
Abstract
Context
Individual species often drive habitat restoration action; however, management under this paradigm may negatively affect non-target species. Prioritization frameworks which explicitly consider benefits to target species while minimizing consequences for non-target species may improve management strategies and outcomes.
Objectives
We examined extents to which conifer removal, an approach frequently implemented to restore sagebrush ecosystems, can be conducted without detrimental effects to conifer-associated species, including the imperiled Pinyon Jay (Gymnorhinus cyanocephalus). Additionally, we prioritized sites for conifer removal, and predicted abundance responses for multiple species following simulated conifer removal at selected sites to achieve variable management objectives.
Methods
We used model-predicted changes in species’ densities following simulated conifer removal to identify optimal removal sites under single species, multi-species (ecosystem), and multi-ecosystem management scenarios. We simulated conifer removal at prioritized sites and evaluated resulting changes in abundance for six passerine species.
Results
Management prioritized for a single species (Brewer’s Sparrow) provided the greatest per-unit-effort benefits for that species but resulted in the lowest population outcomes for all other species considered. In comparison, prioritizations for multiple species within a single ecosystem (i.e., pinyon–juniper or sagebrush) resulted in larger population benefits for species associated with that ecosystem and reduced detrimental effects on non-target species associated with another ecosystem. For example, single species management for Brewer’s Sparrow resulted in an average increase of 1.38% for sagebrush-associated species and a 4.58% decrease for pinyon–juniper associated species. In contrast, when managing for multiple sagebrush-associated species sagebrush-associated songbird populations increased by 3.98% and pinyon–juniper associated species decreased by 2.36%, on average.
Conclusions
Our results illustrate single species management can result in detrimental outcomes and/or opportunity costs for non-target species compared to management designed to benefit multiple species. Our framework can be used to balance undesired consequences for non-target species and is adaptable for other systems and taxa.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Worldwide, habitat loss and fragmentation have drastically increased the number of species recognized as extinct, at-risk, or in-decline, and this trend is expected to continue (Kerr and Deguise 2004; Haddad et al. 2015; Tilman et al. 2017). With so many species of conservation concern, monitoring all potentially at-risk populations is impractical. As such, conservation efforts largely focus on single species or subsets of species (e.g., umbrella, flagship, and keystone species; Simberloff 1998; Caro and O’Doherty 1999), tracking their populations or responses to management to infer outcomes for communities and ecosystems. Despite the common use of single species to guide conservation planning, growing evidence indicates such management may only rarely confer adequate benefits for all co-occurring species (Simberloff 1998; Roberge and Angelstam 2004; Roberge et al. 2008; Carlisle et al. 2018; Wang et al. 2021).
Conservationists have been shifting from single species management paradigms towards multiple species approaches (Lambeck 1997; Link 2002; White et al. 2013). Though daunting in terms of data collection and management actions, considering multiple species provides exciting opportunities to conserve biodiversity, maintain ecosystem function, and enhance resistance and resilience of ecosystems (Allen et al. 2011; Harvey et al. 2017). Unfortunately, as the footprint of intact ecosystems decreases globally (Theobald et al. 2020), it is increasingly difficult to adequately address the needs of species with disparate habitat requirements where they overlap geographically. Balancing the needs of declining species associated with differing habitats within the same landscape represents a “wicked problem” (see DeFries and Nagendra 2017). A special issue in a 2001 Wildlife Society Bulletin provides a description of one such scenario:
Certainly the loss of old-growth forests and the degradation and fragmentation of second-growth forests in eastern North America are major concerns, but another legitimate concern is the decline of early successional habitats dominated by grass, shrubs, or young trees (Askins 2001).
In other words, conservation efforts must address the full suite of successional stages and ecosystems occurring on the landscape. While fundamentally simple, this presents a challenge both in terms of a cultural paradigm shift (i.e., not favoring a single ecosystem or successional stage over another) and in coordinating conservation planning, given the dynamic process of succession and the tendency for resources to be allocated to address local conservation or management issues.
Recent advances in multi-species optimizations (e.g., Ball et al. 2009; Watts et al. 2009; Duchardt et al. 2021) provide a potential solution for balancing disparate needs of wildlife species by optimizing management actions based on expected multi-species outcomes. These systematic conservation planning approaches rely on algorithms (e.g., integer linear programming, simulated annealing) to evaluate multiple spatial configurations of selected parcels when prioritizing sites for conservation action (Ball et al. 2009; Hanson et al. 2021). These prioritization approaches represent an objective, quantifiable, and repeatable way to prioritize management sites across ecosystems, thus providing a viable framework for decision-making when stakeholders have multiple opposing objectives, or management actions have inherent trade-offs.
Ecotones between pinyon–juniper woodlands and sagebrush (Artemisia spp.) shrublands of western North America represent an emerging instance where conservation actions aimed at benefitting one suite of species (sagebrush-associates) may have undesired cross-ecosystem consequences. Land managers are increasingly attempting to restore sagebrush communities by removing pinyon pine (Pinus edulis and Pinus monophylla) and juniper (Juniperus spp.; hereafter, “conifer removal”) from sagebrush and pinyon–juniper ecotones (Reinhardt et al. 2020). Conifer removal treatments are largely conducted to combat the encroachment of pinyon–juniper, which degrades sagebrush systems, and aid in the recovery of the declining Greater Sage-Grouse (Centrocercus urophasianus; hereafter, “Sage-Grouse”) (Miller et al. 2019; Reinhardt et al. 2020). Conifer removal enhances habitat suitability, use, survival, and breeding success of Sage-Grouse (Baruch-Mordo et al. 2013; Coates et al. 2017; Cook et al. 2017; Severson et al. 2017a, b). Additional research indicates conifer removal may help restore the sagebrush ecosystem function by increasing soil moisture and perennial herbaceous groundcover (Roundy et al. 2014a, b), as well as increasing sagebrush-associated songbird abundances (Holmes et al. 2017).
Unfortunately, recent studies indicate these treatments may have unintended negative consequences for a suite of pinyon–juniper associated species, which are also of conservation concern (Holmes et al. 2017; Magee et al. 2019; Zeller et al. 2021). The Pinyon Jay (Gymnorhinus cyanocephalus) demonstrates a particularly close mutualistic relationship to pinyon–juniper woodlands—trees supply jays with food and the jays effectively disperse seeds (Johnson and Balda 2020). The Pinyon Jay has been exhibiting range-wide, long-term, population declines (Sauer et al. 2020), which led to a recent petition to list the species under the Endangered Species Act (ESA; Defenders of Wildlife 2022). Recently, density–habitat relationships indicate Pinyon Jay may occur at highest densities in early successional phases of pinyon–juniper woodland expansion (Van Lanen et al. 2023a), where they forage and cache food (Boone et al. 2021). These early successional phases of pinyon–juniper expansion are often preferentially targeted for conifer removal to recover Sage-Grouse because they exhibit low tree density and frequently possess intact sagebrush understories (Roundy et al. 2014a; Natural Resource Conservation Service 2015; Miller et al. 2019). These characteristics allow managers to quickly revert early successional pinyon–juniper woodlands to sagebrush habitats at relatively low expense. Thus, land managers are now challenged with maintaining viable sagebrush ecosystems capable of supporting Sage-Grouse, while avoiding the removal of conifer at sites supporting high abundances of Pinyon Jay and other at-risk pinyon–juniper associated wildlife.
