
Abstract
Animal domestication is a profound change for human societies, economies, and worldviews. The shifting definitions of animal domestication reflect its varying and process-like nature. Reindeer is one of the species whose domestication is not easily pinned down using standard definitions and research methodologies of animal domestication. In recent years, advances in archaeological methodology and the conceptual understanding of animal domestication have opened new avenues for research on this topic. This review summarizes recent research on the archaeology of reindeer domestication among the Indigenous Sámi of northern Fennoscandia. It compiles a chronological framework of reindeer domestication with an emphasis on the development of reindeer-herding practices and human–reindeer relationships. I argue that while a major transition to reindeer herding occurred among the Sámi from the 15th century onward, small-scale reindeer herding characterized by interspecies sociality, cooperation, and care developed earlier during the Late Iron Age, with regional variations in the timing and details of the events. By focusing on reindeer-herding practices and the human–reindeer relationships embedded in them, I also argue that reindeer domestication, and animal domestication in general, is a relationship constructed and constantly renegotiated in everyday interactions with the animals.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Reindeer herding is an emblematic way of life for much of northern Eurasia, where reindeer-herding peoples are encountered from Mongolia in the east through the Siberian tundra and taiga zones to the northern parts of Finland, Sweden, and Norway. The domestication of the reindeer and the connections between the different reindeer-herding traditions have been discussed by historians, archaeologists, and ethnographers for more than 100 years (Kortesalmi 2008, pp. 20–21; Stépanoff 2017; Storli 1996). One reason for the constant fascination with the topic is due, in part, to the fact that the relationships between people and reindeer escape easy labels and straightforward domestication narratives.
Animal domestication is one of the major shifts in human prehistory, transforming economies, societies, and worldviews (e.g., Cauvin 2000; Hodder 2011; Vigne 2011). According to the classic paradigm, livestock was first domesticated in the Near East for primary products such as meat, hide, and bone, whereas secondary products such as milk, wool, and traction were only later exploited on a larger scale (Greenfield 2010; Sherratt 1981). Some have argued that livestock domestication and the adoption of agriculture led to the development of permanent settlement, population growth, the rise of social inequality, and religious change (e.g., Bocquet-Appel 2002; Cauvin 2000; Hodder 2011; Horwitz et al. 1999; Kohler et al. 2017). It has long been believed that the relationship with animals and nature changed from trust and reciprocity to domination and control during the domestication process (Ingold 2000), and classic definitions of animal domestication tend to emphasize human control over the lives and reproductive cycles of animals (e.g., Bökönyi 1989; Clutton-Brock 1992, p. 21). However, as the domestication histories of reindeer and many other species show, animal domestication processes are complex and unique processes with varying motives, starting points, and outcomes (e.g., Bogaard et al. 2021; Zeder 2012). Although the variation in animal domestication processes has long been acknowledged by archaeologists, the classic narratives and definitions can haunt the methodological and conceptual approaches archaeologists take to study animal domestication.
Probably the most comprehensive definition of animal domestication, by Zeder (2015, p. 3191), defines it as “a sustained multigenerational, mutualistic relationship in which one organism assumes a significant degree of influence over the reproduction and care of another organism in order to secure a more predictable supply of a resource of interest, and through which the partner organism gains advantage over individuals that remain outside this relationship, thereby benefitting and often increasing the fitness of both the domesticator and the target domesticate.” Traditional archaeological approaches to domestication have focused largely on aspects of reproductive control by searching for morphological characteristics associated with the domestication syndrome, which is a set of morphological, physiological, and behavioral features linked to domestication, probably via changes in neural crest development (Wilkins et al. 2014). These morphological characteristics such as craniofacial changes and reduction in body size (e.g., Arbuckle and Makarewicz 2009; Clutton-Brock 1992, pp. 21–25; Germonpré et al. 2009) are not necessarily evolved in incipient domestication (Harbers et al. 2020; Pelletier et al. 2020). Yet human-induced changes in locomotor patterns, for example, corralling and draught use, induce plastic changes such as changes in body size, morphology, and cross-sectional properties that can be indicative of early-stage domestication (Harbers et al. 2020; Pelletier et al. 2020).
