
Abstract
Context
Connectivity between habitat patches is vital for ecological processes at multiple scales. Traditional metrics do not measure the scales at which individual habitat patches contribute to the overall ecological connectivity of the landscape. Connectivity has previously been evaluated at several different scales based on the dispersal capabilities of particular organisms, but these approaches are data-heavy and conditioned on just a few species.
Objectives
Our objective was to improve cross-scale measurement of connectivity by developing and testing a new landscape metric, cross-scale centrality.
Methods
Cross-scale centrality (CSC) integrates over measurements of patch centrality at different scales (hypothetical dispersal distances) to quantify the cross-scale contribution of each individual habitat patch to overall landscape or seascape connectivity. We tested CSC against an independent metapopulation simulation model and demonstrated its potential application in conservation planning by comparison to an alternative approach that used individual dispersal data.
Results
CSC correlated significantly with total patch occupancy across the entire landscape in our metapopulation simulation, while being much faster and easier to calculate. Standard conservation planning software (Marxan) using dispersal data was weaker than CSC at capturing locations with high cross-scale connectivity.
Conclusions
Metrics that measure pattern across multiple scales are much faster and more efficient than full simulation models and more rigorous and interpretable than ad hoc incorporation of connectivity into conservation plans. In reality, connectivity matters for many different organisms across many different scales. Metrics like CSC that quantify landscape pattern across multiple different scales can make a valuable contribution to multi-scale landscape measurement, planning, and management.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
One of the most challenging problems for landscape ecology, and its applications in conservation, has been to find reliable ways of quantifying spatially structured ecological processes (Calabrese and Fagan 2004; Minor and Urban 2007; Moilanen 2011; Magris et al. 2016; Kininmonth et al. 2019; Daigle et al. 2020). Some ecological processes, such as photosynthesis or nitrogen fixation, happen at relatively small scales and can be easily included in models and conservation approaches by the inclusion of appropriate habitat types (e.g., forests or wetlands). Others, however, occur across many different scales and are heavily influenced over long time periods by spatial structure (including both the composition and the configuration of the land- or seascape at any given scale; Turner et al. (2001)). Examples of spatially structured ecosystem processes include dispersal, predation, nutrient subsidies, gene flow, and contagious perturbations such as fire and disease transmission. Although these processes are generally described and analysed under the banner of connectivity problems, available data and ecological understandings of how they occur are often inadequate for multi-scale quantification and conservation planning. For instance, telemetry studies often focus on a small number of larger species (e.g., Gredzens et al. 2014; Lea et al. 2016; Mazor et al. 2016). Habitat preferences during dispersal, and the relevance of matrix properties for dispersal, are poorly understood for most terrestrial organisms (and unknown for many marine organisms; Pittman et al. (2021)) and in addition to their long-standing interest in landscape ecology (Cowen et al. 2006; Prugh et al. 2008; Foster et al. 2012), remain active areas of research (Reis-Filho et al. 2019; San-José et al. 2019; Sanches et al. 2022), making it challenging to transfer results from in-depth analyses between different locations.
Most applications of connectivity analysis are developed at a single scale (grain and extent) of analysis (Lagabrielle et al. 2009; Tognelli et al. 2009; Grantham et al. 2011; Malcolm et al. 2011; Magris et al. 2017). In conservation planning, for example, the choice of planning scale is often determined by available data (Guerrero et al. 2013), such as satellite imagery or oceanographic modelling, that have constraints on grain size, extent, or both. Once a planning unit resolution and plan extent have been selected and implemented, re-extraction of the data and re-estimation of the results at a different scale require a considerable amount of additional GIS or modelling processing and time. Planning exercises thus become rapidly locked in to particular scales of analysis that are appropriate for some but not all relevant ecosystem processes.
Patches that are critical for connectivity may be of at least two different types. One, the traditional highly connected patch, will be frequently colonized by virtue of being near to many other suitable habitat patches (Saura and Pascual-Hortal 2007). The other, a stepping stone patch, may show lower individual occupancy but is critical for overall landscape connectivity because it connects two clusters of patches that would otherwise be separated (Saura et al. 2014). These two different connectivity contributions themselves occur at two different scales in both space and time, with highly connected patches being more likely to be occupied at any point in time (and supporting higher population densities) while stepping-stone patches make their main contributions to the overall occupancy of the entire landscape (Lindenmayer and Fischer 2006). Classical patch-specific nearest neighbour and clumpiness metrics measure the first type of connectivity but not the second.
Other existing metrics provide information about the relative contribution of each patch to the overall connectivity of the landscape (Calabrese and Fagan 2004). They include such measures as the probability of propagule or larval self-retention for each patch (White et al. 2014), the contribution of each patch to metapopulation growth rates (Jacobi and Jonsson 2011), and network-level metrics such as eigenvector centrality (Watson et al. 2011) or betweenness centrality (Magris et al. 2016). Centrality metrics derived from spatial network analysis (Borgatti et al. 2009; Cumming et al. 2010) have been gaining traction in the conservation literature (Treml et al. 2008; Galpern et al. 2011; Saura et al. 2014; Magris et al. 2016; Engelhard et al. 2017). For example, betweenness centrality, which is measured as the number of times a patch occurs on the shortest path between any other two patches in the network (Minor and Urban 2007), has been shown to be a reliable proxy for long-term persistence in marine systems (Magris et al. 2018). However, even these network-based metrics are usually calculated at only one or a few scales based on telemetry or mark-recapture data (Fletcher et al. 2011; Finn et al. 2014) that dictate the degree to which a given array of habitat patches is considered interconnected.