This ecological zero-sum game, where restoration of one target ecosystem may lead to direct losses in another important ecosystem or its services, represents a widespread problem, especially as availability of intact ecosystems continues to decline. To address this conundrum, we applied recently developed density–habitat relationship models (Van Lanen 2022; Van Lanen et al. 2023a) to evaluate expected changes in songbird density following simulated conifer removal. We then incorporated these changes to songbird density in an optimization framework to predict how different magnitudes of conifer removal may affect Pinyon Jay (Objective 1). Next, we applied our optimization framework to compare expected effects to non-target species under conifer removal efforts designed to maximize single species, multi-species (ecosystem), and multi-ecosystem outcomes (Objective 2). Lastly, we identified areas where conifer removal could be conducted to increasingly offset negative effects to the Pinyon Jay (Objective 3). Our approach represents a generalizable framework for land managers in which sites for conservation action can be selected while transparently and quantitatively balancing disparate conservation objectives.
Methods
Study area
We conducted our study throughout Sage-Grouse Priority Areas for Conservation (PACs; U.S. Fish and Wildlife Service 2013) within the Utah portion of the Southern Rockies/Colorado Plateau Bird Conservation Region (BCR16) (United States North American Bird Conservation Initiative Committee 2000) in the western United States (Fig. 1). The Utah portion of BCR16 supports 14,763 km2 of PACs with elevations ranging from 1440 to 3516 m. Land within our study area is managed primarily by federal (55.96%) and private (32.21%) entities. We selected this geographical region due to overlapping conifer woodland and sagebrush habitats, on-going conifer removal activities, and population declines of multiple pinyon–juniper associated songbird species within the region. We limited our inference to Sage-Grouse PACs because managers indicated conifer removal, to benefit sagebrush-associated species, is preferentially conducted within PAC polygons (oral comm. Renee Chi, Megan McLachlan, and Michelle Crist of U.S. Bureau of Land Management, 2021).

Location of Greater Sage-Grouse priority areas for conservation (PACs) within the Utah portion of Bird Conservation Region 16 (Southern Rockies/Colorado Plateau) and pinyon–juniper habitat present within PACs. Base layers were modified from LANDFIRE Existing Vegetation Type (pinyon–juniper cover layer, modified from LANDFIRE 2016a); National Weather Service, 1:2,000,000, 1980 (state polygons); United States Fish and Wildlife Service 2015 Status Review (PAC polygons); and from Bird Studies Canada and NABCI, 2014 (Bird Conservation Region Boundaries) digital data
Data inputs
We developed a systematic conservation planning framework based upon conservation features, planning units, cost, and budget data. Conservation features represent components the user wishes to conserve (e.g., spatially explicit species density), planning units represent potential sites for conservation action, cost information represents spatially-explicit conservation action costs, and budget data represent the project budget and/or the desired conservation outcome (Watts et al. 2009; Hanson et al. 2021) (Fig. 2).

Representation of workflow to develop input data, run optimization problems, and quantify predicted species-specific responses to conifer removal at prioritized sites based upon management paradigms (“Optimization Problems”). Songbird species included Bewick’s Wren (BEWR), Brewer’s Sparrow (BRSP), Gray Flycatcher (GRFL), Pinyon Jay (PIJA), Sagebrush Sparrow (SABS), and Sage Thrasher (SATH). Prioritization efforts included sites within priority areas for conservation (PACs) in the Utah portion of Bird Conservation Region 16 (BCR16). Planning units represented areas with pinyon–juniper habitat (PJ [0,1]) based upon a reclassified binary pinyon–juniper layer developed using LandFire 2.0.0 Existing Vegetation Type product (LANDFIRE 2016b). Van Lanen et al. models were developed and described by Van Lanen et al. (2022)
Conservation features
We applied recently-developed density–habitat models (Van Lanen 2022; Van Lanen et al. 2023a) to develop conservation features for Brewer’s Sparrow (Spizella breweri), Sagebrush Sparrow (Artemisiospiza nevadensis), Sage Thrasher (Oreoscoptes montanus), Bewick’s Wren (Thryomanes bewickii), Gray Flycatcher (Empidonax wrightii), and Pinyon Jay. These hierarchical density–habitat models estimate species-specific density relationships as a function of topographic (ruggedness and elevation), anthropogenic (linear and point disturbance), climatic (Palmer Drought Severity Index, Normalized Difference Vegetation Index), and landcover (amount of cropland, sagebrush, pinyon–juniper, annual herbaceous, perennial herbaceous, litter) conditions on the landscape while accounting for spatial scales of selection (Frishkoff et al. 2019). The density–habitat models we used indicate Brewer’s Sparrow, Sagebrush Sparrow, and Sage Thrasher are negatively associated with pinyon–juniper cover and Bewick’s Wren, Gray Flycatcher, and Pinyon Jay are positively associated with pinyon–juniper cover (Fig. 3).

Point estimates (dots) and associated 95% credible intervals (whiskers) for Bayesian hierarchical model parameters associated with the influence of pinyon–juniper cover on songbird densities in the Western United States of America; 2008–2020. Density–habitat relationships are adapted from Van Lanen et al. (2022, 2023a)
We included Brewer’s Sparrow, Sagebrush Sparrow, and Sage Thrasher to represent three sagebrush-associated species within our study. Brewer’s Sparrow are often classified as sagebrush-obligate species and frequently represent the most abundant species within sagebrush ecosystems (Rotenberry et al. 2020). Long-term trend estimation indicates Brewer’s Sparrow may be declining within BCR 16. Density distribution modeling of Sagebrush Sparrow indicates they are an uncommon, but potentially locally abundant, species. They are believed to be closely associated with big sagebrush (Artemisia tridentata) throughout most of their range (Martin and Carlson 2020), however, some investigations indicate their densities may be higher in areas with low sagebrush cover and ample bare ground (Timmer et al. 2019; Van Lanen 2022). Long-term Sagebrush Sparrow population trends appear to be slightly negative or stable within BCR 16 (Sauer et al. 2020). Sage Thrasher are thought to be strongly associated with both sagebrush cover and ecosystems (Reynolds et al. 2020; Van Lanen 2022). Sage Thrasher populations appear stable within BCR 16 from 2008 to 2020 (Van Lanen 2022) but there is some evidence of long-term population declines (Sauer et al. 2020). All three of the sagebrush-associated species have been identified for conservation action within our study area (Partners in Flight 2021).
Similarly, our inclusion of Bewick’s Wren, Gray Flycatcher, and Pinyon Jay served to represent pinyon–juniper associated species within our modeling efforts. Bewick’s Wren are frequently associated with brushy scrub within open country, open woodlands (including pinyon–juniper), and chaparral (Kennedy and White 2020). Recent density–habitat relationship modeling indicates Bewick’s Wren may occur at high densities within all successional phases of pinyon–juniper woodlands in the InterMountain West and their densities tend to increase with pinyon–juniper cover (Van Lanen 2022). Bewick’s Wren populations appear to be increasing within BCR 16 over both the long- (Sauer et al. 2020) and short-term (Van Lanen 2022). Gray Flycatcher are associated with semi-arid woodlands and shrublands of the InterMountain West (Schlossberg and Sterling 2020). Recent density–habitat relationships indicate Gray Flycatcher may occur at high densities within early successional pinyon–juniper woodlands (Van Lanen 2022). Long-term Gray Flycatcher population trends within BCR 16 appear stable (Sauer et al. 2020) but there is evidence of more recent population declines (Van Lanen 2022). As previously mentioned, Pinyon Jay are frequently associated with early successional pinyon–juniper woodlands (Van Lanen et al. 2023a) and have been exhibiting both short- (Van Lanen et al. 2023a) and long-term (Sauer et al. 2020) population declines within BCR 16.
