
Abstract
Context
The forestry industry provides important goods, services and economic benefits, but timber harvest can adversely impact ecosystem services, including wildlife habitat. Timber harvest planning can integrate wildlife habitat quality through multi-objective optimization for timber harvest and wildlife habitat suitability.
Objectives
Our objective was to develop a method to find optimal solutions for timber harvest and wildlife habitat suitability individually and concurrently, then apply the method to Roosevelt elk (Cervus elaphus roosevelti) on Afognak Island, Alaska.
Methods
We developed three seasonal habitat suitability models using elk locations and landscape variables including historical timber harvest on Afognak Island, Alaska. We used threshold-accepting optimization over a 50-year planning horizon to maximize timber harvest yield and habitat suitability in each season, then used multi-objective goal-deviation optimization to simultaneously maximize timber harvest volume and seasonal habitat suitability.
Results
The optimal solution for timber yield decreased seasonal average habitat suitability by 5.7%. Elk habitat suitability and corresponding optimal solutions varied seasonally; elk generally selected open landcovers and early- to mid-successional timber stands over late-successional and mature stands. Therefore, in the optimal solutions, stands were harvested before they reached maximum volume and few stands were harvested in early planning periods, resulting in a seasonal average loss of 17.5% yield. Multi-objective optimization decreased seasonal average suitability by 3.9% and yield by 1.4% compared to single-objective optimization.
Conclusions
Our multi-objective optimization approach that incorporates data-driven habitat suitability models using open-source software can enable managers to achieve desired quantity and quality of wildlife habitat while providing for resource extraction.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Forests represent 30% of global land area (40 million km2), and over 50% of forests are managed for timber harvest (FAO 2016). Timber harvest provides essential resources such as fuel, pulp and paper, and construction materials (Pepke 2010; FAO 2016). The forestry sector delivers about 117 billion United States dollars in global domestic product and employs over 12 million people (FAO 2016). Though timber has great economic value, timber harvest can adversely affect ecosystems (Nelson et al. 2009; Lawler et al. 2014). Forests provide carbon uptake and storage services which may be reduced when timber is harvested (Díaz-Balteiro and Romero 2003; Lawler et al. 2014). Unharvested forests also increase groundwater retention, reducing runoff and increasing water quality (Nelson et al. 2009).
Timber harvest also effects biodiversity through its impacts on wildlife habitat quality (Steventon et al. 1998; Fisher and Wilkinson 2005). Many wildlife species experience positive effects from early successional habitat following timber harvest (Fisher and Wilkinson 2005; Kellner et al. 2019). During post-harvest forest regeneration, abundant herbaceous vegetation can increase herbivore food availability, and dense woody debris supports moist microclimates and provides food for small mammals and birds (Fisher and Wilkinson 2005). Additionally, later successional stages have denser intermediate cover, which can provide shelter from extreme weather conditions and may allow prey species to avoid predators (Fisher and Wilkinson 2005; Frair et al. 2005).
However, timber harvest can also have negative effects on wildlife, as some species require large, intact mature forests with little to no disturbance (Rettie and Messier 2000; Fisher and Wilkinson 2005; Forsman et al. 2005). Mature forests can provide unique forage resources (Rettie and Messier 2000; Kellner et al. 2019), as well as roost and den sites that are often destroyed during timber harvest (Steventon et al. 1998; Fisher and Wilkinson 2005). Mature forests also may provide thermal or hiding cover for large mammals (Skovlin et al. 1989; Barbknecht et al. 2011; Rearden et al. 2011). Additionally, spatial configuration of timber harvest affects habitat suitability, as timber harvest creates edge effects that have contrasting effects on wildlife (Fisher and Wilkinson 2005; Frair et al. 2005). For example, elk benefit from open habitat with ample forage that is adjacent to hiding and thermal cover (Skovlin et al. 1989; Rumble and Gamo 2011), while birds use post-harvest shrub habitat adjacent to mature forests for food resources and cover (Kellner et al. 2019). Therefore, not only are the effects of timber harvest on habitat suitability substantial, but they are also spatially and temporally dynamic. Timber harvest planning should not only consider economic returns, but also how forest regeneration and spatial and temporal landscape heterogeneity impacts wildlife.
One method of timber harvest planning accounts for wildlife habitat quality through habitat suitability models, which determine the importance of habitat variables by comparing habitat at locations where a species or individual is observed to available habitat (Larson et al. 2004; Thomas and Taylor 2006). These models may use animal location data to provide specific estimates of habitat suitability across timber harvest planning scenarios (Larson et al. 2004; Kellner et al. 2019), which enables timber planning optimization specifically for habitat suitability. Consequentially, an optimal solution can be achieved that balances wildlife habitat and timber harvest goals (Bettinger et al. 1999; Yemshanov et al. 2020). However, multi-objective optimization may be difficult in wildlife or timber harvest management scenarios due to costly software, insufficient computing power, or coding requirements. We present a method for multi-objective optimization incorporating habitat suitability using open-source software and data often available to managers, and apply it to Rooselevelt elk (Cervus elaphus roosevelti) on Afognak Island, Alaska.
Roosevelt elk are a good model species for multi-objective optimization for timber harvest and habitat suitability because their habitat needs are directly impacted by timber harvest (Skovlin et al. 1989; Bettinger et al. 1999). Elk use open areas, such as meadows or recently harvested timber stands, for forage, especially during summer or parturition (Irwin and Peek 1983; Barbknecht et al. 2011). However, elk also require cover, such as younger, denser forests, or alternatively, mature forests, to shelter from severe weather and predators (Skovlin et al. 1989; Unsworth et al. 1998; Rumble and Gamo 2011). Therefore, elk habitat needs may conflict with timber harvest goals because elk may require high quality foraging areas as well as mature forests (Wisdom et al. 1986; Skovlin et al. 1989).
We created a timber harvest plan that balanced seasonal elk habitat suitability with timber yield. First, we used elk location data to develop three seasonal elk habitat suitability models which incorporated landscape variables including temporally dynamic landcover categories. We then used coefficients from the habitat suitability models and timber harvest volume estimates to create optimization models in each season using a threshold-accepting heuristic framework across a 50-year planning horizon separately for timber harvest yield and habitat suitability. Finally, we used multi-objective goal-deviation optimization to balance timber harvest yield with temporally dynamic elk habitat suitability.
