
Abstract
Compared to larger lakes, ponds have rarely been the focus of palaeoecological studies. A common feature of ponds, especially those subject to eutrophication, is mass surface coverings of lemnoid Araceae (duckweed) which have severe implications for ecological processes in small waterbodies, in particular lowered oxygen content. To help understand the implications of duckweed dominance for the long-term ecology of ponds, and to determine the potential for palaeoecological studies in ponds more generally, we develop a new diatom-based Lemna-indicator metric. Recent studies of diatom host-plant relationships have shown significant associations between duckweed and the epiphytes Lemnicola hungarica and Sellaphora saugerresii (formally known as Sellaphora seminulum). To determine the potential of these species as palaeo-indicators of long-term duckweed dynamics in ponds, we investigated the diatom composition of surface sediment assemblages in sets of duckweed and non-duckweed-dominated ponds in Norfolk, eastern England. In addition, we undertook diatom analysis of two cores from a small farmland pond (Bodham Rail Pit) subject to a known duckweed dominance event (1999–2005). Both L. hungarica and S. saugerresii were significant predictors of past Lemna dominance in the surface sediments. Further, in the core study, both diatom species accurately and closely tracked the documented “on–off” duckweed cycle. Our study suggests huge potential for using ponds in palaeoecological studies and for diatom-based investigations of floating plant histories.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
To date, both ecological and palaeoecological research has focused almost entirely on large, deep and shallow lakes, with minimal attention devoted to the ‘poor cousins’ of shallow lakes, namely ponds. The definition of a pond is currently fluid (Biggs et al. 2005), but in this study, we use it to refer to waterbodies with a maximum diameter of less than 100 m, where insufficient wind-mixing permits chemical stratification of the water-column (Sayer et al. 2013). In terms of palaeoecology, the lack of studies in ponds is particularly acute (Edwards et al. 2011; Emson 2015). While a number of palaeo-studies have been undertaken in arctic ponds (Douglas and Smol 1994; Keatley et al. 2011; Bouchard et al. 2012; Stewart et al. 2013), few if any studies have focused on temperate European lowland ponds—as in this study. Nonetheless, in recent years, a number of studies have shown the predominantly agricultural pond resource of lowland Europe to be of major importance in terms of aquatic biodiversity conservation (Ruggiero et al. 2008; Sayer et al. 2012), aquatic—terrestrial energy subsidies (Davies et al. 2016) and carbon sequestration (Downing et al. 2008), among other roles. Given increasing interest in pond conservation, palaeoecology could have an important role in inferring long-term changes in pond ecology and function in response to environmental impacts.
A key impact, particularly in ponds subject to eutrophication and/or enhanced shading (terrestrialisation), is the development of dense mats of lemnoid Araceae (formally Lemnaceae, and hereafter referred to as duckweed) which typically results in severe water-column anoxia (Lewis and Bender 1961; Pokornŷ and Rejmánková 1983; Janes 1998) in turn causing fish kills (Lewis and Bender 1961), diurnal temperature stratification (Dale and Gillespie 1976), reductions in pH (McLay 1976) and loss of submerged macrophytes and macroinvertebrates (Janse and Van Puijenbroek 1998). The invasion of dense mats of free-floating plants is acknowledged as an important threat to the functioning and biodiversity of freshwater systems (Scheffer et al. 2003), with this especially true of ponds (Sayer et al. 2011). In this study, we test the potential for using sedimentary diatoms to indirectly infer the past presence of duckweed dominance in ponds.
For many diatom species, preferences for particular plant structures are generally poorly understood (Bennion et al. 2010). Indeed, despite several studies, potential species-habitat associations between epiphytic diatoms and different macrophyte species have remained relatively elusive, and it is probable that species-specific plant-diatom relationships are rare (Eminson and Moss 1980). There are some studies which do show strong host-plant links, however. For example Reavie and Smol (1997) revealed a close association between Cocconeis pediculus Ehrenb., and the filamentous algae Cladophora sp. in the St. Lawrence River, Canada. Additionally, Comte and Cazaubon (2002) showed a significant degree of homogeneity of the diatom communities associated with Chara vulgaris L., Groenlandia densa L. and Cladophora glomerata Kütz., in a large Mediterranean river, likely linked to substantial variation in plant surface micro-habitat.