Since a continuous distance (rather than a bimodal variable, patches connected or not connected) can be used to estimate connectivity in network analysis, scale can be incorporated in geographic network analysis as a continuous variable by changing the threshold distance beyond which a dispersal event is considered possible or likely. For example, if an animal has been shown to disperse up to 10 km away from suitable habitat, distances from 0 to 10 km can be used to define a colonisation probability; patches further than 10 km apart can only be connected via a third patch which falls within 10 km of each. If another animal using the same habitat network is capable of dispersing up to 30 km away from suitable habitat, it will perceive the network as being more connected. Ideally, measures of connectivity that are intended to support community-level analyses or outcomes (such as habitat conservation by a network of protected areas, or maintenance of ecological functions that are underpinned by animals with a range of body sizes and dispersal capabilities) should incorporate a range of different perspectives on what constitutes connectivity rather than focusing on connectivity for a single species (Maciejewski and Cumming 2015).
Estimates of patch contributions to landscape connectivity have important real-world implications for conservation. Connectivity measures derived from network analysis have already been included via some conservation planning support tools, such as Marxan, prior to generating spatial conservation plans (e.g., Jacobi and Jonsson 2011; Watson et al. 2011; White et al. 2014; Magris et al. 2018; Friesen et al. 2019). This approach has the potential benefit of including connectivity as one of several alternative conservation priorities in an optimization algorithm. Doing so enables connectivity to be treated as another goal and traded off against other objectives (e.g., high local biodiversity or inclusion of endangered species) in conservation plans (Magris et al. 2017). The inclusion of connectivity via Marxan has not, however, been tested across multiple scales or compared to results from network analysis.
In this paper we propose that rather than seeking to identify locations that contribute optimally to network connectivity at a single scale, the patch that is most important for the integrity of a network will be the one that offers the greatest overall contribution to network connectivity over a full range of biologically meaningful scales. The relative multi-scale contributions of different patches can therefore be measured by calculating network centrality across a range of different scales and summing the outcome. The variance in these values provides an index of the variation in the contributions of the patch across scales. To explore the validity and a potential application of this approach we undertook a validation exercise using (1) a metapopulation simulation model and (2) a comparison to the most promising alternative approach, using Marxan. In both cases the application and value of cross-scale centrality (CSC) measures was strongly supported, suggesting that they can offer a useful and relatively simple, efficient approach to measuring and incorporating connectivity in landscape ecology research and applications, such as analyses of metapopulation dynamics or conservation planning.
Methods
We combined simulation models and conservation planning software (Fig. 1) to (1) determine whether CSC correlates significantly with overall patch occupancy; and (2) explore how CSC metrics relate to alternative approaches, specifically the use of Marxan conservation planning software.

Summary of our approach. The analysis included two phases: first, establishing whether cross-scale centrality measures for individual patches offer a reasonable surrogate for metapopulation persistence, based on proportional patch occupancy across each landscape; and second, comparing results from cross-scale betweenness centrality to those from an alternative approach to identifying ecologically critical patches, using Marxan conservation planning software. Red and blue arrow indicated nested loops. We first obtained patch occupancy estimates across a range of dispersal distances, then repeated the analysis at the same distances on a different (randomly generated) landscape
Simulation model
We used the Metalandsim package (Mestre et al. 2016) in R software (R Core Team 2020) to simulate metapopulation growth through different hypothetical landscapes and igraph (Csardi and Nepusz 2006) to measure patch (node) centrality for each habitat patch. Metalandsim provides a relatively easy translation between network and spatially explicit renderings of landscapes, making it easy to both calculate network metrics and simulate population growth and persistence in the same modelling environment. The package creates simulated landscapes using a set of parameters that define various aspects of landscape structure. To test the value of cross-scale network measurements under different conditions, we kept the total area and number of patches constant (respectively, 5000 m × 5000 m and 200 patches) across the 100 different simulated landscapes used for the study. Other parameters were either fixed or varied as described in Table 1, based on starting values suggested in the Metalandsim documentation. In particular, different organisms living within and using the same landscape at different scales were simulated using different dispersal parameters. To maintain transparency and interpretability, we did not attempt to modify colonisation, extinction, or patch size-dependent properties.
Metalandsim functions were applied using two slightly different sequences of operations. First, we defined a simulated landscape as a network using Rland.graph (Fig. 2). We altered dispersal distances within the network using convert.graph to redefine linkages between habitat patches. Each resulting graph was converted to an igraph network for measurement of the different patch centrality properties.

Example of a simulated 5 × 5 km landscape showing 200 habitat patches of different sizes. Patches that are interconnected by a hypothetical organism with a maximum dispersal distance of 500 m are joined by links. X and Y axes (in km) indicate the coordinates of each patch
Second, to determine the role of each habitat patch in metapopulation viability, the network from the first step was used in span.graph and simulate.graph to simulate the metapopulation processes of increase and dispersal. The simulate.graph function was run twenty times per individual landscape, covering each of the same dispersal distances for which network measures were calculated. The occupancy of each patch across all twenty of these runs was summed. Values are summed across different scales, not within a single scale, and so the sum reflects the full range of dispersal distances contingent on each individual dispersal distance. This can be viewed as a process of integrating across the uncertainty in the dispersal and colonisation parameters for dispersal distance; we effectively consider all possible distances, and the uncertainty is higher for longer distances. The sum is therefore the appropriate metric for integration.