Methods
Study area
Afognak Island (1,809 km2; 58.3279°N, 152.6415°W) is 5 km north of Kodiak Island in the Kodiak Archipelago, Alaska, USA (Fig. 1). Afognak Island is primarily owned by Native corporations (64%), followed by state (27%) and federal (9%) ownership. It contains gradual sloping mountains from 300 to 800 m above sea level. The archipelago has a subpolar oceanic climate with average annual high and low temperatures of 8.0 and 2.1 °C, respectively, and average annual rainfall and snowfall are 174 and 172 cm, respectively (Menne et al. 2012).

Afognak Island, Alaska. World and state map outlines from NASA open data portal, Kodiak and Afognak Island outlines from Kodiak Island Borough maps and data center
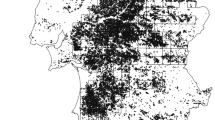
The eastern portion of Afognak Island is dominated by Sitka spruce (Picea sitchensis) which occurs to 365 m in elevation with an understory containing blueberry (Vaccinium ovalifolium), devil’s club (Oplopanax horridus), salmonberry (Rubus spectabilis), and elderberry (Sambucus racemose; Troyer 1960). Other portions of the island are dominated by alder (Alnus fruiticosa) and willow (Salix spp.) interspersed with open herbaceous areas containing forbs such as bluejoint (Calamagrostis canadensis) and fireweed (Epilobium angustifolium) (Troyer 1960). Small areas of Afognak Island were clearcut intermittently during 1930–1965 (Harris 1972), with extensive commercial clearcut logging on south-central Afognak Island since 1976 (Fig. 2). Sitka spruce productivity on Afognak Island is relatively low compared to productivity in most of its range, with a site index of about 24 m at 100 years old (Nesheim, unpublished report; Hansen 2018).

Timber harvest stands and roads, Afognak Island, Alaska, 1943–2022
In 1929, eight Roosevelt elk were introduced to Afognak Island by the Alaska Game Commission to establish a harvestable population (Troyer 1960). The population increased to over 1000 animals by 1965 and has since ranged from 700 to 1200 individuals (Batchelor 1965; Schooler et al. 2022). The elk population on Afognak Island has considerable economic, cultural, and recreational value (Troyer 1960; Schooler et al. 2022). Elk can be legally harvested with permits issued by the Alaska Department of Fish and Game (ADFG) (Alaska Department of Fish and Game 2022).
Elk capture and handling
We captured 70 elk (41 female, 29 male; Online Resource 1 Table S1) during June–August 2014–2020 on Afognak Island using standard aerial darting techniques (Finnegan et al. 2021). We fit elk with Global Positioning System (GPS) radiocollars (model TGW-4677; Telonics, Inc, Mesa, Arizona, USA or model Vertex Plus-4; Vectronic, Berlin, Germany) programmed to attempt a location every 60 min; the collar was programmed to release 21–24 months after deployment to ensure sufficient battery life for retrieval (Finnegan et al. 2021). We attached leather links designed to degrade after 20–24 months as a secondary drop-off mechanism (Finnegan et al. 2021). Animal capture and handling procedures were approved by the State University of New York College of Environmental Science and Forestry (SUNY ESF) (IACUC; protocol 180503) and Alaska Department of Fish and Game (ADFG; IACUC; protocol 0030-2018-37) Institutional Animal Care and Use committees.
Data sources
We compiled 79 years (1943–2022) of timber harvest polygon shapefiles from the United States Forest Service (Harris 1972), Afognak Native Corporation, Koniag Inc., Koncor, Natives of Kodiak Native Corporation, and Ouzinkie Native Corporation (“Native corporations”). We verified, corrected, and augmented these data using historical reports, Google Earth, ESRI ArcGIS Pro imagery, aerial photography, and Landsat satellite imagery (Fig. 2; Online Resource 2; Harris 1972; U.S. Geological Survey 1988, 2006, 2022; Esri et al. 2022; Google Earth 2022). Data included attributes for year of harvest, treatment (e.g., replanted or not replanted), and land ownership. We separated timber harvest data into management units (“stands”) based on these attributes for consistency with original harvest data (Native corporations, unpublished data). Our study area contained 1,272 timber harvest stands ranging from 0.13 to 525 hectares.
Most timber stands harvested before 1990 were left to naturally regenerate (Native corporations, unpublished data). During 2000–2002 61 timber stands harvested during 1980, 1989, and 1995–2002 were replanted with Afognak Island strain Sitka spruce seedlings (Native corporations, unpublished data). After 2002, Sitka spruce seedlings of a more rapidly-growing genetic strain from southeast Alaska (“Juneau strain”) were replanted in 265 stands (“Juneau stands”), with number of replanted stands increasing since 2002. All stands harvested since 2012 have been replanted with Juneau strain Sitka spruce (Native corporations, unpublished data).
Stands replanted with Afognak strain grew at similar rates as naturally-regenerating stands, but due to recolonization time, were about 15 years more mature (Online Resources 2–4; Nesheim, unpublished report; Alden 1998). Therefore, we added 15 years to Afognak planted stand age and grouped them with naturally-regenerating stands (henceforth referred to collectively as naturally-regenerating stands). Because Juneau strain Sitka spruce grows more rapidly than the native Afognak Sitka spruce (Nesheim, unpublished report; Koniag Inc., unpublished data), we used separate age classifications for Juneau and naturally-regenerating stands (Table 1). Planted Juneau stands are economically profitable to harvest after 50 years of age and naturally regenerating stands are profitable after 70 years of age (Koniag Inc., personal communication).
We characterized forest stands based on their habitat value for elk using year of harvest or year of replanting, incorporating differences in growth for planted and unplanted stands (Table 1; Online Resource 2). Using information from Native corporation land managers, peer-reviewed literature, Normalized Difference in Vegetation Index (NDVI) in 2015 and 2021, and canopy cover in 2015 (Online Resources 2–4; Sexton et al. 2013; Garroutte et al. 2016; U.S. Geological Survey and Google 2021), we combined timber harvest stands into seven categories: no cover, no forage; no cover, low forage; no cover, medium forage; medium cover, medium forage; medium cover, low forage; high cover, no forage; and forest (Table 1; Jenkins and Starkey 1996; Visscher and Merrill 2009). For interpretation, we also translated these categories into successional stages: recently harvested; early-, mid-, and late-succession; and mature forest. We calculated cubic meters of timber per hectare for 5-year age classes for naturally-regenerating and Juneau stands using a height-growth model and published literature (Koniag Inc. unpublished data; Hansen 2018). Stands had the greatest volume per hectare at ages 135–145 for naturally-regenerating and 70–80 years for Juneau stands (Online Resource 5).
