves. Notably, turfgrass exhibited higher Q10 value (2. 59) compared to wooded areas (2.50). While there were no discernible differences detected between parks for turfgrass (with Q10 values of 2.41, 3.00, 2.22, 3.06, and 2.34, respectively), significant variability was observed among 5 parks for wooded area (with Q10 values of 2.72, 3.32, 2.25, 1.77, and 2.72, respectively) (Fig. 5a, b). SWC did not follow temperature or flux patterns and was a very poor predictor, only accounting for 4% of the variability in CO2 flux in turfgrass and none in wooded areas (Fig. 5c, d).

Comparison of urban vs. edge parks CO2. a Seasonal patterns of soil CO2 efflux (mean ± standard error, n = 15) and soil temperature (mean ± standard error, n = 15) for turfgrass and wooded sites from parks in town (urban) and on the urban edge of town. b Scatter plot shows the CO2 flux difference between edge and urban vs. soil temperature difference

Relationships between soil CO2 flux and air temperature for grasses and woods, respectively (a and b). Relationships between soil CO2 flux and soil water content (SWC, %) for turfgrass and wooded area, respectively (c and d). All depicted regressions are significant at p < 0.05. Note that only significant regression lines (p < 0.05) were shown
Diurnal variation of urban CO2 flux
Diurnal variation in urban CO2 fluxes was stronger in the turfgrass compared to the wooded areas (note that we only measured diurnal pattern of CO2 flux at park U-3, Fig. 6). In the turfgrass areas of the U-3 park, the diurnal variation of CO2 fluxes was strongest during the summer (Fig. 6c), moderate during the fall (Fig. 6e), and minimal during spring (Fig. 6a). Ts in the turfgrass rose and fell in a pattern similar to CO2 flux; however, SWC maintained similar levels during the 24-hour period. Similar to the seasonal patterns, Ts and SWC explained some of the diurnal variation of CO2 flux in turfgrass and wooded area (Fig. 7a). For soil CO2 flux diurnal variation in turfgrass, we detected a positive relationship between CO2 flux and Ts in turfgrass for all three seasons (Fig. 7a). Temperature was more strongly correlated with CO2 fluxes during the spring and fall, than in the summer (Fig. 7a).

Diurnal variation in soil CO2 flux rates, air temperature, and soil water content (SWC) for turfgrass (left panels) and wooded area (right panels) in Spring 2016 (a, b), Summer 2016 (c, d), and Autumn 2017 (e, f), respectively. Vertical bars represent standard error (n = 5 to 8)

Diurnal variation in CO2 fluxes response to soil temperature at 10 cm (a and b), and diurnal variation of CO2 flux response to soil water content change at 10 cm (c and d). In wooded areas, CO2 flux was negatively correlated with soil temperature at spring, but when the data were analyzed on a per-collar basis (small panel in panel b), CO2 flux either positively correlates with soil temperate or showed no correlation. We detected negative relationships between CO2 flux and soil water content in spring for turfgrass, and detected a negative relationship between CO2 flux and soil water content in spring for wooded area. When analyzing the data on a per-collar basis (small panels in panels c and d), either no relationship or a positive relationship was detected. Note that we only plotted the regression line when the linear model is significant (p < 0.05)
In wooded areas, the diurnal variations of CO2 were positively related with Ts at summer and autumn, but correlated negatively with Ts at spring (solid dots and line in Fig. 7b). We grouped the CO2 flux data in spring by collars (small panel in Fig. 7b), and found that the Ts measured from one of the collars spanned a relatively large range, and CO2 flux measured at each collar positively related with Ts change (we only plotted the regression line when the linear model is significant, p < 0.05, Fig. 7b).
In turfgrass, we detected negative relationships between CO2 flux and SWC in spring, but positive relationship was detected in autumn and summer (Fig. 7c). In wooded area, we detected negative relationships between CO2 flux and SWC in spring and autumn (Fig. 7d). However, upon analyzing the data on a per-collar basis, either no relationship, or a positive relationship, was detected (small panels in Fig. 7c and d). When the data were analyzed on a per-collar basis, the results showed that even within 10 m×10 m area, there is significant sub-site heterogeneity in the CO2 flux temporal dynamics. These results indicated that when CO2 flux, Ts, and SWC diurnal fluctuation is very small, the collar to collar variation is bigger than the diurnal fluctuation, and thus data must be carefully analyzed.