Perhaps the best known example of diatom-habitat specificity is between Lemnicola hungarica (Grunow) Round and P.W. Basson (Synonym: Achnanthes hungarica (Grunow) Grunow in Cleve and Grunow) and free-floating lemnoid (duckweed) plants (Goldsborough 1993; Round and Basson 1997). Grunow first described A. hungarica in 1863 (hereafter referred to as L. hungarica), and it was noted that this species was found in lakes where duckweed was present. Hustedt (1930) commented that L. hungarica had a predisposition towards Lemna spp., and Round (1973) confirmed the observed association between Lemna spp. and L. hungarica. This habitat association was also used by Round and Basson (1997) to justify the establishment of Lemnicola as a new genus distinct from Achnanthes sensu lato. Associations between L. hungarica and free-floating plants were also observed by Bowker and Denny (1980), Goldsborough and Robinson (1985), and Goldsborough (1993, 1994) from field studies of diatom epiphytes sampled from Lemna minor L. Further, Zuberer (1984) identified L. hungarica in Scanning Electron Microscopy (SEM) studies of L. minor, as did Buczkó (2007) from herbarium specimens of various free-floating plants including L. minor, and Desianti (2012) from field collections of diatom epiphytes on free-floating plants. In a host-diatom study of 131 macrophyte samples from 63 sites, covering North and South America, northern Europe, southern Africa, Australia and Asia (Emson 2015), L. hungarica showed a significant association with free-floating plants, as did Sellaphora seminulum (recently named Sellaphora saugerresii (Desm.) C.E. Wetzel et D.G. Mann comb. nov., and hereafter referred to as S. saugerresii—Wetzel et al. 2015). Thus, several studies suggest a strong and robust association between L. hungarica and duckweed, and potentially S. saugerresii also.
Given the poor preservation of Lemna spp. fronds in sediments, and because species in the lemnoid Araceae rarely produce flowers, pollen or seeds (Hillman 1961; Landolt 1986), it is likely infeasible to reconstruct past duckweed dominance directly. Hence, this study will investigate the potential for using diatoms, specifically L. hungarica and S. saugerresii, as indirect indicators of duckweed in ponds. Firstly, we conducted a study of surface sediment diatom assemblages from Lemna-covered ponds and from ponds with no Lemna surface coverage to establish contemporary diatom-duckweed relationships and thereby assess the potential for inferring changes in free-floating macrophyte cover, in small waterbodies. Secondly, we tested the ability of diatoms to track a documented duckweed “on–off” cycle in a small pond (Bodham Rail Pit) through diatom analysis of a sediment core.
Study sites
The surface sediment study focuses on a set of small ponds in Norfolk, eastern England. For the most part, the ponds are situated in arable agricultural land and have their origins as ‘marl pits’ being formed by marl extraction largely in the eighteenth–nineteenth centuries (Prince 1964). Other ponds have their origins as pingos formed at the end of the Last Glaciation 12.7–11.5 cal kyr BP (Flemel 1976). The ponds were all small (< 40 m max. diameter), shallow (generally < 1 m) and, in the case of the farmland ponds, largely alkaline and eutrophic (Table 1). Further details on the history, geography and ecology of ponds in the study area are given in Sayer et al. (2012).
Some 12 Lemna-dominated ponds were selected for study based on local knowledge of duckweed occurrence. In these ponds L. minor and/or Lemna minuta Kunth., was known to have been prevalent over at least 2–3 years at between 25 and 100% of water-surface cover. At the time of sampling (2012) 1, 3 and 8 sites were dominated by L. minor, L. minuta and co-dominated by L. minor and L. minuta respectively. For comparison 14 non-Lemna ponds were selected at random from a comparable study area. In all cases, it was known that these sites had not experienced duckweed domination in the recent (about the last 5 years) past.
The focus of the palaeoecological investigation was the Bodham Rail Pit (52° 54′20. 62′′N; 1°09′21. 23′′E), hereafter referred to as Rail Pit, a typical small (350 m2), shallow (mean depth 103 cm) pond located in Bodham, North Norfolk (Fig. 1). The Rail Pit is located in arable farmland and surrounded by a 10 m rough grassland buffer zone. The pond is primarily ground-water fed but also receives surface runoff via a small road. Water levels in the pond fluctuate seasonally by about 1 m but it never completely dries out. The age of the pond is unknown, but it is shown on a tithe map dating to 1841, suggesting that it is at least 150–200 years old. Importantly, the pond underwent a major duckweed dominance event over 1999–2005, before and after which it is known to have been largely duckweed free. Over the period 1994–1998, prior to high duckweed coverage, the Rail Pit had abundant stands of Potamogeton natans L., but more recently (2008-present) it has been dominated by Ceratophyllum submersum L (Fig. 2).

Bodham Rail Pit, Norfolk, eastern England (a), and site map showing distribution of contemporary aquatic, emergent and riparian vegetation (b). Bodham Rail Pit showing Lemna dominance in 2003 (c) and Lemna absence in 2010 (d). Core RAIL1 (e). Note the dark organic silt above lighter grey marl at the base of the core (Photographs: CD Sayer)

Recent history of aquatic macrophytes recorded from the Bodham Rail Pit. Macrophyte abundances are presented using the DAFOR scale (D Dominant-5, A Abundant-4, F Frequent-3, O Occasional-2, R Rare-1). Note that Lemna minor was generally co-dominant with Lemna minuta
Materials and methods
Historical plant records: Rail Pit
A recent history of aquatic macrophyte abundance for the Rail Pit is available from 1994 to 2016 (Sayer and Emson, unpublished data). In each year, plants were recorded on the DAFOR scale (Dominant—5, Abundant—4, Frequent—3, Occasional—2, Rare—1) based on visual observations assisted by plant collections made with a double–headed rake.
