
Abstract
Underwater light availability and exposure of ultraviolet radiation (UV) in mountain lakes is mainly controlled by dissolved organic matter and ice cover. However, both of these factors are affected by climate warming and other anthropogenic pressures. Still, little is known of the impacts of long-term fluctuations in underwater light conditions onto functional distribution of diatoms, species sensitive to changes in climate and UV penetration in mountain lakes. Two mountain lakes in the Italian Alps were analysed using paleolimnological approaches to investigate impacts of anthropogenic pressures (e.g., climate warming, acidification) on underwater light availability, exposure to UV and diatom functional group distributions, focusing on the post-industrial era. Contemporary diatom communities were collected from the main habitats in the shallow, high irradiance littoral zones of these lakes and geochemical proxies describing the development of lake-water transparency and carbon dynamics were analyzed from the sediment cores covering the time period from ca. 1400 CE to present. The geochemical data indicate that cultural acidification decreased lake-water organic carbon concentrations in both lakes, which suggests increased light availability and UV exposure during the past century. The responses of the studied lakes to anthropogenic pressures varied, as the lake situated at a lower altitude with a larger catchment showed only a few biotic changes indicating higher resilience, whereas the lake situated at higher altitude showed distinct changes in its ecological status. In this alpine lake, almost 30% of the diatom functional groups shifted from benthic to planktic during the post-industrial era. The role of increased light availability and UV exposure as a driver of diatom functional group distributions could not be unambiguously separated, and such diatom assemblage changes have been shown in many regions and most closely linked to a warming climate. However, low guild functional groups, prevalent also in the contemporary samples from the shallow littoral zone, dominated the diatom communities throughout the studied period, suggesting high tolerance to UV radiation.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
In mountain lakes, underwater light conditions, consisting of photosynthetically active radiation (PAR) and ultraviolet radiation (UV), are controlled mainly by lake-water dissolved organic matter (DOM; Sommaruga 2001; Williamson et al. 2009). Brown colored, fulvic and humic acids and other high molecular weight compounds, comprising primarily allochthonous DOM from terrestrial sources, effectively absorb light, whereas autochthonous DOM from aquatic primary production is typically colorless with weaker light-absorbing capacity (McKnight et al. 2001; McGowan et al. 2016; Wauthy et al. 2017). The accelerating increase of air temperature in high altitude areas (Pepin et al. 2015), such as the European Alps (Auer et al. 2007), drives changes in catchment soils and vegetation that shape DOM inflow and its production, transformation, and degradation in the water column (Laurion et al. 2000; Häder et al. 2014; Nevalainen et al. 2020).
In addition to a warming climate, other anthropogenic pressures, such as atmospheric deposition of sulphur (S) and nitrogen (N), can impact remotely located mountain lakes. Widespread lake acidification driven by anthropogenic emissions has changed the biochemistry of many lakes and their catchments, which further impacts their biological communities. Several studies have observed decreases in lake-water total organic carbon (TOC) concentrations, particularly around the mid-twentieth century, followed by increasing TOC trends after the mitigation of anthropogenic S emission in the 1980s (Monteith al. 2007; Meyer-Jacob et al. 2019). These changes in lake-water TOC likely increase and decrease light availability and UV exposure in mountain lakes, respectively, though only a few studies have investigated associated biotic responses (Bothwell et al. 1993; Bothwell et al. 1994; Leavitt et al. 2003).
In addition to affecting the quality and amount of lake-water DOM, anthropogenic climate warming may impact underwater light availability and the ecology of mountain lakes through changes in the ice and snow cover period. Transmittance of PAR and UV radiation to the water column through ice is dependent on the thickness of the snow and ice-cover (Bolsenga et al. 1996). Even though in some conditions the ice might be rather transparent to PAR and UV radiation, the high albedo and light attenuation capacity of even a small accumulation of snow reduces the amount of solar irradiance reaching the water column (Belzile et al. 2001). As UV radiation increases with elevation, and since mountain lakes have generally low DOM concentrations, organisms inhabiting these lakes are particularly vulnerable to changes in UV exposure (Sommaruga 2001). These changes can affect primary production through increased PAR availability but also through increased UV exposure (Williamson et al. 2009). Decreases in the ice-cover period and hence prolonged growing season may also affect benthic microalgae habitat availability and community composition (Griffiths et al. 2017). Warming and thermal stratification of previously well-mixed lakes can further lead to shifts from benthic to planktic microalgae community (Sorvari et al. 2002; Rühland et al. 2008, 2015; Saros et al. 2016).
Tolerance of algae to UV radiation depends on the species, but their survival is also linked to access to physical refugia (Vinebrooke and Leavitt 1999) and photoprotective strategies such as pigmentation (Williamson et al. 2009). Diatoms (Bacillariophyceae) have been shown to have trade-offs between motility and photoprotective capacity under extreme light conditions (Barnett et al. 2015), which points to the importance of underwater light for the functional composition of the phototrophic community. In addition, earlier findings show UV radiation may result in decreased primary production in benthic algal mats (Vinebrooke and Leavitt 1999; Tanabe et al. 2019) and UV-induced photoinhibition in diatoms (Bothwell et al. 1994), which point to potentially adverse effects of UV on algal carbon sequestration.
Diatoms are widely used in paleoecological studies to reconstruct past environmental conditions and current ecological status of aquatic environments (Dixit et al. 1992; Smol and Stoermer 2010). Traditionally, the most precise taxonomic identification of diatoms has been used when assessing changes in environmental conditions or ecological status of waterbodies (Rimet and Bouchez 2012). However, recently applications based on ecological traits and functional groups have been introduced (Tapolczai et al. 2016). These approaches have been applied especially in river and stream ecological status assessment (Passy 2007; Berthon et al. 2011; Rimet and Bouchez 2012; B-Béres et al. 2016, 2017; Soininen et al. 2016) and are being used increasingly in lake studies (McGowan et al. 2018; Saros et al. 2019; Ruwer and Rodrigues 2022; Rantala et al. 2022) but are still underutilized in studies of long-term lake history. In this study we used functional groups in addition to diatom species to detect possible broader trait-based adaptations to UV radiation.
The diatom guild classification we use was described originally by Passy (2007) and consisted of three diatom ecological guilds (high, low, motile) based on diatom life-form and species’ ability to resist physical disturbance and to tolerate different nutrient levels. Later, Rimet and Bouchez (2012) modified the guild classification and added a separate planktic guild. The eco-morphological group classification of B-Béres et al. (2016) combines four ecological guilds and further divides diatoms within each guild to five size categories (1: 5–99 μm3; 2: 100–299 μm3; 3: 300–599 μm3; 4: 600–1499 μm3; 5: ≥ 1500 μm3) as in Berthon et al. (2011). This combined guild and cell volume-based classification is a useful method for studying important environmental factors (for instance, nutrient levels, physical disturbance, light; B-Béres et al. 2016, 2017). To our knowledge, eco-morphological groups, herein referred to as diatom functional groups, described by B-Béres et al. (2016), have not been used in paleolimnological studies thus far.
The aim of this study was to investigate the impacts of anthropogenic pressures on underwater-light availability, exposure to UV radiation, and diatom functional group distributions. The study focuses on the post-industrial era and on baseline conditions before the onset of intensive anthropogenic pressures. This study was carried out in two mountain lakes, located in the Italian Alps, with clear differences in altitude and catchment properties. The main hypotheses are that: (i) underwater PAR availability and exposure to UV radiation has changed during the past ~ 200 years due to increased anthropogenic pressures (climate warming, acidification) that may have resulted in shortening of the ice-cover period and changes in the lakes’ carbon dynamics; and (ii) that these changes are reflected in the diatom functional group distributions.
Materials and methods
Study area
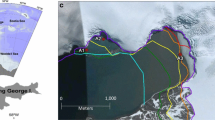
Lake Paione Superiore (LPS, N 46°10′ 14″, E 08°11′25″, 2269 m a.s.l) and Lake Paione Inferiore (LPI, N 46°10′14″, E 08°11′25″, 2002 m a.s.l.) are located in the Bognanco Valley, Central Alps in Piedmont, Italy (Fig. 1).

Map of study area and Paione lakes
LPS and LPI are the highest and lowest lakes in a cascading system of three glacially formed oligotrophic lakes located on the southern slope of the Italian Alps. The main characteristics of the lakes are listed in Table 1. The most distinct differences between the Paione lakes are that LPS is located at a higher altitude, it has a significantly smaller catchment and a history of heavy acidification compared to LPI. Otherwise, the lakes are relatively similar by their limnological properties, mean and maximum depths, and bowl-shaped lake basins (Cammarano and Manca 1997). Due to their shapes, UV irradiance was expected to be highest in the shallow littoral zone with a decreasing gradient towards the bottom of the lake. Water in the Paione lakes is highly transparent, with our coring device easily visible at the bottom of the lakes. The bottom of LPS is dominated by rocks; submerged vegetation was not recorded during sampling. Meanwhile, in LPI, submerged mosses were growing at the bottom of the lake. Small fish were observed in LPI but not in LPS during the sampling.