Elk habitat suitability model
We used landcover classifications from the 2016 National Land Cover Database (Jin et al. 2019) overlaid with timber harvest data. We categorized non-timber landcover data to generalized landcovers (shrub, herbaceous, dwarf shrub, wetland, water, roads, non-vegetated, and forest) for analysis; timber stands over 60 years old (n = 8) were grouped with forest land cover (Online Resource 1 Table S2). To estimate the effects of terrain on elk habitat suitability, we used 30-m digital elevation models for elevation, slope, and aspect (Danielson and Gesch 2011). Because slope and aspect interact in complex ways, we incorporated slope and aspect into a site severity index (Nielsen and Haney 1998; Boyce et al. 2003). This metric incorporates slope and aspect into an index of site moisture and solar insulation from mesic to xeric conditions (Nielsen and Haney 1998):
We also examined the impact of distance from streams and roads (ADFG, unpublished data) on elk habitat suitability.
To reduce spatial autocorrelation, we thinned elk GPS points to one random observation every 12 h (e.g., one each during 0:00–11:59 and 12:00–23:59; “used” points; Swihart and Slade 1985). We excluded years of GPS data for each elk from the habitat suitability analysis if the elk was not within the timber harvest area (Online Resource 2). As elk habitat use varies seasonally (Irwin and Peek 1983; Unsworth et al. 1998), we grouped elk locations into three seasons: autumn and winter (1 October–20 May), parturition (21 May–12 June; Batchelor 1965), and summer (13 June–30 September). We used continuous time movement models to estimate 85% elk home ranges (Roloff and Kernohan 1999; Powell 2012) using gaussian reference function kernel density estimates with small sample size bias reduction for each elk in each season in each year using the R package “ctmm” (Fleming and Calabrese 2017; R Core Team 2021). We generated random points equal to twice the number of thinned GPS locations within the home range (i.e., available points). For each used and available point, we extracted landcover, elevation, site severity index, and distance to nearest stream and road. Because timber stand landcover type varied among years (Table 1), we used year-specific landcovers with the corresponding year of elk location data.
We normalized continuous variables (elevation, site severity index, and distances), then calculated Pearson product-moment correlations between these variables, finding no strong correlations (|r|< 0.7; Dormann et al. 2013). We fit seasonal mixed effects logistic regression models to determine the effects of landcover, elevation, site severity index, and distance to roads and streams on elk habitat suitability (e.g. response variable as point used [1] or available [0]) using random effect terms for individual elk, year, and sex. We determined statistical significance of individual fixed effect and random effect covariates using Wald tests (α = 0.05) and parameter variance (variance > 0.025) respectively (Sommer and Huggins 1996; Burnham and White 2002). As the fixed effect of site severity index and the random effects of year and sex were insignificant (p > 0.05 and variance < 0.025 respectively) and did not improve model performance for any season (difference in Akaiki information criterion corrected for small sample size [ΔAICc] > 0, Online Resource 1 Table S3), we removed those predictors (Online Resources 1 Tables S3–S4 and Online Resource 2; Sommer and Huggins 1996; Burnham and Anderson 2002), which improved model fit (ΔAICc < − 2 for all seasons; Online Resource 1 Tables S3–S4).
Our final habitat suitability model was:
where U represents if the point was used (1) or available (0), α is the model-estimated intercept, β represents the model-estimated coefficients for each variable, c is the landcover type of stand (a dummy variable with seven coefficients representing timber stand categorizations [Table 1]), e is elevation, r is distance to road, and w is distance to water.
We fit habitat suitability models for autumn–winter, summer, and parturition to obtain three sets of coefficients for optimization, averaging coefficients across all individual elk. We tested the predictive accuracy of our models with fivefold cross-validation to calculate area under the receiver operating characteristic curve (AUC; Swets 1988).
Optimization formulation
Using the coefficients from the habitat suitability model (Eq. 2), we calculated habitat suitability indices (HSI) using mean elevation, mean distance to water, mean distance to nearest road, and landcover for each timber harvest stand (y), in each planning period (5-year intervals, t), where land cover type of each stand varied temporally with timber growth. To optimize habitat suitability, we maximized the area-weighted average habitat suitability index for all timber stands averaged across planning periods:
where t is the planning period, u is the total number of planning periods, y represents the timber harvest stand, n is the total number of timber harvest stands, HSIt,y is the temporally complex habitat suitability index (HSI) for Roosevelt elk during period t in stand y (Eq. 2), and Ay is the area of stand y (hectares).
To optimize for timber production, we maximized the objective function:
where Vty is the number of cubic meters per hectare for stand y during period t based on replanting and stand age, and Xty is a binary matrix of current solutions with n rows and u columns which represents whether unit y was (1) or was not (0) harvested during period t.
To maximize elk habitat suitability and timber harvest simultaneously, we minimized deviations from volume (GV) and habitat suitability (GHSI) goals by minimizing a goal-deviation “multi-objective” objective function separately for each season:
We set timber harvest volume and habitat suitability goals as the best objective function values from individual optimizations for timber harvest and habitat suitability in each season. We set a minimum harvest age constraint as 50 years for Juneau and 70 years for naturally-regenerating stands (Koniag Inc., personal communication):
where M is a binary matrix with n rows and u columns that represent whether stand y was harvestable (1; age ≥ 50 years for Juneau strain and ≥ 70 years for natural regeneration) or not-harvestable (0) in period t. Finally, from conversations with Native corporations and analysis of historical harvest (average stands harvested in a 5-year period = 106), we set a maximum harvest constraint where the number of stands harvested in a planning period could not exceed 100:
We determined optimal solutions for timber harvest volume, habitat suitability for each season, and multi-objective optimization functions for each season. Due to the recent harvest of timber on Afognak Island and management goals of Native corporations (Koniag Inc., personal communication), we began our 50-year planning horizon in 2050 by adding 28 years to the 2022 ages of all stands. Due to stand age constraints, only 52 and 92 stands were available for harvest during the first two planning periods, respectively and no stand could be harvested twice in this analysis. Therefore, the maximum number of stands available for harvest during the planning horizon within constraints was 892.