We developed our conservation features by estimating species-specific responses to simulated conifer removal treatments. Doing so allowed the optimization effort to preferentially select areas for treatments likely to result in the largest population increases or smallest population decreases for each species. First, we acquired previously developed 30-m resolution raster layers, representing median posterior model predictions of species densities, throughout the study area (Van Lanen et al. 2023b). These density layers represented predicted densities given 2020 land cover and environmental conditions, summarized at the spatial scale of selection for each covariate and species combination (“baseline layers”; Fig. 2). We also developed a new set of 30-m resolution raster layers of predicted median density, following procedures described by Van Lanen et al. (2023a; hereafter, “conifer removal layers”), in which we simulated complete conifer removal across the landscape. We developed these conifer removal layers by setting the percent of pinyon–juniper cover to zero while retaining all other covariate values used in the original density prediction mapping effort for each species (Fig. 2; Supplemental S1). Next, we subtracted the baseline layer from the conifer removal layer to derive a raster layer representing median expected change in avian density following conifer removal (hereafter, “change layer”) for each species (Fig. 2). Natural history varied widely among target species and, as a result, some species occur at much higher densities than others within our study area. To address this, we normalized the change layer between 0 and 1 for each species using the equation
where zi represents the normalized value for the ith pixel, xi represents the value (i.e., the change in density for a given species) for pixel i within the change layer, min(x) represents the minimum change layer value, and max(x) represents the maximum change layer value. The resulting raster layer for each species represented the normalized median change in avian density (hereafter, “conservation feature”). By normalizing the conservation features, we valued changes to each species’ population in our nine prioritization scenarios (hereafter, “problems”) equally and in proportion to shifts in density for each species, by default. In our application, high conservation feature values represent pixels where densities are expected to increase the most for sagebrush-associated species and decrease the least for the pinyon–juniper associated species.
Planning units
We developed a binary pinyon–juniper raster layer with 30-m resolution from the most recent LANDFIRE existing vegetation type layer (LANDFIRE 2016b), using procedures described in Van Lanen et al. (2023a) and masked this layer using the Sage-Grouse PAC polygons. Pixels within the resulting pinyon–juniper raster layer served as our planning units and represented areas thought to support pinyon–juniper habitat, where conifer removal treatments may realistically be considered within our study area.
Cost information
Since available monetary costs of conifer removal projects were highly variable (Utah Department of Natural Resources 2022), we used percent canopy cover values from the most recent LANDFIRE existing vegetation cover (EVC) 30-m resolution raster layer as our cost layer (LANDFIRE 2016a). We masked this layer using the planning unit layer to generate a final cost layer (hereafter, “canopy cover”) containing pixel values between 1 and 100, where pinyon–juniper woodlands exist. Our use of the existing canopy cost layer reflected our suspicion that the cost of conifer removal would increase with the amount of tree cover present in a planning unit.
Optimization problem design
We designed nine prioritization problems to address our objectives of (1) evaluating how different magnitudes of conifer removal in this region will affect Pinyon Jay abundance; (2) simulating resulting effects to songbird densities from conifer removal conducted under three management paradigms; and (3) developing a prioritization framework to select sites for conifer removal that balances trade-offs for sagebrush and pinyon–juniper associated species with particular attention to the Pinyon Jay (Table 1). To evaluate Objective 1, we designed three separate but similar problems. For each of the three problems we input the canopy cover cost layer and used only the Pinyon Jay conservation feature. We aimed to mimic three levels of pinyon–juniper management, simulating removal of 3%, 6%, and 9% of pinyon juniper in the landscape, respectively. It has been suggested that juniper expansion in portions of the Sage-Grouse range is proceeding at a rate of 0.4–1.6% annually (Reinhardt et al. 2020). Thus, the removal of 3, 6, and 9% of conifer cover represents a magnitude of restoration which would keep pace with conifer expansion over approximately a 3-to-10-year timeframe. We summed the values of all canopy cover pixels within our study area and multiplied that value by 3, 6, and 9% to obtain a value of cover to use as our budget for each problem. We titled these problems Single species: PIJA (3%), Single species: PIJA (6%), and Single species: PIJA (9%) (Table 1).
Next, we developed four problems to explore responses of each of our six species to conifer management based upon single species, ecosystem, and multi-ecosystem management paradigms (Objective 2). For consistency across each of these four problems we fixed the budget at 6% conifer removal within our study area. We included the canopy cover cost layer for all four of these problems and varied the conservation features to reflect desired outcomes under each paradigm. Under a single species management paradigm, conservation action is frequently conducted to maximize the benefits for a single species. To reflect this, we incorporated only the Brewer’s Sparrow density conservation feature to simulate the single species management paradigm. We felt this was reasonable because Brewer’s Sparrow is a species of conservation concern in the region (Partners in Flight 2021), a sagebrush-associated species that uses similar habitats as Sage-Grouse (Timmer et al. 2019), and prior research has shown that 85% of conifer removal conducted for Sage-Grouse recovery has occurred within regions supporting high to moderate Brewer’s Sparrow abundance (Donnelly et al. 2017). Hereafter, we refer to this problem and the associated solution as Single species: BRSP. To simulate conifer management under a sagebrush ecosystem approach, we input conservation features for Brewer’s Sparrow, Sagebrush Sparrow, and Sage Thrasher (hereafter; Sagebrush ecosystem). We simulated conifer management under a pinyon–juniper ecosystem approach using conservation features for Bewick’s Wren, Gray Flycatcher, and Pinyon Jay to investigate species outcomes while prioritizing minimal effects to pinyon–juniper populations (hereafter; Pinyon–juniper ecosystem). Lastly, to simulate conifer management under a multi-ecosystem approach we input the conservation features for all six of our study species (hereafter; Multi-ecosystem) (Table 1). We recognize our use of “ecosystem” and “multi-ecosystem” fails to fully encapsulate all elements of ecosystem function, however, our use of these terms is representative of how management of entire ecosystems is often informed—by the selection of multiple indicators.
Lastly, we designed two problems that emphasize the mitigation of potential negative effects to the Pinyon Jay while maximizing outcomes for the sagebrush-associated species and all investigated species, respectively. For both these problems we again used the canopy cover cost layer and fixed the budget equal to 6% of the total canopy cover within planning units. In both problems, we weighted the Pinyon Jay conservation feature three times more heavily than all other sagebrush or pinyon–juniper species (i.e., multiplied the Pinyon Jay conservation feature data by three), to ensure Pinyon Jay population consequences were valued equally to the sagebrush species. In the penultimate problem, we included conservation features for the three sagebrush-associated species along with the weighted Pinyon Jay density (hereafter; Sagebrush ecosystem + PIJA weighting). For the final problem, we included conservation features for Bewick’s Wren, Brewer’s Sparrow, Gray Flycatcher, Sagebrush Sparrow, and Sage Thrasher as well as the weighted Pinyon Jay conservation feature (hereafter; Multi-ecosystem + PIJA weighting) (Table 1). Thus, these problems were analogous to the Sagebrush ecosystem and Multi-ecosystem problems described above but with the added weighting of the Pinyon Jay conservation feature. These two problems reflected our desire to provide managers with solutions which would mitigate negative effects to Pinyon Jays while simultaneously managing for populations of multiple species.