Domestication is a social and cultural relationship between people and animals (Anderson et al. 2017a; Clark 2007; Ingold 2000; Knight 2012; O’Connor 1997; Oma 2010). In particular, the many forms that human–reindeer entanglements can take challenge traditional understandings of animal domestication, leading scientists to find new vocabularies to describe these entanglements in terms of mutual learning and sharing (e.g., Anderson et al. 2017a; Losey et al. 2021; Stépanoff 2017). Stépanoff (2017) argues that reindeer domestication is a process in which both humans and animals modify their behavior to learn to live together in a shared landscape. Losey et al. (2021) present a related argument that emphasizes the mutual learning aspects in reindeer domestication, claiming that reindeer domestication was a process of enskillment where both people and reindeer learned new skills when engaging with each other. Anderson et al. (2017a) reject clear boundaries between wild and domesticated reindeer and focus on understanding their various relationships with people through architectures and places of encounter in the landscape. Common to these approaches and definitions is that reindeer domestication is understood as an ongoing process in which the relationships between the people and animals are constructed in shared environments and daily interactions (Salmi et al. 2021).
The new vocabularies of domestication are accompanied by new archaeological methodologies and approaches that enable the teasing out of new nuances of human–reindeer relationships from the archaeological data (e.g., Anderson et al. 2019; Heino et al. 2021; Pelletier et al. 2020; Salmi et al. 2020a, b; Seitsonen and Viljanmaa 2021). Promising new possibilities are provided by methods that allow the identification of human–reindeer interactions such as feeding, milking, penning, working together, or shared mobilities that occur in the daily framework of animal management practices (e.g., Jerand and Linderholm 2019; Pelletier et al. 2020; Salmi et al. 2021; Seitsonen and Viljanmaa 2021). Although the emphasis on the social aspects of the domestication relationship may be more descriptive of animal management than domestication as such (Zeder 2015), the addition of human–animal sociality to the definition of the domestication relationship has the potential to add to the archaeological methodological repertoire and the understanding of the shared lives of humans and domesticated animals in the past.
In this paper, I address reindeer domestication and the development of reindeer herding among the Indigenous Sámi of northern Fennoscandia in the light of new studies, methodological possibilities, and conceptual tools that have emerged in recent years. I start by outlining current knowledge on the ecological and genetic aspects of reindeer domestication and the archaeological approaches taken to study it. I then examine four different types of human–animal social engagements more closely that occurred in the process of reindeer domestication: reindeer supplementary feeding, the training and use of draught reindeer, the shared landscapes of mobilities of people and reindeer, and their religious and ritual entanglements. I approach these aspects of the human–reindeer relationship by utilizing information provided by multiple methodologies, the most important being stable isotope analysis, an analysis of ancient DNA, physical activity reconstruction based on skeletal remains, landscape archaeology, and the use of reindeer herders’ traditional knowledge. Through an emphasis on identifying and understanding reindeer-herding practices, I explore the human–reindeer relationships embedded in the domestication relationship. Utilizing the archaeology of these social engagements, I demonstrate that approaches targeting human–animal social interaction can contribute especially to the understanding of reindeer-herding practices, but also to dating and understanding the processes involved in incipient reindeer domestication. I also argue that the study of such human–animal social engagements has the potential to inform us about human–animal relationships in the domestication processes of other animal species. This paper therefore contributes to the understanding of reindeer domestication in the wider context of animal domestication studies.
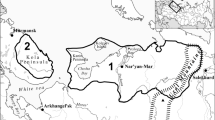
The geographical focus of this paper is the homeland of the Indigenous Sámi people, located in northern Fennoscandia (Norway, Sweden, Finland, and the Kola Peninsula in northwest Russia). There are differences in archaeological research activity and the resulting representativeness of data for northern Fennoscandia (e.g., Seitsonen and Viljanmaa 2021). Furthermore, the Sámi affiliations of many types of archaeological sites located in southern Norway and Sweden and on the coastal areas are still debated, whereas the affiliations of most sites located in northern areas are often less controversial (Amundsen and Os 2020; Baglo 2019; Zachrisson 1997). In practice, many of the analyses covered in this paper of stable isotopes, ancient DNA, and faunal analysis have focused on sites located in the northern parts of the Sámi area (Fig. 1, Table 1). Thus, the primary archaeological data covered in this paper originate from northern Finland, Sweden, and Norway (Fig. 1), with very little data from the Kola Peninsula and Southern Sámi areas.

