
Abstract
Context
Large-scale programs for eradication of pest mammals are confronted with the challenge of managing reinvasion. Exploiting high-elevation landscape features that naturally limit the rate of reinvasion is a strategy that is presumed to improve the success of such initiatives, however, the efficacy of doing so has not yet been investigated.
Objectives
We aimed to assess whether high-elevation landforms limit the movements of 10 species of invasive small mammal in New Zealand to such a degree that they could be exploited in landscape-scale eradication programmes.
Methods
We determined the upper elevation limits of species’ distributions, and made spatial predictions based on occupancy models. We applied these in concert to a 310,000 ha area of rugged mountainous environments and identified landforms that function as dispersal barriers to each species of interest. We validated our predictions with existing presence/absence and GPS movement data, and tested our predictions of high-elevation landform barriers with the GPS movement data of a sample of European hedgehogs (Erinaceus europaeus).
Results
We found that the extent of barriers which limited movement ranged from widespread (5/10 species), to localised, (3/10 species) to limited (2/10 species). Our predictions of hedgehog movement barriers were strongly supported by GPS movement data of 26 hedgehogs that were tracked in the study area.
Conclusions
Our findings show there is enormous potential to advance landscape-scale eradication of invasive small mammals in areas adjacent to high-elevation landforms by identifying and exploiting landscape features that limit the movement of target species in the strategies of eradication programmes.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The eradication of invasive species to protect or restore threatened species populations, native biodiversity, or ecosystem function is a common goal in conservation management and is attempted widely around the world (Simberloff 2009; Robertson et al. 2017). Although unique to their situations, all efforts to eradicate invasive species encounter a common challenge of preventing, or at least severely limiting, invasive species from reinvading after successful eradication from an area (Myers et al. 2000). Programmes targeting invasive terrestrial mammals on offshore islands enjoy high rates of success as the risk of reinvasion can be extremely low due to their isolation from mainland reinvasion sources (Jones et al. 2016). Mainland eradication projects, commonly defined as terrestrial eradication zones surrounded by untreated areas, are seldom isolated from surrounding pest populations and therefore require management to prevent or limit reinvasion. Defence of these areas has been achieved at relatively small-scales (several thousand hectares) by installing dispersal-limiting infrastructure such as fences (Burns et al. 2012; Innes et al. 2019), by implementing buffer zones of intensified trapping or treatment areas, and/or by exploiting naturally-occurring barriers to dispersal, such as inshore bodies of water and coastal peninsulas (Innes et al. 2019), glaciated terrain (Robertson and Gemmell 2004), and high-volume, fast flowing rivers (Cook et al. 2021). A barrier to dispersal or ‘movement barrier’ can be defined as any landscape feature that severely limits or prevents the movement of a species through space (Caplat et al. 2016). Successful mainland eradication projects may employ multiple dispersal-limiting approaches in concert.
Integrating landscape-scale features that naturally limit the movement of terrestrial invasive species offers tremendous opportunities to protect very large areas from the impacts of pests. Such features function by constraining the movement of invasive species, with the exact nature of the barrier being specific to the biological characteristics of the species in question (Caplat et al. 2016). Improved understanding of what comprises a landscape feature that effectively limits dispersal for a particular species is needed to guide the integration of such features into eradication strategies. In steep mountainous regions, high-elevation mountain ranges extend into the alpine and nival zones that are likely unfavourable areas for many terrestrial mammal species. This is due to a decrease in vegetation biomass with increasing elevation and therefore decreasing opportunities for food and shelter; lower temperatures and adverse climatic conditions; or strong seasonal changes (e.g. winter snow loading and low temperatures). Mountains can be considered as impermeable barriers to a species with narrow habitat requirements, such as aquatic taxa (Perrigo et al. 2020), and have been shown to limit the movement of species that have strong associations with riparian habitat e.g. American mink (Neovision vision) (Zalewski et al. 2009; Fraser et al. 2013). Similarly, Haran et al. (2015) attributed the prevention of gene flow of nematode-infested beetles across the Pyrenean Mountain chain to the lower temperatures at high elevations. Given the size and extent of mountain ranges, it is possible that they are one of the most significant barriers to terrestrial species, as evidenced by their role in allopatric speciation (Coyne and Orr 2004), yet their utility in controlling the movements of invasive small mammal species remains largely unexplored.
New Zealand is a country plagued by a suite of invasive small mammals, each of which play a part in the ongoing degradation of native biodiversity values and populations of vulnerable native species (Russell and Stanley 2018). Following the successful eradication of rodents from many of its offshore islands (Russell and Broome 2016) and after some success in early initiatives to remove invasive predators, much of the nation’s attention has shifted to the establishment of mainland predator-free sanctuaries, which serve as the current “maximum-practicable” step on the path to a predator-free New Zealand (Innes et al. 2019). As regional predator-free initiatives grow larger in size and ambition, larger tracts of increasingly complex and inaccessible terrain will need to be cleared of exotic predators. Because of the practical, logistical, and budgetary constraints of sustained pest suppression or fence installation across such areas, any landscape feature that limits invasive predator movements will be valuable in reducing costs and ensuring programme success. An opportunity to use naturally-occurring movement barriers can be found in the South Island’s extensive mountainous terrain, the most significant of which is the main axial range, the Southern Alps. While it is well documented that almost all of New Zealand’s invasive small mammal species occur above the tree line (O’Donnell et al. 2017), there is evidence that some of these species’ distributions are limited by large, mountainous landforms. For example, there is genetic divergence of feral cat populations (Felis catus) separated by large mountain ranges (Cross 2016); capture rates of ship rats (Rattus rattus) decrease to zero above 1000 m a.s.l. (Christie et al. 2017); and there are upper elevation limits to the distributions of 10 invasive species in the eastern dryland zone of New Zealand’s South Island (Foster et al. 2021a). The combination of the high-elevation mountainous topography of the Southern Alps and the presence of 10 out of 12 of New Zealand’s invasive small mammal species presents an opportunity to investigate to what extent high-elevation landforms might limit the movement of invasive small mammals, and how these limitations can be integrated into management strategies.