Discussion
In this study, we found that the CO2 flux of parks in Blacksburg, VA was temporally variable, differed with turfgrass and wooded areas, and influenced by their relative location in town. CO2 fluxes from Blacksburg are similar to other measurements in other larger urban areas. Our average annual mean CO2 flux from wooded areas in the five parks (4.72 g C day−1 m−2) was similar to CO2 flux rates in a temperate urban forest in Beijing, China (Wu et al. 2015) and a mixed forest in an urban park in South Korea (Bae and Ryu 2017). However, our measurements were higher than the CO2 flux rate (2.62 g C day−1 m−2) from an urban forest in Boston, USA (Decina et al. 2016) and an urban forest in Hefei, China (2.60 g C day−1 m−2) (Tao et al. 2016). We observed lower CO2 flux rates compared to urban lawns in drier climates, including Fort Collins, CO (7.61 g C day−1 m−2) and Phoenix, AZ (8.18 g C day−1 m−2) (Koerner and Klopatek 2002) and an urban tropical grassland in Singapore (8.19 g C day−1 m−2) (Ng et al. 2015). Compared to the wooded areas, we observed higher CO2 fluxes from turfgrass in the parks because we did not remove near-surface plants during gas sampling. Thus, CO2 fluxes from turfgrass included both soil respiration and respiration from the grass leaves and roots, i.e., total ecosystem respiration. In contrast, measurements from wooded areas were taken below the canopy and did not include leaf respiration within collars; therefore, CO2 fluxes were primarily root and soil respiration, which is only a part of ecosystem respiration (Barba et al. 2018).
Variation among parks
Despite the uniformity of management provided by the local government there were differences in annual fluxes among parks, opposite our initial predictions. Parks on edges of urban development (R-1 and R-2) had larger annual CO2 efflux compared with parks surrounded by primarily urban land covers (U-1, U-2, and U-3). Soil from edge parks tended to have more soil carbon and nitrogen, and lower bulk densities (Figure S4), which explained some of the pattern in CO2 efflux. One of the edge parks (R-2) was the most recently developed, approximately 25 years before this study; however, both edge parks show similar observations regardless of differences in age. Increased compaction in urban parks compared to edge parks may be limiting the soil microbial community and carbon sequestration. A study observed similar patterns where the annual soil CO2 flux at the primary rural site was 79% higher than the urban site, and 49% higher than the suburban site (Koerner and Klopatek 2010). The authors suggested that lack of microhabitats in the urban soils may be responsible for the lower respiration rates observed at the urban sites (Koerner and Klopatek 2010). In our study, the edge parks may also have lower concentrations of trace metals and other pollutants, not measured here, that are impacting the soil microbiome.
Soil TN emerged as an important factor in the variability of the annual mean soil CO2 flux rate across the five parks included in this study (Fig. 3). Urbanization, via fertilization and combustion, can be a large source in N inputs causing N and C in soil to converge across US cities (Trammell et al. 2020). It is noteworthy that soil TN explained 54% and 75% of the variability of annual soil CO2 flux among parks for turfgrass and wooded areas, respectively. Despite the likely increased in N inputs within cities from N pollution, the two parks situated in the surrounding agricultural and ex-urban land cover (R-1 and R-2) displayed higher TN levels (with TN content of 0.277% for R-1 and 0.224% for R-2, respectively, Figure S4). Three parks located within the urban land cover (U-1, U-2, U-3) have TN level of 0.275%, 0.209%, and 0.146%, respectively. This disparity underscores the role of soil TN as a driving force behind the observed spatial variability in soil CO2 flux. It is possible that legacy effects from historical agriculture could account for the increased soil carbon and nitrogen content for parks on the edge of town (Lewis et al. 2014). The entire town was once farmed, but the farms that became urban parks would have been abandoned long before the edge parks and thus may still contain legacy nitrogen and carbon. Soil TN and its effects on carbon cycling also exhibits variability over the course of a year (Wang et al. 2018). Unfortunately, in our study, we did not collect soil samples at various times throughout the year. This limitation may introduce some uncertainty into the results, therefore, it is advisable for future research to consider the seasonal variability of TN when investigating similar phenomena.