Optimization procedures and evaluation
We used prioritizr package’s integer linear programming algorithm (Package version 7.1.1; Hanson et al. 2021) in program R (R Development Core Team 2020) to address these conservation problems. We employed a commercial solver (Gurobi Optimization 2022) to facilitate faster analysis, given the large number of planning units (Schuster et al. 2020; Duchardt et al. 2021). We selected the “maximum utility objective” to solve all problems (Hanson et al. 2021), which seeks to maximize the total value of the summed conservation features while attempting to minimize cost and stay within the budget amount. We included a boundary penalty of 0.2 and used the default edge factor of 0.5 to aid in clustering selected parcels (Hanson et al. 2021) because we wanted to ease potential on-the-ground implementation and develop clear project areas for land managers. We selected the boundary penalty of 0.2 because initial tests with this penalty resulted in slightly aggregated solutions, represented by combinations of planning units selected to receive conifer removal treatments, in our application which is more representative of actual conifer treatments. In contrast, we found boundary penalties < 0.2 resulted in exceedingly dispersed solutions and higher values resulted in a single cohesive treatment area. We allowed an optimality gap of 0.01 for all solutions, to reduce processing time (Hanson et al. 2021).
We used the solutions from each optimization problem to identify sites for conifer removal. We modified the binary pinyon–juniper layer to simulate removal at the planning unit pixels selected in each problem and recreated the moving window rasters of pinyon–juniper cover for each species. We then predicted avian densities for each species using the new pinyon–juniper moving window raster layers for each of the nine solutions following procedures described by Van Lanen et al. (2023a). We then recalculated predicted abundances for each species and calculated median predicted densities and 75% credible intervals (CrI) pertaining to predicted density for each species and scenario (for details see Supplemental Material S1). Next, we calculated changes in density for each solution by subtracting the baseline pixel values from the new predicted density values (Fig. 2). We converted predicted density values for this layer to abundance (number of individuals present) by multiplying densities (in 1-km2) by 0.0009, which represents the fraction of 1 km2 represented by a 30-m resolution pixel. Finally, we summed the pixel values for this layer to obtain the change in the number of expected individuals resulting from conifer removal at locations selected for each solution.
Results
The optimal solutions for the Pinyon Jay problems with pinyon–juniper canopy cover reduction budgets of 3%, 6%, and 9% (Objective 1) resulted in removal of pinyon–juniper from disproportionately larger numbers of planning units (Table 2). These solutions corresponded to reductions in overall conifer cover of 3.1%, 6.2%, and 9.4%, respectively. The mean canopy cover within selected planning units declined as the problem budget increased (Table 2). Conifer removal resulted in losses of 0.31% (Single species: PIJA [3%]) to 1.18% (Single species: PIJA [9%]) of the Pinyon Jay population within the study area (Fig. 4) and losses per km2 of canopy cover treated increased with increasing budgets (Fig. 5).

The median predicted percent change in overall population size following simulated pinyon–juniper removal treatments at prioritized locations within Sage-Grouse priority areas for conservation (PACs) in the Utah portion of Bird Conservation Region 16. The total expected number of individuals gained or lost if conifer removal was conducted at all sites selected in the solution for each optimization problem is shown at the margin of each bar. Simulated pinyon–juniper removal was targeted to optimize changes in Pinyon Jay density (PIJA; “Single Species: PIJA” problem) with 3%, 6% and 9% reductions in overall canopy cover. Pinyon–juniper removal was targeted to optimize changes in densities of Brewer’s Sparrow (BRSP; “Single Species: BRSP” problem); Brewer’s Sparrow, Sagebrush Sparrow (SABS) and Sage Thrasher (SATH; “Sagebrush Ecosystem” problem); and Brewer’s Sparrow, Sagebrush Sparrow, Sage Thrasher, Bewick’s Wren (BEWR), Gray Flycatcher (GRFL), and Pinyon Jay (“Multi-ecosystem” problem). Pinyon–juniper removal was targeted to optimize the same conservation features as the “Sagebrush Ecosystem” and “Multi-Ecosystem” problems while including and weighting the Pinyon Jay feature three times more heavily than other conservation features in the “Sagebrush Ecosystem + PIJA Weighted” and “Multi-ecosystem + PIJA Weighted” problems

Predicted change in the median number of individuals for each 1km2 of pinyon–juniper canopy cover removed at prioritized locations within Sage-Grouse priority areas for conservation (PACs) in the Utah portion of Bird Conservation Region 16. The expected number of individuals gained or lost per 1km2 of pinyon–juniper cover removed in each solution is shown at the margin of each bar. Simulated pinyon–juniper removal was targeted to optimize changes in Pinyon Jay density (PIJA; “Single Species: PIJA” problem) with 3%, 6% and 9% reductions in overall canopy cover. Pinyon–juniper removal was targeted to optimize changes in densities of Brewer’s Sparrow (BRSP; “Single Species: BRSP” problem); Brewer’s Sparrow, Sagebrush Sparrow (SABS) and Sage Thrasher (SATH; “Sagebrush Ecosystem” problem); and Brewer’s Sparrow, Sagebrush Sparrow, Sage Thrasher, Bewick’s Wren (BEWR), Gray Flycatcher (GRFL), and Pinyon Jay (“Multi-ecosystem” problem). Pinyon–juniper removal was targeted to optimize the same conservation features as the “Sagebrush Ecosystem” and “Multi-Ecosystem” problems, while including and weighting the Pinyon Jay feature three times more heavily than other conservation features in the “Sagebrush Ecosystem + PIJA Weighted” and “Multi-ecosystem + PIJA Weighted” problems
For Objective 2 we applied our optimization framework to compare expected effects to non-target species under conifer removal efforts designed to maximize single species, multi-species (ecosystem), and multi-ecosystem outcomes. Results from problems evaluating species-specific responses to four management scenarios (Single species: BRSP, Sagebrush ecosystem, Pinyon–juniper ecosystem, and Multi-ecosystem) generally yielded increasing numbers of selected planning units as the number of conservation features increased, given the same budget (Table 2). Correspondingly, mean canopy cover values of selected planning units decreased with increasing numbers of conservation features (Table 2). Net population responses for the four problems examined in Objective 2 followed similar patterns for all three pinyon–juniper associated species. Bewick’s Wren, Gray Flycatcher, and Pinyon Jay all experienced the smallest population reductions from conifer removal based upon the Pinyon–juniper ecosystem solution followed, in order, by the Multi-ecosystem, Sagebrush ecosystem, and Single species: BRSP solutions (Fig. 4). Brewer’s Sparrow realized the largest net population benefits from the Sagebrush ecosystem solution, followed by the Single species: BRSP, Multi-ecosystem, and Pinyon–juniper ecosystem solutions (Fig. 4). In contrast, Sagebrush Sparrow and Sage Thrasher experienced the largest net population increases in the Multi-ecosystem solution, followed by the Sagebrush ecosystem, Pinyon–juniper ecosystem, and Single species: BRSP solutions (Fig. 4). When comparing the solutions to the four problems addressing Objective 2, all three of the pinyon–juniper associated species exhibited the smallest reductions in the number of individuals per 1 km2 of pinyon–juniper removed in the Pinyon–juniper ecosystem solution, followed by the Multi-ecosystem, Sagebrush ecosystem, and Single species: BRSP solutions (Fig. 5). For solutions associated with Objective 2, Brewer’s Sparrow experienced the largest increases in individuals, per 1 km2 of pinyon–juniper removed, from the Single species: BRSP solution followed by the Sagebrush ecosystem, Multi-ecosystem, and Pinyon–juniper ecosystem solutions (Fig. 5). For Objective 2, both Sagebrush Sparrow and Sage Thrasher experienced the largest increases in the number of individuals per 1 km2 of pinyon–juniper removed from the Sagebrush ecosystem solution, followed by the Multi-ecosystem, Pinyon–juniper ecosystem, and Single species: BRSP solutions (Fig. 5).