Surface sediment and core sampling
Surface sediments (upper 0–1 cm) were collected from the Lemna-covered (n = 12) and non-Lemna ponds (n = 14) variously using a Glew gravity corer (Glew 1991) and an improvised ladle attached to a long metal rod (“Pond Putter”).
A 118-cm sediment core (RAIL1) was collected from the Rail Pit using a ‘Big Ben’ wide-bore piston corer (Patmore et al. 2014) from a southern central location at a depth of 108 cm (Fig. 1). The core was sliced on site at 1-cm intervals. On extrusion the core length was reduced to 75 cm, likely due to compaction of the very soft upper sediments which were very flocculent. Given the fluid upper section of RAIL1, and as the key aim was to track recent duckweed coverage, a short (22 cm) Glew core was also collected from an adjacent location. This core (RAIL2) was sliced at 0.5-cm intervals throughout.
All sediment samples were placed in sealed plastic bags and subsequently stored at 4 °C prior to analysis.
Radiometric analysis
Dried sediment samples from cores RAIL1 and RAIL2 were analysed for 210Pb, 226Ra, 137Cs and 241Am by direct gamma assay at the Bloomsbury Environmental Isotope Facility (BEIF), University College London, using an ORTEC HPGe GWL series well-type coaxial low background intrinsic germanium detector. 210Pb was determined via its gamma emissions at 46.5 keV, and 226Ra using the 295 keV and 352 keV gamma rays emitted by its daughter isotope 214Pb following three weeks storage in sealed containers to allow radioactive equilibrium. 137Cs and 241Am were measured by their emissions at 662 keV and 59.5 keV respectively (Appleby et al. 1986). The absolute efficiencies of the detector were determined using calibrated sources and sediment samples of known activity. Corrections were made for the effect of self-absorption of low energy gamma rays within each sample (Appleby et al. 1992).
Chronologies and sediment accumulation rates for both of cores RAIL1 and RAIL2 were calculated using the constant rate of 210Pb supply (CRS) model (Appleby and Oldfield 1978). Use of the constant initial 210Pb concentration (CIC) model was precluded because of irregular declines in unsupported 210Pb activities resulting in non-monotonic features in the 210Pb profiles of both cores (Fig. 3).

Chronologies of cores RAIL1 (a) and RAIL2 (b) showing CRS model-inferred 210Pb dates and sedimentation rates. Solid lines show chronological age with depth, and dashed lines show sedimentation rates
Diatom analysis
A total of 48 sediment samples were analysed for diatoms from core RAIL1. The uppermost 10–20 cm of the core was analysed at contiguous 1-cm intervals and below this samples were analysed at 2-cm intervals. A total of 44 contiguous samples were analysed for diatoms in core RAIL2 at 0.5-cm intervals covering the full length of the 22-cm core. Samples were prepared for diatom analysis using standard methods (Battarbee 1986; Battarbee et al. 2001). Given that eutrophication can potentially swamp rare species by abundance increases of meso-eutraphentic and hyper-eutraphentic species (Van Dam and Mertens 1993), and as the aim was to determine how well the diatom stratigraphy tracked past duckweed dominance using two indicator species, large diatom counts (range 500–4700 valves per sample) were undertaken to reduce the dominance of common diatom species, whilst allowing responses of rarer taxa to be more accurately captured. All samples were mounted on microscope slides using Naphrax™ and absolute numbers of diatoms present in 0.1 g of sediment were counted at 1000× magnification with oil immersion under phase-contrast illumination using a Leitz ‘Laborlux S’ light microscope. Diatoms were identified to species level and diatom data for both cores were presented as percentage relative abundances. Images of the two indicator species used in the study are given in Fig. 4.

Light microscopic images of the two Lemna-indicator diatom species: Lemnicola hungarica (a–h) and Sellaphora saugerresii (i–m). Lemnicola hungarica rapheless valves (a–d) and raphe valves (e–h). Scale bars = 10 μm (Photographs: P Rioual)
Data manipulation and analysis
Binomial logistic regression was performed using SPSS version 22 (2013) to determine whether L. hungarica and S. saugerresii could faithfully predict Lemna dominance in the ponds.
Pearson’s correlation analysis was applied to L. hungarica and S. saugerresii relative abundances to determine the strength of their bivariate relationship in both of cores RAIL1 and RAIL2. Diatom core stratigraphic data were plotted using C2 software (Juggins 2007).