To test the ability of CSC measures to capture highly connected patches, we then compared the proportional occupancy of each patch (i.e., proportion of model runs in which the patch was occupied) to its nodal centrality (summed across all hypothetical dispersal distances) within the network. We did this by calculating both parametric and non-parametric correlation coefficients and p values for the relationship between proportional occupancy and a selection of four nodal centrality measures (closeness, degree, eigenvector, and betweenness centrality respectively) for each patch across each of the 100 simulated landscapes.
To further clarify, because confusion is possible here, we aggregated individual nodal centrality measures across dispersal distances for each individual landscape but we did not aggregate patch centrality measures between different landscapes. Instead, we compared centrality to patch occupancy for each landscape and aggregated results across landscapes using correlation coefficients. The mean and deviation of the correlation coefficient and p value across all 100 model runs were used as a measure of how well each cross-scale centrality measure predicted patch occupancy.
In interpreting the simulation results, it is important to note that we have simulated only a simple, classical random patch distribution. The relative value of different nodal centrality measures will depend to some extent on the properties of the landscape being analysed. Comparisons of occupancy individually by patch do not test directly for stepping stone connectivity. Theory suggests that stepping stone patches should exhibit high betweenness centrality (i.e., the patch should appear on the shortest path between two other patches more frequently than expected). Distinguishing between high direct connectivity and stepping stone connectivity influences on metapopulation simulation results (in the real world, where stepping stones are often islands in a sea of cleared land, individual patches are unlikely to have high scores in both metrics) is challenging without the use of patch centrality measures that have been explicitly designed for this purpose. Thus, we have presented both degree centrality and betweenness centrality in our worked example even though the simulation results favour degree centrality, trusting in a body of widely accepted theory that indicates that betweenness centrality is a superior measure of stepping stone connectivity (Bodin and Norberg 2007; Zetterberg et al. 2010; Boulanger et al. 2020). We would continue to recommend the usage of betweenness centrality as a CSC metric in situations where stepping stone connectivity is important; we did not simulate highly non-random networks, in which stepping stones are important.
Real-world data and connectivity metrics
To compare the uses of CSC in conservation planning to the strongest alternative, we used a real-world data set derived from Brazilian coral reefs. These reefs are among the highest conservation priority areas in the Atlantic Ocean, due to their high levels of endemism and unique geomorphologic formation, which are significantly different from the well-known coral reef ecosystems of the Caribbean and Indo-Pacific regions (Leão and Dominguez 2000). We used spatial data about demographically significant dispersal links (i.e., those that maintain populations) between coral reefs to represent connectivity in Marxan (Magris et al. 2016) and for this scale of analysis, treated each coral reef as a single habitat patch. Connectivity is defined as the likelihood that, for each modelled species, larvae from a natal reef are able to reach a neighbouring or nonadjacent reef. Connectivity was modelled using daily data on ocean currents from 2008 to 2012 (Atlantic RTOFS) and reproductive strategy information (timing of reproduction and pelagic larval duration) for four model species that captured a range of species dispersal potential (a brooder coral, a broadcast spawning coral, a roving herbivorous fish, and a large carnivorous fish). The yearly connectivity matrices were averaged to produce an asymmetric connectivity matrix for each modelled species. These reef-based connectivity matrices were summarised at the patch (reef) level by considering all individual reef polygons adjacent to each other. The network of potential habitat for the four modelled species comprised 42 patches; their adjacency was measured in GIS from the centres of polygons overlaid on each reef. Further details of the parameterization of larval simulations can be found in Magris et al. (2016).
Three metrics were calculated from the connectivity matrix for each modelled species: (i) outflux, which is related to the source strength of a patch and its ability to sustain the populations of surrounding units through its outgoing connections; (ii) local retention, which is associated with the degree to which a patch is self-sustaining in isolation and, hence, should be protected; and (iii) betweenness centrality, which is related to the ability of stepping-stone patches to control fluxes, and help spread risk against disturbances. The approach assumes that simultaneously representing all these connectivity surrogates will promote persistence in a proposed spatial configuration of reserves; specifically, patches where individuals replace themselves as closed populations and sites that are critical for the movement of individuals in a network.
Connectivity matrices from all four model species were used to calculate larval fluxes, adapting the formula of Urban and Keitt (2001):
where, \({f}_{ij}\) is the expected dispersal flux from patch \(i\) to patch \(j\), \({p}_{ij}\) is the probability of settling on patch \(j\) from \(i, {s}_{i}\) is relativized as the proportion of total habitat area \(s{\text{tot}}\). We then summed all fluxes for all outgoing links to determine out-flux for each particular patch. We measured local retention for each patch as the diagonal elements of connectivity matrices. Betweenness centrality was measured as indicated above (i.e., the number of times an area occurs on the shortest path between any other two areas in the network (Minor and Urban 2008)).
