
Abstract
Context
Agricultural intensification is contributing to a global species decline. Underlying mechanisms include toxic effects of pesticides on non-target organisms and reductions in habitat and food availability. However, the effects of agricultural intensification on body condition, particularly of ecosystem service-providing arthropods, are poorly understood.
Objectives
Here, we investigated whether variations in the body condition of common lady beetle species (Coleoptera: Coccinellidae) can be explained by the composition and configuration of the surrounding landscape. Assuming strong seasonal variation in food availability in intensively farmed regions, we included the entire period of lady beetle activity in our study.
Methods
Lady beetles were collected from April to September 2011 in 30 landscapes in southern Wisconsin, USA. We examined how body size, body density, and lipid content of the beetles responded to the percentage of intensive cropland, habitat diversity, and edge density in the surrounding landscape.
Results
The strongest predictor of body condition was the percentage of intensive cropland. For every 10% increase in cropland, body density decreased by about 3.9% and lipid content by 6.4%. Landscape diversity and edge density correlated with body condition of individual species.
Conclusions
In agriculturally intensified landscapes, lady beetles with reduced body condition may produce fewer offspring, have lower survival rates, and exert less effective pest control. Thus, our results suggest a mechanistic link between landscape patterns and observed declines in lady beetle populations. Our results also show that the expansion of monocultures affects even common cropland-associated species such as Harmonia axyridis, suggesting a long-term decline in biocontrol services in simplified agricultural landscapes.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Landscape structure affects species’ abilities to move, survive, and persist in their environments (Fahrig 2007; Tscharntke et al. 2012; Gámez-Virués et al. 2015; Martin et al. 2019; Seibold et al. 2019). This is consequential not only for the conservation of biodiversity generally, but also the maintenance of service-providing organisms such as pollinators and natural enemies of crop pests. Agricultural landscape simplification including loss of semi-natural habitat and reduction of crop diversity is often found to have a negative effect on the abundance and diversity of natural enemies and the biological control services they provide (Chaplin-Kramer et al. 2011; Liere et al. 2015; Rusch et al. 2016; Dainese et al. 2019). However, the count data on natural enemy abundance collected in most studies usually cover only one or a few growing seasons (but see Lamb et al. 2019) and cannot adequately capture population trajectories. This may result in incomplete or misleading conclusions about the effects of landscape structure on species responses in the long-term (Kuussaari et al. 2009). The integration of landscape ecology with conservation physiology constitutes a promising direction for resolving such limitations and advancing from pattern- to process-based understanding of species and service distributions (Ellis et al. 2012; Cooke et al. 2013).
Physiological indicators of individual body condition and fitness can offer valuable insights into the status of animal populations in disturbed landscapes before a numerical response is observed. For example, stress biomarkers in birds (Latimer et al. 2020) and limb malformation in amphibians (Guerra and Aráoz 2016) respond to agricultural landscape composition and configuration. For arthropods, characteristics such as body size, mass, and lipid content are important indicators of intraspecific variations in body condition. Larger individuals are more likely to be better competitors for resources and mates, as well as have higher fecundity and reproductive success (Kajita and Evans 2010; Vargas et al. 2013a; Beukeboom 2018). In addition, body size correlates with dispersal ability and thus can strongly influence the response of species and individuals to landscape features (Gámez-Virués et al. 2015; Boetzl et al. 2019). While body size of holometabolous insects is determined by feeding conditions at the larval stage (Hodek 1973; Vargas et al. 2012), body mass is modulated by food availability to adults and is thought to reflect actual nutrient storage (Knapp and Knappová 2013). Lipid content is an important measure of energy reserves, which determine the ability of individuals to survive during periods of resource scarcity (Arrese and Soulages 2009). Direct measurements of body composition, such as lipid content, also tend to be better indicators of body condition than proxy indices such as body size (Knapp and Knappová 2013; Wilder et al. 2016).
Relatively few studies have investigated the consequences of landscape structure on arthropod body condition. Early studies showed that the body size and fecundity of a generalist ground beetle was positively correlated with field perimeter-to-area ratio and percentage of perennial crops across cereal farms (Bommarco 1998, Östman et al. 2001). Effects of landscape structure on body condition have also been demonstrated for wolf spiders (Öberg 2009; Drapela et al. 2013) and dung beetles (Salomão et al. 2018). Flying organisms are highly mobile and thus may be most susceptible to landscape-related drivers. While there is also a growing literature base examining the effects of landscape on body condition of wild and managed pollinators (Renauld et al. 2016; Alaux et al. 2017; Grab et al. 2019; Dolezal et al. 2019; Mogren et al. 2020), surprisingly little is known about the consequences for flying predators.
Lady beetles (Coleoptera: Coccinellidae) are highly mobile predators that provide valuable biological control services in agricultural landscapes (Obrycki et al. 2009). Despite their association with crops, population sizes are often reduced in regions that are dominated by intensive agriculture (Gardiner et al. 2009a, b; Grez et al. 2014; Woltz and Landis 2014). Tiede et al. (2017) showed that the gut microbiome of native and introduced lady beetle species in Wisconsin (USA) varied by species, prey diversity, local habitat type, and landscape context. They also found that landscape context mediated the effect of gut microbiota on beetle lipid content, but the study’s relatively small sample size and qualitative measurement of lipid content makes it difficult to draw strong conclusions and left other aspects of body condition unexplored.
Here we ask whether variation in lady beetle body condition can be explained by features of the surrounding landscape. Specifically, we investigated how body size, body density, and lipid content responded to variation in landscape composition (percentage of intensive cropland, landscape diversity) and configuration (edge density) in agricultural landscapes. In a mensurative field experiment (Hadley and Betts 2016), we sampled populations of five lady beetle species at 30 sites along a gradient from low to high agricultural intensification in southern Wisconsin, USA. The sampling period extended from beetle emergence from hibernation in May to September when they allocate energy for hibernation, thus including periods when the region's large-scale row crops (corn and soybeans) are not yet present or are already harvested, and thus do not provide food resources. We assumed that lady beetles in agriculturally intensified landscapes—characterized by a high percentage of row crops, a low diversity of habitat types, and a low density of habitat edges—have less access to food resources than conspecifics in more complex landscapes, especially in the early and late seasons. We therefore predicted that lady beetles sampled in landscapes with high agricultural intensification will have a reduced body condition compared to beetles collected in less intensified landscapes. Because different lady beetle species have different habitat preferences and food requirements (Lundgren and Wiedenmann 2004; Berkvens et al. 2008; Gardiner et al. 2009a; Diepenbrock and Finke 2013), we also expected species-specific differences in the strength of response and relative importance of different landscape factors.