Results
Recent macrophyte history
The Rail Pit macrophyte data show a cyclical shift between submerged macrophytes and lemnid dominance (Fig. 2). The submerged C. submersum and P. natans were abundant in the early 1990s during years of low Lemna abundance (1994–1998), but rapidly declined and disappeared in the late 1990s (1999–2005) with the onset of a Lemna-dominance phase (typically > 90% surface cover). This Lemna phase ended relatively abruptly in 2005 leaving the Rail Pit resembling a ‘bacterial soup’ (C.D. Sayer pers. observ.) until 2008 when C. submersum reappeared as abundant (DAFOR = 4 in 2008) together with P. crispus (DAFOR = 3 in 2009). During this second phase of submerged plant dominance, P. natans was absent, Lemna trisulca L. was recorded for the first time as a rare species (DAFOR = 1), and Cladophora sp. became more prevalent (DAFOR = 3). At the time of core collection (April 2010) the only macrophytes present were C. submersum (DAFOR = 4), L. minor (DAFOR = 1) and L. minuta (DAFOR = 1) the latter both found in marginal areas (Fig. 2).
Surface sediments
L. hungarica and S. saugerresii were significantly more abundant in the Lemna sites (Fig. 5). Both L. hungarica (max. = 54%, min. = 5%, mean = 16%) and S. saugerresii (max. = 8%, min. = 1%, mean = 3%) were recorded from the surface sediments of all Lemna-dominated sites, with the exception of both L. hungarica and S. saugerresii in Priory Pond 1 and S. saugerresii alone in Old Church Barn Pond 1 (Table 1). Only one non-Lemna site, Sayer’s Black Pit (Site 21), contained L. hungarica in its surface sediments but at a very low percentage abundance (0.003%). There were three non-Lemna sites namely, Pond Farm Pond 2, Sayer’s Black Pit and Otom Pit (Sites 15, 21 and 26 respectively) which recorded S. saugerresii, again at very low percentage abundances (0.002, 0.01 and 0.006% respectively). Logistic regression analysis indicated that duckweed-covered sites successfully predicted the presence of both L. hungarica (p = 0.0001, r2 = 0.903) and S. saugerresii (p = 0.002, r2 = 0.758) confirming their potential as indicator species. The predictive model was equally accurate for both the duckweed (93% correct) and non-duckweed sites (92% correct).

Percentage relative abundances of Lemnicola hungarica (a) and Sellaphora saugerresii (b) in surface sediment samples from Lemna-covered (n = 12) and non-Lemna covered (n = 14) ponds. 25–75 percent quartiles (excluding outliers) are shown using a box, median is the horizontal line, whiskers represent the highest/lowest data points, and outliers are shown as stars
Radiometric dating of cores RAIL1 and RAIL2
The simple CRS model places the 1963 maximum nuclear weapons fallout layer (137Cs peak) at 23.5 cm, slightly above the 137Cs peak (26.5 cm) in core RAIL1. There is a sudden and dramatic increase in sedimentation rate at 34.5 cm (1940s) to 0.47 g cm−2 year−1 which may be due to a sediment slumping event (Fig. 3a). This is followed by a relatively uniform period covering the 1950s to the 1970s of 0.04–0.07 g cm−2 year−1 and an increased relatively uniform sedimentation rate of 0.08–0.1 g cm−2 year−1 for the last 30 years. For core RAIL2 the CRS dating model places 1963 at 12.5 cm, just above that suggested by the 137Cs and 241Am records at 15.5 cm. This may be due to non-continuous sedimentation before 1963. The CRS calculated chronology indicates that sediment accumulation rates gradually increased from 0.02 g cm−2 year−1 in the 1940s to 0.06 g cm−2 year−1 since the 1980s (Fig. 3b).
RAIL1 and RAIL2 diatom stratigraphies and correlation
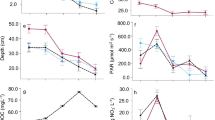
There was high temporal concordance between relative abundance peaks of L. hungarica and S. saugerresii and the observed Lemna-covered period for 1999–2005 in both the RAIL1 (7–3 cm; 1999–2005) and RAIL2 (6–3 cm; 1999–2005) cores (Phase 4 in Fig. 6). Similarly, peaks of both diatoms occurred for the 1950s (Phase 3 in Fig. 6), with this better defined in RAIL1 (32–27 cm) with increases in L. hungarica (2.5%) and S. saugerresii (3.5%), and increases of L. hungarica (11%) and S. saugerresii (7%) for RAIL2 (19–17 cm).

Percentage abundance stratigraphies of Lemnicola hungarica, Sellaphora saugerresii and both species combined, for cores RAIL1 (a) and RAIL2 (b). Grey shading denotes the 1999–2005 observed Lemna dominance period
A further Lemna-dominance phase was evident in core RAIL1 with L. hungarica and S. saugerresii peaks over 48–40 cm (Phase 2 in Fig. 6a). Finally, a potential earlier Lemna phase was evidenced by increases in S. saugerresii (2–4%) and to a lesser extent L. hungarica (< 1%) at around 68 cm (Phase 1 in Fig. 6a). Despite some scatter and a lesser degree of agreement, especially in the lower sections of RAIL1, a positive linear relationship was found between percentages of L. hungarica and S. saugerresii in both of cores RAIL1 (r = 0.57, p < 0.01, Fig. 7a) and RAIL2 (r = 0.72, p < 0.01, Fig. 7b).