Based on 10-year measurements (2011–2021), the warmest month is July (mean temperature: 11 °C) and the coldest month is January (mean temperature: − 5 °C; Regional Agency for Environmental Protection of the Piedmont Region ARPA Piemonte). The Paione lakes have relatively short growing seasons with the duration of ice cover usually from November to June (Marchetto et al. 2004). Monitored data of air temperature from the area collected since the 1930s have shown an average annual increase rate of + 0.011 °C and average summer temperature increase of + 0.015 °C (Rogora et al. 2004). The June air temperature anomaly in the Paione lakes in 1991–2015 compared to baseline temperature in 1961–1990 was + 1.41 °C (Rogora et al. 2018).
Land cover in the catchments of LPS and LPI is mainly bare acidic rock consisting of clear banded orthogneiss and grey gneiss (Mosello et al. 1999), with thin soil cover of 33% and 39% of the catchment area, respectively (Rogora et al. 2003). The acidic soil has low base saturation, and the ion-exchange complex consists mainly of hydrogen and aluminum ions (Rogora et al. 2001).
Vegetation in the catchment of the lakes is scarce and consists of alpine meadows, Carex species and mosses.
Paione lakes have an acidification history as a consequence of heavy anthropogenic emissions in the region (Guilizzoni et al. 1992, 1996; Rogora et al. 2013). Based on diatom, chrysophyte and pigment-ratio reconstructions, pH in LPS started to decrease in the 1930s and the lake acidified by the 1960s compared to baseline conditions (Guilizzoni et al. 1992, 1996). From LPI, previous paleolimnological records are not available for comparison. Monitoring data of the Paione lakes since 1984 show that LPS’s pH increased from 5.5 to 6.5 by 2021, while pH in LPI increased from 6.1 to 6.7 within the same time frame (Supplementary Fig. 1). The more pronounced acidification in LPS is related to lower buffering capacity compared to LPI (Mosello et al. 1999). Even though the catchments of the Paione lakes share multiple similar features, the difference of pH values, alkalinity and base cation concentrations in the lakes are a result of LPI having small amounts of more soluble minerals (e.g., micaschists) in its catchment, which has resulted in neutralization of acidifying inputs (Mosello et al. 1999). SO4 emissions in the area have been estimated to have decreased up to 70% since the 1980s but atmospheric deposition of nitrogen (N) compounds has remained relatively high between 1990 and 2010 (Rogora et al. 2013). Lake-water nitrogen and silica concentrations and pH during the monitoring period 1984–2020 are presented in the supplementary material (Supplementary Fig. 1.) In addition to atmospheric human perturbations, fish (Onchorynchus mykiss) have been introduced to LPI since the 1960s (Cammarano and Manca 1996).
Sample collection and preparation
Two sediment cores from LPS and LPI were collected from the deepest basins in August 2020 using a UWITEC gravity corer (UWITEC Ltd., Austria). The cores were subsampled at 0.5-cm intervals for the first 20 cm of the core and at 1-cm intervals for the rest of the core. For the diatom analysis, the first 2 cm were analysed at 0.5-cm intervals followed by thicker (1 to 2 cm) intervals. In total, 21 sediment samples (to 37.5 cm) from LPS and 25 sediment samples (to 46 cm) from LPI were subjected to diatom analysis. Percentages of carbon (C) and nitrogen (N) and stable isotopes of C and N (δ13C, δ15N) were analysed from bulk sediment at the NIWA Environmental and Ecological Stable Isotope Facility (Wellington, New Zealand) at the same intervals as diatoms. Spectral analyses of inferred lake-water total organic carbon (TOC), sediment chlorophyll-a (CHLa) and pigments of primary producers’ chlorophylls and their derivates (CD) were analysed at 0.5-cm intervals.
Contemporary epilithon samples were collected from three depths (30, 80, 130 cm) by pooling samples from five 5–15 cm diameter stones at each depth. The diatom community inhabiting the surface of the stone were scraped into a vial using a toothbrush. Submerged vegetation was not found in LPS but in LPI submerged mosses were collected from 30 and 70 cm depths as epiphyton samples. The sediment cores and contemporary habitat samples were preserved in 70% ethanol and stored in darkness at + 5 °C.
Diatom slide preparation followed standard procedures as described by Renberg (1990) where wet sediment was added to hydrogen peroxide (H2O2) to degrade organic material. 37% hydrochloric acid (HCl) was added and the samples were centrifuged for four minutes at 4000 rpm after which the supernatant was decanted, and the tubes were filled again with MilliQ water. The washing was repeated five times, checking for the absence of diatom valves in the discarded supernatant. The samples were mounted on microscopic slides with Naphrax (Brunel Microscopes Ltd). A minimum of 300 diatoms were identified to the species or subspecies level from each sample using a light microscope with 1000-fold magnification. Identification was based mainly on the flora of Krammer and Lange-Bertalot (1986, 1988, 1991a, 1991b) with nomenclature updated where relevant. The identified diatoms were further divided into eco-morphological groups as described in B-Béres et al. (2017).
Sediment core chronology
Radiometric dating methods were used to construct a temporal framework for the sediment cores. The analyses were carried out at the Paleoecological Environmental Assessment and Research Laboratory (PEARL, Queen's University, Canada). Age estimates were obtained from lead (210Pb) dating of the recent (0–17 cm LPS, 0–19 cm LPI) sediments testing three different models, including the constant flux constant sedimentation (CFCS), constant initial concentration (CIC), and constant rate of supply (CRS) models. Additionally, the activity of cesium (137C) was analysed.
Plant macrofossil samples at 26.5 cm (terrestrial leaf) and 38 cm (aquatic moss) in LPS and at 31.5 cm (aquatic moss) and 37.5 cm (aquatic vascular plant) sediment depth in LPI were used for accelerator mass spectrometry (AMS) radiocarbon (14C) dating carried out at the Radiochronology Laboratory (Laval University, Canada). Terrestrial plant macrofossil remains were very few in the sediments, however, moss samples may be considered suitable for radiocarbon dating in the studied lakes owing to their soft and well-mixed waters (Wolfe et al. 2004; Oswald et al. 2005).
Age-depth models were built using clam version 2.4.0 (Blaauw 2010) in R (R Development Core Team 2022). IntCal20 (Reimer et al. 2020) Northern Hemisphere terrestrial calibration curve was used for radiocarbon-age calibration and the age-depth models were built with smooth spline with spar (smooth 0.7 LPS, smooth 0.6 LPI). Calculations were done at 95% confidence ranges with 1000 iterations and with calendar-age-point estimates based on the weighted average of all age-depth curves.
Geochemical analyses
Sediment samples were analysed by visible near infrared (VNIR) spectroscopy at PEARL (Queen’s University, Canada) to infer lake-water TOC and sediment CHLa. The resulting data were used to examine the past trends in lake-water transparency and to track trends in aquatic primary production. To prevent interference caused by water or particle-size differences, sediment samples were freeze dried and sieved through 125 µm mesh prior to spectral analysis. The method described by Michelutti et al. (2010) based on CHLa absorbance at 650–700 nm was used for the quantification of sediment CHLa; however, sediment chlorophyll-a concentrations were inferred using log-transformed data from Michelutti et al. (2010) with the equation: Chlorophyll-a + derivatives = EXP(0.83784*LN(peak area 650–700 nm) + (− 2.48861)). VNIR scanning of the samples was performed by a Model 6500 series Rapid Content Analyzer (FOSS NIRSystems Inc., Hillerød, Denmark) through the base of glass vials using diffuse reflectance mode. Wavelengths of 400–2500 nm were measured at 2 nm resolution.
Past lake-water TOC concentrations were determined using a partial least squares regression (PLSR) model between VNIR spectral information (400–2500 nm) of surface sediments and corresponding surface water TOC measurements from 345 northern lakes, with a TOC gradient from 0.5 to 41 mg L−1 (Meyer-Jacob et al. 2017). Applications of the model to lakes located outside of the model’s geographic calibration range accurately captured trends in TOC monitoring data, indicating a widespread applicability of the approach (Meyer-Jacob et al. 2017). TOC can be interpreted as nearly equivalent to DOC as DOC strongly dominates the TOC pool in lake water TOC calibration model (~ 87%; Meyer-Jacob et al. 2017).
Pigments of primary producers’ chlorophylls and their derivatives (CD) were extracted using the methods described by Lami et al. (2009) to provide supporting estimates of long-term trends in lake primary production. HPLC-grade solvents were used to extract pigments overnight in dark at 4 °C and afterwards the extracts were clarified by centrifugation. Sample aliquots were read in UV–Vis spectrophotometer (SAFAS, Monaco). Pigment data were expressed as units per g of organic carbon (Lami et al. 2009).
Data analysis
Unconstrained ordination analysis was conducted to investigate variation in diatom functional group distribution through time. Detrended correspondence analysis (DCA) showed short (< 1 SD) gradient lengths and therefore linear based principal component analysis (PCA) was selected. The analysis was conducted with square-root-transformed diatom functional group relative abundances. R (R Core Development Team 2022) with vegan (Oksanen et al. 2022) and ggbiplot (Vu 2011) packages were used for PCA and figures of the sediment cores' diatom functional group distributions. Spearman correlation analysis was carried out between the PCA axis scores (axes 1 and 2) and geochemical proxies for the estimation of the main environmental gradients.
Results
Age-depth model
210Pb ages modelled with constant rate of supply models (CRS) were chosen for the Paione lakes’ age-depth models (Fig. 2). Based on the age-depth model, the sediment core from LPS covers a time period of ca. 1450–2020 CE and in LPI ca. 1300–2020 CE. Total and supported 210Pb and 137Cs activities in the sediments of LPI and LPS are shown as supplementary material (Supplementary Fig. 2). 14C ages between plant macrofossil types may give different age estimates (Oswald et al. 2005) and therefore the age estimates of each sample were compared to each other within and between the lakes. LPI’s 14C sample at 31.5 cm depth was rejected from the model as the result was significantly older compared to the other dating results. 14C results are shown as supplementary material (Supplementary Table 1).