Marxan analysis
Our connectivity metrics were related to replenishment of larvae, increased potential recovery, and the capacity of reefs to be self-sustaining. We computed a cumulative distribution curve for the values of each conservation feature and used the top third of reef patches for each of the three connectivity metrics to derive the minimum amounts to be targeted in the prioritisation (i.e., targets or objectives). The prioritisation objectives were defined by calculating the percentage that top-ranked reef patches contributed to the total values of each metric.
We ran Marxan under two different sets of assumptions (Table 2) to explore whether different understandings about what needs to be protected would influence the patches identified by Marxan as critical for connectivity. The program uses a simulated annealing algorithm to find the most parsimonious combination of sites (usually specified as planning units of a regular size and shape) defined in a planning exercise. It bases its site selection on both accumulating desired proportions of individual features (e.g., conserving 40% of known occurrences of particular species or habitats) and avoiding costs (e.g., minimizing land price or opportunity cost). In finding solutions it incrementally solves a multivariate optimization problem, starting with a random selection of planning units and gradually improving on its starting point by including or excluding planning units. Simulated annealing includes a level of stochasticity in its incremental approach to help the program to avoid becoming stuck at a locally optimal solution and instead, to identify a global optimum. Since Marxan does not necessarily identify the same optimal solution every time it is run, we followed standard practice of running Marxan 100 times under each set of assumptions and counting the number of times each patch was selected for inclusion in the reserve network as a measure of that patch’s contribution to ecological processes. These values were used to evaluate differences in the spatial distribution of priority areas by comparing the spatial distribution of areas identified through the betweenness centrality scenario against the scenario targeting all metrics. Patches with selection frequencies > 75 were identified as high priorities.
Network analysis for real-world data
To quantify changes in connectivity with scale, we used the geographic distance between individual reef patches to generate networks at different scales ranging from 20 to 1800 km in steps of 20 km (i.e., at the 20 km scale a link was assigned to every pair of reef patches located within 0–20 km from one another; at 40 km, 0–40 km; and so on). This generated 90 different networks. The mean distance between all pairs of individual patches was 627 km; by 1800 km, each patch was directly connected to all other patches.
We defined ecological scale as the potential for a dispersal event beyond a given distance. In geographic analyses, centrality usually changes with scale because changing the distance over which patches are considered to be connected alters the pattern of connections. We therefore calculated centrality measures for each of our different patches for each individual scale. The selection of a range of sampling distances is based on the properties of individual landscapes. Typically, a plot of the value of a network centrality measure against the distance at which patches are considered connected will increase gradually with increasing dispersal distance and then reach an asymptote. We see little value in including measures at distances beyond this asymptotic value, which sets a natural upper limit on the range of dispersal distances to include in a given analysis.
As a single, multi-scale measure of each patch’s contribution to network connectivity, we summed all centrality measures for that patch across all networks, and hence across all ecological scales. This provided a single Cross-Scale Centrality (CSC) value for each of the 42 patches (individual reefs) in the analysis.
Comparing results
To contrast CSC and the results from Marxan, we compared the number of times each patch (reef) was selected for inclusion into the reserve network by Marxan (from 100 runs) against its CSC value. The Marxan results for our coral reef data set were mostly binary (i.e., individual patches were either selected in 100% of runs or 0% of runs, although not in every case). We thus used boxplots and a standard ANOVA, rather than correlations, to compare the means and deviations of CSC between included and excluded patches. If Marxan were actively selecting patches with higher connectivity across multiple scales, we would expect that included patches had significantly greater CSC values than excluded patches.
Lastly, we calculated the Spearman's correlation between the centrality metric at a certain dispersal threshold and the proportion of times the patch was included in the reserve network by Marxan. We plotted these correlation values against all dispersal thresholds. If Marxan genuinely captured network connectivity across multiple scales, we would expect this line to be high and flat, indicating a strong correlation with centrality at all potential dispersal distances.
Results
Simulation analyses
The simulation analyses suggested that simple cross-scale degree centrality (i.e., the number of links incident on a patch) was the strongest correlate of the frequency of individual patch occupancy (Fig. 3 and Table 3). An ANOVA by groups indicated that there were significant differences in the correlations of different CSC metrics to patch occupancy (F = 81.7, Df = 3, p < 0.0001). A Tukey’s multiple comparison of means test showed that all groups were significantly different from each other (p < 0.000) except for eigenvector and betweenness centrality, which were not significantly different (p < 0.12).

Boxplot showing the mean and deviation of Spearman’s correlations between different patch centrality measures and proportional patch occupancy in a metapopulation model. The width of each small horizontal bar is proportional to the number of points it contains. All metrics correlated significantly with metapopulation occupancy (Table 3); letters above bars indicate similarities or differences between metrics based on an ANOVA and Tukey’s HSD Test, as explained in the text
This result provides clear support for the hypothesis that patches that are highly connected across a range of different scales will also contribute the most to the persistence of a diversity of metapopulations, meeting typical conservation objectives. Keeping in mind that our simulation networks did not deliberately include critically important stepping stone patches, the weaker performance of betweenness centrality here is presumably due to its focus on overall network flow and the higher weighting that it will give to stepping stone patches relative to those that are in the centres of clumps of patches.
Marxan analysis of reef data
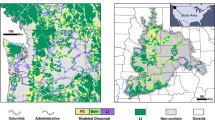
The Marxan analysis showed that targeting either betweenness centrality or all metrics simultaneously resulted in a large number of patches being selected for conservation (Fig. 4). About 73% of areas identified as high-priority overlapped between the two Marxan scenarios. When we included all connectivity metrics in Marxan, 80% of the study region was identified as having priority for conservation.