Solutions weighting Pinyon Jay outcomes more heavily (Objective 3) resulted in relatively large numbers of selected planning units which generally had lower mean conifer cover values compared to the single species solutions (Table 2). Of the two problems designed to mitigate negative effects on the Pinyon Jay (Sagebrush ecosystem + PIJA Weighted and Multi-ecosystem + PIJA Weighted), more Bewick’s Wren and Gray Flycatcher individuals were lost, overall and per unit conifer removed, in the Sagebrush ecosystem + PIJA Weighted solution compared to the Multi-ecosystem + PIJA Weighted solution. In contrast, the number of Pinyon Jay individuals lost, overall and per unit of conifer removed, was lower in the Sagebrush ecosystem + PIJA Weighted solution (Fig. 4) compared to the Multi-ecosystem + PIJA Weighted solution. All three sagebrush-associated species experienced larger net and per unit conifer removed population increases from the Sagebrush ecosystem + PIJA Weighted solution compared to the Multi-ecosystem + PIJA Weighted solution (Fig. 4).
We provide a ranked consequences table using the outcomes for each species, per 1 km2 canopy cover removed (Supplemental Table S1.1), to compare species-specific and overall outcomes across solutions. In this table, lower values indicate the solution resulted in larger populations compared to solutions with higher values. Of the solutions we generated, the Sagebrush ecosystem + PIJA weighting solution ranked the highest when considering the summed species ranks (15) and the Single species: BRSP solution ranked the lowest summed species ranks (31), when considering outcomes for all species. There was little difference in the total rank value of the other seven solutions, with values ranging from 19 to 21.
Discussion
Despite growing concern regarding the effects of single species management on non-target species (Zipkin et al. 2010; White et al. 2013; Bombaci and Pejchar 2016; Gallo and Pejchar 2016), non-game wildlife management within the United States is largely focused on recovering listed species, and preventing new listings, under the Endangered Species Act. For instance, the Natural Resource Conservation Service’s Working Lands For Wildlife Program states “Individual species are used as barometers for healthy, functioning landscapes…” (United States Department of Agriculture (USDA) Natural Resources Conservation Service 2023) and the U.S. Fish and Wildlife Service received more than $43 million dollars for fiscal year 2023 in support of the Endangered Species Program (United States Department of Interior 2022), indicating single species management remains the modus operandi among some federal agencies within the United States. Our results indicate management for a single target species, although cost effective for that species, may result in poor multi-species outcomes compared to management aimed at benefiting suites of species. In our study, the Single species: BRSP solution resulted in the worst net and proportional outcomes (per km2 of canopy cover treated) for all five other species we investigated. Gray Flycatcher, a species declining within our study area (Van Lanen 2022), experienced an estimated 5% reduction in the total regional population, compared to just a 2.7% increase in the overall Brewer’s Sparrow population under the Single species: BRSP solution; illustrating target species benefits may be outweighed by trade-offs to non-target species. Such instances are concerning given the growing number of declining bird species in North America (Rosenberg et al. 2019). Considerations regarding effects on non-target species may be particularly important when ranges of declining non-target species largely overlap target species, as in the case we highlight. We suspect substantial range overlap between declining species with disparate habitat requirements will become more frequent as the area of undisturbed ecosystems continues to contract (Theobald et al. 2020) and the number of declining species continues to increase (Butchart et al. 2010).
Our simulations of conifer removal illustrated the negative consequences for pinyon–juniper associated species, even when sites were selected to avoid unwanted effects (as in the Pinyon–juniper ecosystem solution), supporting experimental findings of songbird response to such management actions. A study in central Colorado found occupancy of Gray Flycatcher and Pinyon Jay was negatively affected by conifer removal treatments (Magee et al. 2019), while a separate study in Oregon found Gray Flycatcher occurred on approximately 30% fewer point count stations in treated sites compared to untreated sites (Holmes et al. 2017). Thus, both experimental findings and model-based inference indicate considerable negative effects to these declining species resulting from pinyon–juniper management. The different configurations of treatments we investigated indicate some negative effects may be reduced by incorporating current distributions and predicted responses of species to treatments in a prioritization framework.
Previous work, based upon observational data, indicates Pinyon Jay density is influenced by pinyon–juniper cover at large spatial extents (Van Lanen et al. 2023a). Thus, it will likely be difficult to conduct conifer removal while completely mitigating detrimental effects to jay populations where they occur. Research indicates the removal of ~ 1340 km2 of conifer cover (assuming 1% annual increase in pinyon–juniper cover) is needed to retain the current footprint of sagebrush habitat (Reinhardt et al. 2020). Despite these expansions of pinyon–juniper woodlands in North America, Pinyon Jay populations are continuing to decline (Sauer et al. 2020; Van Lanen et al. 2023a), indicating that infilling or succession of early-successional woodlands may be degrading Pinyon Jay habitat quality. Our work indicates it would be difficult to conduct this scale of restoration in regions where Pinyon Jay are present without substantial losses to the species. The limited ability of our prioritization efforts to mitigate these effects was partially the result of our study area (selected because of specific interest from management stakeholders), which is predicted to support relatively high densities of Pinyon Jays. It remains possible that a more coordinated prioritization approach, across a larger spatial extent, could yield greater multi-species conservation outcomes and further reduce negative effects on declining pinyon–juniper associated species. Thus, a large-scale prioritization effort to evaluate where conifer removal could be conducted at broader jurisdictional levels, rather than prioritizing sites within states or BCRs (e.g., Utah portion of BCR16), may better guide conservation. That said, developing conservation priorities across nested hierarchical scales (O’Donnell et al. 2022) could result in better coordinated management applications capable of improving the efficiency and effectiveness of pinyon–juniper removals for both target and non-target species.
Our results indicate targeted restoration to minimize negative effects to non-target species (Pinyon–juniper ecosystem solution) resulted in better outcomes for some sagebrush-associated species compared to the Single species: BRSP scenario (Figs. 4, 5). This indicates that prioritizations aimed at benefiting a single sagebrush-associated species may not effectively prioritize the needs of all sagebrush associated songbirds and, instead, could result in a potential opportunity cost. We recognize that current management strategies are largely focused on Sage-Grouse, not Brewer’s Sparrow. Although Sage-Grouse and Brewer’s Sparrow have been shown to use similar habitats (Donnelly et al. 2017; Timmer et al. 2019), additional efforts to evaluate songbird response to conifer management, prioritized to achieve Sage-Grouse conservation, would better assess the actual cost of single species management in this system.
Spatial heterogeneity is an important driver of ecological processes at landscape scales (Turner 2005) and represents an important aspect of wildlife management to satisfy species’ requirements throughout daily, seasonal, and annual cycles (Law and Dickman 1998; Davis et al. 2020). Our results indicate that management to benefit a suite of sagebrush-associated species will result in restoration occurring across a wider range of resource conditions than management targeted to benefit a single species (Brewer’s Sparrow, in our application). For instance, the density–habitat relationships developed by Van Lanen (2022) indicate Brewer’s Sparrow occurs in regions with intermediate Normalized Difference Vegetation Index (NDVI) values while Sage Thrasher occurs at higher densities at sites with low NDVI. Thus, the solution from our Sagebrush ecosystem problem likely resulted in selected sites representing a range of environmental characteristics, contributing to a more heterogeneous landscape, compared to management for Brewer’s Sparrow alone.