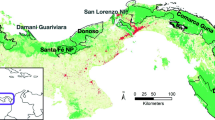
The Te Manahuna Aoraki project seeks to create a 310,000 ha sanctuary free from invasive mammalian predators in the upper Mackenzie Basin. This sanctuary concept relies on the extensive chain of mountain ranges to prevent or significantly limit the reinvasion of invasive species once they are removed. The project area is bounded by the Ben Ohau Range to the west, the Main Divide of the Southern Alps to the north, and the Two Thumb Range to the east (Fig. 1). The southern (open) edge of the project area is proposed to abut the edges of Lake Takapō/Tekapo and Lake Pūkaki, the Takapō-Pūkaki hydroelectric canal and a series of short predator-proof wing fences that are planned to join the ends of the mountain barrier to the lake and canal features to defend the ‘open’ edge of the sanctuary from reinvasion. The project boundary is 277 km long, of which 212 km is along the crest of three main mountain ranges. This study investigates the potential value of the Ben Ohau Range, the Main Divide and the Two Thumb Range in limiting reinvasion by small mammal species, and will explore how the internal landforms of the area might isolate populations of invasive species within the Te Manahuna Aoraki project area.
Methods.
Study area
This study was based in the uplands of the Mackenzie Basin, situated in the high country of South Canterbury in New Zealand’s South Island (Fig. 1). Low altitude areas (500–700 m above sea level, a.s.l.) in this region are comprised of expansive dryland fescue tussock (Festuca novae-zelandiae) grasslands, braided rivers, and intermontane valleys with dense tracts of subalpine shrub and small, isolated patches of beech forest. Snow tussock grasslands, alpine herbfields, and scree slopes dominate the landscape above 1300 m a.s.l., before giving way to bare rock, gravel and permanent ice/snow in the nival zone above 2150 m a.s.l. New Zealand’s highest peaks and most mountainous and glacial terrain are found in this region, with all but one of the country’s 37 peaks higher than 2900 m a.s.l. occurring within 16 km of Aoraki/Mount Cook (3724 m a.s.l.), New Zealand’s highest mountain.

Map of the Te Manahuna Aoraki Project area with bounding landforms of the Ben Ohau Range (BO) and Two Thumb Range (TT) marked with bold, dashed lines, and the Main Divide (MD) marked with a bold, solid line and initials in black circles. The internal ranges of Mount Cook Range (MC), Malte Brun Range (MB), Liebig Range (L), Burnett mountains (BM), Gamack Range (G), Hall Range (H), Sibbald Range (S) and the range south of Graf Spee (GS) are marked with thin, grey lines and initials in white circles. Internal valleys of the Hooker (H), Tasman (T), Murchison (Mu), Jollie (J), Forks/Fraser (F), Cass (C), Mistake (Mi), Godley (G), Macaulay (Ma) and North East Gorge (N) are marked with their respective initials
General approach
The aim of this study was to determine whether high-elevation landforms limit the movement of invasive small mammal species, and if so, to provide examples of how high-elevation landforms create defendable areas. Instead of attempting to designate landscape features as ‘hard barriers’ which, by definition, completely prevent the physical passage of animals by presenting an insurmountable or impermeable landform, we took a more conservative and quantifiable approach: we assumed that members of all species were physically capable of passing through any landscape features present and investigated whether high-elevation landforms segregate tracts of suitable habitat by presenting tracts of extremely low-to-zero value habitat, thereby limiting the movement of the populations across the landform. The movement-limiting mechanisms of extremely low-to-zero value habitat are assumed to function in multiple ways:
-
Extremely low-to-zero value habitat bounds a settled individual’s home range, as large tracts of extremely low-to-zero value habitat would be too energetically costly to regularly traverse in order to access other areas containing valuable habitat.
-
Extremely low-to-zero value habitat prevents home ranges from shifting as established individuals would not become aware of, or could not access, segregated patches of valuable habitat because exploratory foraging forays would be unlikely to cross extremely low-to-zero value habitat to discover new areas and add them to an animal’s cognitive map of their surroundings (Gautestad 2011).
-
When searching for new areas of habitat, dispersing individuals that encounter extremely low-to-zero value habitat are more likely to disperse in the direction of and ultimately settle in what they perceive to be more suitable habitat, and less likely to move through extremely low-to-zero value habitat.
We defined extremely low-to-zero value habitat as either terrain above the highest known upper elevation limit of a species, or terrain with an occupancy probability prediction of less than 0.05 for a species. We used these criteria to identify landforms in the upper Mackenzie Basin that limit the movement of the invasive species that occur in this region.
Camera trapping
To produce occupancy models (MacKenzie et al. 2002), we used data from Foster et al. (2021a). In that study, they conducted camera trap surveys over the summer/autumn periods (January to May) of 2019 and 2020 across 185 sites. Camera sites were positioned in a stratified random design across eight landcover types (rock/gravel, alpine herbfield, tall tussock grassland, subalpine shrubland, beech forest, matagouri grassland, low-producing grassland and riverbed) and spanned an elevation range of 500 to 2250 m (i.e., from the lowest valley floor to the permanent snow line). We included data from 10 of the 12 invasive species in New Zealand (excluding the ship rat, Rattus rattus, and Kiore, Rattus exulans, which do not occur in this region). These were the Norway rat (Rattus norvegicus), house mouse (Mus musculus, hereafter ‘mouse’), stoat (Mustela erminea), least weasel (Mustela nivalis, hereafter ‘weasel’), ferret (Mustela furo), European hedgehog (Erinaceus europaeus, hereafter ‘hedgehog’), brushtail possum (Trichosurus vulpecula, hereafter ‘possum’), feral cat (Felis catus, hereafter ‘cat’), European rabbit (Orcytolagus cuniculus, hereafter ‘rabbit’), and European hare (Lepus europaeus, hereafter ‘hare’). Because there were insufficient camera data for the Norway rat (two records), it was not possible to determine occupancy or to confidently designate upper elevation limits for this species, and so this species was not included in our analysis of movement barriers.
Occupancy modelling and model averaging
We used the presence/absence detection histories of Foster et al. (2021a), with assigned detection and site-level covariates to each of the 31 sampling occasions. We built a set of candidate models for each species using all possible combinations of the four detection covariates: ‘effort’, ‘camera type’, ‘days since site service’ and ‘period’, and the three site-level covariates: ‘elevation’, ‘landcover type’ and ‘normalised difference vegetation index’ (hereafter referred to as NDVI) which represented vegetation productivity. The eight landcover types were assigned dummy variables to permit the spatial projection of a categorical covariate. As permanent ice/snow was not sampled during camera trap surveys, this landcover type was designated as rock, despite the likelihood of it being lower value habitat to all species. We included only the most parsimonious function term of continuous covariates based on the model ranking reported by Foster et al. (2021a).