Methods
Sampling locations & collection
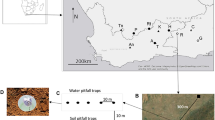
Adult lady beetles were sampled in 30 locations in Wisconsin, USA, between April and September 2011 (Fig. 1). The beetles were collected opportunistically based on the phenology of local vegetation and the abundance of food resources. In the early growing season when lady beetles emerged from hibernation, beetles were collected by sweep netting and hand collection in alfalfa fields, grasslands, and field margins. In June beetles were collected in small grain crops (wheat, oats, rye), and from late June onwards in corn and soybean fields. In addition, yellow sticky traps (Great Lakes IPM GL-3310–00, Vestaburg, MI, USA) were placed in semi-natural habitats such as hedgerows for 48 h in June. The sampling method had no effect on body condition metrics (data not shown) and therefore was not addressed in the statistical analysis. In total we collected five aphidophagous lady beetle species in high enough numbers to be included in the analysis. An overview on the species, number of individuals collected, and sites covered is given in Table 1. Collected beetles were placed in a cooler to limit their activity during transport to the lab, where they were preserved at −20 °C.

Location of the 30 sites (black points) sampled across the state of Wisconsin from April to September 2011. Inset maps show examples of 1 km buffers around the sampling locations with the corresponding land cover classification based on USDA data
Analysis of body condition
Beetles were sexed and dissected under a stereoscopic microscope (10–40 × magnification). The presence of mite or fungus parasites was documented in 153 individuals (11.9% of samples). Excluding these specimens did not qualitatively change the results (data not shown), so we present results based on the analysis of all samples. Additionally, 92 beetles (7.7% of the samples) had soft and pale elytra and were recorded as teneral (newly hatched). The presence or absence of eggs was also noted for female beetles.
We assessed the body condition of beetles by analyzing body size, body density, and lipid content. As an indicator of structural body size, we measured elytra length and pronotum width to the nearest 0.001 mm (ProgRes camera and CapturePro imaging system, JenOptik, Jena, Germany) and multiplied them. The use of multiple size measurements has been shown to be more informative of body condition than single size measurements (Knapp and Knappová 2013). In preparation for the measurement of lipid content, the elytra, wings, head and pronotum were removed from the torso (thorax and abdomen) and processed separately. All samples were dried at 50 °C for 72 h and cooled to room temperature in a desiccator for at least 1 h before being weighed to the nearest 0.001 mg (Mettler Toledo MT5, Mettler-Toledo, Columbus, OH, USA) (Anderson 1981). The torso was weighed separately from the other body parts, and the values were added to obtain the total dry mass. Body mass was corrected for individual size to standardize body condition, as the two measures are closely correlated (Knapp and Knappová 2013). We divided mass by size to determine "body density" (i.e., size-corrected dry mass).
Lipid content was determined gravimetrically by the difference in dry mass before and after total lipid extraction (Plaistow and Siva-Jothy 1996; Östman et al. 2001; Östman 2005). Only the torso was used (pronotum and abdomen without wings, elytra and legs) as wings and elytra contain no measurable amount of lipids, and no metabolically usable lipids are stored in the head (Plaistow and Siva-Jothy 1996). As a lipid solvent, 1 ml of dichloromethane-methanol solution (2:1) was added to the torso in individual glass vials. After 72 h at room temperature, the lipid-containing solution was discarded, and the torso was washed in another 1 ml of dichloromethane-methanol solution for 10 min and dried for 72 h at 50 °C before re-weighing. Percentage lipid content was calculated by dividing the lipid-free dry weight by the total dry weight of the torso and multiplying by 100 (Morita et al. 1999). Lipid content data was collected for all species except Coccinella septempunctata. Although there were significant correlations between body condition metrics (especially size and density), sufficient variation existed to test the effects of landscape on each metric separately (see Online Supplement 1 Fig. S1).
Landscape metrics
All landscape analyses were conducted at a 1 km radius around sampling sites. Landscape features within a 0.5 km to 2 km radius have been ecologically most informative for population-level numerical responses of lady beetles in previous studies (Gardiner et al. 2009a; Woltz and Landis 2014; Yang et al. 2018). While lady beetles migrate between habitats and patches during the season to find ephemeral prey, foraging within a patch is characterized by short-distance movements (Hodek et al. 1993; Evans 2003; Schellhorn et al. 2014). We therefore assume that a rather local scale is most relevant for physiological effects at the individual level.
We used the 2011 USDA Cropland Data Layer (CDL, NASS 2011) and the landscape metrics package in R (Hesselbarth et al. 2019) to obtain measures of landscape composition and configuration at a resolution of 30 m. We calculated the percentage of land categorized as corn and soybean, the dominant crops in the region, as a metric of intensively managed annual monoculture (hereafter “intensive cropland”). Intensive cropland was negatively correlated (Pearson’s r = −0.91, p < 0.001) with semi-natural habitat (grassland and woodland classes) and was used in our analysis because this type of habitat is easier to categorize unambiguously than semi-natural habitats.
Landscape diversity was quantified as the diversity of land cover classes (consolidated from USDA data to corn, soybean, alfalfa, small grains, other crops, grassland, woodland, developed, and water) using Simpson’s diversity index (Simpson 1949), which is less sensitive to rare classes than other metrics such as Shannon’s diversity index (McGarigal 1995). We included all habitat types, i.e., cultivated, semi-natural, and urban habitats, because due to their broad foraging range and high dispersal ability, lady beetles can exploit a variety of habitat types during their active season, including resources in residential gardens, and parks (Liere et al. 2019). Edge density was calculated as the length of the habitat boundaries of all land cover classes. High edge density is not only a measure of field size but is in the study region commonly associated with an increase in resources, as road verges and field boundaries often had relatively diverse year-round vegetation. Pearson correlation coefficients between pairs of these three landscape metrics used in our analysis were r < 0.4 (see Online Supplement 1 Fig. S2 for Pearson’s correlation coefficient distributions of landscape metrics).
Statistical analyses
We used R (v4.0.5; R Core Team 2020), and RStudio (v1.4.1103; RStudio Team 2020) for all statistical analysis and the ggplot2 package (Wickham 2016) for the visualization of results. Data and code are provided (Online Supplement 2 to 7).
We tested the effects of landscape metrics on the body condition of lady beetles with linear mixed-effects models (lme function, nlme package; Pinheiro et al. 2021). For each combination of body condition measurements (body size, body density, and lipid content) and landscape metrics (percentage of intensive cropland, landscape diversity, and edge density) we used separate models, resulting in nine predictor and response variable combinations.