Map of northern Fennoscandia with archaeological sites mentioned in the text. Filled square Settlement site filled circle Offering place filled triangle Mining site filled triangle Marketplace (illustration by A. Salmi)
The archaeological data I discuss consist of different data sets from Sámi dwelling sites (Salmi et al. 2021; Seitsonen and Viljanmaa 2021), marketplaces (Fjellström et al. 2020), and ritual offering places (Núñez et al. 2020; Salmi et al. 2018, 2020a). The main data sets used to discuss the central themes of this paper are listed in Table 1, including faunal assemblages from dwelling sites located in Norway, Sweden, and Finland, and dating from ca. AD 1000 to the 18th century. There also are faunal data from marketplaces, which have a long history in northern Fennoscandia, at least from the 12th century onward (Kuusela et al. 2016; Wallerström 1983; Ylimaunu 2007, pp. 24–25). The northernmost inland marketplaces were established in the 16th century, when the King of Sweden ruled that a church and a marketplace were to be established in every Lapp village (Korpijaakko 1989, pp. 139–145; Slunga 1993). Marketplaces served various social, economic, legal, and religious purposes for the Sámi (Korpijaakko 1989, pp. 139–145; Ylimaunu 2007, pp. 26–28). There also are archaeological data from Silbojokk, representing the living facilities of the workers of the Nasafjäll silver mine. Many of the workers were Sámi, and the Sámi communities of the surrounding areas played an important role in providing for the mining community (Roslund 1989; Sten 1989). A large set of archaeological data originates from Sámi offering places located in Norway, Finland, and Sweden. Offering places were natural features such as boulders, waterbodies, or mountains, usually unshaped by people, and they served in negotiating the relationships between people, the land, and its resources (Salmi et al. 2018). Various goods such as metal objects, animal body parts, tobacco, and alcohol were offered at these places, beginning with animals in ca. AD 500–700 (Salmi et al. 2018). In addition to these data sets (see Table 1), spatial data regarding various Sámi sites dating from ca. AD 700 onward have been utilized to analyze hunting and herding landscapes through GIS analysis (Seitsonen and Viljanmaa 2021). The timeline of the data thus extends approximately from the eighth to the 19th century AD. The data sets are patchy spatially and temporally, the most comprehensive data set in terms of spatial and temporal representativeness perhaps being that from Sámi offering places (Heino et al. 2021; Salmi et al. 2018, 2020a; Table 1). Despite the patchiness, the data sets offer insights into both more general and local processes of reindeer domestication and the development of reindeer herding among the Sámi. Yet the limitations the patchiness pose, especially to understanding the development of reindeer herding outside the core study area, must be borne in mind.
Background
Ecological, Morphological, and Genetic Aspects of the Reindeer and Its Domestication
The reindeer (or caribou) (Rangifer tarandus) is a pan-arctic ungulate species extant in the northern parts of Eurasia and North America. The morphology and ecology of the reindeer vary across the area. Different subspecies or ecotype divisions have been made, but they can generally be divided into high arctic, tundra, and forest ecotypes. Ecological and ethological characteristics such as mobility, diet, and herd structure vary between populations and ecotypes (Banfield 1961). These characteristics also vary seasonally. Two subspecies, wild forest reindeer (R. t. fennicus) and mountain reindeer (R. t. tarandus), live in Fennoscandia. Wild forest reindeer herds are relatively small, and they do not exhibit large-scale migratory behavior between seasonal habitats as mountain reindeer do. Mountain reindeer live in larger herds, although the herd composition varies seasonally, and they can migrate over large distances between seasonal habitats. Today, mountain reindeer include small wild populations in southern Norway and the Kola Peninsula in Russia, and the domestic populations in Sweden, Norway, and northern Finland. Wild forest reindeer are extant today in central and eastern Finland and in northwestern Russia (Banfield 1961; Weldenegodguad et al. 2020).