We then used model averaging in the Multi-Model Inference package ‘MuMIn’ (Burnham and Anderson 2002) in RStudio software (RStudio Team 2020) to make spatial predictions of occupancy probability. Model averaging considers a list of candidate models and computes a weight for each individual model based on how well it fits the data. We used Akaike’s Information criterion (Akaike 1974) corrected for small sample sizes (AICc) to rank the list of candidate occupancy models built with the ‘unmarked’ package (Fiske and Chandler 2011) from best to worst fit, and all models with delta AICc values < 100 were included in model averaging. The model-averaging process weighted the effect of each individual detection and occupancy covariate based on the sum of weights of models in which each covariate features, and assigned to each covariate a weight between 0 and 1. We used ‘full’ beta coefficients of the resulting model average rather than the ‘subset’, as the latter only considers the effect of each covariate in models in which it features and potentially biases the effect of weak covariates away from zero.
Spatial projection of occupancy probabilities
Next, we projected model-averaged occupancy probabilities for each invasive mammal species in the Te Manahuna Aoraki project area. Environment data comprised of an 8 m digital elevation model raster (Land Information New Zealand, 2012), a 20 m NDVI raster from composite sentinel-2 A2A cloud-free imagery from January 2nd and March 12th 2020 (Copernicus Sentinel data [2020], processed by ESA) using the raster calculator function in QGIS software (QGIS.org 2021) and a 20 m landcover type raster made with the semi-automatic classification plugin (Congedo 2020) in QGIS software which also used the composite sentinel-2 A2A cloud-free imagery. The three environment rasters were aligned at a 20 m × 20 m pixel size using the nearest-neighbour resampling method of the QGIS raster alignment tool. The model-averaged projections for each species were created using the vignette provided by Chandler (2019) for producing distribution maps of unmarked occupancy models. This resulted in each 20 × 20 m pixel within the 310,000 ha area being assigned an occupancy probability estimate for each of the 10 invasive mammal species.
Model validation
We then assessed model performance for each species using Continuous Boyce Indices (CBI), which test predicted values using independent presence-only records (Boyce et al. 2002; Hirzel et al. 2006). This analysis produces a value between −1 and 1, depending on how well model predictions match real data collected in the local area. Values close to zero indicate that model predictions are random, negative values indicate that model predictions are worse than random, and positive predictions indicate that the model predictions are consistent with real-life data: a value of 1 representing perfect model fit. CBIs compute a Spearman’s rank correlation coefficient between a gradient of habitat suitability and an F-ratio, which is a proportion of predicted data points (test data) compared to expected data points (typically randomly generated pseudo-absences in the immediate landscape) within a moving class window, which by default is 1/10th of the habitat suitability range.
We used two types of independent test data: global positioning system (hereafter GPS) tracking data from previous and recent studies in the local area (available for the cat, hare and hedgehog); and capture data from kill-trapping networks in the Te Manahuna Aoraki area (all other species). To produce expected data points in the immediate landscape where GPS data were collected, we randomly generated 10,000 pseudo-absences (Barbet-Massin et al. 2012) for each species within a buffer of the presence test data for each species in question. For test data derived from GPS location fixes of highly mobile species, the hare (27 individuals, Foster, unpublished data) and cat (26 individuals, Recio et al. 2010, 2014), we generated pseudo-absences in polygons with a 2 km buffer around all data points. We used a smaller buffer of 1 km around GPS location fix test data of the hedgehog (n = 26, Foster, unpublished data) as this species is less mobile. GPS data are ideal for model evaluation as tracked animals are free to move and select/avoid habitat in their immediate area. The fix rate for the GPS-tracked hares was set at 4- to 5-h intervals, whereas the fix rate for the tracked hedgehogs and cats were 5- and 15-min intervals. To avoid autocorrelation, GPS fix data of the tracked hedgehogs and cats were subset to 2-h intervals.
Instead of using randomly-generated pseudo absences for the trapping data of the remaining pest species, we used all trap locations as the background ‘expected data’, which directly compared the presence data to expected data derived from locations where there was an opportunity to record a species. This was done because trap devices are typically placed at locations where they are likely to encounter the target species, resulting in a reduced opportunity to record species in areas that are perceived to be unsuitable. Additionally, trap lines are often established in easily accessed areas, with trappers avoiding features such as tracts of dense scrub or overly-steep areas. Doing so may result in a reduced opportunity to record some species in their preferred habitats and would likely bias the F-ratio of higher habitat suitability towards zero. Extensive trapping networks within the Te Manahuna Aoraki project area cover a variety of landcover types, varying degrees of vegetation productivity, and a wide elevation range, and provide good background pseudo-absence data for most species. However, there was a lack of test data for the possum (Trichosurus vulpecula) owing to their large body size and the small trap openings of common predator traps (60 × 60 mm and 80 × 80 mm), and an overlap of high-density possum populations with the distribution of an inquisitive native parrot, kea (Nestor notabilis), prohibiting the use of any trap that put kea at risk.
Identifying and evaluating movement barriers
First, we investigated how each species’ upper elevation limit (hereafter abbreviated to ‘UEL’) related to the elevation of the bounding landforms. We sampled the elevation every 1 m of an 8 m resolution digital elevation model raster (Land Information New Zealand, 2012) along the 212 km length of the combined Ben Ohau, Main Divide and the Two Thumb Range using QGIS software (QGIS.org 2021). Each measurement point was assigned as being either above or below the elevation of the highest known record for each species of interest, and was plotted using the geom_raster function in ggplot2 (Wickham 2016) from the southernmost limit of the Ben Ohau Range boundary (west; Fig. 1) to the southernmost limit of the Two Thumb Range boundary (east). We used the highest elevation records reported by Foster et al. (2021a) as these were the highest known records of invasive small mammals for the central eastern region of New Zealand’s Southern Alps. However, we raised the upper elevation limit of the rabbit to 1300 m based on anecdotal observations of rabbits up to this elevation in this region.
Second, we produced occupancy probability estimates along the apex of the bounding landforms of the study area to identify areas of extremely low to zero value habitat. We designed this analysis to discriminate between sections of the bounding landforms that supported widespread or continuous high-quality habitat, and sections with patchy and poor/zero quality habitat. We sampled predictions of occupancy probability within a 500 m-wide strip along the apex of the Ben Ohau, the Main divide and the Two Thumb Ranges. We divided the 212 km-long chain of mountains into 500 m lengths and calculated the mean pixel value in each of the 424 sampling polygons. A sampling polygon size of 500 × 500 m was used as it encompassed a large number of raster pixels for which a mean could be calculated without sampling too far from the apex of the mountain range, as doing so would sample lower-elevation terrain in areas where mountain ranges have high vertical relief. We took the 540 pixels with the highest occupancy values in each sampling polygon and plotted them from bottom to top in descending occupancy probability and arranged sampling polygons from the western end to the eastern end of the chain of mountains left-to-right using the geom_raster function in ‘ggplot2’ package (Wickham 2016). We overlayed a line of the mean pixel value of each sampling polygon on this raster, allowing the mean of each sampling polygon to be expressed while providing a visual representation of the composition of each sampling polygon. Mean occupancy probability of sampling polygons along the length of the boundary was evaluated as very low (< 0.25), low to moderate (0.25–0.50), moderate to high (0.50–0.75), and very high (> 0.75).