Optimization technique
Because our optimization problem was nonlinear and temporally dynamic, we used a heuristic threshold accepting technique (Dueck and Scheuer 1990). Though heuristic techniques do not guarantee a global optimal solution, they produce feasible and efficient solutions to complex problems (Bettinger et al. 2002; Gilli et al. 2019). Our formulation used a neighborhood function that changed the harvest period of one stand at a time subject to the constraints (“1-opt moves”; Online Resources 2, 6; Bettinger et al. 1999). We implemented the threshold-accepting algorithm using the “nmof” package in program R (Online Resources 2, 6; Gilli et al. 2019; Schumann 2020; R Core Team 2021). As this optimization method required minimization (Schumann 2020), we minimized the negative value of the timber harvest and habitat suitability objective functions.
Threshold-accepting optimization requires the user to provide a sequence of thresholds that decline to zero and a number of steps for each threshold (Online Resource 2). We determined thresholds using the default method in the “nmof” package (Gilli et al. 2006; Schumann 2020), which requires three user-defined parameters (number of steps, quantile for starting threshold, and number of thresholds). To determine the best combination of parameters, we ran each optimization eight or more times at several parameter levels, then selected the combination of parameters that provided the best objective function value (Online Resources 2, 6; Bettinger et al. 2009). Using these parameters, we repeated the timber harvest optimization 500 times and the remaining optimizations 50 times (due to computer processing time constraints). As alternative plans included varying objective variables with different model coefficients, we used coefficients of variation to measure the consistency of optimization results (Online Resource 2; Hurme et al. 2007; Bettinger et al. 2009).
The optimization model required several assumptions. First, we assumed no timber harvest would occur on Afognak Island during 2023–2050. As most harvestable timber land owned by Native corporations has been harvested, most timber harvest was completed on Afognak Island by 2021. Afognak Native Corporation continues limited timber harvest on the island, but data is not available on future timber harvest stand locations. Second, we assumed that all harvested stands were replanted with Juneau strain Sitka spruce during the planning period in which they were harvested; since 2012 Native corporations have replanted stands within 5 years of harvest. Finally, we assumed no new roads would be created on Afognak Island, landcover on Afognak Island would not change other than timber harvest, and elk habitat selection and timber value would not change over time.
Results
Habitat suitability
Elk habitat suitability model fit based on K-fold cross validation was modest (autumn–winter AUC = 0.654, parturition AUC = 0.652, summer AUC = 0.586). During all seasons, elk habitat suitability decreased with increasing elevation (Fig. 3a, Online Resource 1 Table S4). During autumn–winter, elk habitat suitability increased with increasing distance from roads, during parturition, habitat suitability was unrelated to road distance, and during summer, elk habitat suitability decreased with increasing distance from roads (Fig. 3b). During autumn–winter and summer, elk habitat suitability increased with increasing distance from streams, while during parturition distance from streams had no effect on elk habitat suitability (Fig. 3c).

Elk habitat suitability model results showing the effects of elevation (a), distance to road (b), distance to water (c), and landcover (d; including timber harvest successional categories: no cover, no forage [NCNF]; no cover, low forage [NCLF]; no cover, medium forage [NCMF]; medium cover, medium forage [MCMF]; medium canopy, low forage [MCLF]; and high canopy, no forage [HCNF]) during autumn–winter (1 October–20 May), parturition (21 May–12 June), and summer (13 June–30 September), with 95% confidence intervals, Afognak Island, Alaska, 2016–2021
During all seasons, elk selected shrub and herbaceous landcovers frequently, avoided water and older-aged timber harvest stands with high cover and no forage, and exhibited intermediate use of wetlands (Fig. 3d; Online Resource 1 Table S4). During autumn–winter, elk selected timber harvest regrowth with low to no cover and low to medium forage, avoided dwarf shrub landcover and timber harvest regrowth with medium and high cover and low to no forage, and demonstrated intermediate use of other landcovers. During parturition, elk selected young to intermediate-aged timber regrowth with no to medium cover and low to high forage, avoided newly harvested stands with no cover and no forage, dwarf shrub, and forest landcovers, and demonstrated intermediate use of other landcovers. During summer, elk selected dwarf shrub landcover and avoided non-vegetated areas, and demonstrated intermediate use of other landcovers.
Optimization
The timber harvest optimization resulted in the highest timber yield (21.2% increase over habitat suitability optimization timber yield [averaged across seasons]) and the lowest habitat suitability (5.7% loss compared to maximum suitability [averaged across seasons]; Table 2; Figs. 4 and 5; Online Resource 1 Fig. S1). Conversely, the habitat suitability optimizations had the lowest timber yield (17.5% loss compared to maximum yield [averaged across seasons]), and the highest habitat suitability (6.1% increase over timber harvest habitat suitability [averaged across seasons]). The multi-objective optimizations reached intermediate solutions between the timber harvest and habitat suitability maxima (1.4% loss compared to optimum yield; 3.9% loss compared to optimum suitability [averaged across seasons]). The habitat suitability and multi-objective optimization solutions varied among seasons, reflecting respective seasonal changes in elk habitat use (Figs. 3, 4, 5 and 6; Online Resource 1 Figs. S1–S3).