Age-depth model for LPI (a) and LPS (b) representing estimated ages (year common era (CE)) at different depths within the two studied sediment cores. Smooth splines were used in both lakes with a smoothing level of 0.7 in LPS and smoothing level of 0.6 in LPI. Age-depth models include constant rate of supply (CRS) modelled 210Pb ages (grey rectangle) with error bars and AMS 14C ages (Supplementary table 1; dark grey rectangle) with error bars. The light grey envelope represents the 95% confidence interval, with best age estimates in dark grey line
In both lakes, cesium (137Cs) activities showed an increasing trend initiated at around 5 cm, with peak values occurring near the sediment surface (1.25 cm in LPS, 0.25 cm in LPI). Previously, Guilizzoni et al. (1996) could not detect an activity peak related to the 1960s atomic bomb testing in LPI; however, a strong 137Cs activity peak corresponding to the 1986 Chernobyl accident was detected. In our sediment cores, the 137Cs results were rejected from the final age-depth models as the peaks were too young, likely influenced by post-fallout deposition of 137Cs from the catchment soils (Klaminder et al. 2012). Recent research also suggests that, in areas heavily influenced by Chernobyl fallout, delayed input of hot radioactive particles may result in anomalous 137Cs peaks in lake-sediment records (Appleby et al. 2022).
Several lines of evidence confirm that the sediment cores were not disturbed; visual examination in the field and the laboratory showed that the surfaces of the sediment cores were intact, and the cores further displayed irregular but frequent horizontal laminae that showed no signs of disturbance. In addition, 210Pb activities showed the expected consistent increasing trends towards topmost sediment samples and the geochemical proxies (in particular, CHLa, TOC, δ15N) displayed consistent directional changes near the sediment surface.
Sediment core geochemistry
Mean δ13C in the sediments of LPS was lower (− 26.8‰) compared to LPI (− 24.8‰). The δ13C values in LPS stayed fairly stable until the 1950s and thereon declined toward the surface (Fig. 3). In LPI, δ13C values showed a small and gradual increasing trend over the study period. δ15N trends in LPS and LPI (mean 2.3‰ and 2.4‰, respectively) were somewhat similar, starting around 3‰ during ca. 1600s followed by a synchronous decline to values below 0‰ from ca. 1940s onward. A peak was recorded around the 1940s, which was especially pronounced in LPS. δ15N declined in both lakes to − 1‰ in the topmost sample (Fig. 3).

Analysed geochemical proxies from the Paione lakes’ sediment-cores. Stable isotopes of carbon and nitrogen (δ13C and δ15N), carbon content (C%), UV–Vis inferred chlorophyll derivatives (CD µg/g C org−1) and VNIR spectroscopically inferred chlorophyll-a (CHLa mg L−1), carbon and nitrogen ratio (C/N atomic ratio), nitrogen content (N%) and inferred lake-water total organic carbon (TOC mg L−1)
C% in the Paione lakes was 4–5% in the beginning of the studied period. Around 1700 CE, C% increased to 7.5% followed by a decline to 4% by 1800 CE. This was followed in both lakes by a C% increase to 5% around 1900 CE, followed by a decrease to nearly 3% in LPS and 4% in LPI in the 1940s (Fig. 3). After an increase to 5% in the 1960s in both lakes, C% increased to 6% in LPS and decreased to 4% in LPI. Patterns in N% were similar to those of C%.
The mean C/N atomic ratio was 12.9 in LPI and 15 in LPS. In LPS, C/N ratios were highly variable but showed an overall declining trend from ratios around 16 to ratios around 13 at the sediment surface (Fig. 3). In LPI, C/N ratios were relatively steady between 12.5 and 14 over the studied period and had the lowest C/N ratios around the mid-eighteenth century.
Sediment inferred CHLa patterns were similar between the Paione lakes but LPI had a higher mean concentration (0.02 mg g−1) compared to LPS (0.01 mg g−1; Fig. 3). CHLa showed few consistent trends in the lower sections of the sediment cores but a subtle increase was observed in both lakes beginning around the 1940s towards the topmost samples. A steep increase near the surface could be at least partially related to diagenetic processes, which generally occur during the first decade after deposition (Rydberg et al. 2020).
Previous applications of VNIR spectroscopy to sediment records showed that inferred TOC trends closely correspond to trends of water-chemistry monitoring data, highlighting the ability of the technique to track long-term TOC trends (Meyer-Jacob et al. 2017, 2019). However, absolute concentrations should be interpreted with care because they are frequently over/underestimated compared to measured data. In Paione lakes, contemporary TOC levels are overestimated by ~ 2 to 3 mg L−1 (Table 1; Fig. 3). After ca. 1700 CE, LPS showed an increase of TOC before starting to decline in the 1940s (Fig. 3). LPI showed a decline of inferred TOC around 1850 CE and an increase in the 1940s. After the 1940s, lake-water TOC concentrations decreased with lowest value around 2000 CE. In the topmost sample, TOC showed a subtle increase in both lakes (Fig. 3).
Contemporary habitat samples from high irradiance littoral zone
In LPS, habitat samples consisted of epilithon samples from three different depths (30, 80, 130 cm). LPI had similar epilithon samples and additional epiphyton habitat samples from 30 and 70 cm depths. The aim with these samples was to support the interpretation of the sediment data by detecting diatom taxa and traits in the main habitats of the lakes that are likely tolerant to high UV radiation. In the epilithon samples of LPS, the dominant functional group was L2 (49–55%) followed by L3 (19–20%) and M5 (6–13%). Other functional groups observed were present in smaller relative abundances (0–5%). Commonly observed diatom taxa were Psammothidium marginulatum (L2), P. helveticum (L3) and Pinnularia microstauron (M5).
In the epilithon samples of LPI, dominant functional groups were L1 (30–41%), L2 (14–18%), L3 (8–20%), H4 (8–12%) and M3 (7–13%). Other observed functional groups were present in small relative abundances (0–6%).
Same diatom functional groups as in epilithon-habitat samples dominated in epiphyton samples, including L1 (18–23%), L2 (18–24%), L3 (6–20%), H4 (8–12%) and M3 (7–8%). Other observed functional groups represented 0–10%.
Commonly observed diatom taxa in both LPI habitat types were Brachysira vitrea (L2), Achnanthidium minutissimum (L1), Psammothidium microscopicum (L1), P. helveticum (L3) and Fragilaria arcus (H4).
Diatom functional group stratigraphy
Paione Superiore
Among the functional groups representing at least 1% of total taxa abundance of the identified diatoms from LPS, taxa of the high group (H) were represented in size classes 1, 2, 4 and 5, taxa of the low group (L) in size classes 1–4, taxa of the motile group (M) in size classes 2, 4 and 5, and taxa of the planktic group (P) in size class 2 (Fig. 4). The full list of analysed diatom taxa with corresponding functional group is provided in Supplementary Table 2 and the diatom-species stratigraphy of LPS is provided in Supplementary Fig. 3.

Stratigraphic diagram of relative abundances (%) of the most common functional diatom groups representing at least 1% of total abundance of LPS’s sediment core as a function of sediment age (CE). Letter in the abbreviation stand for name of the group (i.e., H = high, L = low, M = motile, P = planktic) and number for size class (1–5) based on diatom-cell biovolume
For the high groups (i.e., taxa with a long stature and more vulnerable to physical disturbance), the common size classes were relatively stable throughout the studied period (H1 relative abundance 0–8%, H2 1–10%, H4 4–12%, H5 0–6%; Fig. 4). The most common high group taxa were Encyonema silesiacum (H4), Eunotia exigua (H1), E. intermedia (H2) and E. praerupta (H5).
Two of the low group (i.e., taxa tolerant to physical disturbance and low nutrient concentrations and tightly attached to their substrate) size classes, L1 and L2, were represented as smaller relative abundances through the core (2–9% and 0–3%, respectively), whereas L2 and L3 had a higher relative share (5–22% and 10–31%, respectively). L2 had small peaks up to 10% around ca. 1600 CE and up to 17% ca. 1950 CE followed by a decline to 8% in the 1980s (Fig. 4). During the 2000s, L2 represented 20% of the functional groups. L3 peaked at 29% in ca. 1540 CE, followed by a decline to 10% around 1890 CE, and then increasing again to 27% in ca. 1790 CE. Around 1890 CE, L3 declined to 12% and peaked again in the 1970s reaching 17%. From the 1980s to the present, L3 declined to 16% (Fig. 4). Frequently observed low group species were Psammothidium helveticum and its varieties (L3), P. marginulatum (L2), P. microscopicum (L1), P. scoticum (L1), Encyonopsis cesatii (L4) and Cymbella gaeumannii (L2).
Motile groups M2 and M4 were present at relative abundances of 4–10% and 1–6% throughout the sediment core (Fig. 4). Group M5 was represented as relative abundance of 9 to 49%. The highest abundance of 49% occurred around 1610 CE and was followed by a gradual decline to 20% in ca. 1860 CE. In ca. 1890 CE, M5 increased to 45% and since then declined reaching its lowest abundance of 9% in the topmost sample representing present time. Common identified motile groups taxa were Pinnularia microstauron (M5), Neidium alpinum (M2), N. ampliatum (M5) and N. affine (M4).