Spatial distribution of high priority coral reefs for each scenario using Marxan. In A, patches coloured in red represent priorities (selection frequency > 75) when maximizing betweenness centrality metric only. In B, patches coloured in black represent priorities (selection frequency > 75) when maximizing all connectivity metrics. Unfilled patches were not selected as priorities by Marxan. Inset map shows the location of our study region in the southwestern Atlantic Ocean
Network analysis of reef data
The connectivity of the network changed substantially with scale (Fig. 5). As would be expected, cross-scale betweenness centrality within the network was unevenly distributed, showing network-specific idiosyncrasies.

Network graph generated using the geographic distance matrix at four different scales: a 100 km, b 500 km, c 1000 km, and d 1500 km. This figure shows how both overall network connectivity and the contributions of individual patches to network connectivity depend on assumptions about the scale of dispersal. Our approach integrates over the ‘nuisance variable’ of scale by summing contributions to connectivity over a range of scales
Comparing results for reef data
The Marxan and CSC results showed relatively little correspondence, with Marxan excluding several patches (e.g., patch numbers 8, 32, 31) with high CSC values. Our results show a largely bimodal pattern of conservation importance when using Marxan in the prioritisation: with a few exceptions, patches were mostly either selected or excluded from the spatial solutions (Fig. 6). Reefs selected for inclusion by Marxan did not differ significantly in their CSC from those that were excluded (Fig. 7). The lowest p value, for degree centrality in scenario 1, was still an unconvincing 0.28 (F = 1.22, df = 40). Across all different measures the mean F-statistic and p value were respectively F = 0.63, p = 0.50 (Scenario 1) and F = 0.26, p = 0.66 (Scenario 2). Marxan thus did not preferentially select patches with higher CSC and hence, based on the independent validation provided by our simulation results, did not reliably capture multi-scale metapopulation patch occupancy.

Comparisons of a the average of 100 Marxan runs focusing on betweenness centrality and the CSC metric, calculated independently using network analysis; and b the average of 100 Marxan runs focusing on optimizing all different criteria and the CSC metric. Marxan produces nearly identical results in both scenarios. Similar outcomes also occur when Marxan results are compared to other centrality metrics, here using Marxan optimization across all criteria: c degree centrality, d closeness centrality, and e eigenvector centrality


Boxplots comparing CSC values (y axis) for different reefs and centrality measures against the number of times they were selected by Marxan (x axis) for inclusion in the reserve network. Data are shown for Scenario 1: a betweenness centrality; b degree centrality; c closeness centrality; and d eigenvector centrality. For Scenario 2: e betweenness centrality; f degree centrality; g closeness centrality; and h eigenvector centrality. As reported in the text, ANOVA tests indicated that none of the means of CSC values for the different groups were significantly different
Finally, consideration of the Spearman’s correlation between centrality and Marxan runs (using all criteria, but assuming that dispersal was limited to particular distances), showed that Marxan provided good inclusion of connectivity at a few scales but performed poorly at other scales (Fig. 8). There was reasonable inclusion of connectivity at coarser scales (over 1700 km) but considerable variation in between, and in relation to different centrality measures.

Comparison of the correlation between different centrality metrics and the results from Marxan Scenario 2 for different assumed dispersal distances. This figure shows how the Marxan approach captured connectivity unevenly across scales as well as demonstrating some of the differences between different centrality measures. Line colours are red, betweenness centrality; dark blue, closeness centrality; pale blue, eigenvector centrality; and gold, degree centrality
Discussion
Overall, our results show that measures of cross-scale connectivity derived from network analysis can identify the habitat patches that are most critical for the persistence of an entire community of organisms with differing dispersal capabilities. Individual patches with higher cross-scale centrality (CSC) metrics were proportionally more occupied in metapopulation simulations covering a full range of different dispersal capabilities. The distinctions between different kinds of network measures and their ability to capture different kinds of connectivity are important to recognize and understand, and we propose that the choice of metric should be based primarily on theory and the specific details of the kind of connectivity that is of greatest interest in a given study. Cross-scale degree centrality emerged here as the most promising centrality measure, suggesting that the ability of a patch to form individual linkages at multiple scales is more important for its individual metapopulation occupancy than its overall contribution to landscape connectivity. It is likely, however, that degree centrality is less effective than other measures in non-random landscapes, particularly where several larger clusters of patches have just a few critically important ‘stepping stone’ connections. Testing how our findings apply under different levels of patch dispersion and randomness would be an obvious next step in determining the optimal measure(s) of CSC to apply in real-world studies.
Although progress has been made in improving connectivity modelling in the last decade (Bodin and Saura 2010; Kool et al. 2013), many regions lack spatially explicit connectivity information and the required skill set involved is often daunting. The results of our simulation models demonstrate that CSC measures provide a reasonable surrogate for metapopulation occupancy in realistic situations where metapopulations of many species, with different dispersal capabilities, co-occur within a single landscape. Although it would be possible to apply our approach to a specific landscape and repeat the simulations under the specific geographic configuration of a given location, our analysis suggests that this is unnecessary; the vast majority of simulations indicated a significant correlation between CSC and metapopulation occupancy over time.