Relationships between Lemnicola hungarica and Sellaphora saugerresii percentage relative abundances for cores RAIL1 (a) and RAIL2 (b)
Discussion
The first formal documented record of a diatom was in 1703 (thought to be Tabellaria flocculosa) sampled from the roots of Lemna (Round et al. 1990), but on Christmas day in 1702 Van Leeuwenhoek looked at the roots of Lemna and also probably saw diatoms, although he described his findings as ‘animalcula’ in the Philosophical Transactions of the Royal Society (Van Leeuwenhoek 1703). Since these early days, many workers have investigated potential host-macrophyte relationships for diatoms (Prowse 1959; Siver 1977; Eminson and Moss 1980; Millie and Lowe 1983). Round (1973), Bowker and Denny (1980), Goldsborough and Robinson (1985) and Goldsborough (1993, 1994) were amongst the first researchers to acknowledge the association between L. minor and L. hungarica. Following these pioneering studies other workers have investigated the consistency and robustness of diatom-duckweed relationships. Buczkó (2007) found L. hungarica dominated diatom assemblages attached to L. minor, Lemna gibba L., Spirodela polyrhiza L. Schleid., and Wolffia arhiza L. Horkel ex Wimm., in marked contrast to assemblages found on L. trisulca which were dominated by Cocconeis placentula Ehrenb. Buczkó (2007) concluded that L. hungarica was tightly attached to well definable taxa of the lemnoid Araceae. Desianti (2012) also reported L. hungarica for L. minor, S. polyrhiza and Wolffia spp.
Clearly, several studies have concluded that L. hungarica has a strong habitat preference for duckweed, while Emson (2015) showed a similar association for S. saugerresii (Synonym: Sellaphora seminulum (Grunow) D.G. Mann). This good understanding of diatom ecology begs the question: is it possible to employ these diatoms as palaeo-indicators of past Lemna dominance? To this end, our study clearly shows both L. hungarica and S. saugerresii to be successfully transferred from duckweed mats to the sediment strongly suggesting a positive answer to this question. Further, this study shows that, even at low percentages (< 5%), these diatoms can indicate high duckweed coverage (Fig. 6). Importantly, this indicator status for L. hungarica and S. saugerresii applies not only to the lemnoid Araceae (with the exception of L. trisulca) but can also be applied to other free-floating plants such as Azolla spp. and Riccia spp. (Emson 2015).
Confirmation of a strong Lemna-diatom association comes from comparing the RAIL1 and RAIL2 diatom stratigraphies with historical observations in the Rail Pit. In both cores the documented arrival and termination of high Lemna coverage over 1999–2005 is clearly tracked by peaks of both L. hungarica and S. saugerresii (Phase 4 in Fig. 6). Thus, even despite highly flocculated upper sediments for RAIL1 after collection, the Lemna signal remained intact. Core RAIL1, which extended to the beginning of sedimentation in the Rail Pit, showed some degree of bloom versus low-abundance (boom-bust) cyclicity for both L. hungarica and S. saugerresii, with three further potential Lemna-covered phases (Fig. 6a). For the penultimate peak (Phase 3 in Fig. 6) reasonable temporal accordance was evident between cores RAIL1 (32–27 cm = 1950–1960) and RAIL2 (19–17 cm = 1948–1958) giving faith in the reconstruction. The two earlier inferred duckweed peaks in RAIL1 (Phase 2—48–40 cm; Phase 1—68–60 cm) were less well defined for S. saugerresii compared to L. hungarica, however, potentially suggesting that L. hungarica is the most reliable Lemna indicator. On this basis low percentages of L. hungarica for Phase 1 make an inference of duckweed-dominance more questionable. Indeed, despite a significant positive correlation between L. hungarica and S. saugerresii in both cores, differences in their responses in RAIL1 suggest that these species may have slightly different ecological preferences and further studies in this respect could be fruitful. Given this, the merit of combining L. hungarica and S. saugerresii to provide a Lemna-indicator metric (Fig. 6) is open to debate.