The planktic group P2 was represented from 0 to 29% in the sediment core. Since ca. 1790 CE, P2 was present at 1 to 5% and after the 1970s started to rapidly increase reaching its highest relative abundance of 29% in the top most sample (Fig. 4). The most common species observed were Aulacoseira nivalis (P2), A. alpigena (P2) and A. lirata (P2).
Paione Inferiore
Among functional groups representing at least 1% of total taxa abundance of the identified diatoms from LPI, the high group (H) was present in size classes 2–4, the low group (L) in size classes 1–3, the motile group (M) in size classes 1–5, and the planktic group (P) in size class 5 (Fig. 5). The full list of analysed diatom taxa and the corresponding functional group is provided in Supplementary Table 2 and species stratigraphy of LPI is provided in Supplementary Fig. 4.

Stratigraphic diagram of relative abundances (%) of the most common functional diatom groups representing at least 1% of total abundance of LPI’s sediment core as a function of sediment age (CE). Letter in the abbreviation stand for the name of the group (i.e., H = high, L = low, M = motile, P = planktic) and number for size class (1–5) based on diatom-cell biovolume
The high group with its classes was observed throughout the sediment core (H2 4–12%, H3 1–6%, H4 4–11%; Fig. 5). Common high group taxa observed were Fragilaria capucina and its variations (H2), Fragilaria arcus (H4), Gomphonema parvulum (H4), and Eunotia naegelii (H4).
L1 was the dominant group representing relative abundance of 19–47% in the sediment core. L1 showed peaks of 47% in ca. 1900 CE and the 1960s. After this it declined to 22% in the ca. 1990s and then increased to 37% in ca. 2015 CE but declined in the topmost sample to 19% (Fig. 5). L2 was represented in smaller amounts (3–18%) compared to L1 (19–47%) but followed generally similar patterns with the exception that L2 reached its highest abundance of 18% in the topmost sample. L3 and L5 were represented as smaller amounts (2–10% and 1–10%, respectively). L3 had a peak of 10% in the ca. 1980s followed by a decline to 2% in the topmost sample. L5 was present throughout the sediment core as 2–4% with the exception of a peak of 10% in the topmost sample (Fig. 5). Commonly observed taxa were Achnanthidium minutissimum (L1), Psammothidium microscopicum (1), P. helveticum (L1), P. subatomoides (L2), Brachysira vitrea (L2), Amphora libyca (L5) and Cymbella proxima (L5).
Groups M1 and M2 represented relative abundances of 0–3% and 0–4%, respectively. M3, M4 and M5 had larger share of 5–14%, 4–11% and 3–16%, respectively (Fig. 5). Common observed taxa were Pinnularia microstauron (M5), Stauroneis neohyalina (M3), Navicula cryptotenella (H3), Nitzschia recta (H4) and N. palea (H3).
P5 was represented as 1–12% in the sediment core. It had peaks of 11% in ca. 1900 CE, 12% in ca. 1990 CE and 10% in the topmost sample (Fig. 5). Common observed taxa were Aulacoseira valida (P5) and A. nivaloides (P5).
Paione Superiore PCA
Principal component 1 (PC1) explained 57.5% of variation of the LPS's analysed sediment depths and diatom functional groups and principal component 2 (PC2) explained 13.2% of the variation (Fig. 6). Sediment inferred lake-water TOC, δ15N and δ13C values had positive correlations with PC1 (R = 0.86; p < 0.001, R = 0.93; p < 0.001 and R = 0.87; p < 0.001, respectively) and sediment inferred CHLa had a negative correlation (R = −0.79; p < 0.001). PC2 did not have significant correlations with the geochemical proxies.

Principal component analysis (PCA) of LPS’s most common diatom functional groups representing at least 1% of total abundance and estimated sediment ages (CE). Letters in the abbreviation stand for the name of the group (i.e., H = high, L = low, M = motile, P = planktic) and number for size class (1–5) based on diatom-cell biovolume
Paione Inferiore PCA
Principal component 1 (PC1) explained 27.9% of the variation of the LPI's analysed sediment depths and diatom functional groups and principal component 2 (PC2) explained 18.1% of the variation (Fig. 6).
Sediment CHLa, N%, and C% had positive correlations with the main axis PC1 (R = 0.75, p < 0.001; R = 0.66, p < 0.001; R = 0.66, p < 0.001, respectively). Sediment inferred lake-water TOC and δ15N had a negative correlation with PC1 axis (R = −0.62, p < 0.001; R = −0.73, p < 0.001). PC2 axis had significant negative correlations with CHLa, N% and C% (R = −0.41, p = 0.04; R = −0.43, p = 0.03; R = −0.46, p = 0.02, respectively).
Discussion
Impacts of anthropogenic pressures on Paione lakes’ underwater light availability and UV radiation
The distinct decline in lake-water TOC levels (Fig. 3) during the post-industrial era suggests increased UV exposure and underwater light availability in the Paione lakes during the past century, and particularly from the 1940s onward, confirming our hypothesis (i). Lake-water DOC, the main component of inferred lake-water TOC (Meyer-Jacob et al. 2017) is mainly controlling underwater light availability (Williamson et al. 2009) and even a small decrease, particularly in the allochthonous fraction, can lead to significant increases in UV exposure in lake ecosystems where the baseline organic carbon concentrations are low (Schindler et al. 1996; Laurion et al. 1997). In transparent, shallow alpine lakes, this may lead to a situation where UV radiation reaches even the bottom parts of lakes leaving aquatic organisms without a possibility to escape the harmful irradiance (Schindler et al. 1996). The role of light in these ecosystems is complex, as PAR fuels life and carbon sequestration through photosynthesis, but especially the shorter wavelength UV radiation has adverse effects on primary production through photoinhibition (Tanabe et al. 2019) and it modifies lakes’ dissolved carbon pools by increasing carbon availability for microbial degradation (Nevalainen et al. 2020).
The geochemical data indicate that the steep decline in TOC around the 1940s is primarily a consequence of declining terrestrial carbon inputs. C/N ratios were generally elevated deeper in the sediment-cores, indicative of higher contribution of terrestrial organic carbon (Meyers 1994), compared to the recent period characterized by declining lake-water TOC. In addition, the sediment chlorophyll concentrations, indicative of aquatic primary production, showed a subtle increase in both lakes coincident with the declining trend in lake-water TOC (Fig. 3). Due to differences in optical characteristics, carbon from terrestrial sources absorbs light and UV radiation more efficiently compared to carbon from aquatic primary production (McKnight et al. 1997; Sommaruga et al. 1999). A decrease in terrestrial carbon input could be related to acidification reducing organic carbon mobility in catchment soils (Evans et al. 2012).
Considering the timing of the lake-water TOC decline in the ~ 1940s in the Paione lakes (Fig. 3), the pattern is likely related to acidification, which has caused widely observed decreases in lake-water DOC/TOC levels in lakes subjected to atmospheric acidifying inputs (Meyer-Jacob et al. 2019). In the earlier reconstructions of the pH of LPS, the onset of acidification occurred in the ~ 1930s (Guilizzoni et al. 1992, 1996). The PCA analysis (Figs. 6, 7) also suggests a pronounced impact of anthropogenic acidification and pH decline in LPS as the time periods before and after the onset of the acidifying emissions were plotted on the opposite sides along the main axis. In the PCA analysis of LPI (Fig. 7), a similar distinct pattern was not seen. Changes in long-term TOC trends have been observed in other relatively well-buffered systems without an acidification history (Hruska et al. 2009; Meyer-Jacob et al. 2019). Acidic deposition can change soil organic matter solubility and thus mobility through its effects on soil acidity and ionic strength of the soil solution (Monteith et al. 2007). This may explain the similar declining TOC trends in both Paione lakes, even though LPI has not undergone a pronounced pH decline as LPS.

Principal component analysis (PCA) of LPI’s most common diatom functional groups representing at least 1% of total abundance and estimated sediment ages CE. Letters in the abbreviation stand for the name of the group (i.e., H = high, L = low, M = motile, P = planktic) and number for size class (1–5) based on diatom-cell biovolume
The distinct decreasing trend in sediment δ15N values in the Paione lakes after the ~ 1940s could as well be related to industrial emissions (Fig. 3), as declines in δ15N have been associated with increased anthropogenic reactive nitrogen deposition (Enders et al. 2008, Holtgrieve et al. 2011). The low observed values of the δ15N values are consistent with recent findings that the area still receives relatively high concentrations of nitrogen input, regardless of the mitigation of industrial emissions since the 1980s (Rogora et al. 2016). However, other processes likely contributed to the sustained decline in δ15N particularly over the past couple of decades since the levels of nitrogen deposition measured at nearby meteorological stations have remained relatively stable. Post-depositional alteration may be one factor as diagenetic processes have been shown to shape δ15N profiles in sediment organic matter (Gälman et al. 2009). Moreover, investigation of the lake-water TOC, which has not increased back to pre-industrial levels in either lake, suggests that the Paione lakes may not have fully recovered from the earlier acidifying deposition, even though modern measurements indicate that acidifying sulphur emissions in the area have decreased (Rogora et al. 2016).