The second section of our analysis, the real-world case study application, suggested that including small amounts of dispersal data in conservation planning tools does not provide an adequate solution to including multiple scales of ecological connectivity in conservation planning exercises. The value of including dispersal data from telemetry or other sources may be higher when landscape structure is complex and dispersal pathways are convoluted. Conservation plans are often developed at a single grain and extent of analysis (Leslie 2005; Guerrero et al. 2013; Álvarez-Romero et al. 2018). Methods to design protected area networks using connectivity information have proliferated in the literature (Luque et al. 2012; Kool et al. 2013; Burgess et al. 2014; Magris et al. 2014; Balbar and Metaxas 2019). Although this body of literature has dealt with challenging practical problems of managing ecological processes in a regional setting, there has been little evidence for how patterns of conservation importance vary across multiple scales. In our analysis, seeking to protect ecological processes that are important for coral reefs without capturing multiple-scale variability did not appear to consistently achieve good outcomes. A more robust prioritization approach for coral reef conservation that has no limit on how many scales can be addressed in the same prioritization problem is more likely to provide protection for species over different spatial scales.
Multi-scale approaches to landscape connectivity measurement and conservation planning are necessary for conservation to maintain ecologically functional landscapes that protect species with different habitat needs and dispersal abilities (Poiani et al. 2000) as well as to understand multi-scale impacts arising from land-use change and global warming (Dilts et al. 2016). Although this need has been recognized conceptually for more than 20 years (Keitt et al. 1997; Poiani et al. 2000; Sanderson et al. 2002), rigorous multi-scale measures of habitat connectivity have proven elusive. Cross-scale centrality measures can quantify consistent connections over time (Treml et al. 2008), including both classical and stepping stone connections, and help identify critical pathways for maintaining functionally connected local populations even of long-distance dispersers (Saura et al. 2014; Magris et al. 2016). Our approach could be further extended to evaluate how the relative importance of each patch would change with temporal variability in connectivity in response to disturbances (Bodin and Saura 2010); further research is needed to determine whether CSC, or a time series of CSC based on fluctuations in landscape permeability, can offer a robust tool to inform conservation across temporal scales. Additional metrics, such as nestedness and modularity, may also be necessary to capture other aspects of habitat morphology (Cumming 2002; Moore et al. 2016). Lastly, replacing a geographic straight line with an estimate of habitat resistance or permeability can allow the approach to be tailored to situations in which the properties of the matrix may be a key element of system dynamics (Calder et al. 2015; Peterman et al. 2019).
For landscape measurement and conservation planning we therefore derive two clear recommendations. First, estimating CSC for each patch offers a useful guideline to the multi-scale contributions of each individual patch to connectivity and metapopulation persistence within the system. Second, unless the scales of connectivity between patches are very well known and clearly defined, including connectivity estimates via Marxan (or other conservation-planning tools) is insufficient. Our approach can easily be used with Marxan in a complementary manner, with the analyst including CSC metrics transparently in the development of a plan by adding or excluding locations that respectively make important or trivial contributions to connectivity. CSC measures are fast to estimate and intuitive to understand; and since they are not strongly tied to a single grain of analysis, they can be easily recalculated at different grains and under different assumptions. CSC thus meets the main criteria required for a useful landscape metric: it is easily measured, efficient to calculate, interpretable, and can be genuinely related to a mechanism. Transparency and interpretability in conservation planning, and its ability to easily consider a range of alternative solutions, will be particularly important in trying to balance different planning objectives (e.g., the tradeoffs between financial and ecological considerations). As the discipline of landscape ecology advances, we anticipate that measures that consider the multi-scale nature of the real world and seek to quantify landscape properties across a range of different scales will be increasingly important.
Data availability
The article does not present new data. Code is available on request.
References
Álvarez-Romero JG, Mills M, Adams VM et al (2018) Research advances and gaps in marine planning: towards a global database in systematic conservation planning. Biol Conserv 227:369–382
Balbar AC, Metaxas A (2019) The current application of ecological connectivity in the design of marine protected areas. Glob Ecol Conserv 17:e00569
Bodin Ö, Norberg J (2007) A Network Approach for Analyzing Spatially Structured Populations in Fragmented Landscape. Landscape Ecol 22:31–44
Bodin Ö, Saura S (2010) Ranking individual habitat patches as connectivity providers: integrating network analysis and patch removal experiments. Ecol Model 221(19):2393–2405