For each response variable, we tested whether the data fitted a normal or lognormal distribution using the fitdistrplus package (function fitdist; Delignette-Muller and Dutang 2015) in combination with Akaikés information criterion (AIC). In addition, we visually checked the error distribution of the models to verify whether a ln-transformation reduced heteroskedasticity compared to untransformed response variables. Body size data was analyzed untransformed, while body density and lipid content data were ln-transformed. For models with body density as a response variable, teneral beetles (newly emerged adults) were excluded since they were significantly lighter than non-teneral beetles (data not shown). In models of body size or lipid content teneral beetles were statistically indistinguishable and were thus included (data not shown).
Because the scales of different landscape and body condition metrics varied considerably, we standardized the regression coefficients by transforming the predictor (% intensive cropland, Simpson’s diversity index, and edge density) and response variables (body size, body density, and lipid content) to z-scores. In all models the fixed effects were beetle species, collection date and their interactions with the landscape metrics. In the body density and lipid content models, a quadratic term was used for the collection date (allowing for a peak in mid-season), as this improved model fitting (lower AICc values; http://christoph-scherber.de/stepAICc.txt). Sampling site, species, and sex were included as nested random intercepts to account for repeated measures and species and sex specific differences in body traits. Females in our study were larger and tended to have a higher body density and lipid content than males. Mean and standard deviation of species-specific body measurements are shown in Table 1. A variance function with different estimates for each species was also used to account for variance heterogeneity between species.
We applied weighted effects coding using the wec package (Nieuwenhuis et al. 2017) to test the effects of landscape variables and their interactions with lady beetle species on body condition metrics. Weighted effects coding is a way to build contrasts for linear models to explicitly specify a regression intercept which represents the overall mean response across groups. It is an alternative to conventional “dummy coding” of categorical variables in regression analysis and is well-suited to analyzing unbalanced observational data. In weighted effects coding, the effect of each category (here, beetle species) represents the deviation of that category from the sample mean (te Grotenhuis et al. 2017). This allowed us to test whether there is an overall effect of each landscape variable on the body condition of all beetles sampled, as well as whether there are species for which the relationship between body condition and landscape metric is statistically different from the overall trend. The significance of all predictor terms in the linear mixed-effects models was assessed at α = 0.05 using type II Wald chi-square tests (Anova function, car package; Fox and Weisberg 2018). These tests follow the principle of marginality, where each term is tested after all others, but ignores each term’s interactions. A summary of the main effects of landscape predictors on all body condition metrics can be found in Table 2, and a graphical representation of selected effects in Fig. 2.

Effect of landscape predictors within 1 km of sampling locations on the body condition metrics of lady beetles, presented as partial residual plots (back-transformed from ln-transformation for illustrative purposes). The panels show: (a) the relationship between the amount of intensive cropland and body density; (b) the relationship between habitat edge and body density; (c) the relationship between the amount of intensive cropland and lipid content; (d) the relationship between landscape diversity and body density. Species-specific effects (individual lines) are shown for illustrative purposes only; the interaction between landscape metrics and lady beetle species was not significant in any model (see also Table 2 and Online Supplement 1 Table S1)
To investigate the effects of landscape on individual species, we extracted regression coefficients calculated with the generic summary function in R that is based on t-statistic using type-III sums of squares. The significance of predictor terms did not differ from the ones determined by Wald chi-square tests. The values from this test indicate whether the responses of individual species differed from the weighted means for all species (Table S1, Fig. 3). In addition, we were interested in testing whether individual species were affected by landscape even when all lady beetles as a group were not (i.e., whether the effect for each species was different than zero). To do this, we calculated means and 95% confidence intervals for each species by adding the marginal species effects to the all-species mean, then calculated p values from these confidence intervals (Table S2, Fig. 3).

Effect of landscape predictors (% intensive cropland, landscape diversity, and edge density) on lady beetle body condition (body size, body density, and lipid content). The x-axis shows standardized (z-scored) beta coefficients of the specific landscape predictors in linear mixed effects models. Black diamonds represent the overall effect of the landscape predictor on the body condition metric. Colored dots represent species-specific effects, whose estimates were calculated from interaction terms between landscape predictor and species. Error bars represent 95% confidence intervals. Filled symbols and lines highlight confidence intervals that do not cross the zero line, representing significant landscape effects. See Table S2 in Online Supplement 1 for corresponding summary statistic values
For four beetle species with sufficient numbers of non-teneral female individuals for statistical analysis (C. maculata, n = 83; C. munda, n = 32; C. septempunctata, n = 51; H. axyridis, n = 247) we examined the relationship between body condition metrics and the presence of eggs. To do this, we used logistic regression models using the glmmTMB package (Brooks et al. 2017). Models included egg presence/absence (1/0) as a response variable and one of the body condition metrics (body size, body density, and lipid content), collection date and their interactions with species as fixed effects, and sampling location as a random intercept. We used the weighted effects coding described above to determine the overall effect of body density on egg presence and possible interactions with the species. The significance of predictor terms was assessed using Wald chi-square tests (Anova function, car package; Fox and Weisberg 2018) as described above (see Online Supplement 1: Fig. S3, Table S3 and S4).
Results
All three landscape predictors studied had a significant effect on the body condition of at least one lady beetle species (Table 2; Figs. 2, 3). As the percentage of intensively managed cropland in a landscape increased there was a small, but significant decrease in body density and lipid content of lady beetles. For every 10% increase in landscape area covered by intensively managed cropland, there was an approximately 3.9% decrease in beetle body density and 6.4% decrease in beetle lipid content (Table 2; Fig. 2a, c). At the species level, an increasing amount of intensively managed cropland reduced body density of two introduced species, the multicolored Asian lady beetle Harmonia axyridis (Fig. 3, orange points) and the variegated lady beetle Hippodamia variegata (Fig. 3, blue points) and lipid content of H. axyridis. Intensive cropland was also negatively associated with body size of the native polished lady beetle Cycloneda munda (Fig. 3, yellow points).
Landscape diversity and edge density did not have an overall effect for any of the body condition metrics (Table 2) but had effects on individual species. The native pink spotted lady beetle Coleomegilla maculata (Fig. 3; green points) had reduced lipid content in more diverse landscapes, and C. maculata and H. variegata collected from landscapes with higher edge density had greater body density.