Third, we combined information on the upper elevation limit of species with the spatial projection of their occupancy probabilities to identify defendable areas within the Te Manahuna Aoraki project area. This was done by overlaying a line of each species’ upper elevation limit on a spatial projection of occupancy probability across the Te Manahuna Aoraki project area. Terrain that was above the upper elevation limit of a species and/or had an extremely low occupancy probability in (< 0.05) across substantial areas (> 500 m wide) was considered to significantly limit the movement of the species in question. Using either criterion or both, we identified ‘movement barriers’ and mapped these along the Te Manahuna Aoraki project boundary and internal ranges, for each species of interest.
Finally, as a test of our movement barrier predictions, we overlaid hedgehog GPS tracks collected at mid-to-high elevations (1200–1950 m a.s.l) in one internal valley (Mistake) and one external boundary (Two Thumb Range) with occupancy probability projections and lines of hedgehog UELs. Small GPS backpacks were made from i-gotU GT120 GPS travel loggers (Mobile Action Technology, Taipei, Taiwan) and 11 g VHF transmitters (Lotek V2G 152 C) and were attached to spines on the backs of hedgehogs with hot-melt adhesive. Stationary tests revealed these devices to have a mean location error of 4.17 m (± 0.10) on bare ground, and 5.28 m (± 0.17) in dense tussock (Foster et al. 2021b). GPS movement tracks of 26 individual hedgehogs were acquired across two 6-month periods (December–June). Each individual was tracked for an average of 42 days (SD = 20.89) with fixes recorded every 5 min from dusk until dawn, the hedgehog being a nocturnal animal (72,856 locations in total, Foster, unpublished data). We investigated how this movement data interacted with predicted movement barriers.
Results
Upper elevation limits
The upper elevation limits (UEL) of the 10 species varied greatly (Table 1). The UEL of the rabbit, weasel and ferret fell nearly entirely below the highest elevations of the three bounding mountain ranges, with only a short length of the terminus of the barrier (< 500 m) being lower than the UEL of the distributions of the weasel and ferret (Table 1). The higher UEL of the hedgehog resulted in 16.7 km (7.9% of the total bounding landform length) being lower than hedgehog UEL. These areas were mainly situated at the ends of the proposed barrier and in several occupied regions along the Two Thumb Range (Fig. 2). The same areas were occupied by the mouse, possum, cat, stoat and hare, but instead of being confined to lower passes, occupied sections were increasingly wide and connected with increasing UELs. Nearly the entire length of the Two Thumb Range fell below the UEL of the species with the highest distribution limits (cat, stoat and hare). The higher elevation of the Ben Ohau Range resulted in fewer occupied areas, which were concentrated around the heads of the major catchments (Fig. 2). The Main Divide appeared to be almost completely secure for all species, with only a small number of low points falling below the UEL of the mouse, possum, cat, stoat and hare (Fig. 2).

A visualisation of whether the elevation at the apex of the bounding landforms fell above (purple) or below (yellow) the upper elevation limit of each species of interest. Data are presented left-to-right from the western terminus of the landscape barrier formed by the Ben Ohau Range to the terminus on the eastern side formed by the Two Thumb Range
Occupancy probability estimates
Model-averaged estimates of occupancy probability of the rabbit, weasel and ferret indicated very low probability of occurrence along all three of the mountain ranges that bound the Te Manahuna Aoraki project area (Fig. 3). Model-averaged occupancy models of the hedgehog identified the same barriers as the UEL approach, but predicted a near-zero probability of occurrence on the glaciated main divide. There was a low probability of hedgehog occurrence at a number of sections of the Two Thumb Range (Fig. 3). Although presenting low mean occupancy probabilities, these polygons did include pixels with moderate probabilities of hedgehog occupancy.
Mouse occupancy probability along the TMA boundary varied between the three ranges. Averaged values along the Ben Ohau Range fluctuated between 0.20 and 0.25 with higher values predicted for lower elevation areas at the heads of the major catchments and a clear heightened probability of occurrence at Barron Saddle. The Main Divide was found to have very low probabilities of mouse occupancy along its length (Fig. 3). Aside from the elevated terrain surrounding The Thumbs (2546 m), the probability of mice occurring along the apex of the Two Thumb Range was consistently low/low to moderate. The possum and cat were predicted to have near-identical landform barriers, with probability of possum occupancy generally being slightly higher than that of the cat. For both species, there were isolated points along the Ben Ohau Range that had slightly raised but very low probabilities of occurrence. The Barron Saddle and Copland Pass areas were identified as presenting terrain with very low probabilities of occurrence, and between Copland Pass and Mt Huss there was a near-zero probability of occurrence for both species (Fig. 3). The lower elevation areas of The Two Thumb Range presented terrain with the highest likelihood of possum and cat occurrence. While this did not exceed a mean pixel value of 0.25 occupancy probability, it did present occasional pixels with moderate (0.50) occupancy for possums.
Model-averaged predictions of stoat occupancy probability heavily weighted ‘landcover type’ (0.99) and ‘NDVI’ (0.90) but only lightly weighted ‘elevation’ (0.06, Table 2). With negligible weighting of ‘elevation’ in model-averaged predictions, the dominant ‘landcover type’ factor of rock (which included permanent ice/snow) at high elevations resulted in a widespread moderate-to-high stoat occupancy probability of ~ 0.60 along the mountain boundary (Fig. 3). In individual model suites in Foster et al. (2021a), the probability of a hare occurring at a site peaked at ~ 1250 m a.s.l. and steadily decreased but only reached very low probabilities of occurrence above 2500 m. This resulted in an undulating low to moderate probability of hares occurring at the apex of the Ben Ohau Range, very low occupancy probabilities along the high-elevation landforms of the Main Divide, and a moderate to high probability along the lower elevation sections of the Two Thumb Range (Fig. 3).