Borgatti SP, Mehra A, Brass DJ, Labianca G (2009) Network analysis in the social sciences. Science 323(5916):892–895
Boulanger E, Dalongeville A, Andrello M, Mouillot D, Manel S (2020) Spatial graphs highlight how multi-generational dispersal shapes landscape genetic patterns. Ecography 43(8):1167–1179
Burgess SC, Nickols KJ, Griesemer CD et al (2014) Beyond connectivity: how empirical methods can quantify population persistence to improve marine protected-area design. Ecol Appl 24(2):257–270
Calabrese JM, Fagan WF (2004) A comparison-shopper’s guide to connectivity metrics. Front Ecol Environ 2(10):529–536
Calder J-L, Cumming GS, Maciejewski K, Oschadleus HD (2015) Urban land use does not limit weaver bird movements between wetlands in Cape Town, South Africa. Biol Cons 187:230–239
Cowen R, Paris C, Srinivasan A (2006) Scaling of connectivity in marine populations. Science 311(5760):522–527
Csardi G, Nepusz T (2006) The igraph software package for complex network research. InterJournal 1695(5):1–9
Cumming G (2002) Habitat shape, species invasions, and reserve design: Insights from simple models. Conserv Ecol 6(1):06103
Cumming GS, Bodin Ö, Ernstson H, Elmqvist T (2010) Network analysis in conservation biogeography: challenges and opportunities. Divers Distrib 16(3):414–425
Daigle RM, Metaxas A, Balbar AC et al (2020) Operationalizing ecological connectivity in spatial conservation planning with Marxan Connect. Methods Ecol Evol 11(4):570–579
Dilts TE, Weisberg PJ, Leitner P et al (2016) Multiscale connectivity and graph theory highlight critical areas for conservation under climate change. Ecol Appl 26(4):1223–1237
Engelhard SL, Huijbers CM, Stewart-Koster B, Olds AD, Schlacher TA, Connolly RM (2017) Prioritising seascape connectivity in conservation using network analysis. J Appl Ecol 54(4):1130–1141
Finn J, Brownscombe J, Haak C et al (2014) Applying network methods to acoustic telemetry data: modeling the movements of tropical marine fishes. Ecol Model 293:139–149
Fletcher RJ, Acevedo MA, Reichert BE, Pias KE, Kitchens WM (2011) Social network models predict movement and connectivity in ecological landscapes. Proc Natl Acad Sci USA 108(48):19282–19287
Foster NL, Paris CB, Kool JT et al (2012) Connectivity of Caribbean coral populations: complementary insights from empirical and modelled gene flow. Mol Ecol 21(5):1143–1157
Friesen SK, Martone R, Rubidge E, Baggio JA, Ban NC (2019) An approach to incorporating inferred connectivity of adult movement into marine protected area design with limited data. Ecol Appl 29(4):e01890
Galpern P, Manseau M, Fall A (2011) Patch-based graphs of landscape connectivity: a guide to construction, analysis and application for conservation. Biol Conserv 144(1):44–55
Grantham HS, Game ET, Lombard AT et al (2011) Accommodating dynamic oceanographic processes and pelagic biodiversity in marine conservation planning. PLoS ONE 6(2):e16552
Gredzens C, Marsh H, Fuentes MM, Limpus CJ, Shimada T, Hamann M (2014) Satellite tracking of sympatric marine megafauna can inform the biological basis for species co-management. PLoS ONE 9(6):e98944
Guerrero AM, McAllister R, Corcoran J, Wilson KA (2013) Scale Mismatches, Conservation Planning, and the Value of Social-Network Analyses. Conserv Biol 27(1):35–44
Jacobi MN, Jonsson PR (2011) Optimal networks of nature reserves can be found through eigenvalue perturbation theory of the connectivity matrix. Ecol Appl 21(5):1861–1870
Keitt TH, Urban DL, Milne BT (1997) Detecting critical scales in fragmented landscapes. Ecol Soc 1(1):1
Kininmonth S, Weeks R, Abesamis RA et al (2019) Strategies in scheduling marine protected area establishment in a network system. Ecol Appl 29(1):e01820
Kool JT, Moilanen A, Treml EA (2013) Population connectivity: recent advances and new perspectives. Landscape Ecol 28(2):165–185
Lagabrielle E, Rouget M, Payet K et al (2009) Identifying and mapping biodiversity processes for conservation planning in islands: a case study in Réunion Island (Western Indian Ocean). Biol Conserv 142(7):1523–1535
Lea JS, Humphries NE, von Brandis RG, Clarke CR, Sims DW (2016) Acoustic telemetry and network analysis reveal the space use of multiple reef predators and enhance marine protected area design. Proc R Soc B 283(1834):20160717
Leão ZM, Dominguez JM (2000) Tropical coast of Brazil. Mar Pollut Bull 41(1–6):112–122
Leslie HM (2005) A synthesis of marine conservation planning approaches. Conserv Biol 19(6):1701–1713
Lindenmayer DB, Fischer J (2006) Habitat fragmentation and landscape change. Island Press, Washington
Luque S, Saura S, Fortin M-J (2012) Landscape connectivity analysis for conservation: insights from combining new methods with ecological and genetic data. Landscape Ecol 27(2):153–157
Maciejewski K, Cumming GS (2015) Multi-scale network analysis shows scale-dependency of significance of individual protected areas for connectivity. Landscape Ecol 31(4):761–774
Magris RA, Andrello M, Pressey RL et al (2018) Biologically representative and well-connected marine reserves enhance biodiversity persistence in conservation planning. Conserv Lett 11:e12
Magris RA, Pressey RL, Mills M, Vila-Nova DA, Floeter S (2017) Integrated conservation planning for coral reefs: designing conservation zones for multiple conservation objectives in spatial prioritisation. Glob Ecol Conserv 11:53–68
Magris RA, Pressey RL, Weeks R, Ban NC (2014) Integrating connectivity and climate change into marine conservation planning. Biol Conserv 170:207–221