Responses of individual species did not generally differ from that of all lady beetles together; the sole exception was that the body density of C. maculata responded more negatively to landscape diversity than the all-species mean effect (Table S1), but neither the effect of landscape diversity on C. maculata nor the overall effect were statistically significant (Table S2). See Online Supplement 1 Table S1 for summary statistics on deviations of species-specific responses from the all-species mean effect, and Table S2 for effects of landscape predictors on individual species.
Lady beetle species was a strong and significant predictor of body condition in all models (Table 2). That is, there were inherent differences among species after accounting for landscape metrics. The collection date itself had an effect on body density, with higher body density in the middle of the season. The interaction between collection date and beetle species was a significant predictor of body density and lipid content (Table 2), indicating that these body condition metrics increased in some species and decreased in others over the season.
In all four species analyzed, female beetles with greater body density were more likely to have developing eggs (Online Supplement 1 Fig S3, Table S3, S4). This suggests that body density (i.e., standardized body mass) is a meaningful index of fecundity. No relationship was found between the presence of eggs and body size or lipid content (Online Supplement 1 Table S3, S4).
Discussion
Landscape composition and configuration explained small but significant portions of the variation in the body condition of multiple lady beetle species across southern Wisconsin’s agricultural landscapes. Lady beetles were generally lighter and leaner in simplified agricultural landscapes dominated by annual row crop fields. Previous studies have shown that coccinellid abundance and biological control services decline in simplified landscapes in the midwestern United States and other regions of the world (Gardiner et al. 2009b; Grez et al. 2014; Woltz and Landis 2014; Yang et al. 2018, 2019). Our results provide a possible mechanistic link for the observed relationship between landscape structure and lady beetle numeric response, and consequently ecosystem services.
Likely reasons for the effects of landscape composition and configuration on lady beetle body condition include differences in spatial and temporal resource continuity and quality. Corn and soybean fields can be heavily colonized by aphids and thus provide abundant prey for lady beetles, but this is only the case during limited time periods; over long periods such fields are characterized by resource scarcity, e.g., before crop emergence, after crop maturity, or after pesticide application (Rand et al. 2006; Schellhorn et al. 2014; Iuliano and Gratton 2020). In landscapes that are strongly dominated by a few crop monocultures, these periods are synchronized over large areas and habitat patches with alternative food sources are often far away and only accessible at high energetic costs (Bonte et al. 2012; Stowe et al. 2021b). In contrast, in more heterogeneous landscapes where a variety of annual and perennial crops are grown and natural or semi-natural habitat patches are available, landscape complementation (in the sense of Dunning et al. 1992) for mobile natural enemies may occur. Landscapes with more intensive cropland may also mean greater risk of pesticide exposure (Meehan et al. 2011), which has potential lethal and sublethal consequences for lady beetles and other beneficial insects (Santos et al. 2017; Stuligross and Williams 2021), though little is known about pesticide effects at the landscape scale (Mancini et al. 2020).
Landscape structure also shapes the diversity and composition of plants and arthropods (Jonsen and Fahrig 1997; Clough et al. 2007; Seibold et al. 2019; Le Provost et al. 2021) and consequently the quality of food resources for lady beetles, which in turn may influence their performance. Nutritional studies conducted in the laboratory have shown that growth rate, body composition, and fecundity can vary significantly depending on the prey and non-prey foods consumed. For example, C. maculata can complete its life cycle entirely on corn pollen, but then achieved a lower body weight and laid fewer eggs than conspecifics that feed on aphids (Lundgren and Wiedenmann 2004). In H. axyridis, known primarily as aphid predators, a diet of Lepidoptera eggs (Ephestia kuehniella Zeller) resulted in higher body weight, lipid content, reproductive output, and longevity compared to a diet of pea aphids (Acyrthosiphon pisum Harris) (Specty et al. 2003; Berkvens et al. 2008). In this context, it appears to be species-specific which food types are best suited for predators to achieve high body condition. However, a mixed diet of complementary foods has been shown to be beneficial for several generalist predators and is actively sought by them (Soares et al. 2004; Raubenheimer et al. 2007; Harwood et al. 2009; Marques et al. 2015; Stowe et al. 2021a). As such, landscapes with fewer diversity of crops or landscape diversity may manifest as decreases in the physiological condition of mobile predatory arthropods such as lady beetles, leading to longer-term population declines, as has been found for other beneficial insects (Hemberger et al. 2021).
Generalist natural enemies are likely to achieve a balanced diet more easily in landscapes with high resource diversity. Locally high plant diversity and habitat heterogeneity increased the dietary diversity and composition of generalist predatory ground arthropods (Tiede et al. 2016; Staudacher et al. 2018; Macé et al. 2019). In a study from Germany and Switzerland, H. axyridis and another common aphid predator, the lacewing Chrysoperla carnea (Stephens), supplemented their diets throughout the season with pollen that, even in agriculture-dominated landscapes, came largely from non-crop plants (Bertrand et al. 2019). Local semi-natural habitats can supplement the food supply for natural enemies of plant pests, but the availability of resources also depends on the surrounding landscape. Seibold et al. (2019) demonstrated a decline in arthropods over nine years in grasslands that was particularly pronounced when the areas were embedded in intensively farmed landscapes.
In our study, we examined the correlation of three different aspects of landscape simplification with three different characteristics of coccinellid body condition. The two measures of landscape composition, the percentage of predominant annual monocultures of soybean and corn, and landscape diversity were not correlated. This means that a landscape with few soybean and corn crops is not necessarily a landscape with many different other habitat types, and vice versa. The third metric, edge density, is a measure of landscape configuration and provides information about the size of fields and other habitats. In addition, high edge density is often associated with an increase in resources, as road verges and field boundaries are often covered with natural vegetation year-round. Its independence from the other two landscape metrics implies that, for example, landscapes with a high percentage of intensively used farmland may consist of many small fields with correspondingly high edge density, and high edge density is not necessarily associated with high diversity of habitat types (see also Martin et al. 2019).
Of these three factors, the percentage of intensive cropland was the strongest predictor of lady beetle body condition and the only one for which a general effect on all species was found. A high percentage of intensive cropland was generally negatively correlated with lipid content and body density of coccinellids. In a former study, lady beetles collected from semi-natural habitats had qualitatively higher lipid content than conspecifics from monocultures, but a direct effect of the amount of intensive cropland in the surrounding landscape could not be demonstrated (Tiede et al. 2017). The survey period of our present study exceeds that of the former one by far, and thus covers a larger part of the activity period of coccinellids, ranging from awakening from hibernation to reproduction of (multiple) new generations to preparation for overwintering. This includes periods when annual row crops may be productive food sources for natural enemies, as well as periods when crop pests are not yet or no longer present. While high pest abundance can have a positive short-term effect on the development of natural enemy populations (Rand et al. 2006), our results suggest longer-term negative consequences of agricultural intensification for beneficial insect populations.