The ability to infer periods of free-floating plant dominance through time is of particular relevance to small waterbodies where duckweed proliferation, often symptomatic of high-nutrient loading, results in extensive surface coverings of duckweed (Portielje and Roijackers 1995). These dense mats physically prevent oxygen from being dissolved from the atmosphere and increase carbon dioxide levels (Janes 1998) causing a reduction in pH (McLay 1976; Janes 1998) and increased conductivity (Greaves et al. unpublished data). In addition, lemnid dominance can attenuate photosynthetic active radiation (PAR) by up to 99% with associated temperature fluctuations which can lead to diurnal temperature stratification (Dale and Gillespie 1976; Goldsborough 1993). Dense duckweed mats can cause significant species losses in submerged macrophyte and invertebrate communities (Janse and Van Puijenbroek 1998), and associated cold, anoxic conditions can cause fish kills (Lewis and Bender 1961). Thus, a diatom-Lemna inference tool has much utility in terms of reconstructing the past ecology of ponds, with the same potentially true of shallow lakes and indeed larger lakes under certain circumstances across the globe (Duarte and Kalff 1990) where high Lemna-coverage occurs. For example, dominance of free-floating mats of lemnids have also occurred in Lake Maracaibo, Venezuela and Lake Titicaca, Peru due to increased nutrient inputs (Cruz et al. 2006), and seasonal blooms of duckweeds have also been observed in cut-off meanders in large rivers (Beauger et al. 2015). Significant relationships between L. hungarica and duckweed have been demonstrated for a variety of climatic conditions and waterbody types in many parts of the world including Canada (Goldsborough 1993, 1994), USA (Zuberer 1984; Desianti 2012), Brazil (Garcia and Fonseca de Souza 2006) and substantial tracts of Europe (Buczkó 2007; Emson 2015). Consequently, with the caveat that more work on habitat preferences is needed for S. saugerresii, compared to L. hungarica, we strongly suggest that both diatoms might be reliably used to infer past free-floating plant dominance much more widely. Will it be “mission possible” in other waterbodies in the world? Certainly, we urge other workers to go out and test this idea.
Conclusions
This study suggests that ponds in lowland European agricultural landscapes have considerable palaeolimnological potential. Indeed, the close agreement of diatom-Lemna signals in two cores from the Rail Pit strongly suggest that palaeo-techniques can be confidently applied to these often over-looked waterbodies, at least where ponds are permanent.
The study also shows the huge advantage of incorporating ‘rare’ species in diatom analysis and of undertaking large (> 500 valves per slide) counts to better characterize diatom signals in cores. In this case the two Lemna-indicator diatoms occurred at relatively low percentages (< 5%) in both stratigraphic sequences and in the surface sediments, even where duckweed was entirely dominant. This is particularly pertinent given that conventional diatom studies, based on counts of 300 or at best 500 diatom valves, tend to delete species at < 5% relative abundance from summary plots. Thus, as a ‘rule of thumb’, the duckweed-covered signal afforded by L. hungarica and S. saugerresii would typically be missed.
Finally, in developing two diatom-indicators of free-floating plants, our research highlights the huge value of understanding diatom species ecologies for interpreting palaeo-data. In this respect, similar to previous studies (Bennion et al. 2010; Sayer et al. 2010), we suggest that a greater understanding of diatom responses to habitat and chemical variation across and between different littoral zone structures in lakes and ponds would greatly improve the interpretation of sediment records dominated by non-planktonic diatoms.
References
Appleby PG, Oldfield F (1978) The calculation of 210Pb dates assuming a constant rate of supply of unsupported 210Pb to the sediment. Catena 5:1–8
Appleby PG, Nolan PJ, Gifford DW, Godfrey MJ, Oldfield F, Anderson NJ, Battarbee RW (1986) 210Pb dating by low background gamma counting. Hydrobiologia 141:21–27
Appleby PG, Richardson N, Nolan PJ (1992) Self-absorption corrections for well-type germanium detectors. Nucl Instrum Meth B 71:228–233
Battarbee RW (1986) Diatom analysis. In: Berglund BE (ed) Handbook of holocene palaeoecology and palaeohydrology. Wiley, New York, pp 527–570
Battarbee RW, Jones VJ, Flower RJ et al (2001) Diatoms. In: Smol JP, Birks HJB, Last WM (eds) Tracking environmental change using lake sediments, volume 3: terrestrial, algal and siliceous indicators. Kluwer, Dordrecht
Beauger A, Delcoigne A, Voldoire O, Serieyssol K, Peiry J (2015) Distribution of diatom, macrophyte and benthic macroinvertebrate communities related to spatial and environmental characteristics: the example of a cut-off meander of the River Allier (France). Cryptogam Algol 36(3):323–355