Area of harvested timber stands (hundreds of hectares) across 5-year planning periods by stand age (years) and replanting type (Juneau strain or natural regeneration [natural regen.]) from optimal solutions for timber harvest, elk habitat suitability (suitability) during autumn–winter (1 October–20 May), parturition (21 May–12 June), and summer (13 June–30 September), and multi-objective suitability and timber harvest (multi-objective) during three seasons, Afognak Island, Alaska, 2050–2100

Elk habitat suitability across 5-year planning periods showing percent area of each landcover type across timber stands under optimal solutions for timber harvest (average suitability across three seasons), suitability during autumn–winter (1 October–20 May), parturition (21 May–12 June), and summer (13 June–30 September), and multi-objective suitability and timber harvest (multi-objective) during three seasons, Afognak Island, Alaska, 2050–2100

Timber stands selected for harvest during each period from optimal solutions for timber harvest; elk habitat suitability (suitability) during autumn–winter (1 October–20 May), parturition (21 May–12 June), and summer (13 June–30 September); and multi-objective timber harvest and suitability (multi-objective) during the three seasons, Afognak Island, Alaska, 2050–2100
Timing and frequency of timber harvest varied between optimal solutions for different seasonal objectives (Figs. 4, 5 and 6; Online Resource 1 Figs. S1–S3, Online Resource 2). When timber harvest alone was maximized, the maximum number of stands was selected for harvest during each period within constraints and larger-area stands were harvested later in the 50-year planning horizon. The optimal solution for elk habitat suitability during autumn–winter had the most stands harvested and therefore the highest yield among the habitat suitability optimizations (15.5% yield loss compared to optimum; Table 2). The optimal solution for parturition habitat suitability resulted in an intermediate number of stands harvested and similarly intermediate loss of yield (16.8%), while the optimal solution for summer habitat suitability had the fewest stands harvested and therefore the lowest yield (20.0% loss). The optimal solution from the multi-objective optimization for summer elk habitat suitability resulted in the lowest number of stands harvested among the multi-objective optimizations, but the highest yield (1.6% loss compared to optimum), because larger-area stands were harvested later. The multi-objective optimizations for elk habitat suitability during autumn–winter and parturition resulted in similar solutions, with the same number of stands harvested and similar yield loss (1.5% and 2.0% respectively). The optimal solution for timber harvest yield resulted in the greatest loss of elk habitat suitability during parturition (6.5% loss compared to optimum), followed by autumn–winter (5.6% loss), then summer (5.0% loss). The multi-objective optimizations resulted in similar elk habitat suitability losses across seasons (all seasons 3.9% loss compared to optimal solutions).
Discussion
Our aim was to develop a method for timber harvest scheduling that balanced seasonal elk habitat suitability with timber harvest volume for a 50-year planning horizon. Optimization of the timber harvest, habitat suitability, and multi-objective functions resulted in expected variations in timber volume yield and habitat suitability. During all seasons, elk selected herbaceous vegetation and shrub landcovers over forests, indicating that in this system, forage may be more limiting for elk than cover.
Optimizing for timber harvest volume resulted larger-area older stands harvested later in the planning horizon. However, the optimal solution for timber harvest resulted in lower elk habitat suitability than other solutions because older stands have lower forage availability and therefore lower elk habitat value than earlier successional stages (Skovlin et al. 1989; Visscher and Merrill 2009). Though extensive timber harvest may increase elk habitat suitability in the short-term, a homogenous landscape of mid- to late-successional stages later in forest regeneration reduces long-term habitat value for elk (Starkey et al. 1982; Irwin and Peek 1983; Wisdom et al. 2004).
In autumn–winter, timber harvest increased elk habitat value because elk selected recently harvested early-successional timber stands over mid- to late-successional and mature forest stands. Elk may spend more time in open habitat during winter than during other seasons because forage is less palatable and more difficult to obtain, increasing the foraging time required to acquire sufficient energy (Starkey et al. 1982; Unsworth et al. 1998; Christianson and Creel 2007). Elk selected mature forest stands over mid- to late-successional stands, indicating that forests may provide better shelter from predation and severe weather as compared to late-successional timber stands (Starkey et al. 1982). Severe winters on Afognak Island can cause high elk mortality (Alexander 1972), and elk may mitigate effects of winter by using mature forest, especially after heavy snowfall (Troyer 1960; Batchelor 1965). During parturition, rather than reducing predation risk by using forest habitat with high canopy cover (Troyer 1960; Witmer and DeCalesta 1983), elk used early to mid-successional habitat, leading to an optimal solution with intermediate timber harvest. During calving, elk may select shrubby early-successional habitat over forests as it may offer denser vegetation, which provides “hiding” cover and additional foraging resources (Barbknecht et al. 2011; Rearden et al. 2011). The optimal solution for elk habitat suitability during summer had the lowest timber yield because elk used forests similarly to early- to mid-successional timber stands. Elk may use forests to shelter from vehicular traffic or logging operations during summer (Unsworth et al. 1998; Wisdom et al. 2004), or for thermal cover, as forests may provide a cooler microclimate which protects elk from high temperatures (Starkey et al. 1982; Long et al. 2014).
By using multi-objective optimization, we found solutions that achieved approximately 99% of optimal timber yield while maintaining greater elk habitat suitability than the single-objective timber yield optimization. To improve elk habitat suitability during the next timber harvest planning horizon, our optimizations suggest harvesting smaller stands before they reach their maximum yield potential while leaving larger stands to be harvested at peak yield, and also leaving some stands unharvested (Skovlin et al. 1989; Visscher and Merrill 2009; Rumble and Gamo 2011). This harvest system may better mimic forest dynamics following natural disturbances and provide continuous regeneration that prevents a decline in elk habitat suitability (Irwin and Peek 1983; Skovlin et al. 1989; Jenkins and Starkey 1996). Additionally, we suggest that before the beginning of our planning horizon (i.e., over the next 23 years), managers may consider harvesting small areas within late-successional stands to create gaps in the forest canopy and allow for increased forage production (Starkey et al. 1982; Skovlin et al. 1989).
While there are multiple complex drivers of elk population dynamics (Brodie et al. 2013; Lukacs et al. 2018; Schooler et al. 2022), in northern regions elk populations are generally limited by weather during winter and forage availability during summer and autumn (Brodie et al. 2013; Cook et al. 2013; Lukacs et al. 2018; Schooler et al. 2022). Additionally, because elk demonstrated increased use of higher elevation dwarf shrub habitat during summer, they may be less affected by timber harvest during summer compared to the other seasons. Therefore, managers may consider prioritizing the multi-objective optimization solution that incorporates autumn–winter habitat suitability over other solutions. This solution recommends limited harvest of stands early in the planning horizon and harvesting smaller Juneau strain stands before they reach peak maturation, while leaving larger-area stands to be harvested later in the planning horizon.