Both subspecies are thought to have been present in Fennoscandia since deglaciation (Rankama and Ukkonen 2001). Genetic differences between the Fennoscandian reindeer populations indicate that mountain reindeer colonized the area from the south after deglaciation, whereas wild forest reindeer colonized the area from the east (Røed et al. 2008; Weldenegodguad et al. 2020). It is likely that the distribution of the subspecies has fluctuated according to climatic and environmental changes over time (Rankama and Ukkonen 2001). Furthermore, the reindeer subspecies can interbreed with one another (e.g., Røed et al. 2014). Genetic research conducted mainly on mitochondrial DNA indicates that reindeer were domesticated separately in Fennoscandia and Siberia. Mitochondrial genetic lineages common in today’s domestic reindeer herds in Fennoscandia (haplotypes II and Ib) are present but less prevalent in Siberia, implying two separate domestication centers (Røed et al. 2008; Weldenegodguad et al. 2020). These genetic lineages first appear in the archaeological record in Fennoscandia ca. AD 1400–1600 (Bjørnstad et al. 2012; Heino et al. 2021; Røed et al. 2018), but they were present in Siberia as early as ~3900 BC (Røed et al. 2020).
Although the morphological and behavioral characteristics of wild and domestic reindeer are not as distinct as in many other species, several morphological and behavioral changes have occurred in the process of reindeer domestication. The skeletal morphology of the reindeer varies according to subspecies, sex, age, population, and various environmental factors (e.g., Puputti and Niskanen 2009; Weinstock 1997; Weladji and Holand 2006). Recent studies have shown that although the morphologies and body proportions of the subspecies extensively overlap (Nieminen and Helle 1980), there are several differences in skeletal morphology according to not only subspecies and sex, but also human influence in the domestication process (Pelletier et al. 2020, 2021a, b). The postcranial skeletal elements of wild forest reindeer are generally significantly larger than those of mountain reindeer and have more robust bony epiphyses. Moreover, due to the sexual dimorphism of the reindeer, males have significantly more massive skeletal elements, with larger epiphyses than females, which tend to have a slenderer skeletal morphology (Pelletier et al. 2020, 2021a, b). Furthermore, castration affects the size and proportions of those long bones that fuse after the age at which the individual is castrated (van den Berg et al. in press). Reduced mobility due to captivity also leads to decreased body size without directed selection (Pelletier et al. 2020; 2021a), a phenomenon also observed in other domestic species (e.g., Zohary et al. 1998). Although reindeer have never been kept in complete captivity, apart from zoo-kept individuals, practices such as part-time penning and other human-influenced changes in reindeer mobility patterns may cause skeletal morphology changes. There also are other functional changes such as working that affect skeletal morphology: Bone cross-sectional properties differ between working and non-working reindeer; and working reindeer also tend to be larger and more robust than non-working reindeer, although the latter is a product of the selection of suitable individuals for training, not domestication-related selection pressure (Niinimäki et al. 2021; Pelletier et al. 2020, 2021b).
Previous Research on Reindeer Domestication in Fennoscandia
Reindeer have been hunted by various human groups since the Paleolithic period, with hunting practices varying with local knowledge, reindeer behavior, and the local landscape (Helskog 2011a). In Fennoscandia, wild reindeer were hunted from the Mesolithic until the early 19th century as part of varying subsistence patterns (Halinen 2005, pp. 82–94; Helskog 2011b; Lundmark 1982, p. 161; Rankama and Ukkonen 2001; Virrankoski 1973, pp. 271–272). For example, pitfall systems for catching wild reindeer are known throughout prehistory in northern Fennoscandia, with peaks in the intensity of their use in the Iron Age and early medieval period (Hennius 2020; Jordhøy 2008). Although rock art dating from the Mesolithic depicts enclosures, reindeer tied to a rope, and people riding reindeer, it is more likely to depict hunting techniques, rituals, and mythology than incipient domestication (Helskog 2011b, 2012).