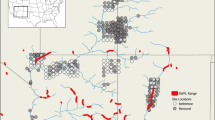
Species-specific population outcomes resulting from our solutions demonstrated the inherent trade-offs when managing for species with disparate requirements. Management prioritized to reduce effects to Pinyon Jay resulted in the best outcomes for Pinyon Jay. The same pattern was true in the case of the Brewer’s Sparrow, with the Single species: BRSP solution resulting in the best per-unit-effort results for Brewer’s Sparrow. The mapped solutions in Fig. 6 (see also Fig. S3.1 for solutions throughout the study area) specifically illustrate how the inclusion of Pinyon Jay outcomes (as in the Sagebrush ecosystem + PIJA Weighted solution) pushes selected sites away from regions currently predicted to support high densities of Pinyon Jays, as in the southwestern portion of our study area. This pattern in trade-offs extends to the ecosystem solutions, with the Sagebrush ecosystem solution yielding more Brewer’s Sparrow, Sagebrush Sparrow, and Sage Thrasher individuals than the Pinyon–juniper ecosystem solution. Conversely, the Pinyon–juniper ecosystem solution yielded more Bewick’s Wren, Gray Flycatcher, and Pinyon Jay individuals than the Sagebrush ecosystem solution. Thus, optimal management is almost certain to depend upon specific management objectives.

Predicted median densities for Brewer’s Sparrow (a, b) and Pinyon Jay (c, d) in relation to sites prioritized for pinyon–juniper removal treatments (PJ Removal Sites, shown in red) within southwest Utah. Removal sites were selected using an integer linear programming optimization solver designed to maximize species-specific outcomes for Brewer’s Sparrow (a, c) or for three sagebrush-associated species (Brewer’s Sparrow, Sagebrush Sparrow, and Sage Thrasher) and triple-weighted Pinyon Jay outcomes (b, d)
The enormous proliferation of species distribution and/or niche models provides ample products that can be integrated into conservation decision-making (Guisan et al. 2013). Although our approach suffered from several limitations [including an inability to explicitly incorporate uncertainty, using models informed by observational data, and merging prioritization efforts with models which incorporate spatial scales of selection (Supplemental S2)], we encourage regional land managers to consider these niche modeling products when prioritizing regions for conservation action. Density-distribution maps indicate southern Utah supports high densities of Pinyon Jay and Gray Flycatcher, while areas in Wyoming are strongholds for Brewer’s Sparrow (Van Lanen 2022; Van Lanen et al. 2023a). Such information raises the question of whether management in southern Utah should be conducted to support pinyon–juniper communities rather than the current effort in the region to restore sagebrush-associated communities. A large-scale prioritization effort, as previously suggested, could help evaluate where conifer removal may result in the largest multi-species benefits.
We suspect the luxury of managing to recover one species without consideration of non-target species will become increasingly rare, given the rising number of at-risk species (Butchart et al. 2004, 2010). Our case study serves as a cautionary example for managers implementing restoration for single species. We urge managers to consider effects to non-target species, particularly when data on the distribution and abundance of non-target species are available. Past global conservation efforts, which focused on identifying and protecting regions representing biodiversity hotspots (Wilson et al. 2006), may ultimately contribute to even more inherent trade-offs among species given the sheer number of species within protected regions. Management within these protected places will be hard-pressed to positively influence the full suite of species inhabiting these hotspots and difficult decisions will need to be made.
Ultimately, land management decisions address specific objectives and societal values. We provide an example of how decisions using model-based inference and optimization techniques can better meet well-defined objectives in a transparent and equitable way. We also demonstrate how weighting of conservation features can be incorporated to explicitly reflect stakeholder values and preferences. Our approach is transferrable for use across a wide variety of taxa, ecosystems, and conservation applications. Despite some limitations, transparent tools like we describe, may help build trust and result in more equitable solutions among stakeholders (Law et al. 2018) when trade-offs are unavoidable. Results from frameworks such as ours can be used to allocate resources for conservation action at regional and local scales, identify local-scale project areas for restoration, and inform environmental impact assessments for proposed projects.
Data availability
The baseline predicted songbird density layers are available via two U.S. Geological Survey data releases (Van Lanen et al. 2022, 2023b). Raster layers of the selected planning units under the scenarios we investigated in this manuscript are available via a U.S. Geological Survey data release at https://doi.org/10.5066/P9QFTK1D (Van Lanen et al. 2023c).
References
Allen CR, Cumming GS, Garmestani AS, Taylor PD, Walker BH (2011) Managing for resilience. Wildl Biol 17:337–349
Askins RA (2001) Sustaining biological diversity in early successional communities: the challenge of managing unpopular habitats. Wildl Soc Bull 29:407–412
Ball IR, Possingham HP, Watts ME (2009) Marxan and relatives: software for spatial conservation prioritization. In: Moilanen A, Wilson KA, Possingham HP (eds) Chapter: pages 185–195 in spatial conservation prioritization: quantitative methods and computational tools. Oxford University Press, Oxford
Baruch-Mordo S, Evans JS, Severson JP, Naugle DE, Maestas JD, Kiesecker JM, Falkowski MJ, Hagen CA, Reese KP (2013) Saving sage-grouse from the trees: a proactive solution to reducing a key threat to a candidate species. Biol Conserv 167:233–241
Bombaci S, Pejchar L (2016) Consequences of pinyon and juniper woodland reduction for wildlife in North America. For Ecol Manag 365:34–50
Boone JD, Witt C, Ammon EM (2021) Behavior-specific occurrence patterns of Pinyon jays (Gymnorhinus cyanocephalus) in three Great Basin study areas and significance for pinyon–juniper woodland management. PLoS ONE 16:e0237621
Butchart SH, Stattersfield AJ, Bennun LA, Shutes SM, Akcakaya HR, Baillie JE, Stuart SN, Hilton-Taylor C, Mace GM (2004) Measuring global trends in the status of biodiversity: red list indices for birds. PLoS Biol 2:e383
Butchart SHM, Walpole M, Collen B, van Strien A, Scharlemann JPW, Almond REA, Baillie JEM, Bomhard B, Brown C, Bruno J, Carpenter KE, Carr GM, Chanson J, Chenery AM, Csirke J, Davidson NC, Dentener F, Foster M, Galli A, Galloway JN, Genovesi P, Gregory RD, Hockings M, Kapos V, Lamarque J, Leverington F, Loh J, McGeoch MA, McRae L, Minasyan A, Morcillo MH, Oldfield TEE, Pauly D, Quader S, Revenga C, Sauer JR, Skolnik B, Spear D, Stanwell-Smith D, Stuart SN, Symes A, Tierney M, Tyrrell TD, Vie J, Watson R (2010) Global biodiversity: indicators of recent declines. Science 328:1164–1168