We recognize several limitations of our study. Though ungulates are strongly influenced by landscape complexity and edge effects (Skovlin et al. 1989; Rowland et al. 2018), we were unable to incorporate adjacent habitat into suitability because our study landscape was relatively large, our problem was temporally complex, and we used a large number of planning periods. We also did not consider balance of economic benefits between Native corporations, or economic advantages of harvesting spatially continuous stands. Because very few forest stands on Afognak Island were older than 50 years, we were unable to separate mature timber stands from unharvested old growth forest in our habitat suitability model and optimizations, even though previously-harvested mature timber stands likely have different habitat characteristics than unharvested old growth (Arris and Farr 1974). Additionally, though thinning, burning, fertilization, and alternative harvesting strategies may be beneficial for elk habitat suitability because they encourage early successional stages and maximize forage productivity (Starkey et al. 1982; Skovlin et al. 1989), we were unable to account for these management scenarios due to problem complexity and lack of data. Similarly, though we could not account for replanting schedules in our optimization, managers could consider delaying replanting harvested stands, as immediate replanting may decrease habitat suitability by reducing the length of forage-rich early successional stages (Starkey et al. 1982). Climate change has increasing effects on habitats (e.g. changes in vegetation phenology and growth; Root et al. 2003) and animal behavior (Beever et al. 2017), which may change elk habitat use and timber growth and therefore optimal solutions.
Though elk benefited from timber harvest, optimizing for either timber harvest or habitat suitability alone resulted in a decrease of the other objective. However, our goal-deviation multi-objective optimization reached a compromise between optimal timber harvest and elk habitat suitability. Though wildlife management and financial goals may conflict, multi-objective heuristic optimization using habitat suitability models is a technique that can be implemented across wildlife species. As over 15% of the earth’s terrestrial surface is managed for timber harvest (FAO 2016), there is great concern about the impacts of timber harvest on biodiversity (Fisher and Wilkinson 2005; Kellner et al. 2019). Multi-objective optimization that incorporates data-driven habitat suitability models provides a framework to maintain adequate habitat suitability while allowing for necessary resource extraction. Given the availability of animal location data (e.g., MoveBank [Wikelski et al. 2022], Global Biodiversity Information Facility [GBIF Secretariat 2022]) and our use of open source software (R [Schumann 2020; R Core Team 2021]), our approach can be more broadly applied for wildlife-oriented timber harvest management.
References
Alaska Department of Fish and Game (2022) 2022–2023 Alaska hunting regulations: governing general, subsistence, and commercial uses of Alaska’s wildlife. Alaska Department of Fish and Game, Juneau
Alden JN (1998) Effects of seed shelters on emergence, survival, and growth of Sitka spruce seedlings on Afognak island. In: Wheeler R (ed) Alaska Extension Review 1997 Trial Season. Alaska Cooperative Extension, Fairbanks, pp 44–49
Alexander JE (1972) Seasonal movements of elk. Alaska Department of Fish and Game, Juneau
Arris AS, Farr WA (1974) Forest ecology and timber management. In: The Forest Ecosystem of Southeast Alaska. United States Forest Service Pacific Northwest Research Station, Portland
Barbknecht AE, Fairbanks WS, Rogerson JD et al (2011) Elk parturition site selection at local and landscape scales. J Wildl Manage 75:646–654. https://doi.org/10.1002/jwmg.100
Batchelor RF (1965) The Roosevelt elk in Alaska: its ecology and management. Alaska Department of Fish and Game, Juneau
Beever EA, Hall LE, Varner J et al (2017) Behavioral flexibility as a mechanism for coping with climate change. Front Ecol Environ 15:299–308
Bettinger P, Boston K, Sessions J (1999) Combinatorial optimization of elk habitat effectiveness and timber harvest volume. Environ Model Assess 4:143–153. https://doi.org/10.1023/A:1019051931272
Bettinger P, Sessions J, Boston K, Murray AT (2009) A review of the status and use of validation procedures for heuristics used in forest planning. Math Comput for Nat Sci 1:26–37
Bettinger P, Graetz D, Boston K et al (2002) Eight heuristic planning techniques applied to three increasingly difficult wildlife planning problems. Silva Fenn 36:561–584. https://doi.org/10.14214/sf.545
Boyce MS, Mao JS, Merrill EH et al (2003) Scale and heterogeneity in habitat selection by elk in Yellowstone National Park. Ecoscience 10:421–431
Brodie J, Johnson H, Mitchell M et al (2013) Relative influence of human harvest, carnivores, and weather on adult female elk survival across western North America. J Appl Ecol 50:295–305. https://doi.org/10.1111/1365-2664.12044
Burnham KP, Anderson DD (2002) Model selection and multimodel inference: a practical information-theoretic approach, 2nd edn. Springer, New York
Burnham KP, White GC (2002) Evaluation of some random effects methodology applicable to bird ringing data. J Appl Stat 29:245–264. https://doi.org/10.1080/02664760120108755
Christianson DA, Creel S (2007) A review of environmental factors affecting elk winter diets. J Wildl Manage 71:164–176. https://doi.org/10.2193/2005-575
Cook RC, Cook JG, Vales DJ et al (2013) Regional and seasonal patterns of nutritional condition and reproduction in elk. Wildl Monogr. https://doi.org/10.1002/wmon.1008
Danielson JJ, Gesch DB (2011) Global multi-resolution terrain elevation data 2010 (GMTED2010)
Díaz-Balteiro L, Romero C (2003) Forest management optimisation models when carbon captured is considered: a goal programming approach. For Ecol Manage 174:447–457. https://doi.org/10.1016/S0378-1127(02)00075-0
Dormann CF, Elith J, Bacher S et al (2013) Collinearity: a review of methods to deal with it and a simulation study evaluating their performance. Ecography (cop) 36:27–46. https://doi.org/10.1111/j.1600-0587.2012.07348.x