Ts showed notable variability among parks in both turfgrass and wooded area, which was correlated with the percentage of surrounding tree coverage (Figure S3). Parks within urban areas had greater tree coverage surrounding our sampling plots tend to have lower soil CO2 flux and TS. Tree canopy coverage has been shown to have a strong effect on the air and surface temperature in urban areas via evapotranspiration and shading which alleviates the heat island effect (Lin and Lin 2010). Wang et al. (2023) found that land surface temperature of green spaces was highly dependent on tree cover and was not mitigated by grass in the city of Leeds, UK. Soil temperature in forests has been shown to depend on the leaf area index (Paul et al. 2004). While park area itself was not important, it may be indirectly impacting CO2 fluxes via the tree cover. Parks within town were considerably smaller and the tree canopy covered a greater percentage of the park. Larger edge parks also had trees lining the park, but there was considerably more open area and space between. Local shading from vegetation is an important tool to influence the microclimate and offset surrounding urbanicity, which may extend to biogeochemical processes in the soil under nearby turfgrass as well.
Seasonal and daily CO2 efflux
CO2 fluxes from urban greenspaces in this study were strongly correlated with variations in temperature across seasons and days, which supports many studies on temperate ecosystems (Davidson et al. 2006) and urban ecosystems (Zhou et al. 2012). We found, however, the correlation of CO2 flux with soil moisture was relatively poor (Fig. 5). A possible reason is due to the plentiful rainfall and good drainage, so the soils seldom underwent long-term saturation or frequent prolonged drought (SWC maintain at 5–10% all year around, Fig. 5c and d), thus seasonal variation of soil moisture was relatively small and may not limit activities of microbe in most months. In addition, during wet or normal years in temperate regions, root respiration from trees and grasses may not respond to seasonal variation of soil moisture because the root activity is seldom limited by soil water availability (Yang et al. 2007). Generally, our results indicated that the major driver of CO2 flux seasonal variation was Ts rather than SWC (Fig. 5) in both urban turfgrass and urban wooded areas in our chosen parks.
In wooded areas, CO2 flux diurnal variation was positively related with Ts in summer and autumn, but negatively correlated with Ts at spring (Fig. 7b). We did not detect any significant relationship between Ts and SWC at diurnal scales (Table S2), thus it is unlikely that Ts and SWC interaction cause negative relationships between CO2 flux and Ts in spring. We grouped CO2 flux measurements in spring by collars (different colors in the small panel in the Fig. 7b), and found that within each collar, the soil respiration either positively related with Ts change or show no relationship to Ts (small panel in the Fig. 7b). Results indicated that in spring, when CO2 flux, Ts, and SWC diurnal fluctuation is very small, collar-to-collar variation is larger than the diurnal fluctuation. A negative correlation between soil CO2 flux and soil temperature was detected (Fig. 7b–d) between CO2 flux and environmental factors is an unusual relationship and require additional experimentation to ascertain the underlying factors contributing to these unexpected relationships.
Implications for urban ecosystems
The results of this study indicate that when management is constant there is still sufficient heterogeneity across the landscape of a medium-sized town to affect CO2 efflux. CO2 in these urban greenspaces is a temperature dependent flux, which drives variation over seasons and days. Landscape level factors surrounding the parks did not change the relationship between temperature and respiration rates. Rather the surrounding landscape and site level factors affected temperatures directly. Contrary to expectations, urban heat island effects associated with urbanicity did not increase CO2 fluxes. Soil temperatures were lower in parks closer to the center of town, likely due to the shade provided by local tree canopy shade. These results also suggest increased N deposition associated with urbanicity is also less influential than other factors, such as legacy stores of nutrients and soil compaction. This may be good news for town ecosystems, as it is easier for municipalities to manage soil compaction and increase shade trees in greenspaces than it would be to reduce the larger urban heat and N pollution. Similar studies in larger cities would be needed to determine if there is a threshold population size or traffic level at which N pollution signatures may be observed.