While the percentage of soybean and corn fields can be easily and unambiguously categorized, landscape diversity as a measure of heterogeneity is less straightforward, as the composition of habitat types can vary greatly even with the same level of diversity. Some habitat types, although contributing to greater diversity at the landscape level, may not provide many resources for coccinellids, e.g., impervious surfaces, water bodies, or intensively pesticide-treated crops. As landscape diversity in our study includes both, crops and semi-natural habitats, low landscape diversity could be attributed to large semi-natural habitats. Preferences for habitat types are often species-specific. For example, native species are often found in grasslands, whereas forests play a more important role for introduced species (Gardiner et al. 2009a; Werling et al. 2011; Diepenbrock and Finke 2013). We detected a significant negative effect of landscape diversity on lipid content but only for the native C. maculata. Other studies have already shown that this species differs in its response to landscape diversity from the response of the lady beetle community: C. maculata achieved higher abundances in landscapes with low habitat diversity and a high proportion of grassland and corn, while the total abundance of lady beetles and their biological control efficiency were greatest when landscape diversity was high (Gardiner et al. 2009a, 2010).
High edge density had a positive effect on the body density of C. maculata and H. variegata, but again no overall effect could be detected. C. maculata and H. variegata are the smallest species in our study and a small field size could possibly help them overcome size-related dispersal limitations due to the presence of nearby alternative resources (Gámez-Virués et al. 2015; Boetzl et al. 2019). In addition, C. maculata and H. variegata might also benefit most from pollen from flowering plants on field margins and along roadsides (Lundgren and Wiedenmann 2004; Bertolaccini et al. 2008).
Landscape classification using the USDA Cropland Data Layer (CDL, NASS 2011) provides a relatively coarse 30 m × 30 m resolution of land cover classes. A finer-grained landscape analysis that also captures small edge structures and habitat islands, as well as field margins between the same habitat types could potentially reveal stronger impacts of edge density on coccinellid performance. Small-scale structures that can be accessed with minimal energy expenditure such as field margins can be an attractive source of food and shelter for coccinellids (Hodek et al. 2012). Recent studies have found that edge density increases abundance of many pollinators and natural enemy species, improves pollination and pest control, and increases yields in arable landscapes (Martin et al. 2019). Alternatively, other measures of landscape configuration such as connectivity may reveal additional landscape features that translate to physiological outcomes for lady beetles (Koh et al. 2013).
We used three different physiological parameters to estimate body condition in lady beetles to account for the fact that an animal's phenotype is a consequence of many integrated and interdependent traits (Wilder et al. 2016). Body density and lipid content were more informative for the influence of landscape structure on body condition than body size. This is of little surprise given that body size can be influenced by two different mechanisms with opposite effects: Dispersal limitation and resource limitation. Smaller individuals may benefit from landscape complexity because it provides many patches of resources within short distances, so landscape simplification may increase body size by filtering out the small individuals (Gámez-Virués et al. 2015; Boetzl et al. 2019). On the other hand, complex landscapes offer more resources, so landscape simplication may decrease body size by limiting resources. In our study, we only found evidence of the latter mechanism; body size of C. munda was smaller in landscapes with high proportions of agriculture. Additionally, more pronounced effects may be constrained by the fact that body size is determined during larval development (Vargas et al. 2013b) but adult lady beetles often travel repeatedly longer distances in search of suitable foraging, oviposition, or overwintering sites (Hodek et al. 1993; Schellhorn et al. 2014). Thus, the landscape within a 1 km radius of sampling sites may not be representative of larval foraging habitat.
Body density and lipid content are better predictors of recent access to resources. Lipid content is a direct measure of body composition and a good estimate of energy reserves in arthropods (Lease and Wolf 2011; Knapp and Knappová 2013). Body density, calculated as size-corrected body weight, also comprises total lipid weight, but in addition includes the content of other body substances, e.g., protein. Protein is not directly related to the energy reserves of individuals but has been shown to be of considerable importance for insect reproductive fitness (Knapp and Knappová 2013; Wilder et al. 2016). Consistently, body density was also the only body parameter we measured that was correlated with the presence of eggs. We conclude that, taken together, the chosen parameters provide good insight into the effects of landscape simplification on the body condition and fitness of coccinellids.
Continuous sampling of local lady beetle populations is challenging because the highly mobile beetles move between different habitat patches in search of food or egg-laying sites. We often found that a collection site that previously had high lady beetle abundance was devoid of lady beetles on the next visit. In the resulting dataset, landscape structure and collection date are somewhat confounded. As such, for species that could only be collected at a limited subset of sites or dates, the full extent of landscape effects on body metrics may not have been fully measured. However, lipid content and body density could reflect nutritional conditions over a longer time span. Thus, better access to resources in the early season would give beetles a "head start" that could lead to overall better body condition in the peak season, explaining the negative correlation with intensive agriculture after accounting for time.
Samples were dominated by the native C. maculata and the introduced H. axyridis, while other species comprised < 20% of the specimens. This, however, is consistent with species distributions in agricultural landscapes of the upper Midwestern United States (Werling et al. 2011; Lamb et al. 2019). The dramatic decline of native coccinellid species in the United States, (Harmon et al. 2007; Gardiner et al. 2011; Lamb et al. 2019), Europe (Brown and Roy 2018) and South America (Grez et al. 2013, 2014) since the late twentieth century has often been linked to the introduction of invasive competitors such as H. axyridis or H. variegata, but an important mediating effect of landscape structure has also been emphasized (Gardiner et al. 2009a, 2011; Werling et al. 2011; Grez et al. 2013; Bahlai et al. 2015). Since reliable conclusions about the relationship between landscape characteristics and body condition require a robust sample size, rare species had to be excluded from the analysis. Thus, it can only be speculated whether a reduction in body condition due to landscape intensification may have contributed to their low abundance. An enormous collection effort would be required to cover rare species as well. Our results, however, demonstrate that even highly mobile species closely associated with cropland and pest control, such as H. axyridis, exhibit the effects of agricultural intensification on physiological performance.
While the landscape effects on lady beetle body condition documented here are modest, they suggest the potential loss of biological pest control due to a yet-to-be fully realized numerical response, since lighter and leaner beetles may be less effective predators, produce fewer offspring and have a lower chance of survival (Arrese and Soulages 2009; Kajita and Evans 2010; Beukeboom 2018). These results are consistent with arguments for maintaining semi-natural habitats, diversifying cropping systems, and increasing the number of edges (e.g., by reducing the size of fields) in agricultural landscapes where conservation of beneficial arthropods such as lady beetles and their ecosystem services are desirable.