Dueck G, Scheuer T (1990) Threshold accepting: a general purpose optimization algorithm appearing superior to simulated annealing. J Comput Phys 90:161–175. https://doi.org/10.1016/0021-9991(90)90201-B
Esri, Maxar, Earthstar Geographics (2022) World Imagery [basemap]. https://www.arcgis.com/home/item.html?id=10df2279f9684e4a9f6a7f08febac2a9. Accessed 4 Jan 2022
FAO (2016) Global forest resources assessment 2015. Food and Agriculture Organization of the United Nations, Rome
Finnegan SP, Svoboda NJ, Fowler NL et al (2021) Variable intraspecific space use supports optimality in an apex predator. Sci Rep. https://doi.org/10.1038/s41598-021-00667-y
Fisher JT, Wilkinson L (2005) The response of mammals to forest fire and timber harvest in the North American boreal forest. Mamm Rev 35:51–81. https://doi.org/10.1111/j.1365-2907.2005.00053.x
Fleming CH, Calabrese JM (2017) A new kernel density estimator for accurate home-range and species-range area estimation. Methods Ecol Evol 8:571–579. https://doi.org/10.1111/2041-210X.12673
Forsman ED, Kaminski TJ, Lewis JC et al (2005) Home range and habitat use of northern spotted owls on the Olympic Peninsula, Washington. J Raptor Res 39:365–377
Frair JL, Merrill EH, Visscher DR et al (2005) Scales of movement by elk (Cervus elaphus) in response to heterogeneity in forage resources and predation risk. Landsc Ecol 20:273–287. https://doi.org/10.1007/s10980-005-2075-8
Garroutte EL, Hansen AJ, Lawrence RL (2016) Using NDVI and EVI to map spatiotemporal variation in the biomass and quality of forage for migratory elk in the greater Yellowstone ecosystem. Remote Sens. https://doi.org/10.3390/rs8050404
Gilli M, Këllezi E, Hysi H (2006) A data-driven optimizaiton heuristic for downside risk minimization. J Risk 8:1–19. https://doi.org/10.2139/ssrn.910233
Gilli M, Maringer D, Schumann E (2019) Numerical methods and optimization in finance, 2nd edn. Academic Press, London
Google Earth (2022) Google Earth Pro V 7.3.4.8248. 1984–2022. Afognak Island, USA. 58°13;18.30″N, 152°30;22.91″ W, eye alt 50,000 ft. Landsat and Copernicus. Accessed 1 May 2022
Hansen D (2018) Growth and yield of Sitka spruce on Afognak Island, Alaska is low, or is it? Thesis, State University of New York College of Environmental Science and Forestry
Harris AS (1972) Natural reforestation after logging on Afognak Island. United States Forest Service Pacific Northwest Research Station, Portland
Hurme E, Kurttila M, Mönkkönen M et al (2007) Maintenance of flying squirrel habitat and timber harvest: a site-specific spatial model in forest planning calculations. Landsc Ecol 22:243–256. https://doi.org/10.1007/s10980-006-9019-9
Irwin LL, Peek JM (1983) Elk habitat use relative to forest succession in Idaho. J Wildl Manage 47:664–672
Jenkins K, Starkey E (1996) Simulating secondary succession of elk forage values in a managed forest landscape, western Washington. Environ Manage 20:715–724. https://doi.org/10.1007/BF01204142
Jin S, Homer C, Yang L et al (2019) Overall methodology design for the United States national land cover database 2016 products. Remote Sens. https://doi.org/10.3390/rs11242971
Kellner KF, Renken RB, Millspaugh JJ et al (2019) Effects of forest management on vertebrates: synthesizing two decades of data from hardwood forests in Missouri, USA. Ecol Appl 29:1–16. https://doi.org/10.1002/eap.1993
Larson MA, Thompson FR, Millspaugh JJ et al (2004) Linking population viability, habitat suitability, and landscape simulation models for conservation planning. Ecol Modell 180:103–118. https://doi.org/10.1016/j.ecolmodel.2003.12.054
Lawler JJ, Lewis DJ, Nelson E et al (2014) Projected land-use change impacts on ecosystem services in the United States. Proc Natl Acad Sci USA 111:7492–7497. https://doi.org/10.1073/pnas.1405557111
Long RA, Bowyer RT, Porter WP et al (2014) Behavior and nutritional condition buffer a large-bodied endotherm against direct and indirect effects of climate. Ecol Monogr 84:513–532. https://doi.org/10.1890/13-1273.1
Lukacs PM, Mitchell MS, Hebblewhite M et al (2018) Factors influencing elk recruitment across ecotypes in the Western United States. J Wildl Manage 82:698–710. https://doi.org/10.1002/jwmg.21438
Menne MJ, Durre I, Korzeniewski B et al (2012) Global historical climatology network—Daily (GHCN-Daily), Version 3. [USW00025501 1955–2020]. https://mrcc.purdue.edu/CLIMATE/. Accessed 16 Feb 2021
Nelson E, Mendoza G, Regetz J et al (2009) Modeling multiple ecosystem services, biodiversity conservation, commodity production, and tradeoffs at landscape scales. Front Ecol Environ 7:4–11. https://doi.org/10.1890/080023
Nielsen SE, Haney A (1998) Gradient responses for understory species in a bracken grassland and northern dry forest ecosystem of northeast Wisconsin. Trans Wisconsin Acad Sci Arts Lett 86:149–166
Pepke E (2010) Global wood markets: consumption, production, and trade. In: International Forestry and Global Issues. UNECE/FAO Timber Section, Geneva
Powell RA (2012) Movements, home ranges, activity, and dispersal. In: Boitani L, Powell RA (eds) Carnivore ecology and conservation: a handbook of techniques. Oxford University Press, New York, pp 189–217
R Core Team (2021) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Version 4.0.4. https://www.r-project.org/
Rearden SN, Anthony RG, Johnson BK (2011) Birth-site selection and predation risk of Rocky Mountain elk. J Mammal 92:1118–1126. https://doi.org/10.1644/09-MAMM-A-019A.1
Rettie WJ, Messier F (2000) Hierarchical habitat selection by woodland caribou: its relationship to limiting factors. Ecography (Cop) 23:466–478. https://doi.org/10.1111/j.1600-0587.2000.tb00303.x
Roloff GJ, Kernohan BJ (1999) Evaluating reliability of habitat suitability index models. Wildl Soc Bull 27:973–985
Root TL, Price JT, Hall KR et al (2003) Fingerprints of global warming on wild animals and plants. Nature 421:57–60. https://doi.org/10.1038/nature01333
Rowland MM, Wisdom MJ, Nielson RM et al (2018) Modeling elk nutrition and habitat use in western Oregon and Washington. Wildl Monogr 199:1–69. https://doi.org/10.1002/wmon.1033
Rumble MA, Gamo RS (2011) Habitat use by elk (Cervus elaphus) within structural stages of a managed forest of the northcentral United States. For Ecol Manage 261:958–964. https://doi.org/10.1016/j.foreco.2010.12.005