Based on the geochemical data (Fig. 3), Paione lakes’ lake-water carbon pools have undergone distinct changes because of anthropogenic acidification. Indications of the effects of warming climate on the lakes' carbon dynamics were more difficult to discern and could be either muted by or intertwined with the effects of acidification. The gradual increasing trends in both δ13C values and sediment CHLa and CD in LPI could indicate a slight increase in overall primary production (Meyers 2003) over the past centuries (Fig. 3). In LPS, the decline in δ13C values starting after the ~ 1940s (Fig. 3), observed with a parallel increase in planktic diatom life-forms (Fig. 4), may together point to increased pelagic primary production that tends to yield more 13C-depleted organic matter compared to benthic algae with diffusion-limited carbon uptake (Meyers 1994; Hecky and Hesslein 1995). In LPI, primary production has remained mainly benthic (Fig. 5) as suggested by the lower δ13C values (Fig. 3) typical of benthic primary production mixed with terrestrial organic matter signal (Meyers 1994; Hecky and Hesslein 1995).
Catchment greening above the tree line related to increased summer temperatures has been observed widely in the Alps during the past decades, however, the patterns have not been uniform (Choler et al. 2021). The most pronounced greening has been observed on the northern slope of the Alps in the very scarcely vegetated areas (Choler et al. 2021), whereas the Paione lakes are located on the southern slope. The non-uniform distribution of the greening and the location of Paione lakes could explain why evidence of lake-water browning linked to catchment greening were not found in the geochemical proxies, even though the lakes are in a sensitive ecotonal area just above the tree line.
Anthropogenic climate warming is projected to exceed a global temperature rise of 1.5–2 °C compared to pre-industrial era within the twenty-first century, unless remarkable CO2 reductions are made within the upcoming few decades (IPCC 2021). The timberline in the greater Alpine region, currently situated at 1880 m a.s.l., is projected to move higher up between 2120 and 2820 m by the end of the twenty-first century, depending on the GHG scenario used (RCP 2.6, RCP 8.5, respectively; Rubel et al. 2017). Based on the projections, anthropogenic warming will have major effects on the vegetation of the Alpine region in less than 100 years. In addition to direct impacts of climate warming on mountain lakes, the changes in the catchment can cascade to the tightly catchment-connected lakes in similar altitudes as the Paione lakes (LPS, 2269 m a.s.l, LPI 2002 m a.s.l.) For example, climate change can increase the terrestrial organic carbon load following warming-induced changes in vegetation type and soil formation as well as by possible changes in hydrology.
UV-tolerant diatom functional groups
Littoral zones of mountain lakes are often stressful environments, with diatoms inhabiting the shallow littoral subjected to multiple pressures in addition to UV exposure. Especially in LPS, intensive acid shock during spring snow melt (Cammarano and Manca 1999) will likely affect diatom community composition and favor taxa tolerant to acidic conditions. The shallow littoral area is also characterized by high physical turbulence (water-level fluctuations, waves) and desiccation during winter (Cantonati et al. 2009; Leira et al. 2015).
The contemporary diatom samples from the Paione lakes consisted of samples of the lakes’ main habitats in the shallow littoral zone and were dominated by low guild functional group diatoms. These taxa are attached tightly to different substrates, often associated with the epipsammon and epilithon, and can tolerate physical turbulence and low nutrient conditions (Passy 2007). This group has also been found to be indicative of earlier stages of biofilm development, whereas the abundance of motile and high group taxa have been shown to increase in well-developed biofilms (B-Béres et al. 2016). Due to the fast regeneration time of small low guild diatoms, they have the capacity to recover quickly after disturbances like water-level fluctuations (Stenger-Kovács et al. 2013). The motile and high group diatom taxa are more sensitive to physical disturbances, but tolerant to higher nutrient levels compared to low group taxa (Passy 2007). The motile group also has the competitive advantage of moving into more favorable positions in the biofilm (Passy 2007).
The same functional groups (LPS: L2, L3, M5; LPI: L1, L2, L3, H4, M3) dominated regardless of sampling depth (30–130 cm), suggesting environmental conditions being somewhat similar in this short depth gradient. In LPI, where epiphyton samples were collected in addition to epilithon samples, the same functional groups dominated in both habitats. The high guild taxa typically have long stature and are therefore vulnerable to turbulence, and plant surfaces may offer them shelter (Passy 2007). This could explain their occurrence in the LPI habitat samples, even though the observations were not limited to epiphyton samples. In LPS, only three functional groups (L2, L3, M5) and in LPI five groups (L1, L2, L3, H4, M3) dominated both habitat types and the rest of the functional groups were represented between 0 and 6% in all analysed habitats.
The low guild diatom taxa mainly formed the modern biofilm in the lakes with smaller proportions of motile and high guild taxa. Based on the community composition and characteristics of the observed functional groups, the results suggest that physical disturbance (waves, water-level changes) have impacted diatom community composition. In addition, low group taxa may be more tolerant to high solar irradiance (Stenger-Kovács et al. 2013). Even though UV radiation is not the only parameter limiting the occurrence of other diatom functional groups in the high irradiance littoral zone, the low guild taxa must withstand high UV exposure in the absence of physical UV refugia and given the typically limited mobility of low guild diatoms.
Long-term diatom functional group responses
Low guild diatom taxa, which showed tolerance to high UV radiation based on contemporary habitat sampling, dominated in both of study lakes across the past centuries. Low guild diatoms are taxa with short stature consisting of prostrate, adnate and erect diatoms typically attached to their substrate (Passy 2007). This guild tolerates nutrient limitation in biofilms and high physical disturbance (Passy 2007) and their sustained dominance in the Paione lakes is therefore not surprising. Diatom functional groups in the sediment cores mainly represented the three smallest size classes. Small diatom frustule size has been previously associated with more efficient nutrient uptake and small cells may be less sensitive to physical disturbance (Smol et al. 1984; Tapolczai et al. 2016).
Based on the diatom records (Figs. 4, 5), differences in lake resilience were apparent in the biological communities. However, the hypothesis related to diatom responses to light and UV exposure (hypothesis ii) could not be confirmed as the responses in the lakes varied despite the uniform trajectories in lake-water TOC. Moreover, the role of light and UV radiation could not be reliably distinguished from other drivers. Although the diatom communities of LPI remained almost the same throughout the studied period (Fig. 5), one third of the diatom community of LPS shifted from benthic to planktic within the past century (Fig. 3). The timing of this change suggests it is caused by anthropogenic drivers. The difference in responses to anthropogenic acidification in the Paione lakes was also evident in the PCA (Figs. 6, 7), where the PCA analysis of LPS showed a clear trend related to the onset of acidification (Fig. 6).
The most pronounced change in the Paione lakes' diatom functional groups analysed from the sediment cores was the gradual increase in the planktic (P2) group. A parallel and distinct decline in the motile group (M5) in LPS and a decline in the M2 and M4 groups within the past decades could also be observed (Fig. 3). The planktic life-forms in the LPS sediment core were representing Aulacoseira genus, which has been related to turbulent and well-mixed water columns (Rühland et al. 2008, 2015), whereas motile life-forms do not tolerate turbulence well (Passy 2007) which could contribute to the opposite trends of the traits in LPS (Fig. 2). Furthermore, the main taxon representing the P2 group was Aulacoseira nivalis, which has a lower pH optimum compared to the dominating taxa of the declining motile groups, namely Pinnularia microstauron (M5), Neidium alpinum (M2) and Neidium affine (M4; Van Dam et al. 1994; Cameron et al. 1999). Legacy effects of heavy acidification in LPS could therefore be one of the explaining factors behind the increase of the P2 group. The M5 group, namely P. microstauron, showed an increasing trend in the early seventeenth century followed by a decline (Fig. 4). Therefore, it is possible that the decreasing trend within the past century is related to other environmental factors (like localized plant or moss growth), which were not monitored in this study.
Aulacoseira taxa are tychoplanktonic and able to live also in benthic habitats. Since the ice cover melts first from the shoreline of lakes, the plasticity of Aulacoseira taxa may have given them competitive advantage during earlier ice-offs and also climate warming-induced longer growing season, here inferred from the subtle increase in primary production, could have been favorable for planktic life-form. Additionally, as an acidophilic taxon, Aulacoseira nivalis would not suffer from snow melt-induced acid shock during spring, which has been especially pronounced in LPS (Cammarano and Manca 1997). Even though the lake-water pH in LPS has increased during the past decades (Supplementary Fig. 1), the relative abundances of the acidophilous Aulacoseira taxa of the P2 group have continued to increase. Biological recovery of lakes from severe acidification is often slower compared to the chemical recovery, and biological communities may not recover in the same state that they were before acidification (Keller et al. 2019). Previous diatom studies from acidified regions have shown that recovery to pre-disturbance assemblages is rarely happening (Sivarajah et al. 2017).
Shifts in diatom communities linked to limnological changes associated with climate-warming effects (e.g., changes in snow regime, shortening of ice-cover period, increased habitat availability, changes in thermal stability) are widely observed phenomena (Rühland et al. 2008, 2015; Griffiths et al. 2017). Contrary to our findings, earlier lake studies have often found increases in small cyclotelloid diatoms concurrent with Aulacoseira declines as a consequence of warming-related strengthening of thermal stratification in lakes (Sorvari et al. 2002; Rühland et al. 2008, 2015; Edlund et al. 2022). However, the Paione lakes are not stratified and tychoplanktonic Aulacoseira could be expected to benefit from habitat expansion due to longer open water season and increased wind exposure leading to well mixed water column. The increase in planktic Aulacoseira found in LPS may be therefore linked to a climate warming-driven transition from a mainly benthic community to one sustaining also planktic life-forms (Smol 1988).