Bennion H, Sayer CD, Tibby J, Carrick H (2010) Chapter 8: Diatoms as indicators of environmental change in shallow lakes. In: Smol JP, Stoermer EF (eds) The diatoms: applications for the environmental and earth sciences. Cambridge University Press, Cambridge, pp 152–173
Biggs J, Williams P, Whitfield M, Nicolet P, Weatherby A (2005) 15 years of pond assessment in Britain: results and lessons learned from the work of Pond Conservation. Aquat Conserv 15:693–714
Bouchard F, Pienitz R, Ortiz JD, Francus P, Laurion I (2012) Palaeolimnological conditions inferred from fossil diatom assemblages and derivative spectral properties of sediments in thermokarst ponds of subarctic Quebec, Canada. Boreas 42:575–595
Bowker DW, Denny P (1980) The seasonal succession and distribution of epiphytic algae in the phyllosphere of Lemna minor L. Arch Hydrobiol 90:39–55
Buczkó K (2007) The occurrence of the epiphytic diatom Lemnicola hungarica on different European Lemnaceae species. Fottea Olomouc 7(1):77–84
Comte K, Cazaubon A (2002) Structural variations of epiphytic diatom communities on three macrophytes in a regulated river (Durance), in South-East of France. Ann Limnol 38(4):297–305
Cruz MA, Terrazas EM, Northcote T (2006) Worsening water quality conditions at Inner Puno Bay, Lake Titicaca, Peru, and their effect on Lemna spp. biomass. Freshwater Forum 26:46–57
Dale HM, Gillespie TJ (1976) The influence of floating vascular plants on the diurnal fluctuations of temperature near the water surface in early spring. Hydrobiologia 49:245–256
Davies SR, Sayer CD, Greaves H, Siriwardena GM, Axmacher JC (2016) A new role for pond management in farmland bird conservation. Agr Ecosyst Environ 233:179–191
Desianti N (2012) Interactions between duckweeds and their diatom epiphytes. MSC thesis, University of Oklahoma
Douglas MSV, Smol JP (1994) Limnology of high arctic ponds (Cape Herschel, Ellesmere Island, N.W.T.). Arch Hydrobiol 131:401–434
Downing JA, Cole JJ, Middelburg JJ, Striegl RG, Duarte CM, Kortelainen P, Prairie YT, Laube KS (2008) Sediment organic carbon burial in agriculturally eutrophic impoundments over the last century. Glob Biogeochem Cy 22:GB1018
Duarte CM, Kalff J (1990) Patterns in the submerged macrophyte biomass of lakes and the importance of scale of analysis in the interpretation. Can J Fish Aquat Sci 47(2):357–363
Edwards KJ, Schofield JE, Kirby JR, Cook GT (2011) Problematic but promising ponds? Palaeoenvironmental evidence from the Norse Eastern Settlement of Greenland. J Quat Sci 26(8):854–865
Eminson D, Moss B (1980) The composition and ecology of periphyton communities in freshwaters I. The influence of host type and external environment on community composition. Br Phycol J 15:429–446
Emson D (2015) The ecology and palaeoecology of diatom-duckweed relationships. Unpublished Ph.D. thesis, University College London and Botany Department, NHM, London
Flemel RC (1976) Pingos and Pingo Scars: their characteristics, distribution and utility in reconstructing former permafrost environments. Quat Res 6(1):37–53
Garcia M, Fonseca de Souza V (2006) Lemnicola hungarica (Grunow) Round and Basson from southern Brazil: ultra-structure, plastid morphology and ecology. Diatom Res 21(2):465–471
Glew JR (1991) Miniature gravity corer for recovering short sediment cores. J Paleolimnol 5:285–287
Goldsborough LG (1993) Diatom ecology in the phyllosphere of the common duckweed (Lemna minor L.). Hydrobiologia 269(270):463–471
Goldsborough LG (1994) Heterogeneous spatial distribution of periphytic diatoms on vertical substrata. J N Am Benthol Soc 13(2):223–236
Goldsborough LG, Robinson GGC (1985) Seasonal successional of diatom epiphyton on dense mats of Lemna minor. Can J Bot 63:2332–2339
Hillman WS (1961) The Lemnaceae or duckweeds: a review of the descriptive and experimental literature. Bot Rev 27:221–318
Hustedt F (1930) Bacillariophyta (Diatomeaea) Heft. 10. Die Süsswasserflora von Mitteleuropas. G. Fischer, Jena. pp 466
Ibm, SPSS Statistics for Windows. Version 22.0 (2013) Armonk. IBM Corp, NY
Janes R (1998) Growth and survival of Azolla filiculoides in Britain. I. Vegetative reproduction. New Phytol 138:367–375
Janse JH, Van Puijenbroek PJTM (1998) Effects of eutrophication in drainage ditches. Environ Pollut 102:547–552
Juggins S (2007) C2 Version 1.5: Software for ecological and palaeoecological data analysis and visualization (program). University of Newcastle, Newcastle upon Tyne
Keatley BE, Blais JM, Douglas MSV, Gregory-Eaves I, Mallory ML, Mitchelutti N, Smol JP (2011) Historical seabird population dynamics and their effects on Arctic pond ecosystems: a multi-proxy paleolimnological study from Cape Vera, Devon Island, Arctic Canada. Fund Appl Limnol 179:51–66
Landolt E (1986) Biosystematic Investigations in the Family of Duckweeds (Lemnaceae) Vol. 2. The Family of Lemnaceae—a Monographic Study, Vol. 1. Veroff Geobot Inst ETH, Zurich
Lewis WM, Bender M (1961) Effect of a cover of duckweeds and the alga Pithopora on the dissolved oxygen and free carbon dioxide on small ponds. Ecology 42:602–603