Magris RA, Treml EA, Pressey RL, Weeks R (2016) Integrating multiple species connectivity and habitat quality into conservation planning for coral reefs. Ecography 39(7):649–664
Malcolm H, Foulsham E, Pressey R et al (2011) Selecting zones in a marine park: early systematic planning improves cost-efficiency; combining habitat and biotic data improves effectiveness. Ocean Coast Manag 59:1–12
Mazor T, Beger M, McGowan J, Possingham HP, Kark S (2016) The value of migration information for conservation prioritization of sea turtles in the Mediterranean. Glob Ecol Biogeogr 25(5):540–552
Mestre F, Cánovas F, Pita R, Mira A, Beja P (2016) An R package for simulating metapopulation dynamics and range expansion under environmental change. Environ Model Softw 81:40–44
Minor ES, Urban DL (2007) Graph theory as a proxy for spatially explicit population models in conservation planning. Ecol Appl 17(6):1771–1782
Minor ES, Urban DL (2008) A graph-theory framework for evaluating landscape connectivity and conservation planning. Conserv Biol 22(2):297–307
Moilanen A (2011) On the limitations of graph-theoretic connectivity in spatial ecology and conservation. J Appl Ecol 48(6):1543–1547
Moore C, Grewar J, Cumming GS (2016) Quantifying network resilience: comparison before and after a major perturbation shows strengths and limitations of network metrics. J Appl Ecol 53(3):636–645
Peterman WE, Winiarski KJ, Moore CE, Carvalho CdS, Gilbert AL, Spear SF (2019) A comparison of popular approaches to optimize landscape resistance surfaces. Landscape Ecol 34(9):2197–2208
Pittman S, Yates K, Bouchet P et al (2021) Seascape ecology: identifying research priorities for an emerging ocean sustainability science. Mar Ecol Prog Ser 663:1–29
Poiani KA, Richter BD, Anderson MG, Richter HE (2000) Biodiversity conservation at multiple scales: functional sites, landscapes, and networks. Bioscience 50:133–146
Prugh LR, Hodges KE, Sinclair ARE, Brashares JS (2008) Effect of habitat area and isolation on fragmented animal populations. Proc Natl Acad Sci USA 105:20770–20775
R Core Team (2020) R: a language and environment for statistical computing. R Core Team, Vienna
Reis-Filho JA, Schmid K, Harvey ES, Giarrizzo T (2019) Coastal fish assemblages reflect marine habitat connectivity and ontogenetic shifts in an estuary-bay-continental shelf gradient. Mar Environ Res 148:57–66
San-José M, Arroyo-Rodríguez V, Jordano P, Meave JA, Martínez-Ramos M (2019) The scale of landscape effect on seed dispersal depends on both response variables and landscape predictor. Landscape Ecol 34(5):1069–1080
Sanches VQA, Menezes JFS, Prevedello JA, Almeida-Gomes M, Oliveira-Santos LGR (2022) Can matrix structure affect animal navigation between fragments? A dispersal experiment using release platforms. Biotropica 54(2):370–380
Sanderson EW, Redford KH, Vedder A, Coppolillo PB, Ward SE (2002) A conceptual model for conservation planning based on landscape species requirements. Landsc Urban Plan 58(1):41–56
Saura S, Bodin Ö, Fortin MJ (2014) Stepping stones are crucial for species’ long-distance dispersal and range expansion through habitat networks. J Appl Ecol 51(1):171–182
Saura S, Pascual-Hortal L (2007) A new habitat availability index to integrate connectivity in landscape conservation planning: comparison with existing indices and application to a case study. Landsc Urban Plan 83(2–3):91–103
Tognelli MF, Fernández M, Marquet PA (2009) Assessing the performance of the existing and proposed network of marine protected areas to conserve marine biodiversity in Chile. Biol Cons 142(12):3147–3153
Treml EA, Halpin PN, Urban DL, Pratson LF (2008) Modeling population connectivity by ocean currents, a graph-theoretic approach for marine conservation. Landscape Ecol 23:19–36
Turner MG, Gardner RH, O’Neill RV (2001) Landscape ecology in theory and practice: pattern and process. Springer-Verlag, Berlin
Urban D, Keitt T (2001) Landscape connectivity: a graph-theoretic perspective. Ecology 82(5):1205–1218
Watson JR, Siegel DA, Kendall BE, Mitarai S, Rassweiller A, Gaines SD (2011) Identifying critical regions in small-world marine metapopulations. Proc Natl Acad Sci USA 108(43):E907–E913
White JW, Schroeger J, Drake PT, Edwards CA (2014) The value of larval connectivity information in the static optimization of marine reserve design. Conserv Lett 7(6):533–544
Zetterberg A, Mörtberg UM, Balfors B (2010) Making graph theory operational for landscape ecological assessments, planning, and design. Landsc Urban Plan 95(4):181–191
Acknowledgements
This research was supported by the ARC Centre of Excellence for Coral Reef Studies at James Cook University and a James S. McDonnell Foundation complexity scholar award to G.S. Cumming. RAM acknowledges support from CNPq/Brazil.
Funding
Open Access funding enabled and organized by CAUL and its Member Institutions. This research was supported by the ARC Centre of Excellence for Coral Reef Studies.
Author information
Authors and Affiliations
Contributions
GC and RM conceived the study. GC, KM and RM ran analyses. The first draft was written by GC and all authors contributed to the final version. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors have no relevant competing interests to disclose.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Cumming, G.S., Magris, R.A. & Maciejewski, K. Quantifying cross-scale patch contributions to spatial connectivity. Landsc Ecol 37, 2255–2272 (2022). https://doi.org/10.1007/s10980-022-01497-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10980-022-01497-7