In seasonally snow-covered ecosystems, earlier snow melting and rapid flushing during spring affects a lake’s hydrology, as well as the nutrient and carbon dynamics through transportation of nutrients, acidic compounds and DOC (Miller and McKnight 2015; Sadro et al. 2018). The increase in tychoplanktic Aulacoseira may be expected with warming, but it could also be driven by a combination of increased N deposition and warming (Oleksy et al. 2020). The δ15N trends (Fig. 3) suggested anthropogenic N deposition has reached the Paione lakes during post-industrial era, but the concentrations of analysed lake-water NO3 showed no increase within the monitoring period 1984–2020 (Supplementary Fig. 1) indicating that at least the recent increase of Aulacoseira from 20 to 30% within the past two decades in LPS is not driven by increased overall nutrient input (Supplementary Fig. 1). Aulacoseira could also benefit from increased flushing of catchment silicates though modern measurements (1984–2020) of dissolved silica provide no evidence of increasing trends (Supplementary Fig. 1). The modern measurements of NO3 and silica were conducted in late autumn and thus do not exclude the possibility of warming-induced changes in the timing of snow melt and related nutrient pulse with possibly increasing flushing rates due to rapid snow melt, which also could contribute to the increase of spring blooming Aulacoseira. However, the gradual increase of Aulacoseira during the last century (Fig. 4) likely carries a signal of the earlier atmospheric deposition and related acidification coupled with the multiple effects of the accelerating anthropogenic climate warming.
Conclusions
We tracked a decline in lake-water TOC over the past century, which was followed by a recent increasing trend within the past two decades. This trend is likely linked to anthropogenic acidification and subsequent recovery of the lakes following emission mitigation, which was supported by long-term monitoring data. However, the TOC levels are still low compared to the pre-industrial era. The results of the sedimentary C/N ratios and CHLa suggest that underwater light availability and UV radiation have increased within the past century driven by the decline in terrestrial TOC inputs.
Even though the Paione lakes can be considered sensitive to climate warming, the warming-induced changes were subtle. The results suggest that multiple anthropogenic pressures (climate warming, former acidification) are driving the diatom community towards planktic functional groups in the lake situated at higher altitude (LPS), whereas the diatom community in the lake located at lower altitude (LPI) with a larger catchment and better buffering capacity did not show remarkable changes within the study period. The exact role of light availability and UV exposure as a driver of diatom functional group distributions could not be distinguished, but the dominance of low guild diatoms in the contemporary habitats in the shallow littoral zone as well as in the sediment cores suggests that the low guild is tolerant to high UV radiation and the other stressors in the littoral zones of these lakes (physical disturbance, and acid shock during spring in LPS).
The study benefited from the combination of contemporary diatom sampling with a long-term paleolimnological perspective, as well as the assessment of diatom responses at various levels of organization (species, genus, functional group). The combination of these data with multiproxy geochemical analysis helped reveal the impacts of the multiple anthropogenic pressures on the lakes’ development during the post-industrial era. However, resolving the role of underwater light in driving diatom functional group distributions will need further studies to separate the tightly intertwined effects of changing availability of habitats and light.
Change history
08 April 2023
The original article has been revised
References
Allan RP, Hawkins E, Bellouin N, Collins B (2021) IPCC, 2021: summary for policymakers
Appleby PG, Piliposyan G, Hess S (2022) Detection of a hot 137Cs particle in marine sediments from Norway: potential implication for 137Cs dating. Geo-Mar Lett 42:2
Auer I, Böhm R, Jurkovic A, Lipa W, Orlik A, Potzmann R, Schöner W, Ungersböck M, Matulla C, Briffa K, Jones P, Efthymiadis D, Brunetti M, Nanni T, Maugeri M, Mercalli L, Mestre O, Moisselin J-M, Begert M, Müller-Westermeier G, Kveton V, Bochnicek O, Stastny P, Lapin M, Szalai S, Szentimrey T, Cegnar T, Dolinar M, Gajic-Capka M, Zaninovic K, Majstorovic Z, Nieplova E (2007) HISTALP—historical instrumental climatological surface time series of the Greater Alpine Region. Int J Climatol 27:17–46
Barnett A, Méléder V, Blommaert L, Lepetit B, Gaudin P, Vyverman W, Sabbe K, Dupuy C, Lavaud J (2015) Growth form defines physiological photoprotective capacity in intertidal benthic diatoms. ISME J 9:32–45
Belzile C, Vincent WF, Gibson JA, Hove PV (2001) Bio-optical characteristics of the snow, ice, and water column of a perennially ice-covered lake in the High Arctic. Can J Fish Aquat Sci 58:2405–2418
B-Béres V, Lukács Á, Török P, Kókai Z, Novák Z, T-Krasznai E, Tóthmérész B, Bácsi I (2016) Combined eco-morphological functional groups are reliable indicators of colonisation processes of benthic diatom assemblages in a lowland stream. Ecol Indic 64:31–38
B-Béres V, Török P, Kókai Z, Lukács Á, T-Krasznai E, Tóthmérész B, Bácsi I (2017) Ecological background of diatom functional groups: comparability of classification systems. Ecol Indic 82:183–188
Berthon V, Bouchez A, Rimet F (2011) Using diatom life-forms and ecological guilds to assess organic pollution and trophic level in rivers: a case study of rivers in south-eastern France. Hydrobiologia 673:259–271
Blaauw M (2010) Methods and code for ‘classical’ age-modelling of radiocarbon sequences. Quat Geochronol 5:512–518
Bolsenga SJ, Evans M, Vanderploeg HA, Norton DG (1996) PAR transmittance through thick, clear freshwater ice. Hydrobiologia 330:227–230
Bothwell ML, Sherbot D, Roberge AC, Daley RJ (1993) Influence of natural ultraviolet radiation on lotic periphytic diatom community crowth, biomass accrual, and species composition: short-term versus long-term effects1, 2. J Phycol 29:24–35
Bothwell ML, Sherbot DMJ, Pollock CM (1994) Ecosystem response to solar ultraviolet-B radiation: influence of trophic-level interactions. Science 265:97–100
Cameron NG, Birks HJB, Jones VJ, Berge F, Catalan J, Flower RJ, Garcia J, Kawecka B, Marchetto A, Sánchez-Castillo P, Schmidt R (1999) Surface-sediment and epilithic diatom pH calibration sets for remote European mountain lakes (AL:PE Project) and their comparison with the Surface Waters Acidification Programme (SWAP) calibration set. J Paleolimnol 22:291–317
Cammarano P, Manca M (1997) Studies on zooplankton in two acidified high mountain lakes in the Alps. Hydrobiologia 356:97–109
Cantonati M, Scola S, Angeli N, Guella G, Frassanito R (2009) Environmental controls of epilithic diatom depth-distribution in an oligotrophic lake characterized by marked water-level fluctuations. Eur J Phycol 44:15–29
Choler P, Bayle A, Carlson BZ, Randin C, Filippa G, Cremonese E (2021) The tempo of greening in the European Alps: spatial variations on a common theme. Glob Chang Biol 27:5614–5628
Dixit SS, Smol JP, Kingston JC, Charles DF (1992) Diatoms: powerful indicators of environmental change. Environ Sci Technol 26:22–33
Edlund MB, Ramstack Hobbs JM, Heathcote AJ, Engstrom DR, Saros JE, Strock KE, Hobbs WO, Andresen NA, VanderMeulen DD (2022) Physical characteristics of northern forested lakes predict sensitivity to climate change. Hydrobiologia 849:2705–2729
Enders SK, Pagani M, Pantoja S, Baron JS, Wolfe AP, Pedentchouk N, Nuñez L (2008) Compound-specific stable isotopes of organic compounds from lake sediments track recent environmental changes in an alpine ecosystem, Rocky Mountain National Park, Colorado. Limnol Oceanogr 53:1468–1478
Evans CD, Jones TG, Burden A, Ostle N, Zieliński P, Cooper MDA, Peacock M, Clark JM, Oulehle F, Cooper D, Freeman C (2012) Acidity controls on dissolved organic carbon mobility in organic soils. Glob Change Biol 18:3317–3331
Gälman V, Rydberg J, Bigler C (2009) Decadal diagenetic effects on δ13C and δ15N studied in varved lake sediment. Limnol Oceanogr 54:917–924
Griffiths K, Michelutti N, Sugar M, Douglas MS, Smol JP (2017) Ice-cover is the principal driver of ecological change in High Arctic lakes and ponds. PLoS ONE 12:e0172989