The quality of predictions, as indicated by Continuous Boyce Indices (Table 2), ranged from worse than random (possum = − 0.232), random (stoat = 0.006), good (weasel = 0.310, mouse = 0.482, hedgehog = 0.531, ferret = 0.615) to very good (rabbit = 0.772, cat = 0.853, hare = 1.000).

Model-averaged occupancy estimates at the apex of bounding landforms per species, presented as the mean cell value per 500 m sampling polygon overlaying the stacked cell values from highest value (bottom) to lowest value (top). Data are presented left-to-right from the western terminus of the landscape barrier formed by the Ben Ohau Range, to the terminus on the eastern side, formed by the Two Thumb Range
Identification of movement barriers and defendable areas
Both the UEL and OP < 0.05 approaches indicated that the entire TMA boundary and internal ranges could be considered as strong movement barriers to the rabbit, weasel and ferret (Figs. 4a–c, 5a–c), and identified numerous defendable areas. Movement barriers for the rabbit extended down to lower altitudes (~ 1300 m a.s.l.), whereas the lower extents of landform barriers for the ferret and weasel were shortened. Hedgehog UEL fell close to where < 0.05 occupancy probabilities were predicted, so either approach was useful in identifying movement barriers. The Ben Ohau Range, the Main Divide and almost all major internal ranges were predicted to be excellent movement barriers to the hedgehog, but there were many areas of favourable habitat along the apex of the Two Thumb Range (Figs. 4d, 5d).
The UELs of the mouse and possum were used to identify movement barriers, as model predictions where OP < 0.05 were limited to small areas of high elevations of the Upper Tasman and Murchison valleys. Predicted movement barriers and defendable areas for these two species were nearly identical, owing to their similar UELs (2053 and 2079 m). The Ben Ohau Range was predicted to severely limit the movement of both species, with a small occupied area at the head of Fred’s Stream (Figs. 3, 4e, f, 5e, f). The Main Divide was identified as a movement barrier to both the mouse and possum, with a small occupied area at Barron Saddle and at the head of the Godley River (Fig. 2). In contrast, apart from high-elevation areas of Mt D’Archiac and The Thumbs massifs, the Two Thumb Range section of the bounding landforms was unlikely to limit their movements (Fig. 5e, f). Several internal ranges appeared to present significant movement barriers to these two species, including the Mount Cook Range, the Malte Brun Range and the Liebig Range (Fig. 5e, f). Other ranges presented long lengths of terrain that were above their UELs, but were dissected by lower terrain where higher rates of movement might be expected (Fig. 5e, f).
The UELs of the stoat, cat and hare were predominantly used to identify movement barriers for these species, with cat and hare occupancy predictions < 0.05 only occurring at high elevations along the Main Divide. Much of the Ben Ohau Range was identified as a movement barrier to these three species, but lengths of barriers were segmented as a result of low elevation passes at the heads of most valleys (Fig. 5g–i). These gaps were widest for the hare, the species with the highest UEL recorded in camera trap surveys (Foster et al. 2021a). Terrain along the apex of the Main Divide was almost entirely above the UELs of the cat, stoat and hare, with short occupied areas identified around Copland pass and the head of the Godley River (Fig. 5g–i). The Two Thumb Range had the lowest elevation terrain, with only ranges north of Mt D’Archiac and the immediate ranges surrounding The Thumbs presenting terrain above the UELs of these three species. While several internal ranges reached elevations above their UELs, all identified movement barriers except the Mount Cook Range, the Malte Brun Range and the Liebig Range were extremely disjointed, which indicated that only the Hooker, upper Tasman and Murchison Valleys could be defended using continuous high-elevation landforms for these three species.

Predictions of movement barriers based on occupancy probabilities < 0.05 (green) of each species of interest. Plots are arranged in increasing order of upper elevation limit

Predictions of movement barriers based on terrain above the upper elevation limits of each species of interest (green). Plots are arranged in increasing order of upper elevation limit
Model test: European hedgehogs
The UEL and predictions of hedgehog occupancy probability at 0.05 were very similar, with occupancy probability predictions < 0.05 slightly expanding what could be considered as a movement barrier (Figs. 4d, 5d). During the GPS study periods, all movements of adult (n = 20) or juvenile (n = 6) hedgehogs supported our predictions of landforms that significantly limit dispersal (Fig. 6). In total, there were only 3 of 72,856 location fixes that occurred in OP 0.00-0.05 terrain. These resulted from one individual that appeared to move into an area of extremely low-value habitat before returning to higher value habitat and navigating around it (Fig. 7a), though it is possible that the offending fixes could have resulted from GPS location error. Slightly higher value terrain of OP 0.05–0.10 was also minimally used (693 of 72,856 location fixes, < 1% of total fixes), mostly cases where hedgehogs either encountered the low value habitat and went no further (Fig. 7c), traversed short distances to link habitat patches (Fig. 7a–c) or repeatedly returned to den sites via a ‘short cut’, as illustrated by the cluster of points in Fig. 7a. Hedgehog movements were often obstructed by small streams, in some cases as narrow as 1 m across. These were seldom, if ever, crossed and had a strong influence on the shape of home ranges on valley floors (Figs. 6, 7c). Eight of the 26 tracked hedgehog regularly interacted with the edges of waterways and only three of these were recorded to have made a single return crossing each: this equated to three days during which a return crossing occurred across a summed total of 442 hedgehog tracking days.

Hedgehog occupancy predictions in the Mistake valley (‘Mi’, Fig. 1) and the Two Thumb Range (inset) with occupancy probability estimates < 0.05 indicated in green at 70% opacity overlaying a hillshade surface. The UEL of hedgehogs (1937 m) is marked with a dashed white line, and GPS movement data of a sample of hedgehogs (n = 26) are displayed with solid white lines

Examples of hedgehog GPS fixes that occurred in habitat with hedgehog occupancy probability predictions of 0.00–0.05 as red points (A only) and of 0.05–0.10 as blue points (A–C). White lines indicate chronological movement between GPS data points
Discussion
High-elevation landforms as movement barriers
Our findings indicate that high-elevation landforms significantly limit the movement of most invasive small mammal species in New Zealand. We demonstrated that many significant landforms of the Southern Alps and associated ranges reach elevations beyond the highest known occurrence of a species, presented large tracts of extremely low-to zero value habitat, or both, and provided examples of how movement-limiting landforms create defendable areas that can be exploited by landscape-scale eradication programmes. The distributions of species which had associations with low elevation areas (rabbit, weasel, ferret) or had narrow habitat preferences at higher elevations (hedgehog), were limited by high-elevation landforms to such an extent that many landforms might be considered to prevent the movement of individuals in all but the most exceptional circumstances.