Domesticated reindeer have been herded in multiple ways by many cultural groups across northern Eurasia. Today, reindeer-herding peoples exist in the Russian Far East, Mongolia, Siberia, and northern Fennoscandia (Helskog 2011a). In northern Fennoscandia, reindeer herding is practiced by the Indigenous Sámi of Finland, Sweden, Norway, and northwestern Russia. In Finland, reindeer herding is also practiced by ethnic Finns. Studied for more than 100 years by archaeologists, historians, and ethnographers, reindeer domestication and the adoption of reindeer herding in northern Eurasia is one of the central questions in the archaeology of the area (Kortesalmi 2008, pp. 20–21; Stépanoff 2017; Storli 1996). Historical studies of the development of reindeer herding have mostly focused on the post-16th century period (e.g., Hultblad 1968; Kortesalmi 2008; Lundmark 1982; Tegengren 1952). The analysis of changes in settlement patterns and their social implications for the development or herding societies (e.g., Aronsson 1991; Bergman et al. 2013; Halinen et al. 2013; Hedman et al. 2015; Seitsonen and Viljanmaa 2021) have been especially prominent avenues of archaeological research on the matter in the Fennoscandian research tradition. Many studies of reindeer domestication also typically combine ethnographic, historical, and archaeological sources (e.g., Bjørklund 2013; Wallerström 2000). Although these approaches have produced a general chronology of events, many questions about the early stages of reindeer domestication and reindeer-herding practices remain unresolved.
The first piece of historical evidence to describe domesticated reindeer is usually considered to be Ohthere’s account to King Alfred the Great in England in AD 890, which mentions among Ohthere’s possessions tame deer and decoy or draught reindeer (Bately 2007, p. 46). In addition to this piece of historical evidence, archaeological analysis of habitation patterns, in particular the emergence of so-called stállo sites in the Scandinavian mountains, have been used to argue that incipient domestication was underway between AD 700 and AD 1000 (Bergman et al. 2013; Seitsonen and Viljanmaa 2021). In addition, the so-called hearth row sites (also rectangular hearth sites), dating to ca. AD 600/700–1300 and situated in inland northern Fennoscandia, are considered by most researchers to be cold-season settlements of reindeer herders, although the evidence is ambiguous and may also point to the importance of hunting and fishing (Bergman et al. 2013; Halinen et al. 2013; Hedman et al. 2015; Olsen 2019; Seitsonen and Viljanmaa 2021). Researchers have also argued that the roots of the family-based siida organization of Sámi society can be traced back to this period (Bergman et al. 2013; Halinen et al. 2013; Hedman et al. 2015).
Multiple sources of evidence indicate that a clear shift to larger-scale reindeer herding occurred between AD 1400 and AD 1600. Settlement pattern changes indicate mobile pastoralism (Bergman et al. 2013; Seitsonen and Viljanmaa 2021). At the same time, analysis of ancient mtDNA from sites across northern Fennoscandia similarly implies a replacement of other haplotypes by haplotypes common in modern domesticated reindeer (Bjørnstad et al. 2012; Heino et al. 2021; Røed et al. 2018). Furthermore, it seems that the family-based siida system of Sámi society was established at this time (Bergman et al. 2013).