Guilizzoni P, Lami A, Marchetto A (1992) Plant pigment ratios from lakes sediments as indicators of recent acidification in alpine lakes. Limnol Oceanogr 37:1565–1569
Guilizzoni P, Marchetto A, Lami A, Cameron N, Appleby P, Rose N, Schnell Ø, Belis C, Giorgis A, Guzzi L (1996) The environmental history of a mountain lake (Lago Paione Superiore, Central Alps, Italy) for the last c. 100 years: a multidisciplinary, palaeolimnological study. J Paleolimnol 15:245–264
Häder D-P, Williamson CE, Wängberg S-Å, Rautio M, Rose KC, Gao K, Helbling EW, Sinha RP, Worrest R (2014) Effects of UV radiation on aquatic ecosystems and interactions with other environmental factors. Photochem Photobiol Sci 14:108–126
Hecky RE, Hesslein RH (1995) Contributions of benthic algae to lake food webs as revealed by stable isotope analysis. J N Am Benthol Soc 14:631–653
Holtgrieve GW, Schindler DE, Hobbs WO, Leavitt PR, Ward EJ, Bunting L, Chen G, Finney BP, Gregory-Eaves I, Holmgren S (2011) A coherent signature of anthropogenic nitrogen deposition to remote watersheds of the northern hemisphere. Science 334:1545–1548
Hruska J, Krám P, McDowell WH, Oulehle F (2009) Increased dissolved organic carbon (DOC) in Central European streams is driven by reductions in ionic strength rather than climate change or decreasing acidity. Environ Sci Technol 43:4320–4326
Keller W, Heneberry J, Edwards BA (2019) Recovery of acidified Sudbury, Ontario, Canada, lakes: a multi-decade synthesis and update. Environ Rev 27:1–16
Klaminder J, Appleby P, Crook P, Renberg I (2012) Post-deposition diffusion of 137Cs in lake sediment: implications for radiocaesium dating. sedimentology 59:2259–2267
Krammer, K, Lange-Bertalot H (1986) Bacillariophyceae. In: Ettl H, Gerloff J, Heynig H, Mollenhauer D (eds) Süβwasserflora von Mitteleuropa, vol 2/1. Gustav Fisher, Stuttgart/Jena
Krammer K, Lange-Bertalot H (1988) Bacillariophyceae. In: Ettl H, Gerloff J, Heynig H, Mollenhauer D (eds) Süβwasserflora von Mitteleuropa, vol 2/2. Gustav Fisher, Stuttgart/Jena
Krammer K, Lange-Bertalot H (1991a) Bacillariophyceae. In: Ettl H, Gerloff J,Heynig H, Mollenhauer D (eds) Süβwasserflora von Mitteleuropa, vol 2/3. Gustav Fisher, Stuttgart/Jena
Krammer K, Lange-Bertalot H (1991b) Bacillariophyceae. In: Ettl H, Gärtner G, Gerloff J, Heynig H, Mollenhauer D (eds) Süβwasserflora von Mitteleuropa, vol 2/4. Gustav Fisher, Stuttgart/Jena
Lami A, Musazzi S, Marchetto A, Buchaca T, Kernan M, Jeppesen E, Guilizzoni P (2009) sedimentary pigments in 308 alpine lakes and their relation to environmental gradients. Adv Limnol 62:247–268
Laurion I, Vincent WF, Lean DR (1997) Underwater ultraviolet radiation: development of spectral models for northern high latitude lakes. Photochem Photobiol 6:66
Laurion I, Ventura M, Catalan J, Psenner R, Sommaruga R (2000) Attenuation of ultraviolet radiation in mountain lakes: factors controlling the among- and within-lake variability. Limnol Oceanogr 45:1274–1288
Leavitt PR, Cumming BF, Smol JP, Reasoner M, Pienitz R, Hodgson DA (2003) Climatic control of ultraviolet radiation effects on lakes. Limnol Oceanogr 48:2062–2069
Leira M, Filippi ML, Cantonati M (2015) Diatom community response to extreme water-level fluctuations in two Alpine lakes: a core case study. J Paleolimnol 53:289–307
Marchetto A, Mosello R, Rogora M, Manca M, Boggero A, Morabito G, Musazzi S, Tartari GA, Nocentini AM, Pugnetti A, Bettinetti R, Panzani P, Armiraglio M, Cammarano P, Lami A (2004) The chemical and biological response of two remote mountain lakes in the Southern Central Alps (Italy) to twenty years of changing physical and chemical climate. J Limnol 63:77
McGowan S, Anderson NJ, Edwards ME, Langdon PG, Jones VJ, Turner S, van Hardenbroek M, Whiteford E, Wiik E (2016) Long-term perspectives on terrestrial and aquatic carbon cycling from palaeolimnology. Wires Water 3:211–234
McGowan S, Gunn HV, Whiteford EJ, John Anderson N, Jones VJ, Law AC (2018) Functional attributes of epilithic diatoms for palaeoenvironmental interpretations in South-West Greenland lakes. J Paleolimnol 60:273–298
Mcknight DM, Harnish R, Wershaw RL, Baron JS, Schiff S (1997) Chemical characteristics of particulate, colloidal, and dissolved organic material in Loch Vale Watershed, Rocky Mountain National Park. Biogeochemistry 36:99–124
McKnight DM, Boyer EW, Westerhoff PK, Doran PT, Kulbe T, Andersen DT (2001) Spectrofluorometric characterization of dissolved organic matter for indication of precursor organic material and aromaticity. Limnol Oceanogr 46:38–48
Meyer-Jacob C, Michelutti N, Paterson AM, Monteith D, Yang H, Weckström J, Smol JP, Bindler R (2017) Inferring past trends in lake-water organic carbon concentrations in northern lakes using sediment spectroscopy. Environ Sci Technol 51:13248–13255
Meyer-Jacob C, Michelutti N, Paterson AM, Cumming BF, Keller WB, Smol JP (2019) The browning and re-browning of lakes: divergent lake-water organic carbon trends linked to acid deposition and climate change. Sci Rep 9:1–10
Meyers PA (1994) Preservation of elemental and isotopic source identification of sedimentary organic matter. Chem Geol 114:289–302
Meyers PA (2003) Applications of organic geochemistry to paleolimnological reconstructions: a summary of examples from the Laurentian Great Lakes. Org Geochem 34:261–289
Michelutti N, Blais JM, Cumming BF, Paterson AM, Rühland K, Wolfe AP, Smol JP (2010) Do spectrally inferred determinations of chlorophyll a reflect trends in lake trophic status? J Paleolimnol 43:205–217
Miller MP, McKnight DM (2015) Limnology of the Green Lakes Valley: phytoplankton ecology and dissolved organic matter biogeochemistry at a long-term ecological research site. Plant Ecolog Divers 8:689–702
Monteith DT, Stoddard JL, Evans CD, de Wit HA, Forsius M, Høgåsen T, Wilander A, Skjelkvåle BL, Jeffries DS, Vuorenmaa J, Keller B, Kopácek J, Vesely J (2007) Dissolved organic carbon trends resulting from changes in atmospheric deposition chemistry. Nature 450:537–540
Mosello R, Marchette A, Boggero A, Brizzio MC, Tartati GA, Rogora M (1999) Pluriannual evolution of the hydrochemistry of two Alpine lakes (Lake Paione Inferiore and Lake Paione Superiore, Ossola Valley) in relation to atmospheric loads. J Limnol 58:42–48
Nevalainen L, Rantala MV, Kivilä EH, Lami A, Wauthy M, Rautio M, Luoto TP (2020) Biogeochemical and photobiological responses of subarctic lakes to UV radiation. J Photochem Photobiol B 209:111932
Oksanen J, Simpson G, Blanchet F, Kindt R, Legendre P, Minchin P,O'Hara R, Solymos P, Stevens M, Szoecs E, Wagner H, Barbour M, Bedward M, Bolker B, Borcard D, Carvalho G, Chirico M, De Caceres M, Durand S, Evangelista H, FitzJohn R, Friendly M, Furneaux B, Hannigan G, Hill M, Lahti L, McGlinn D, Ouellette M, Ribeiro Cunha E, Smith T, Stier A, Ter Braak C, Weedon J (2022) _vegan: Community Ecology Package_. R package version 2.6-4. https://CRAN.R-project.org/package=vegan
Oleksy IA, Baron JS, Leavitt PR, Spaulding SA (2020) Nutrients and warming interact to force mountain lakes into unprecedented ecological states. Proc R Soc B 287:20200304
Oswald WW, Anderson PM, Brown TA, Brubaker LB, Hu FS, Lozhkin AV, Tinner W, Kaltenrieder P (2005) Effects of sample mass and macrofossil type on radiocarbon dating of arctic and boreal lake sediments. The Holocene 15:758–767
Passy SI (2007) Diatom ecological guilds display distinct and predictable behavior along nutrient and disturbance gradients in running waters. Aquat Bot 86:171–178
Pepin N, Bradley RS, Diaz H, Baraër M, Caceres E, Forsythe N, Fowler H, Greenwood G, Hashmi M, Liu X (2015) Elevation-dependent warming in mountain regions of the world. Nat Clim Change 5:424–430
R Core Team (2022) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. https://www.R-project.org/
Rantala MV, Kivilä EH, Meyer-Jacob C, Atti S, Luoto TP, Smol JP, Nevalainen L (2022) Neoglacial lake-ecosystem changes above and below the subarctic Fennoscandian treeline inferred from changes in diatom functional groups. J Paleolimnol 6:66
Reimer PJ, Austin WEN, Bard E, Bayliss A, Blackwell PG, Bronk Ramsey C, Butzin M, Cheng H, Edwards RL, Friedrich M, Grootes PM, Guilderson TP, Hajdas I, Heaton TJ, Hogg AG, Hughen KA, Kromer B, Manning SW, Muscheler R, Palmer JG, Pearson C, van der Plicht J, Reimer RW, Richards DA, Scott EM, Southon JR, Turney CSM, Wacker L, Adolphi F, Büntgen U, Capano M, Fahrni SM, Fogtmann-Schulz A, Friedrich R, Köhler P, Kudsk S, Miyake F, Olsen J, Reinig F, Sakamoto M, Sookdeo A, Talamo S (2020) The IntCal20 Northern Hemisphere radiocarbon age calibration curve (0–55 cal kBP). Radiocarbon 62:725–757