Our methods predicted large differences in the ability of various landforms to limit the movement of invasive species, ultimately due to differences in elevation and vegetation. Landforms such as the Two Thumb Range were predicted to allow movements of the majority of invasive mammal species because they are lower elevation (between 1900 and 2100 m) and because the low saddles and passes have contiguous tall tussock grasslands and alpine herbfields. In contrast, the Main Divide formed a near continuous barrier to movements because it had high elevations of > 2500 m and near-complete absence of any vegetation near the apex of the landform. The distributions of species that have close habitat associations with the limits of vegetation (i.e. hedgehog) would likely be severely limited by the absence of vegetation at high elevations, often over many kilometres before the apex. The distributions of species that do not appear to have strong associations with vegetation, e.g., stoats, may be strongly influenced by the distribution and availability of prey items upon which they rely. A recent exploration of the dietary niche of the stoat in New Zealand’s alpine zones revealed that they primarily consumed mammals (rats, mice and hares) but seasonally shifted to rely more heavily on insects, and in some locations, skinks formed a substantial portion of their diet (McAulay et al. 2020). For stoats to fulfil their high metabolic requirements, they would need to remain in relatively close proximity to these food sources, and dwindling vegetation biomass with increasing elevation might translate into lower prey biomass for stoats as only a small number of invertebrate species specialise in the barren habitats of the nival zone (Mark 2012; Chinn and Chinn 2020).
Species that are resident in the alpine zone with very limited movement potential (e.g. mouse; MacKay et al. 2011) are likely obstructed by unsuitable areas because the movement of individuals requires relatively continuous available resources of food and shelter. Highly mobile species that occur frequently at high elevation, such as the stoat and hare, are likely less limited by high-elevation landforms near the limit of their altitudinal distribution as they are capable of linking habitat patches separated by greater distances of unsuitable terrain. In comparison, while some species have the capability to move large distances and do occur in the alpine zone, they may be less likely to cross high-elevation landforms because they typically either have home ranges centred in mid to low areas, as indicated by GPS tracking (cat; Recio et al. 2014) or are infrequently detected at high-elevation sites. In cases where a landform does not significantly limit a species’ distribution but where habitat becomes less suitable along a gradient of an environmental variable (e.g. the possum having a preference for higher vegetation structure, which decreases with increasing elevation) that limits a species’ abundance, the number of individuals moving across a landscape feature, or ‘dispersal pressure’ would be lower, even if physical habitat of some value is continuous across the landscape feature.
We based our investigation of landforms that limit movement on the connectivity or segregation of suitable habitat. It is important to consider that dispersing animals do not necessarily demonstrate the same behaviour as settled animals (Zalewski et al. 2009) and that predictions of habitat suitability are often based on data collected on settled individuals. Without a deep understanding of a species’ dispersal behaviour, including an individual’s intrinsic motivations to move large distances away from genetically similar individuals, search for mates, escape high or low population densities, or abandon areas of limited resources, then even the strongest evidence of landscape barriers should be treated as potentially only limiting dispersal-type movements rather than preventing them. Identifying exactly what is an effective ‘hard barrier’ to a particular species is problematic and requires not only intimate knowledge of the physical limitations of the species in question, but also for researchers to be able to accurately quantify the limiting characteristics of landscape features of interest. Our analyses quantified strong predictors of the presence or absence of a species—the habitat it uses, and at what elevations it occurs. We used these predictions to explore how valued habitat was fragmented by habitat that we identified to be of extremely low value, and thereby limiting movement between fragmented patches on the basis of several mechanisms. It is likely that animals perceive habitat along a gradient of suitability (Manning et al. 2004) and select habitat on the basis of either pre-dispersal habitat imprinting, where an animal searches for habitat similar to where it developed in the early stages of life, or conspecific attraction, where the presence of members of its own species indicates suitable habitat and increases the probability that it will settle (Stamps 2001). Thus, predictions of habitat preferences of established animals are likely to be useful in predicting movement directions and indicating high to low movement permeability. Because data used in this study were collected across a wide time period (January–May), it is likely that dispersing individuals were detected and in turn influenced modelling of habitat suitability, and so predictions of occupancy probability might go some way to approximate actual movement permeability for all species studied. We gave an arbitrary barrier width of > 500 m as presenting a substantial barrier for all small mammal species studied: the true effective width is bound to differ between species relative to their movement potential. Further exploration, such as GPS movement studies as has been done for the hedgehog, or testing for genetic divergence, is required to confirm/refine our understanding of the permeability of landforms for all species of interest.
The limitations of our methods notwithstanding, the case study of the European hedgehog demonstrated the efficacy of using habitat suitability to evaluate landforms that severely limit dispersal. The occurrence of location fixes in habitat with occupancy probabilities between 0.05 and 0.10 indicate permeability where patches of suitable habitat are intermittent across low value habitat (Fig. 6). An occupancy probability of 0.05 was used as an arbitrary cut-off, and it is expected that the same types of movement illustrated in Fig. 7b would occur across zero value habitat if there are intermittent patches of higher value habitat that a hedgehog might seek. Hedgehog movements indicate that the dispersal-limiting mechanisms of the edges of vegetated patches bounding home ranges and discouraging further exploration would only apply where there are large tracts of continuous low-to-zero habitat. However, the instances where 0.05–0.10 occupancy probability habitat was used by the tracked hedgehogs suggested that the lower limit of habitat occupancy probability for hedgehogs is greater than 0.05, effectively extending the areas acting as movement barriers to hedgehogs. This is illustrated in Fig. 7a, where a hedgehog’s movements are neatly bound by a scree slope that was assigned an occupancy probability value of 0.08. We have shown that the hedgehog has relatively narrow habitat requirements in this environment. In addition, juveniles do not disperse great distances from their maternal dens (Sæther 1997; Rasmussen et al. 2019), they are not territorial (Jones and Sanders 2021), and are therefore less likely to disperse in unpredictable ways. These features of hedgehog ecology make them very promising candidates to defend against using high-elevation landscape features.
Suitability of criteria for identifying high-elevation landscape barriers
The two criteria that we used to identify movement barriers were useful and often coincided, but did have clear limitations. The UEL of a species does not take into account suitable habitat above the limit, nor the absence of suitable habitat below the limit of a species’ distribution. It also relies on the assumption that the information that we used is the true habitat use and upper limit to their distributions, which is undoubtedly not the case as exceptional movements are bound to occur for all species. Indeed, there are several anecdotes of the local area that demonstrate this, such as mountaineers discovering the frozen body of a possum at the head of the Tasman Glacier (~ 2500 m a.s.l.), observations of a hedgehog crossing the lower Tasman Glacier, and a sighting of a cat on the Murchison glacier (Neil Bolton, pers. comm.). However, the UELs that we used are representative of normal distribution limits, being based on extensive camera trap surveys across two summer/autumn periods, and are appropriate for use in determining movement-limiting landforms as leaking barriers.