Data availability
Data is available in Online Supplement 2 to 5.
Code availability
Code is available in Online Supplement 6 to 7.
References
Alaux C, Allier F, Decourtye A et al (2017) A ‘Landscape physiology’ approach for assessing bee health highlights the benefits of floral landscape enrichment and semi-natural habitats. Sci Rep. https://doi.org/10.1038/srep40568
Anderson JME (1981) Seasonal field analyses of fat content, live weight, dry weight and water content of the aphidophagous Scymnodes Lividigaster (Mulsant) and mycophagous Leptothea Galbula (Mulsant) (Coleoptera: Coccinellidae). Aust J Zool 29:679–689
Arrese EL, Soulages JL (2009) Insect fat body: energy, metabolism, and regulation. Annu Rev Entomol 55:207–225
Bahlai CA, Colunga-Garcia M, Gage SH, Landis DA (2015) The role of exotic ladybeetles in the decline of native ladybeetle populations: evidence from long-term monitoring. Biol Invasions 17:1005–1024
Berkvens N, Bonte J, Berkvens D et al (2008) Influence of diet and photoperiod on development and reproduction of European populations of Harmonia axyridis (Pallas) (Coleoptera: Coccinellidae). Biol Control Invasion Ladybird Harmon Axyridis Model Species 53:211–221
Bertolaccini I, Núñez-Pérez E, Jorge Tizado E (2008) Effect of wild flowers on oviposition of Hippodamia variegata (Coleoptera: Coccinellidae) in the laboratory. J Econ Entomol 101:1792–1797
Bertrand C, Eckerter PW, Ammann L et al (2019) Seasonal shifts and complementary use of pollen sources by two bees, a lacewing and a ladybeetle species in European agricultural landscapes. J Appl Ecol 56:2431–2442
Beukeboom LW (2018) Size matters in insects—an introduction. Entomol Exp Appl 166:2–3
Boetzl FA, Krimmer E, Krauss J, Steffan-Dewenter I (2019) Agri-environmental schemes promote ground-dwelling predators in adjacent oilseed rape fields: diversity, species traits and distance-decay functions. J Appl Ecol 56:10–20
Bommarco R (1998) Reproduction and energy reserves of a predatory carabid beetle relative to agroecosystem complexity. Ecol Appl 8:846–853
Bonte D, Van Dyck H, Bullock JM et al (2012) Costs of dispersal. Biol Rev 87:290–312
Brooks ME, Kristensen K, van Benthem KJ et al (2017) glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J 9:378–400
Brown PMJ, Roy HE (2018) Native ladybird decline caused by the invasive harlequin ladybird Harmonia axyridis: evidence from a long-term field study. Insect Conserv Divers 11:230–239
Chaplin-Kramer R, O’Rourke ME, Blitzer EJ, Kremen C (2011) A meta-analysis of crop pest and natural enemy response to landscape complexity. Ecol Lett 14:922–932
Clough Y, Kruess A, Tscharntke T (2007) Local and landscape factors in differently managed arable fields affect the insect herbivore community of a non-crop plant species. J Appl Ecol 44:22–28
Cooke SJ, Sack L, Franklin CE et al (2013) What is conservation physiology? Perspectives on an increasingly integrated and essential science. Conserv Physiol. https://doi.org/10.1093/conphys/cot001
Dainese M, Martin EA, Aizen MA et al (2019) A global synthesis reveals biodiversity-mediated benefits for crop production. Sci Adv. https://doi.org/10.1126/sciadv.aax0121
Delignette-Muller ML, Dutang C (2015) Fitdistrplus: an R package for fitting distributions. J Stat Softw 64:1–34
Diepenbrock LM, Finke DL (2013) Refuge for native lady beetles (Coccinellidae) in perennial grassland habitats. Insect Conserv Divers 6:671–679
Dolezal AG, Clair ALS, Zhang G et al (2019) Native habitat mitigates feast–famine conditions faced by honey bees in an agricultural landscape. Proc Natl Acad Sci 116:25147–25155
Drapela T, Frank T, Heer X et al (2013) Landscape structure affects activity density, body size and fecundity of Pardosa wolf spiders (Araneae: Lycosidae) in winter oilseed rape. EJE 108:609–614
Dunning JB, Danielson BJ, Pulliam HR (1992) Ecological processes that affect populations in complex landscapes. Oikos 65:169–175
Ellis RD, McWhorter TJ, Maron M (2012) Integrating landscape ecology and conservation physiology. Landsc Ecol 27:1–12
Evans EW (2003) Searching and reproductive behaviour of female aphidophagous ladybirds (Coleoptera: Coccinellidae): a review. EJE 100:1–10
Fahrig L (2007) Non-optimal animal movement in human-altered landscapes. Funct Ecol 21:1003–1015
Fox J, Weisberg S (2018) An R companion to applied regression. SAGE Publications
Gámez-Virués S, Perović DJ, Gossner MM et al (2015) Landscape simplification filters species traits and drives biotic homogenization. Nat Commun. https://doi.org/10.1038/ncomms9568
Gardiner MM, Landis DA, Gratton C et al (2009a) Landscape diversity enhances biological control of an introduced crop pest in the north-central USA. Ecol Appl 19:143–154
Gardiner MM, Landis DA, Gratton C et al (2009b) Landscape composition influences patterns of native and exotic lady beetle abundance. Divers Distrib 15:554–564
Gardiner MM, O’Neal ME, Landis DA (2011) Intraguild predation and native lady beetle decline. PLoS ONE 6:e23576
Gardiner MA, Tuell JK, Isaacs R et al (2010) Implications of three biofuel crops for beneficial arthropods in agricultural landscapes. BioEnergy Res 3:6–19
Grab H, Brokaw J, Anderson E et al (2019) Habitat enhancements rescue bee body size from the negative effects of landscape simplification. J Appl Ecol 56:2144–2154
Grez AA, Rand TA, Zaviezo T, Castillo-Serey F (2013) Land use intensification differentially benefits alien over native predators in agricultural landscape mosaics. Divers Distrib 19:749–759
Grez AA, Zaviezo T, Hernández J et al (2014) The heterogeneity and composition of agricultural landscapes influence native and exotic coccinellids in alfalfa fields. Agric for Entomol 16:382–390
Guerra C, Aráoz E (2016) Amphibian malformations and body condition across an agricultural landscape of northwest Argentina. Dis Aquat Organ 121:105–116
Hadley AS, Betts MG (2016) Refocusing habitat fragmentation research using lessons from the last decade. Current Landscape Ecology Reports 1(2):55–66