Conclusions
Urban carbon fluxes play crucial role for informing sustainable urban planning in climate-friendly cities, but to date, there is little information about the differences in these fluxes across urban landscapes and independent of management effects. Soil temperature and soil moisture were the main environmental factors controlling CO2 flux diurnal, weekly and seasonal variation. Spatially, we observed a significant difference in CO2 flux among the five parks in this study, where interior parks had lower soil temperature and CO2 fluxes. Soil properties like bulk density, total carbon and total nitrogen contribute to variation in CO2 fluxes from different parks even when management is uniform. These results contribute to our understanding of carbon cycles in heterogeneous urban landscapes and indicate local site scale management could have a strong effect on CO2 efflux in for towns ecosystems.
Data availability
All data used in this analysis are available on request from the corresponding author.
Code availability
All code to reproduce all the results in this analysis are available on request from the corresponding author.
References
Bae J, Ryu Y (2017) Spatial and temporal variations in soil respiration among different land cover types under wet and dry years in an urban park. Landsc Urban Plan 167:378–385
Barba J, Cueva A, Bahn M, Barron-Gafford G, Bond-Lamberty B, Hanson P, ... & Vargas R (2018) Comparing ecosystem and soil respiration: Review and key challenges of tower-based and soil measurements. Agric For Meteorol 249: 434–443
Belmeziti A, Cherqui F, Kaufmann B (2018) Improving the multi-functionality of urban green spaces: relations between components of green spaces and urban services. Sustain Cities Soc 43:1–10
Chen W, Jia X, Zha T et al (2013a) Soil respiration in a mixed urban forest in China in relation to soil temperature and water content. Eur J Soil Biol 54:63–68
Chen Y, Day SD, Wick AF et al (2013b) Changes in soil carbon pools and microbial biomass from urban land development and subsequent post-development soil rehabilitation. Soil Biol Biochem 66:38–44
Chen Y, Day SD, Shrestha RK et al (2014a) Influence of urban land development and soil rehabilitation on soil–atmosphere greenhouse gas fluxes. Geoderma 226–227:348–353
Chen Y, Day SD, Wick AF, McGuire KJ (2014b) Influence of urban land development and subsequent soil rehabilitation on soil aggregates, carbon, and hydraulic conductivity. Sci Total Environ 494–495:329–336
Cleveland CC, Townsend AR (2006) Nutrient additions to a tropical rain forest drive substantial soil carbon dioxide losses to the atmosphere. Proc Natl Acad Sci 103(27):10316–10321
Cramer W, Bondeau A, Woodward FI et al (2001) Global response of terrestrial ecosystem structure and function to CO2 and climate change: results from six dynamic global vegetation models. Glob Chang Biol 7:357–373
Crum SM, Liang LL, Jenerette GD (2016) Landscape position influences soil respiration variability and sensitivity to physiological drivers in mixed-use lands of Southern California, USA. J Geophys Res: Biogeosci 121(10):2530–2543
Davidson EA (2016) Biogeochemistry: projections of the soil-carbon deficit. Nature 540:47–48
Davidson EA, Savage K, Verchot LV, Navarro R (2002) Minimising artifacts and biases in 20 chamber-based measurements of soil respiration. Agric Ecosyst Environ Meteorol 113:21–37
Davidson EA, Richardson AD, Savage KE, Hollinger DY (2006) A distinct seasonal pattern of the ratio of soil respiration to total ecosystem respiration in a spruce-dominated forest. Glob Chang Biol 12:230–239
Decina SM, Hutyra LR, Gately CK et al (2016) Soil respiration contributes substantially to urban carbon fluxes in the greater Boston area. Environ Pollut 212:433–439
Decina SM, Hutyra LR, Templer PH (2020) Hotspots of nitrogen deposition in the world’s urban areas: a global data synthesis. Front Ecol Environ 18(2):92–100
Dorrepaal E, Toet S, van Logtestijn RSP et al (2009) Carbon respiration from subsurface peat accelerated by climate warming in the subarctic. Nature 460:616–619
Drew MC (1983) Plant injury and adaptation tot oxygen deficiency in the root environment: a review. Plant Soil 75(2):179–199
Forman RT (2019) Towns, ecology, and the land. Cambridge University Press, Cambridge
Frey SD, Lee J, Melillo JM, Six J (2013) The temperature response of soil microbial efficiency and its feedback to climate. Nat Clim Chang 3:395–398