Renberg I (1990) A procedure for preparing large sets of diatom slides from sediment cores. J Paleolimnol 4:66
Rimet F, Bouchez A (2012) Life-forms, cell-sizes and ecological guilds of diatoms in European rivers. Knowl Manag Aquatic Ecosyst 1:66
Rogora M, Marchetto A, Mosello R (2001) Trends in the chemistry of atmospheric deposition and surface waters in the Lake Maggiore catchment. Hydrol Earth Syst Sci 5:379–390
Rogora M, Mosello R, Arisci S (2003) The effect of climate warming on the hydrochemistry of alpine lakes. Wat Air Soil Pol 148:347–361
Rogora M, Arisci S, Mosello R (2004) Recent trends of temperature and precipitation in alpine and subalpine areas in North Western Italy. Geogr Fis e Din Quat 27:151–158
Rogora M, Colombo L, Lepori F, Marchetto A, Steingruber S, Tornimbeni O (2013) Thirty years of chemical changes in alpine acid-sensitive lakes in the Alps. Water Air Soil Pollut 224:1746
Rogora M, Colombo L, Marchetto A, Mosello R, Steingruber S (2016) Temporal and spatial patterns in the chemistry of wet deposition in Southern Alps. Atmos Environ 146:44–54
Rogora M, Frate L, Carranza ML, Freppaz M, Stanisci A, Bertani I, Bottarin R, Brambilla A, Canullo R, Carbognani M, Cerrato C, Chelli S, Cremonese E, Cutini M, Di Musciano M, Erschbamer B, Godone D, Iocchi M, Isabellon M, Magnani A, Mazzola L, Morra di Cella U, Pauli H, Petey M, Petriccione B, Porro F, Psenner R, Rossetti G, Scotti A, Sommaruga R, Tappeiner U, Theurillat J-P, Tomaselli M, Viglietti D, Viterbi R, Vittoz P, Winkler M, Matteucci G (2018) Assessment of climate change effects on mountain ecosystems through a cross-site analysis in the Alps and Apennines. Sci Total Environ 624:1429–1442
Rubel F, Brugger K, Haslinger K, Auer I (2017) The climate of the European Alps: shift of very high resolution Köppen–Geiger climate zones 1800–2100. Meteorol Z 26:115–125
Rühland K, Paterson AM, Smol JP (2008) Hemispheric-scale patterns of climate-related shifts in planktonic diatoms from North American and European lakes: planktonic diatom responses to recent warming. Glob Change Biol 14:2740–2754
Rühland KM, Paterson AM, Smol JP (2015) Lake diatom responses to warming: reviewing the evidence. J Paleolimnol 54:1–35
Ruwer DT, Rodrigues L (2022) Simplification and replacement of diatom functional traits from a subtropical floodplain lakes paleolimnological records. Limnology 23:153–163
Rydberg J, Cooke CA, Tolu J, Wolfe AP, Vinebrooke RD (2020) An assessment of chlorophyll preservation in lake sediments using multiple analytical techniques applied to the annually laminated lake sediments of Nylandssjön. J Paleolimnol 64:379–388
Sadro S, Sickman JO, Melack JM, Skeen K (2018) Effects of climate variability on snowmelt and implications for organic matter in a high-elevation lake. Water Resour Res 54:4563–4578
Saros JE, Northington RM, Anderson DS, Anderson NJ (2016) A whole-lake experiment confirms a small centric diatom species as an indicator of changing lake thermal structure. Limnol Oceanogr Lett 1:27–35
Saros JE, Anderson NJ, Juggins S, McGowan S, Yde JC, Telling J, Bullard JE, Yallop ML, Heathcote AJ, Burpee BT, Fowler RA, Barry CD, Northington RM, Osburn CL, Pla-Rabes S, Mernild SH, Whiteford EJ, Andrews MG, Kerby JT, Post E (2019) Arctic climate shifts drive rapid ecosystem responses across the West Greenland landscape. Environ Res Lett 14:074027
Schindler DW, Curtis PJ, Parker BR, Stainton MP (1996) Consequences of climate warming and lake acidification for UV-B penetration in North American boreal lakes. Nature 379:705–708
Sivarajah B, Rühland KM, Smol JP (2017) Are diatoms recovering to pre-acidification assemblages in a warming world? Revisiting Killarney Provincial Park lakes (Ontario, Canada). Fundam Appl Limnol 190:13–28
Smol JP (1988) Paleoclimate proxy data from freshwater arctic diatoms. Verh Int Ver Theor Angew Limnol 23:837–844
Smol JP, Stoermer EF (2010) The Diatoms: Applications for the environmental and earth sciences. Cambridge University Press, Cambridge
Smol JP, Brown S, McIntosh H (1984) A hypothetical relationship between differential algal sedimentation and diatom Succession. Verh Int Ver Theor Angew Limnol 22:1361–1365
Soininen J, Jamoneau A, Rosebery J, Passy SI (2016) Global patterns and drivers of species and trait composition in diatoms: global compositional patterns in stream diatoms. Glob Ecol Biogeogr 25:940–950
Sommaruga R (2001) The role of solar UV radiation in the ecology of alpine lakes. J Photochem Photobiol B 62:35–42
Sommaruga R, Psenner R, Schafferer E, Koinig KA, Sommaruga-Wögrath S (1999) Dissolved organic carbon concentration and phytoplankton biomass in high-mountain lakes of the Austrian Alps: potential effect of climatic warming on UV underwater attenuation. Arc Antarct Alp Res 31:247–253
Sorvari S, Korhola A, Thompson R (2002) Lake diatom response to recent Arctic warming in Finnish Lapland: aquatic change caused by arctic warming. Glob Change Biol 8:171–181
Stenger-Kovács C, Lengyel E, Crossetti LO, Üveges V, Padisák J (2013) Diatom ecological guilds as indicators of temporally changing stressors and disturbances in the small Torna-stream, Hungary. Ecol Indic 24:138–147
Tanabe Y, Hori M, Mizuno AN, Osono T, Uchida M, Kudoh S, Yamamuro M (2019) Light quality determines primary production in nutrient-poor small lakes. Sci Rep 9:4639
Tapolczai K, Bouchez A, Stenger-Kovács C, Padisák J, Rimet F (2016) Trait-based ecological classifications for benthic algae: review and perspectives. Hydrobiologia 776:1–17
Van Dam H, Mertens A, Sinkeldam J (1994) A coded checklist and ecological indicator values of freshwater diatoms from the Netherlands. Neth J Aquat Ecol 28:117–133
Vinebrooke RD, Leavitt PR (1999) Differential responses of littoral communities to ultraviolet radiation in an alpine lake. Ecology 80:223–237
Vu VQ (2011) ggbiplot: a ggplot2 based biplot. R package version 0.55, 755
Wauthy M, Rautio M, Christoffersen K, Forsström L, Laurion I, Mariash H, Peura S, Vincent W (2017) Dissolved organic carbon and related environmental data from ponds and lakes in the circumpolar North, v. 1.0 (2002–2016). Nordicana D31:66
Williamson CE, Saros JE, Vincent WF, Smol JP (2009) Lakes and reservoirs as sentinels, integrators, and regulators of climate change. Limnol Oceanogr 54:2273–2282
Wolfe AP, Miller GH, Olsen CA, Forman SL, Doran PT, Holmgren SU (2004) Geochronology of high latitude lake sediments. In: Smol JP, Pienitz R, Douglas MSV (eds) Long-term Environmental Change in Arctic and Antarctic Lakes. Developments in Paleoenvironmental Research, vol 8. Springer, Dordrecht. https://doi.org/10.1007/978-1-4020-2126-8_2
Acknowledgements
This study was funded by the Academy of Finland (Grants #308954, #314107, #335317). For the long-term monitored data, we would like to thank Michela Rogora and the CNR Water Research Institute of Verbania. The data have been collected through the ICP WATERS programme (International Cooperative Programme for assessment and monitoring of the effects of air pollution on rivers and lakes) and the LTER (Long Term Ecological Research Network) network. We thank the personnel at CNR Water Research Institute for assistance during field work and Marina Manca for help with bathymetric maps. We also thank the two anonymous reviewers of their constructive and helpful comments.
Funding
Open Access funding provided by University of Helsinki including Helsinki University Central Hospital.
Author information
Authors and Affiliations
Contributions
M.R., S.A. and L.N. designed the study and acquired the research material (S.A., M.R, A.L). All authors contributed to sample and data analyses relating to diatoms (S.A., M.R., J.W.), sediment chronologies (C.M.-J., J.P.S.), elemental and isotope biogeochemistry (S.A., M.R.) and spectral biogeochemistry (C.M.-J., J.P.S., A.L). S.A. wrote the manuscript which was reviewed by all the other authors.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
The original online version of this article was revised.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Atti, S., Rantala, M.V., Lami, A. et al. Impacts of anthropogenic pressures on underwater light conditions and diatom functional group distributions in mountain lakes. J Paleolimnol 70, 57–76 (2023). https://doi.org/10.1007/s10933-023-00283-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10933-023-00283-y