Harmon JP, Stephens E, Losey J (2007) The decline of native coccinellids (Coleoptera: Coccinellidae) in the United States and Canada. J Insect Conserv 11:85–94
Harwood J, Phillips S, Anderson J et al (2009) Invertebrate biodiversity affects predator fitness and hence potential to control pests in crops. Biol Control 51:499–506
Hemberger J, Crossley MS, Gratton C (2021) Historical decrease in agricultural landscape diversity is associated with shifts in bumble bee species occurrence. Ecol Lett 24:1800–1813
Hesselbarth MHK, Sciaini M, With KA et al (2019) landscapemetrics: an open-source R tool to calculate landscape metrics. Ecography 42:1648–1657
Hodek I (1973) Biology of coccinellidae. Dr. W. Junk N.V, The Hague
Hodek I, Iperti G, Hodkova M (1993) Long-distance flights in Coccinellidae (Coleoptera). EJE 90:403–414
Hodek I, Honek A, van Emden HF (2012) Ecology and behaviour of the ladybird beetles (Coccinellidae). John Wiley & Sons
Iuliano B, Gratton C (2020) Temporal resource (dis)continuity for conservation biological control: from field to landscape scales. Front Sustain Food Syst. https://doi.org/10.3389/fsufs.2020.00127
Jonsen ID, Fahrig L (1997) Response of generalist and specialist insect herbivores to landscape spatial structure. Landsc Ecol 12:185–197
Kajita Y, Evans EW (2010) Relationships of body size, fecundity, and invasion success among predatory lady beetles (Coleoptera: Coccinellidae) inhabiting alfalfa fields. Ann Entomol Soc Am 103:750–756
Knapp M, Knappová J (2013) Measurement of body condition in a common carabid beetle, Poecilus cupreus: a comparison of fresh weight, dry weight, and fat content. J Insect Sci. https://doi.org/10.1673/031.013.0601
Koh I, Rowe HI, Holland JD (2013) Graph and circuit theory connectivity models of conservation biological control agents. Ecol Appl 23(7):1554–1573
Kuussaari M, Bommarco R, Heikkinen RK et al (2009) Extinction debt: a challenge for biodiversity conservation. Trends Ecol Evol 24:564–571
Lamb RJ, Bannerman JA, Costamagna AC (2019) Stability of native and exotic lady beetle populations in a diverse landscape. Ecosphere 10:e02630
Latimer CE, Smith OM, Taylor JM et al (2020) Landscape context mediates the physiological stress response of birds to farmland diversification. J Appl Ecol 57:671–680
Le Provost G, Thiele J, Westphal C et al (2021) Contrasting responses of above- and belowground diversity to multiple components of land-use intensity. Nat Commun 12:3918
Lease HM, Wolf BO (2011) Lipid content of terrestrial arthropods in relation to body size, phylogeny, ontogeny and sex. Physiol Entomol 36:29–38
Liere H, Egerer MH, Philpott SM (2019) Environmental and spatial filtering of ladybird beetle community composition and functional traits in urban landscapes. J Urban Ecol. https://doi.org/10.1093/jue/juz014
Liere H, Kim TN, Werling BP et al (2015) Trophic cascades in agricultural landscapes: indirect effects of landscape composition on crop yield. Ecol Appl 25:652–661
Lundgren JG, Wiedenmann RN (2004) Nutritional suitability of corn pollen for the predator Coleomegilla maculata (Coleoptera: Coccinellidae). J Insect Physiol 50:567–575
Macé OG, Ebeling A, Eisenhauer N et al (2019) Variations in trophic niches of generalist predators with plant community composition as indicated by stable isotopes and fatty acids. Soil Org 91:45–59
Mancini F, Woodcock BA, Redhead J et al (2020) Chapter three—detecting landscape scale consequences of insecticide use on invertebrate communities. In: Bohan DA, Vanbergen AJ (eds) Advances in ecological research. Academic Press, pp 93–126
Marques RV, Sarmento RA, Lemos F et al (2015) Active prey mixing as an explanation for polyphagy in predatory arthropods: synergistic dietary effects on egg production despite a behavioural cost. Funct Ecol 29:1317–1324
Martin EA, Dainese M, Clough Y et al (2019) The interplay of landscape composition and configuration: new pathways to manage functional biodiversity and agroecosystem services across Europe. Ecol Lett 22:1083–1094
McGarigal K (1995) FRAGSTATS: spatial pattern analysis program for quantifying landscape structure. U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station
Meehan TD, Werling BP, Landis DA, Gratton C (2011) Agricultural landscape simplification and insecticide use in the Midwestern United States. Proc Natl Acad Sci 108(28):11500–11505
Mogren CL, Benítez M-S, McCarter K et al (2020) Diverging landscape impacts on macronutrient status despite overlapping diets in managed (Apis mellifera) and native (Melissodes desponsa) bees. Conserv Physiol. https://doi.org/10.1093/conphys/coaa109
Morita A, Soga K, Hoson T et al (1999) Changes in mechanical properties of the cuticle and lipid accumulation in relation to adult diapause in the bean bug, Riptortus clavatus. J Insect Physiol 45:241–247
Nieuwenhuis R, te Grotenhuis M, Pelzer B (2017) Weighted effect coding for observational data with wec. R J 9:477–485
Öberg S (2009) Influence of landscape structure and farming practice on body condition and fecundity of wolf spiders. Basic Appl Ecol 10:614–621
Obrycki JJ, Harwood JD, Kring TJ, O’Neil RJ (2009) Aphidophagy by Coccinellidae: application of biological control in agroecosystems. Biol Control 51:244–254
Östman Ö (2005) Asynchronous temporal variation among sites in condition of two carabid species. Ecol Entomol 30:63–69
Östman Ö, Ekbom B, Bengtsson J, Weibull A-C (2001) Landscape complexity and farming practice influence the condition of polyphagous carabid beetles. Ecol Appl 11:480–488
Pinheiro J, Bates D, DebRoy S, Sarkar D (2021) nlme: linear and nonlinear mixed effects models. https://cran.r-project.org/web/packages/nlme/nlme.pdf. Accessed 4 Nov 2021
Plaistow S, Siva-Jothy MT (1996) Energetic constraints and male mate-securing tactics in the damselfly Calopteryx splendens xanthostoma (Charpentier). Proc R Soc Lond B Biol Sci 263:1233–1239
Rand TA, Tylianakis JM, Tscharntke T (2006) Spillover edge effects: the dispersal of agriculturally subsidized insect natural enemies into adjacent natural habitats. Ecol Lett 9:603–614