The analysis of faunal assemblages has produced inconclusive or mixed results. Reindeer age profiles from Sámi dwelling sites dating to ca. AD 1000–1650 indicate that mainly adult individuals were slaughtered, with some differences between sites (Hambleton and Rowley-Conwy 1997; Harlin et al. 2019; Hedman et al. 2015; Vretemark 2019). This corresponds to the practices employed in later traditional reindeer herding, in which mostly adult individuals were slaughtered (e.g., Itkonen 1948, p. 259; Korhonen 2008, p. 137). However, the employment of selective hunting strategies such as stalking adult males with the aid of a female reindeer (Itkonen 1948, p. 18) cannot be ruled out as an explanation for the adult-dominated age profiles. An analysis of skeletal frequencies shows that all parts of the reindeer carcass were present at the dwelling sites (Hambleton and Rowley-Conwy 1997; Harlin et al. 2019; Hedman et al. 2015; Vretemark 2019). Even representation of skeletal elements may indicate that the animals were slaughtered close to or on the site, which may in turn mean that the animals originated in domestic herds instead of wild animals hunted and processed at a distance from the dwelling site (Hedman et al. 2015; Indrelid and Hufthammer 2011). It is also likely that many Sámi communities practiced a mixed subsistence of small-scale herding, hunting, and fishing (e.g., Harlin et al. 2019; Vretemark 2019), in which case the reindeer bone assemblages would represent a mix of domesticated and wild reindeer.
The early herders’ reindeer-herding practices remain relatively unclear until historical sources start shedding light on the matter from the 16th century onward. In A Description of the Northern Peoples, Magnus (1996 [1555]) describes reindeer milking and draught reindeer use, and a mobile pastoralism “like that practiced by sheepherders elsewhere.” From the 17th century, the number of historical sources on reindeer herding increases. The Crown of Sweden started to undertake reindeer counts for taxation needs from the early 17th century. These counts show a geographical variation in reindeer-herding practices: Large reindeer herds of more than 100 individuals were common in the mountain areas, but in the forest and lake areas of present-day Finland, reindeer herders were few, and the herds were small at ten reindeer or fewer (Hultblad 1968; Kortesalmi 2008, pp. 22–23). There is also increasing if patchy information about reindeer-herding practices. The 17th-to-18th century sources mention reindeer milking, draught and cargo reindeer use, care-intensive herding, and mobile pastoralism, as well as the different adaptations of the mountain and forest Sámi (Leem 1956 [1767]; von Linné 1969 [1732]; Rheen 1897 [1671]; Schefferus 1979 [1674]; Tornaeus 1900 [1672]).
Historical sources show a wide variation in the reindeer herding and overall subsistence practices of the Sámi. Historical and archaeological sources imply that the Sámi population in eastern Fennoscandia relied on a mixed subsistence strategy of hunting, fishing, gathering, and small-scale reindeer herding, whereas in the mountain areas, large-scale mobile pastoralism was dominant (e.g., Bergman et al. 2013; Hansen and Olsen 2014, p. 231; Harlin et al. 2019; Hultblad 1968; Lundmark 1982). Other regional variations include lake fishing, important in the interior (Hansen and Olsen 2014, pp. 188–191; Itkonen 1948, pp. 536–540), marine hunting, and fishing coupled with small-scale agriculture on the northern coast of Norway (Hansen and Olsen 2014, p. 174), and a combination of farming and fishing in the more southern coastal areas (Hansen and Olsen 2014, pp. 186–188). Sheep or goat husbandry was incorporated into the livelihood of the Sámi in Norway possibly in the Viking period, more certainly in the early medieval period, and cattle and sheep husbandry were part of the livelihood of the Sámi in fertile river valleys from the 18th century (Hansen and Olsen 2014, pp. 177, 190–200).
Modern transformations in reindeer-herding practices include state border closures in the late 19th century, which altered the traditional seasonal mobility patterns of many Sámi communities (e.g., Heikkinen et al. 2012). In the 20th century, Fennoscandian reindeer herding was characterized by the free-ranging system, in which the reindeer are corralled only twice a year (Helle and Jaakkola 2008). Herding practices shifted from the multipurpose herd toward meat production (Holand 2007). In addition, the decrease in availability of lichen pastures and arboreal lichens due to border closures and forestry practices led to growing dependence on supplementary winter feeding during the 20th century (Helle and Jaakkola 2008). Today, reindeer herding is under pressure from other competing land use (forestry, mining, energy production, nature protection, predator conservation, and tourism), an increasing number of predators, rising costs, a decreasing and aging workforce, and the effects of climate change on the annual cycle of reindeer herding; technological changes such as snowmobiles and GPS also have changed herding practices (Sarkki et al. 2018). The colonial legacy of nation-states remains in the reindeer-herding area, as land and resource use continue to be contested between the Sámi and the respective states (e.g., Brännström 2020).