Groffman PM, Cavender-Bares J, Bettez ND et al (2014) Ecological homogenization of urban USA. Front Ecol Environ 12:74–81
Hall SJ, Ogata EM, Weintraub SR et al (2016a) Convergence in nitrogen deposition and cryptic isotopic variation across urban and agricultural valleys in Northern Utah. J Geophys Res: Biogeosci 121(9):2340–2355
Hall SJ, Learned J, Ruddell B et al (2016b) Convergence of microclimate in residential landscapes across diverse cities in the United States. Landsc Ecol 31:101–117
Hansen J, Ruedy R, Sato M, Lo K (2010) Global surface temperature change. Rev Geophys 48:RG4004
Heinemeyer A, Di Bene C, Lloyd AR et al (2011) Soil respiration: implications of the plant-soil continuum and respiration chamber collar-insertion depth on measurement and modelling of soil CO2 efflux rates in three ecosystems. Eur J Soil Sci 62:82–94
Herrmann DL, Schifman LA, Shuster WD (2018) Widespread loss of intermediate soil horizons in urban landscapes. Proc Natl Acad Sci 115:6751–6755
Homer C, Dewitz J, Yang L et al (2015) Completion of the 2011 National Land Cover Database for the conterminous United States–representing a decade of land cover change information. Photogramm Eng Remote Sens 81:345–354
Hu Z, Li H, Yang Y et al (2010) Effects of simulated nitrogen deposition on soil respiration in northern subtropical deciduous broad-leaved forest. Environ Sci 31:1726–1732
Jian J, Steele MK, Day SD et al (2018) Measurement strategies to account for soil respiration temporal heterogeneity across diverse regions. Soil Biol Biochem 125:167–177
Karhu K, Auffret MD, Dungait JAJ et al (2014) Temperature sensitivity of soil respiration rates enhanced by microbial community response. Nature 513:81–84
Kaye JP, McCulley RL, Burke IC (2005) Carbon fluxes, nitrogen cycling, and soil microbial communities in adjacent urban, native and agricultural ecosystems. Glob Chang Biol 11:575–587
Koerner B, Klopatek J (2002) Anthropogenic and natural CO2 emission sources in an arid urban environment. Environ Pollut 116:45–51
Koerner BA, Klopatek JM (2010) Carbon fluxes and nitrogen availability along an urban-rural gradient in a desert landscape. Urban Ecosyst 13:1–21
Lewis DB, Kaye JP, Kinzig AP (2014) Legacies of agriculture and urbanization in labile and stable organic carbon and nitrogen in Sonoran Desert soils. Ecosphere 5(5):1–18
Li R, Tu L, Hu T et al (2010) Effects of simulated nitrogen deposition on soil respiration in a Neosinocalamus Affinis plantation in Rainy Area of West China. Chin J Appl Ecol 21:1649–1655
Lin BS, Lin YJ (2010) Cooling effect of shade trees with different characteristics in a subtropical urban park. HortSci 45(1):83–86
Lorenz K, Lal R (2015) Managing soil carbon stocks to enhance the resilience of urban ecosystems. Carbon Manag 6(1–2):35–50
Lubowski RN, Vesterby M, Bucholtz S et al (2006) Major uses of land in the United States, 2002. United States Department of Agriculture, Economic Research Service, Washington, D.C., USA
Luo Y, Zhou X (2006) Soil respiration and the environment. Elsvevier, Amsterdam
McKinney ML (2008) Effects of urbanization on species richness: a review of plants and animals. Urban Ecosyst 11:161–176
Ng BJ, Hutyra LR, Nguyen H et al (2015) Carbon fluxes from an urban tropical grassland. Environ Pollut 203:227–234
Oh K, Jeong S (2007) Assessing the spatial distribution of urban parks using GIS. Landsc Urban Plan 82:25–32
Oke TR (1967) City size and the urban heat island. Atmos Environ 7(8):769–779
Park MS, Joo SJ, Park SU (2014) Carbon dioxide concentration and flux in an urban residential area in Seoul, Korea. Adv Atmos Sci 31:1101–1112
Paul KI, Polglase PJ, Smethurst PJ et al (2004) Soil temperature under forests: a simple model for predicting soil temperature under a range of forest types. Agric for Meteorol 121(3–4):167–182
Pouyat RV, Parmelee RW, Carreiro MM (1994) Environmental effects of forest soil-invertebrate and fungal densities in oak stands along an urban-rural land use gradient. Pedobiologia 38(5):385–399
Pouyat R, Groffman P, Yesilonis I, Hernandez L (2002) Soil carbon pools and fluxes in urban ecosystems. Environ Pollut 116:S107–S118
Pouyat RV et al (2010) Chemical, physical, and biological characteristics of urban soils. Urban Ecosyst Ecol 55:119–152
Riveros-Iregui DA, McGlynn BL (2009) Landscape structure control on soil CO2 efflux variability in complex terrain: scaling from point observations to watershed scale fluxes. J Geophys Research: Biogeosciences, 114(G2).