Raubenheimer D, Mayntz D, Simpson SJ, Tøft S (2007) Nutrient-specific compensation following diapause in a predator: implications for intraguild predation. Ecology 88:2598–2608
Renauld M, Hutchinson A, Loeb G et al (2016) Landscape simplification constrains adult size in a native ground-nesting bee. PLoS ONE 11:e0150946
Rusch A, Chaplin-Kramer R, Gardiner MM et al (2016) Agricultural landscape simplification reduces natural pest control: a quantitative synthesis. Agric Ecosyst Environ 221:198–204
Salomão RP, González-Tokman D, Dáttilo W et al (2018) Landscape structure and composition define the body condition of dung beetles (Coleoptera: Scarabaeinae) in a fragmented tropical rainforest. Ecol Indic 88:144–151
Santos KFA, Zanuzo Zanardi O, de Morais MR et al (2017) The impact of six insecticides commonly used in control of agricultural pests on the generalist predator Hippodamia convergens (Coleoptera: Coccinellidae). Chemosphere 186:218–226
Schellhorn NA, Bianchi FJJA, Hsu CL (2014) Movement of entomophagous arthropods in agricultural landscapes: links to pest suppression. Annu Rev Entomol 59:559–581
Seibold S, Gossner MM, Simons NK et al (2019) Arthropod decline in grasslands and forests is associated with landscape-level drivers. Nature 574:671–674
Simpson EH (1949) Measurement of diversity. Nature 163:688–688
Soares AO, Coderre D, Schanderl H (2004) Dietary self-selection behaviour by the adults of the aphidophagous ladybeetle Harmonia axyridis (Coleoptera: Coccinellidae). J Anim Ecol 73:478–486
Specty O, Febvay G, Grenier S et al (2003) Nutritional plasticity of the predatory ladybeetle Harmonia axyridis (Coleoptera: Coccinellidae): comparison between natural and substitution prey. Arch Insect Biochem Physiol 52:81–91
Staudacher K, Rennstam Rubbmark O, Birkhofer K et al (2018) Habitat heterogeneity induces rapid changes in the feeding behaviour of generalist arthropod predators. Funct Ecol 32:809–819
Stowe HE, Michaud JP, Kim T (2021a) The benefits of omnivory for reproduction and life history of a specialized aphid predator, Hippodamia convergens (Coleoptera: Coccinellidae). Environ Entomol 50:69–75
Stowe HE, Michaud JP, Kim TN (2021b) Floral resources enhance fecundity, but not flight activity, in a specialized aphid predator, Hippodamia convergens (Coleoptera: Coccinellidae). Front Ecol Evol 9:619
Stuligross C, Williams NM (2021) Past insecticide exposure reduces bee reproduction and population growth rate. PNAS 118:np
te Grotenhuis M, Pelzer B, Eisinga R et al (2017) When size matters: advantages of weighted effect coding in observational studies. Int J Public Health 62:163–167
Tiede J, Scherber C, Mutschler J et al (2017) Gut microbiomes of mobile predators vary with landscape context and species identity. Ecol Evol 7:8545–8557. https://doi.org/10.1002/ece3.3390
Tiede J, Wemheuer B, Traugott M et al (2016) Trophic and non-trophic interactions in a biodiversity experiment assessed by next-generation sequencing. PLoS ONE 11:e0148781. https://doi.org/10.1371/journal.pone.0148781
Tscharntke T, Tylianakis JM, Rand TA et al (2012) Landscape moderation of biodiversity patterns and processes—eight hypotheses. Biol Rev 87:661–685
Vargas G, Michaud JP, Nechols JR (2012) Larval food supply constrains female reproductive schedules in Hippodamia convergens (Coleoptera: Coccinellidae). Ann Entomol Soc Am 105:832–839
Vargas G, Michaud JP, Nechols JR (2013a) Cryptic maternal effects in Hippodamia convergens vary with maternal age and body size. Entomol Exp Appl 146:302–311
Vargas G, Michaud JP, Nechols JR (2013b) Trajectories of reproductive effort in Coleomegilla maculata and Hippodamia convergens (Coleoptera: Coccinellidae) respond to variation in both income and capital. Environ Entomol 42:341–353
Werling BP, Meehan TD, Gratton C, Landis DA (2011) Influence of habitat and landscape perenniality on insect natural enemies in three candidate biofuel crops. Biol Control 59:304–312
Wickham H (2016) ggplot2: elegant graphics for data analysis. https://ggplot2.tidyverse.org. Accessed 4 Nov 2021
Wilder SM, Raubenheimer D, Simpson SJ (2016) Moving beyond body condition indices as an estimate of fitness in ecological and evolutionary studies. Funct Ecol 30:108–115
Woltz JM, Landis DA (2014) Coccinellid response to landscape composition and configuration. Agric for Entomol 16:341–349
Yang L, Xu L, Liu B et al (2019) Non-crop habitats promote the abundance of predatory ladybeetles in maize fields in the agricultural landscape of northern China. Agric Ecosyst Environ 277:44–52
Yang L, Zeng Y, Xu L et al (2018) Change in ladybeetle abundance and biological control of wheat aphids over time in agricultural landscape. Agric Ecosyst Environ 255:102–110
Acknowledgements
We would like to thank the landowners who allowed us to sample lady beetles on their land, and we also thank Rachel Mallinger, Hannah Gaines-Day, Heidi Liere, Allison Dehnel, and Ken Frost for rides to the sampling sites. This research was supported by the United States Department of Agriculture (USDA) grants 2011-67009-30022 and 2018-67013-28060 to CG.
Funding
Open Access funding enabled and organized by Projekt DEAL. This research was supported by the United States Department of Agriculture (USDA) grants 2011–67009-30022 and 2018–67013-28060 to CG.
Author information
Authors and Affiliations
Contributions
JT and CG conceived of the study. JT conducted field sampling and laboratory analysis. BI and JT analyzed the data. BI wrote the first manuscript draft. All authors contributed to subsequent drafts.
Corresponding author
Ethics declarations
Conflicts of interest
The authors declare no conflict of interest.
Ethical approval
None required.
Consent to participate
None required.
Consent for publication
All authors agree to the content of this manuscript.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Tiede, J., Iuliano, B. & Gratton, C. Agriculturally intensified landscapes are associated with reduced body condition of lady beetles. Landsc Ecol 37, 1921–1936 (2022). https://doi.org/10.1007/s10980-022-01458-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10980-022-01458-0