Animal Feeding in Early Reindeer Herding
Animal feeding is a type of human–animal interaction that often characterizes the domestication relationship, although it also occurs between people and undomesticated species. It is even possible that additional food provided by the human association is the driving force of the domestication of species such as the dog, which probably entered the domestication relationship through the commensal pathway (Zeder 2012). Animal feeding practices in domestication processes and in animal husbandry practices have been studied archaeologically through stable isotope analyses indicative of trophic level and other changes in the animals’ diets (e.g., Gillis et al. 2013; Pickard et al. 2017). Human influence in the diets and foraging behavior of domesticated animals is not restricted to the addition of specific foodstuffs to animals’ diets but can also be understood in the broader, landscape-level context of human influence over animal foraging patterns in pastoral systems, for example (Hörnberg et al. 2018; Miller and Makarewicz 2019).
In natural conditions, reindeer consume a seasonally variable diet consisting of more than 200 plant species, as well as mushrooms and lichens. In the summer, they consume mostly green vegetation, whereas lichen is their main food in the winter, making up about two-thirds of the vegetable mass consumed (Bezard et al. 2015; Nieminen and Heiskari 1989). Domesticated reindeer have probably never been kept in full captivity necessitating a full-time feeding regime. Instead, people have affected their foraging patterns by guiding them to fresh pastures (Helle and Jaakkola 2008), maintaining lichen pastures with the aid of recurrent fires (Hörnberg et al. 2018), and giving them supplementary food in times of need and for taming specific individuals (Itkonen 1948, pp. 83–84; Korhonen 2008, p. 42; Soppela et al. 2020). In early 20th-century reindeer herding, reindeer were given supplementary fodder consisting of dried lichens, branches, grasses, and sedges (Itkonen 1948, pp. 83–84; Korhonen 2008, p. 42). Supplementary feeding was practiced especially during times when the reindeer had difficulty finding food themselves, when a thick crust formed on the snow and made digging for lichen under the snow difficult (Itkonen 1948, pp. 83–84; Korhonen 2008, p. 42). In addition, feeding may have served other purposes related to the relationships between herders and reindeer; today’s reindeer herders use feeding to establish a trusting relationship with reindeer, help with taming and training of reindeer, keep the reindeer in good condition, and monitor their well-being (Salmi et al. 2022).
New archaeological evidence of reindeer feeding has been obtained through stable isotope analysis and an analysis of feeding behavior-related physical activity patterns (Fjellström et al. 2020; Núñez et al. 2020; Salmi et al. 2020a; Salmi and Niinimäki 2021). Lichen is rich in carbohydrates but poor in protein (Nieminen and Heiskari 1989; Soppela et al. 2008), and the nitrogen (δ15N) values of lichens are lower than those of leafy trees, grasses, and sedges in subarctic and arctic environments (e.g., Beck and Mayr 2012; Gustine et al. 2012; Liu et al. 2018; Michelsen et al. 1998; Skrzypek et al. 2015). This is reflected in the stable isotope values of reindeer bone tissue, which is expected to be relatively low in nitrogen and high in carbon in reindeer feeding mostly on lichen (Britton 2009; Evans 2007, p. 85; Fjellström et al. 2020). Based on the differences in the nitrogen and carbon values in the different foods the reindeer consume, these values may reflect differences in reindeer diet and possibly, human influences on it (Fjellström et al. 2020; Salmi et al. 2020a; Takken Beijersbergen et al. 2021). In particular, it has been argued that especially relatively high values of nitrogen (δ15N) reflect foddering in the reindeer, as the replacement of low-nitrogen lichens in the winter diet with grasses, shrubs, and dried tree branches provided by humans would affect the stable isotope composition of the winter diet (Fjellström et al. 2020; Salmi et al. 2020a). However, other factors that affect nitrogen values in reindeer bone, antler, and teeth, such as feeding in different environments (e.g., boreal forest vs. tundra), starvation, seasonal variations in antler shedding between the sexes, a young age (before weaning at ca. 6–7 months of age) (e.g., Barboza and Parker 2006; Parker et al. 2005; Takken Beijersbergen et al. 2021), and their effects must be considered when analyzing the stable isotope values of reindeer tissue samples.