Schindler DW, Bayley SE (1993) The biosphere as an increasing sink for atmospheric carbon: estimates from increased nitrogen depostion. Glob Biogeochem Cycles 7(4):717–733
Seto KC, Guneralp B, Hutyra LR (2012) Global forecasts of urban expansion to 2030 and direct impacts on biodiversity and carbon pools. Proc Natl Acad Sci 109:16083–16088
Spargo JT, Alley MM, Thomason WE, Nagle SM (2009) Illinois soil nitrogen test for prediction of fertilizer nitrogen needs of corn in Virginia. Soil Sci Soc Am J 73(2):434–442
Steele MK, Wolz H (2019) Heterogeneity in the land cover composition and configuration of US cities: implications for ecosystem services. Landsc Ecol 34:1247–1261
Talen E (2010) The spatial logic of parks. J Urban Design 15(4):473–491
Tao X, Cui J, Dai Y et al (2016) Soil respiration responses to soil physiochemical properties in urban different green-lands: a case study in Hefei, China. Int Soil Water Conserv Res 4:224–229
Tateno R, Hishi T, Takeda H (2004) Above-and belowground biomass and net primary production in a cool-temperate deciduous forest in relation to topographical changes in soil nitrogen. For Ecol Manag 193(3):297–306
Tu LH, Hu TX, Zhang J, Li RH, Dai HZ, Luo SH (2011) Short-term simulated nitrogen deposition increases carbon sequestration in a Pleioblastus amarus plantation. Plant and soil, 340:383–396
Trammell TLE, Pataki DE, Pouyat RV et al (2020) Urban soil carbon and nitrogen converge at a continental scale. Ecol Monogr 90:1–13
Wang C, Liu D, Bai E (2018) Decreasing soil microbial diversity is associated with decreasing microbial biomass under nitrogen addition. Soil Biol Biochem 120:126–133
Wang X, Scott CE, Dallimer M (2023) High summer land surface temperatures in a temperate city are mitigated by tree canopy cover. Urban Clim 51:101606
Wu X, Yuan J, Ma S et al (2015) Seasonal spatial pattern of soil respiration in a temperate urban forest in Beijing. Urban for Urban Green 14:1122–1130
Yang YS, Chen GS, Guo JF et al (2007) Soil respiration and carbon balance in a subtropical native forest and two managed plantations. Plant Ecol 193:71–84
Zhang C, Niu D, Hall SJ et al (2014) Effects of simulated nitrogen deposition on soil respiration components and their temperature sensitivities in a semiarid grassland. Soil Biol Biochem 75:113–123
Zhou X, Wang X, Tong L et al (2012) Soil warming effect on net ecosystem exchange of carbon dioxide during the transition from winter carbon source to spring carbon sink in a temperate urban lawn. J Environ Sci (China) 24:2104–2112
Zhou B, Rybski D, Kropp JP (2017) The role of city size and urban form in the surface urban heat island. Sci Rep 7(1):4791
Funding
This research was supported by the by the Hatch Grant from Virginia Tech with award number 160060 and the Strategic Priority Research Program of Chinese Academy of Sciences, with grant number XDA20040202.
Author information
Authors and Affiliations
Contributions
J.J. and M.S. conceived the study and designed the experiment, J.J. performed the field work, J.J. and M.S. analyzed the data and wrote the manuscript.
Corresponding author
Ethics declarations
Conflicts of interest
We declare no conflicts of interest.
Ethical approval
Not applicable.
Consent to participate
Not applicable.
Consent for publication
Not applicable.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Jian, J., Steele, M.K. Heterogeneity of soil CO2 efflux from local parks across an urban landscape. Landsc Ecol 39, 16 (2024). https://doi.org/10.1007/s10980-024-01812-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10980-024-01812-4