Schooler SL, Svoboda NJ, Finnegan SP et al (2022) Maternal carryover, winter severity, and brown bear abundance relate to elk demographics. PLoS One 17:e0274359. https://doi.org/10.1371/journal.pone.0274359
Schumann E (2020) Financial optimisation with R. https://enricoschumann.net/files/NMOFman.pdf. Accessed 30 Jun 2022
GBIF Secretariat (2022) What is GBIF? GBIF: the Global Biodiversity Information Facility. https://www.gbif.org/what-is-gbif
Sexton JO, Song XP, Feng M et al (2013) Global, 30-m resolution continuous fields of tree cover: Landsat-based rescaling of MODIS vegetation continuous fields with lidar-based estimates of error. Int J Digit Earth 6:427–448. https://doi.org/10.1080/17538947.2013.786146
Skovlin JM, Bryant LD, Edgerton PJ (1989) Timber harvest affects elk distribution in the Blue Mountains of Oregon. United States Forest Service Pacific Northwest Research Station, Portland
Sommer S, Huggins RM (1996) Variables selection using the Wald test and a robust CP. J R Stat Soc 45:15–29. https://doi.org/10.2307/2986219
Starkey EE, DeCalesta DS, Witmer GW (1982) Management of Roosevelt elk habitat and harvest. In: Sabol K (ed) Transactions of the 47th North American wildlife and natural resources conference. United States National Park Service, Washington, pp 353–362
Steventon JD, KacKenzie KL, Mahon TE (1998) Response of small mammals and birds to partial cutting and clearcutting in northwest British Columbia. For Chron 74:703–713
Swets JA (1988) Measuring the accuracy of diagnostic systems. Science 240:1285–1293. https://doi.org/10.1126/science.3287615
Swihart RK, Slade NA (1985) Testing for independence of observations in animal movements. Ecology 66:1176–1184. https://doi.org/10.2307/1939170
Thomas DL, Taylor EJ (2006) Study designs and tests for comparing resource use and availability II. J Wildl Manage 70:324–336. https://doi.org/10.2193/0022-541X(2006)70[324:SDATFC]2.0.CO;2
Troyer WA (1960) The Roosevelt elk on Afognak Island, Alaska. J Wildl Manage 24:15–21. https://doi.org/10.2307/3797351
U.S. Geological Survey (1988) Aerial photo single frames high resolution. https://doi.org/10.5066/F7610XKM. Accessed 1 May 2021
U.S. Geological Survey (2006) Aerial photography high resolution orthoimagery (HRO). https://doi.org/10.5066/F73X84W6. Accessed 1 May 2021
U.S. Geological Survey (2022) Landsat 8–9 operational land imager and thermal infrared sensor collection 2 level-1 data. https://doi.org/10.5066/P975CC9B. Accessed 30 Jun 2022
U.S. Geological Survey, Google (2007) Landsat 8 collection 1 tier 1 top of atmosphere reflectance. In: Google Earth Engine Data Cat. https://developers.google.com/earth-engine/datasets/catalog/LANDSAT_LC08_C01_T1_TOA. Accessed 4 Jan 2022
Unsworth JW, Kuck L, Garton EO, Butterfield BR (1998) Elk habitat selection on the Clearwater National Forest, Idaho. J Wildl Manage 62:1255–1263
Visscher DR, Merrill EH (2009) Temporal dynamics of forage succession for elk at two scales: implications of forest management. For Ecol Manage 257:96–106. https://doi.org/10.1016/j.foreco.2008.08.018
Wikelski M, Davidson SC, Kays R (2022) Movebank: archive, analysis and sharing of animal movement data. Max Planck Institute of Animal Behavior. https://www.movebank.org
Wisdom MJ, Bright LR, Carey CG et al (1986) A model to evaluate elk habitat in western Oregon. United States Forest Service Pacific Northwest Region, Portland
Wisdom MJ, Johnson BK, Vavra M et al (2004) Cattle and elk responses to intensive timber harvest. In: Rahm J (ed) Transactions of the 69th North American wildlife and natural resources conference. Wildlife Management Institute, Spokane, pp 197–216
Witmer GW, DeCalesta DS (1983) Habitat use by female Roosevelt elk in the Oregon coast range. J Wildl Manage 47:933–939. https://doi.org/10.2307/3808152
Yemshanov D, Haight RG, Liu N et al (2020) Assessing the trade-offs between timber supply and wildlife protection goals in boreal landscapes. Can J for Res 50:243–258. https://doi.org/10.1139/cjfr-2019-0234
Acknowledgements
We thank the Alaska Department of Fish and Game, Rocky Mountain Elk Foundation, Kodiak Brown Bear Trust, and Camp Fire Conservation Fund for financial support. Afognak Native Corporation, Koniag Native Corporation, Koncor, Natives of Kodiak Native Corporation, Ouzinkie Native Corporation, and Old Harbor Native Corporation provided timber harvest data and logistical support. We appreciated information and advice from P. Olsen and B. Wall on Afognak timber harvest and the assistance of H. Valley, C. Coulter, and N. Hayden in timber harvest data compilation. We thank K. Kellner for statistical advice. Field support was provided by F. Bled, A. Christofferson, J. Crye, J. De La Pena, J. Fieldman, N. Jensen, A. Jones, J. Larivee, J. Mortenson, T. Petroelje, J. Rich, C. Ramsey, L. Tyrrell, L. Van Daele, and K. Wattum.
Funding
This project was supported by a Natural Resources Conservation Service Conservation Innovation Grant and a United States Fish and Wildlife Service Federal Aid in Wildlife Restoration Act under Pittman-Robertson project AKW-12 through the Alaska Department of Fish and Game. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Author information
Authors and Affiliations
Contributions
Conceptualization: SS, JB; Methodology: SS, CK, JB; Investigation: SS, SF, NS; Formal analysis: SS; Writing—original draft preparation: SS; Writing—review and editing: all authors; Funding acquisition: NS and JB; Resources: NS and CK; Supervision: JB.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Conflict of interest
The authors have no relevant financial or non-financial interests to disclose.
Data availability
Elk location data are protected and therefore unavailable for public distribution. Timber harvest data and Juneau strain growth tables are unavailable due to agreements with Native corporations. All other data and code for the optimization model are provided in online resources.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Schooler, S.L., Svoboda, N.J., Kroll, C.N. et al. Multi-objective optimization for timber harvest management incorporating wildlife habitat goals. Landsc Ecol 38, 3097–3113 (2023). https://doi.org/10.1007/s10980-023-01766-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10980-023-01766-z