Carlisle JD, Chalfoun AD, Smith KT, Beck JL (2018) Nontarget effects on songbirds from habitat manipulation for greater sage-grouse: implications for the umbrella species concept. Condor 120:439–455
Caro TM, O’Doherty G (1999) On the use of surrogate species in conservation biology. Conserv Biol 13:805–814
Coates PS, Prochazka BG, Ricca MA, Gustafson KB, Ziegler P, Casazza ML (2017) Pinyon and juniper encroachment into sagebrush ecosystems impacts distribution and survival of greater sage-grouse. Rangel Ecol Manag 70:25–38
Cook AA, Messmer TA, Guttery MR (2017) Greater sage-grouse use of mechanical conifer reduction treatments in northwest Utah. Wildl Soc Bull 41:27–33
Davis KP, Augustine DJ, Monroe AP, Derner JD, Aldridge CL (2020) Adaptive rangeland management benefits grassland birds utilizing opposing vegetation structure in the shortgrass steppe. Ecol Appl 30:e02020
Defenders of Wildlife (2022) Petition to list the pinyon jay (Gymnorhinus cyanocephalus) as endangered or Threatened Under the Endangered Species Act
DeFries R, Nagendra H (2017) Ecosystem management as a wicked problem. Science 356:265–270
Donnelly JP, Tack JD, Doherty KE, Naugle DE, Allred BW, Dreitz VJ (2017) Extending conifer removal and landscape protection strategies from sage-grouse to songbirds, a range-wide assessment. Rangel Ecol Manag 70:95–105
Duchardt CJ, Monroe AP, Heinrichs JA, O’Donnell MS, Edmunds DR, Aldridge CL (2021) Prioritizing restoration areas to conserve multiple sagebrush-associated wildlife species. Biol Conserv 260:109212
Frishkoff LO, Mahler DL, Fortin MJ (2019) Integrating over uncertainty in spatial scale of response within multispecies occupancy models yields more accurate assessments of community composition. Ecography 42:2132–2143
Gallo T, Pejchar L (2016) Improving habitat for game animals has mixed consequences for biodiversity conservation. Biol Conserv 197:47–52
Guisan A, Tingley R, Baumgartner JB, Naujokaitis-Lewis I, Sutcliffe PR, Tulloch AI, Regan TJ, Brotons L, McDonald-Madden E, Mantyka-Pringle C, Martin TG, Rhodes JR, Maggini R, Setterfield SA, Elith J, Schwartz MW, Wintle BA, Broennimann O, Austin M, Ferrier S, Kearney MR, Possingham HP, Buckley YM (2013) Predicting species distributions for conservation decisions. Ecol Lett 16:1424–1435
Gurobi Optimization LLC (2022) Gurobi optimizer reference manual. https://www.gurobi.com
Haddad NM, Brudvig LA, Clobert J, Davies KF, Gonzalez A, Holt RD, Lovejoy TE, Sexton JO, Austin M, Collins CD, Cook WM, Damschen EI, Ewers RM, Foster BL, Jenkins CN, King AJ, Laurance WF, Levey DJ, Margules CR, Melbourne BA, Nicholls AO, Orrock JL, Song D, Townshend JR (2015) Habitat fragmentation and its lasting impact on earth’s ecosystems. Sci Adv 1:e1500052
Hanson JO, Schuster R, Morrell N, Strimas-Mackey M, Edwards BPM, Watts ME, Arcese P, Bennett J, Possingham HP (2021) Prioritizr: systematic conservation prioritization in R, package version 7.1.1
Harvey E, Gounand I, Ward CL, Altermatt F, Cadotte M (2017) Bridging ecology and conservation: from ecological networks to ecosystem function. J Appl Ecol 54:371–379
Holmes AL, Maestas JD, Naugle DE (2017) Bird responses to removal of western juniper in sagebrush-steppe. Rangel Ecol Manag 70:87–94
Johnson K, Balda RP (2020) Pinyon jay (Gymnorhinus cyanocephalus), version 2.0. In: Rodewald PG, Keeney BK (eds) Birds of the world. Cornell Lab of Ornithology, Ithaca. https://doi.org/10.2173/bow.pinjay.2102
Kennedy ED, White DW (2020) Bewick’s wren (Thryomanes bewickii), version 1.0. In: Poole AF (ed) Birds of the world. Cornell Lab of Ornithology, Ithaca. https://doi.org/10.2173/bow.bewwre.2101
Kerr JT, Deguise I (2004) Habitat loss and the limits to endangered species recovery. Ecol Lett 7:1163–1169
Lambeck RJ (1997) Focal species: a multi-species umbrella for nature conservation. Conserv Biol 11:849–856
LANDFIRE (2016a) Existing Vegetation Cover, LANDFIRE 2.0.0, U.S. Department of the Interior, Geological Survey, and U.S. Department of Agriculture. https://landfire.gov/version_download.php. Accessed 22 Apr 2021
LANDFIRE (2016b) Existing Vegetation Type, LANDFIRE 2.0.0, U.S. Department of the Interior, Geological Survey, and U.S. Department of Agriculture. https://landfire.gov/version_download.php. Accessed 22 Apr 2021
Law BS, Dickman CR (1998) The use of habitat mosaics by terrestrial vertebrate fauna: implications for conservation and management. Biodivers Conserv 7:323–333
Law EA, Bennett NJ, Ives CD, Friedman R, Davis KJ, Archibald C, Wilson KA (2018) Equity trade-offs in conservation decision making. Conserv Biol 32:294–303
Link JS (2002) Ecological considerations in fisheries management: when does it matter? Fisheries 27:10–17
Magee PA, Coop JD, Ivan JS (2019) Thinning alters avian occupancy in piñon–juniper woodlands. Condor 121:duy008
Martin JW, Carlson BA (2020) Sagebrush sparrow (Artemisiospiza nevadensis), version 1.0. In: Poole AF (ed) Birds of the world. Cornell Lab of Ornithology, Ithaca. https://doi.org/10.2173/bow.sagspa1.01
Miller RF, Chambers JC, Evers L, Williams CJ, Snyder KA, Roundy BA, Pierson FB (2019) The ecology, history, ecohydrology, and management of pinyon and juniper woodlands in the Great Basin and Northern Colorado Plateau of the Western United States
Natural Resource Conservation Service (2015) Outcomes in conservation: Sage Grouse Initiative. NRCS Progress Report, Washington, D.C., p 57
O’Donnell MS, Edmunds DR, Aldridge CL, Heinrichs JA, Monroe AP, Coates PS, Hanser SE, Wiechman LA (2022) Defining biologically relevant and hierarchically nested population units to inform wildlife management. Ecol Evol. https://doi.org/10.1002/ece1003.9565
Partners in Flight (2021) Avian Conservation Assessment Database, version 2021. http://pif.birdconservancy.org/ACAD. Accessed 1 Oct 2021
R Development Core Team (2020) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org
Reinhardt JR, Filippelli S, Falkowski M, Allred B, Maestas JD, Carlson JC, Naugle DE (2020) Quantifying pinyon–juniper reduction within North America’s sagebrush ecosystem. Rangel Ecol Manag 73:420–432
Reynolds TD, Rich TD, Stephens DA (2020) Sage thrasher (Oreoscoptes montanus), version 1.0. In: Poole AF, Gill FB (eds) Birds of the world. Cornell Lab of Ornithology, Ithaca. https://doi.org/10.2173/bow.sagthr.2101
Roberge J-M, Angelstam P (2004) Usefulness of the umbrella species concept as a conservation tool. Conserv Biol 18:76–85
Roberge J-M, Mikusiński G, Svensson S (2008) The white-backed woodpecker: umbrella species for forest conservation planning? Biodivers Conserv 17:2479–2494
Rosenberg KV, Dokter AM, Blancher PJ, Sauer JR, Smith AC, Smith PA, Stanton JC, Panjabi AO, Helft L, Parr M, Marra PP (2019) Decline of the North American avifauna. Science 366:120–124