There were several limitations of using occupancy probability estimates to identify dispersal-limiting landforms: ‘permanent snow/ice’ was not sampled and instead was treated as ‘rock’, which likely resulted in overestimation of habitat quality and therefore movement permeability across high-elevation landforms with glaciated terrain; over-weighting of covariates in the model-averaging process resulted in poor model performance; and, because of a lack of devices opportunity to record the absence of individuals above 2300 m a.s.l., the occupancy probability estimates at high elevations for species such as stoats are likely overestimates. An advantage of using occupancy probability estimates was that it was possible to discriminate between areas that fell within or outside of what we identified as a barrier. For example, while there were lengths of both the Ben Ohau Range and the Two Thumb Range that were not considered barriers for the hedgehog, possum and cat, sampling, averaging and visualising occupancy probabilities revealed that the probability of these species occurring in these areas was much higher in the Two Thumb Range than in the Ben Ohau Range.
Implications for management
Our findings have a significant application in landscape-scale programs for eradication of invasive small mammals in New Zealand. We provide the first comprehensive examination of how high-elevation landforms can be used to defend against reinvasion of invasive small mammals. The major landforms of the entire intermontane area of the upper Mackenzie Basin were shown to segregate populations of the majority of the studied species, demonstrating the utility of such landforms in eradication programmes. Predictions such as those presented here can identify defendable boundaries and inform strategic rollout of landscape-scale eradication programmes in mountainous habitats elsewhere in New Zealand. In practice, a pest population may be removed from within an area that is defended from reinvasion by its bounding high-elevation landforms. Incursions from the ‘open’ area down-valley would be managed by maintaining a buffer zone of lethal devices until effort can made to remove the reinvading population, and so on. Given the size and extent of the Southern Alps, it is likely that its landforms present one of the most substantial opportunities to create defendable areas on mainland New Zealand, in which the eradication of invasive species could be achieved and maintained. Programmes that aim to suppress rather than eradicate populations of invasive small mammals may also apply this strategy, as removing individuals from areas with low recolonisation rates would result in lower predator population densities, reduced reinvasion potential and therefore more efficient ongoing management costs. Globally, eradication strategies for removal of unwanted pests as part of biosecurity programmes may benefit from applying similar elevation and occupancy modelling approaches, and this may prevent wasted resources resulting from setting up poorly defined or difficult to defend treatment zones.
Data availability
All data are available upon reasonable request.
References
Akaike H (1974) A new look at the statistical model identification. IEEE Trans Autom Control 19:716–723
Barbet-Massin M, Jiguet F, Albert CH, Thuiller W (2012) Selecting pseudo-absences for species distribution models: how, where and how many? Methods Ecol Evol 3:327–338
Bellingham PJ, Richardson SJ, Gormley AM, MacLeod CJ, Forsyth DM (2013) Department of Conservation biodiversity indicators: 2013 Assessment. Landcare Research
Boyce MS, Vernier PR, Nielsen SE, Schmiegelow FKA (2002) Evaluating resource selection functions. Ecol Model 157:281–300
Burnham KP, Anderson DR (2002) Model selection and multimodel inference: a practical information-theoretic approach, 2nd edn. Springer, New York
Burns B, Innes J, Day T (2012) The use and potential of pest-proof fencing for ecosystem restoration and fauna conservation in New Zealand. In: Fencing for conservation, pp 65–90
Caplat P, Edelaar P, Dudaniec RY, Green AJ, Okamura B, Cote J, Ekroos J, Jonsson PR, Löndahl J, Tesson SVM, Petit EJ (2016) Looking beyond the mountain: dispersal barriers in a changing world. Front Ecol Environ 14:261–268
Chandler R (2019) Modeling and mapping species distributions. https://cran.r-project.org/web/packages/unmarked/vignettes/spp-dist.pdf
Chinn WGH, Chinn TJH (2020) Tracking the snow line: responses to climate change by New Zealand alpine invertebrates. Arct Antarct Alp Res 52:361–389
Christie JE, Wilson PR, Taylor RH, Elliott G (2017) How elevation affects ship rat (Rattus rattus) capture patterns, Mt Misery, New Zealand. N Z J Ecol 41:113–119
Clout MN, Gaze PD (1984) Brushtail possums (Trichosurus vulpecula Kerr) in a New Zealand beech (Nothofagus) forest. N Z J Ecol 7:147–155
Coleman JD, Green WQ, Polson JG (1985) Diet of brushtail possums over a pasture- alpine gradient in Westland, New Zealand. N Z J Ecol 8:21–35
Cook B, Mulgan N, Nathan H (2021) Rivers as obstacles to home range expansion by the brushtail possum. N Z J Ecol 45:1–6
Coyne JA, Orr HA (2004) Speciation. Sinauer Associates, Sunderland
Cross CC (2016) Spatial ecology and population genetics of cats (Felis catus) living in or near conservation-sensitive areas. University of Otago
Fiske IJ, Chandler RB (2011) Unmarked: an R package for fitting hierarchical models of wildlife occurrence and abundance. J Stat Softw 43:1–23
Foster NJ, Maloney RF, Seddon PJ, Recio MR, Khan MSI, van Heezik Y (2021a) Altitudinal distribution of the entire invasive small mammal guild in the eastern dryland zone of New Zealand’s Southern Alps. Biol Invasions 23:1837–1857
Foster NJ, Maloney RF, Recio MR, Seddon PJ, van Heezik Y (2021b) European hedgehogs rear young and enter hibernation in New Zealand’s alpine zones. N Z J Ecol 45:1–6
Fraser EJ, Macdonald DW, Oliver MK, Piertney S, Lambin X (2013) Using population genetic structure of an invasive mammal to target control efforts—an example of the American mink in Scotland. Biol Conserv 167:35–42
Gautestad AO (2011) Memory matters: Influence from a cognitive map on animal space use. J Theor Biol 287:26–36