McLay CL (1976) The effect of pH on the population growth of three species of duckweed: Spirodela oligorrhiza, Lemna minor and Wolffia arrhiza. Freshwater Biol 6:125–136
Millie DF, Lowe RL (1983) Studies on Lake Erie’s littoral algae: host specificity and temporal periodicity of epiphytic diatoms. Hydrobiologia 99:7–18
Patmore IR, Sayer CD, Goldsmith B, Davidson TA, Rawcliffe R, Salgado J (2014) Big Ben: a new wide-bore piston corer for multi-proxy palaeolimnology. J Paleolimnol 51:79–86
Pokornŷ J, Rejmánková E (1983) Oxygen regime in a fishpond with duckweeds (Lemnaceae) and Ceratophyllum. Aquat Bot 17:125–137
Portielje R, Roijackers RMM (1995) Primary succession of aquatic macrophytes in experimental ditches in relation to nutrient input. Aquat Bot 50:127–140
Prince HC (1964) The origin of pits and depressions in Norfolk. Geography 49:15–32
Prowse GA (1959) Relationship between epiphytic algal species and their macrophyte hosts. Nature 183:1204–1205
Reavie ED, Smol JP (1997) Diatom-based model to infer past littoral habitat characteristics in the St. Lawrence River. J Great Lakes Res 23:339–348
Round FE (1973) The biology of algae. Edward Arnold, London
Round FE, Basson PW (1997) A new monoraphid diatom genus (Pogoneis) from Bahrain and the transfer of previously described species A. hungarica and A. taeniata to new genera. Diatom Res 12:71–81
Round FE, Crawford RM, Mann DG (1990) The diatoms. Cambridge University Press, NY
Ruggiero A, Céréghino R, Figuerola J, Marty P, Angélbert S (2008) Farm ponds make a contribution to the biodiversity of aquatic insects in a French agricultural landscape. C.R. Biologies 331:298–308
Sayer CD, Davidson TA, Jones JI, Langdon PG (2010) Combining contemporary ecology and palaeolimnology to understand shallow lake ecosystem change. Freshwater Biol 55:487–499
Sayer CD, Copp GH, Emson D, Godard MJ, Zięba G, Wesley KJ (2011) Towards the conservation of Crucian Carp Carassius carassius: understanding the extent and causes of decline within part of its native English range. J Fish Biol 79:1608–1624
Sayer CD, Andrews K, Shilland E, Edmonds N, Edmonds-Brown V, Patmore IR, Emson D, Axmacher J (2012) The role of pond management for biodiversity conservation in an agricultural landscape. Aquat Conserv 22:626–638
Sayer CD, Shilland E, Greaves H, Dawson B, Patmore IR, Emson D, Alderton E, Robinson P, Andrews K, Axmacher J, Wiik E (2013) Managing British ponds-conservation lessons from a Norfolk farm. British Wildlife 25(1):21–28
Scheffer M, Szabó S, Gragnani A, van Nes EH, Rinaldi S, Kautsky N, Norberg J, Roijackers RMM, Franken RJM (2003) Floating plant dominance as a stable state. Proc Natl Acad Sci USA 100(7):4040–4045
Siver PA (1977) Comparison of attached diatom communities on natural and artificial substrates. J Phycol 13:402–406
Stewart EM, Michelutti N, Blais JM, Mallory ML, Douglas MSV, Smol JP (2013) Contrasting the effects of climatic, nutrient, and oxygen dynamics on subfossil chironomids assemblages: a paleolimnological experiment from eutrophic High Arctic ponds. J Paleolimnol 49:205–219
Van Dam H, Mertens A (1993) Diatoms on herbarium macrophytes as indicators for water quality. Hydrobiologia 269(270):437–445
Van Leeuwenhoek A (1703) Part of a letter from Mr. Antony van Leeuwenhoek, F.R.S. concerning green weeds growing in water, and some animalcula found about them. Philos T R Soc Lond 23:1304–1311
Wetzel CE, Ector L, Van De Vijver B, Compère P, Mann DG (2015) Morphology, typification and critical analysis of some ecologically important small naviculoid species (Bacillariophyta). Fottea Olomouc 15(2):203–234
Zuberer DA (1984) Microbial colonization of some duckweeds (Lemnaceae): examination by scanning and transmission electron and light microscopy. Aquat Bot 18:275–285
Acknowledgements
The authors would like to thank the erstwhile landowner Pat Cubitt, and current landowner Peter Seaman, for making the Bodham Rail Pit available for study and many other landowners/land managers who allowed us to collect surface sediment samples from their ponds. A really big thank you to members of the ECRC, Maria Asuncion Ramos Cruz and Kevin Ruddy for kindly providing macrophyte samples with a smile! Our sincere thanks go to Jorge Salgado and Gizem Bezirci for expertise in coring and air-guitars! Grateful thanks to Handong Yang and Neil Rose for radiometric analysis and to Miles Irvine for drafting the figures. We are also grateful to the two reviewers for helpful and constructive comments and to Tom Davidson for advice and encouragement which substantially improved this article. Our heartfelt gratitude goes to June and Derek Sayer for accommodation, patience and everything else.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Emson, D., Sayer, C.D., Bennion, H. et al. Mission possible: diatoms can be used to infer past duckweed (lemnoid Araceae) dominance in ponds. J Paleolimnol 60, 209–221 (2018). https://doi.org/10.1007/s10933-017-0008-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10933-017-0008-6