Rotenberry JT, Patten MA, Preston KL (2020) Brewer’s sparrow (Spizella breweri), version 1.0. In: Poole AF, Gill FB (eds) Birds of the world. Cornell Lab of Ornithology, Ithaca. https://doi.org/10.2173/bow.brespa.2101
Roundy BA, Miller RF, Tausch RJ, Young K, Hulet A, Rau B, Jessop B, Chambers JC (2014a) Understory cover responses to piñon–juniper treatments across tree dominance gradients in the Great Basin. Rangel Ecol Manag 67:482–494
Roundy BA, Young K, Cline N, Hulet A, Miller RF, Tausch RJ, Chambers JC, Rau B (2014b) Piñon–juniper reduction increases soil water availability of the resource growth pool. Rangel Ecol Manag 67:495–505
Sauer JR, Link WA, Hines JE (2020) The North American breeding bird survey, analysis results 1966–2019. U.S. Geological Survey data release. https://doi.org/10.5066/P96A7675
Schlossberg S, Sterling JC (2020) Gray flycatcher (Empidonax wrightii), version 1.0. In: Poole AF (ed) Birds of the World. Cornell Lab of Ornithology, Ithaca. https://doi.org/10.2173/bow.gryfly.2101
Schuster R, Hanson JO, Strimas-Mackey M, Bennett JR (2020) Exact integer linear programming solvers outperform simulated annealing for solving conservation planning problems. PeerJ 8:e9258
Severson JP, Hagen CA, Maestas JD, Naugle DE, Forbes JT, Reese KP (2017a) Effects of conifer expansion on greater sage-grouse nesting habitat selection. J Wildl Manag 81:86–95
Severson JP, Hagen CA, Tack JD, Maestas JD, Naugle DE, Forbes JT, Reese KP (2017b) Better living through conifer removal: a demographic analysis of sage-grouse vital rates. PLoS ONE 12:e0174347
Simberloff D (1998) Flagships, umbrellas, and keystones: is single-species management passe in the landscape era? Biol Conserv 83:247–257
Theobald DM, Kennedy C, Chen B, Oakleaf J, Baruch-Mordo S, Kiesecker J (2020) Earth transformed: detailed mapping of global human modification from 1990 to 2017. Earth Syst Sci Data 12:1953–1972
Tilman D, Clark M, Williams DR, Kimmel K, Polasky S, Packer C (2017) Future threats to biodiversity and pathways to their prevention. Nature 546:73–81
Timmer JM, Aldridge CL, Fernández-Giménez ME (2019) Managing for multiple species: greater sage‐grouse and sagebrush songbirds. J Wildl Manag 83:1043–1056
Turner MG (2005) Landscape ecology: what is the state of the science? Annu Rev Ecol Evol Syst 36:319–344
United States Department of Agriculture (USDA) Natural Resources Conservation Service (2023) Working lands for wildlife. https://www.nrcs.usda.gov/programs-initiatives/working-lands-for-wildlife. Accessed 4 Apr 2023
United States Department of Interior (2022) Budget justifications and performance information fiscal year 2023. U.S. Fish and Wildlife Service, p 480
United States North American Bird Conservation Initiative Committee (2000) Bird conservation regions descriptions: a supplement to the North American Bird Conservation Initiative. Bird conservation regions map. U.S. Fish and Wildlife Service
U.S. Fish and Wildlife Service (2013) Greater sage-grouse (Centrocercus urophasianus) conservation objectives—final report. U.S. Department of the Interior, Fish and Wildlife Service, Denver, p 91
Utah Department of Natural Resources (2022) Utah’s Watershed Restoration Initiative. https://wri.utah.gov/wri/project/search.html. Accessed 24 May 2022
Van Lanen NJ (2022) Songbird trend estimation and density–habitat relationships to inform and prioritize conifer management in the sagebrush and pinyon–juniper ecotone. Dissertation, Colorado State University
Van Lanen NJ, Monroe AP, Aldridge CL (2022) Predicted Pinyon Jay (Gymnorhinus cyanocephalus) densities across the western United States, 2008–2020. U.S. Geological Geological Survey data release. https://doi.org/10.5066/P9NIG4UW
Van Lanen NJ, Monroe AP, Aldridge CL (2023a) A hidden cost of single species management: habitat relationships reveal potential negative effects of conifer removal on a non-target species. Biol Conserv. https://doi.org/10.1016/j.biocon.2023.109959
Van Lanen NJ, Monroe AP, Aldridge CL (2023b) Predicted 2020 densities for 11 songbird species across the western United States. U.S. Geological Survey data release. https://doi.org/10.5066/P9MJHTMQ
Van Lanen NJ, Shyvers JE, Duchardt CJ, Aldridge CL (2023c) Prioritized sites for conifer removal within the Utah portion of bird conservation region 16, 2020. U.S. Geological Survey data release. https://doi.org/10.5066/P9QFTK1D
Wang F, Winkler J, Viña A, McShea WJ, Li S, Connor T, Zhao Z, Wang D, Yang H, Tang Y, Zhang J, Liu J (2021) The hidden risk of using umbrella species as conservation surrogates: a spatio-temporal approach. Biol Conserv 253:108913
Watts ME, Ball IR, Stewart RS, Klein CJ, Wilson K, Steinback C, Lourival R, Kircher L, Possingham HP (2009) Marxan with zones: software for optimal conservation based land- and sea-use zoning. Environ Model Softw 24:1513–1521
White AM, Zipkin EF, Manley PN, Schlesinger MD (2013) Conservation of avian diversity in the Sierra Nevada: moving beyond a single-species management focus. PLoS ONE 8:e63088
Wilson KA, McBride MF, Bode M, Possingham HP (2006) Prioritizing global conservation efforts. Nature 440:337–340
Zeller KA, Cushman SA, Van Lanen NJ, Boone JD, Ammon E (2021) Targeting conifer removal to create an even playing field for birds in the Great Basin. Biol Conserv 257:109130
Zipkin EF, Andrew Royle J, Dawson DK, Bates S (2010) Multi-species occurrence models to evaluate the effects of conservation and management actions. Biol Conserv 143:479–484
Acknowledgements
Our project benefitted from conversations with Brad Udell, Renee Chi, Megan McLachlan, and Michele Crist. The Bird Conservancy of the Rockies, Integrated Monitoring in Bird Conservation Regions partnership, U.S. Bureau of Land Management, Wyoming Landscape Conservation Initiative, and U.S. Geological Survey provided funding. Our manuscript was improved through reviews by Larissa Bailey, Dave Koons, Liba Pejchar, Brad Udell, and an anonymous reviewer. Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the U.S. Government.
Author information
Authors and Affiliations
Contributions
NJVL and CLA conceived the initial research question. NJVL conducted analysis with guidance from all authors. The first manuscript draft was written by NJVL. CJD developed Fig. 2. NJVL developed all other figures. All authors contributed critically to manuscript drafts and gave final approval for publication.
Corresponding author
Ethics declarations
Competing interests
The authors certify they have no competing interests regarding the subject matter or materials discussed in this manuscript.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Van Lanen, N.J., Shyvers, J.E., Duchardt, C.J. et al. A multi-ecosystem prioritization framework to balance competing habitat conservation needs of multiple species in decline. Landsc Ecol 38, 2795–2813 (2023). https://doi.org/10.1007/s10980-023-01712-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10980-023-01712-z