Glen AS, Byrom AE, Pech RP, Cruz J, Schwab A, Sweetapple PJ, Yockney I, Nugent G, Coleman M, Whitford J (2012) Ecology of brushtail possums in a New Zealand dryland ecosystem. N Z J Ecol 36:29–37
Haran J, Roques A, Bernard A, Robinet C, Roux G (2015) Altitudinal barrier to the spread of an invasive species: could the pyrenean chain slow the natural spread of the pinewood nematode? PLoS ONE 10:1–25
Hirzel AH, Le Lay G, Helfer V, Randin C, Guisan A (2006) Evaluating the ability of habitat suitability models to predict species presences. Ecol Model 199:142–152
Innes J, Fitzgerald N, Binny R, Byrom A, Pech R, Watts C, Gillies C, Maitland M, Campbell-Hunt C, Burns B (2019) New Zealand ecosanctuaries: types, attributes and outcomes. J R Soc N Z 49:370–393
Jones C, Sanders MD (2021) European hedgehog. In: King CM (ed) The handbook of New Zealand mammals, 3rd edn. Otago University Press
Jones HP, Holmes ND, Butchart SHM, Tershy BR, Kappes PJ, Corkery I, Aguirre-Muñoz A, Armstrong DP, Bonnaud E, Burbidge AA, Campbell K, Courchamp F, Cowan PE, Cuthbert RJ, Ebbert S, Genovesi P, Howald GR, Keitt BS, Kress SW, Miskelly CM, Oppel S, Poncet S, Rauzon MJ, Rocamora G, Russell JC, Samaniego-Herrera A, Seddon PJ, Spatz DR, Towns DR, Croll DA (2016) Invasive mammal eradication on islands results in substantial conservation gains. Proc Natl Acad Sci USA 113:4033–4038
MacKay JWB, Murphy EC, Anderson SH, Russel JC, Hauber ME, Wilson DJ, Clout MN (2011) A successful mouse eradication explained by site-specific population data. Island invasives: eradication and management. 1989:198–203
MacKenzie DI, Nichols JD, Lachman GB, Droege S, Royle JA, Langtimm CA (2002) Estimating site occupancy rates when detection probabilities are less than one. Ecology 83:2248–2255
Manning AD, Lindenmayer DB, Nix HA (2004) Continua and Umwelt: novel perspectives on viewing landscapes. Oikos 104:621–628
Mark AF (2012) Above the treeline: a nature guide to alpine New Zealand. Craig Potton Publishing, Nelson
McAulay J, Seddon PJ, Wilson DJ, Monks J (2020) Stable isotope analysis reveals variable diets of stoats (Mustela erminea) in the alpine zone of New Zealand. N Z J Ecol 44:1–13
Myers JH, Simberloff D, Kuris AM, Carey JR (2000) Eradication revisited: dealing with exotic species. Trends Ecol Evol 15:316–320
O’Donnell CFJ, Weston KA, Monks JM (2017) Impacts of introduced mammalian predators on New Zealand’s alpine fauna. N Z J Ecol 41:1–22
Perrigo A, Hoorn C, Antonelli A (2020) Why mountains matter for biodiversity. J Biogeogr 47:315–325
Rasmussen SL, Jones OR, Berg TB, Dabelsteen T (2019) The ecology of suburban juvenile European hedgehogs (Erinaceus europaeus) in Denmark. Ecol Evol 9:13174–13187
Recio MR, Mathieu R, Maloney R, Seddon PJ (2010) First results of feral cats (Felis catus) monitored with GPS collars in New Zealand. N Z J Ecol 34:288–296
Recio MR, Mathieu R, Virgós E, Seddon PJ (2014) Quantifying fine-scale resource selection by introduced feral cats to complement management decision-making in ecologically sensitive areas. Biol Invasions 16:1915–1927
Robertson BC, Gemmell NJ (2004) Defining eradication units to control invasive pests. J Appl Ecol 41:1042–1048
Robertson PA, Adriaens T, Lambin X, Mill A, Roy S, Shuttleworth CM, Sutton-Croft M (2017) The large-scale removal of mammalian invasive alien species in Northern Europe. Pest Manag Sci 73:273–279
RStudio Team (2020) RStudio: Integrated development for R. RStudio Inc, Boston
Russell JC, Broome KG (2016) Fifty years of rodent eradications in New Zealand: another decade of advances. N Z J Ecol 40:197–204
Russell JC, Stanley MC (2018) An overview of introduced predator management in inhabited landscapes. Pac Conserv Biol 24:371–378
Sæther HM (1997) Overlevelse og tidlig spredning hos juvenile piggsvin (Erinaceus europaeus). MSc thesis. Norges Teknisk-Naturvitenskapelige Universitet
Simberloff D (2009) We can eliminate invasions or live with them. Successful management projects. Biol Invasions 11:149–157
Stamps JA (2001) Habitat selection by dispersers: integrating proximate and ultimate approaches. In: Clobert J, Danchin E, Dhondt AA, Nichols JD (eds) Dispersal. Oxford University Press, New York, pp 230–242
Wickham H (2016) ggplot2: Elegant graphics for data analysis
Zalewski A, Piertney SB, Zalewska H, Lambin X (2009) Landscape barriers reduce gene flow in an invasive carnivore: geographical and local genetic structure of American mink in Scotland. Mol Ecol 18:1601–1615
Acknowledgements
This research was conducted with University of Otago ethics committee approval (AUP-18-104). We thank all who were involved in the collection of trapping and GPS data that were used for model assessment, and are grateful to the landowners who permitted us to conduct this research across their lands. We thank Andrew Bennett and two anonymous reviewers for their thorough examination of our manuscript and feedback which greatly improved this piece of research. This study was supported by Predator Free 2050 Ltd., Te Manahuna Aoraki Project, Miss E.L. Hellaby Indigenous Grassland Research Trust, and a University of Otago Doctoral Scholarship awarded to NJF.
Funding
Open Access funding enabled and organized by CAUL and its Member Institutions. This work was supported by Predator Free 2050 Ltd., Te Manahuna Aoraki Project, Miss E.L. Hellaby Indigenous Grassland Research Trust, and a University of Otago Doctoral Scholarship awarded to NJF.
Author information
Authors and Affiliations
Contributions
All authors contributed to the study conception and design. Data collection and analyses were performed by NJF. The first draft of the manuscript was written by NJF, to which all authors contributed revisions and refinements. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors have no competing interests to disclose.
Ethical approval
This research was conducted with University of Otago animal ethics committee approval (AUP-18-104).
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Foster, N.J., Maloney, R.F., Seddon, P.J. et al. High-elevation landforms limit the movement of invasive small mammal species. Landsc Ecol 37, 2651–2670 (2022). https://doi.org/10.1007/s10980-022-01496-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10980-022-01496-8