Stable isotope values have been analyzed for reindeer bone samples from Sámi dwelling sites, offering places, and marketplaces dating from the 11th to the 20th centuries (Fjellström et al. 2020; Núñez et al. 2020; Salmi et al. 2020a) (Fig. 2). The stable isotope values show considerable variation between and within sites (Fjellström et al. 2020; Núñez et al. 2020; Salmi et al. 2020a). Some of the nitrogen values clearly fell in the range typical of the bone tissue of adult, non-fed reindeer in the area, ca. 1–3.5 ‰, while others were significantly elevated, between ca. 3.5–6 ‰ (Fjellström et al. 2020; Núñez et al. 2020; Salmi et al. 2020a). The nitrogen values seem not only to reflect the environmental setting, because elevated nitrogen values were observed on sites located in both alpine and boreal vegetation zones (Fig. 2A). Moreover, no major temporal trends are observable, because samples with elevated nitrogen values date to the entire studied period (Fig. 2B). The samples were taken from individuals assessed as adults or past weaning age, which excludes the possible effects of lactation on elevated nitrogen values. It is therefore possible that the diets of at least some of the sampled individuals with elevated nitrogen values were influenced by people in a similar manner to those documented in ethnographic sources. The earliest samples with elevated nitrogen values were date to between the 11th and 13th centuries and originated in the offering places of Unna Saiva and Udtjäure in Sweden (Salmi et al. 2020a), as well as the dwelling site of Vivallen (Fjellström et al. 2020).

Stable isotope values of nitrogen (δ15N) and carbon (δ13C) in reindeer bone samples from Sámi sites. Data from Fjellström et al. (2020) and Salmi et al. (2020a). Site dating is used as dates for samples from the Vivallen dwelling site, Arjeplog marketplace, and Silbojokk mining community (Fjellström et al. 2020). Because offering places are often multi-period sites, dates are presented only for those samples from offering places that were directly radiocarbon dated (Salmi et al. 2020a)
A small data set of physical activity-related skeletal changes backs the conclusion that some of the reindeer offered specifically at Unna Saiva received supplementary winter fodder (Salmi and Niinimäki 2021). Feeding behavior, specifically digging for lichen under the snow in the winter with the aid of the repetitive movement of the elbow joint, affects muscle attachment sites in that joint to the degree that fed reindeer are distinguishable from non-fed reindeer in terms of entheseal site scores (Niinimäki and Salmi 2016). Reindeer offered at Unna Saiva exhibited skeletal markers indicative of a low degree of lichen-digging behavior, probably due to supplementary winter feeding (Salmi and Niinimäki 2021).
Regarding ritual practices, reindeer with human-altered diets were offered at roughly the same time and on the same sites with reindeer individuals with no human-influenced changes in their diet (Salmi et al. 2020a). The variability of the carbon and nitrogen values throughout the studied period and across sites implies that domesticated reindeer were probably offered along with wild reindeer (Núñez et al. 2020; Salmi et al. 2020a). Reindeer supplementary feeding seems to have been part of early reindeer-herding practice, but it was not universal among Sámi reindeer herders. Rather, it was probably practiced locally and at times of need. It is also possible that the fed animals were specific individuals, for example, those intended for draught reindeer training and kept in closer human contact than other reindeer. It is also likely that feeding was part of the establishing and maintaining of trust and closeness between herders and reindeer, and that the contacts related to reindeer feeding were occasions for the herders to monitor the animals’ well-being.
Working with Reindeer
Traction, along with wool and milk, is one of the secondary products of animal domestication. The role of secondary products, as opposed to primary products such as meat and hides, in animal domestication has been debated since Sherratt (1981) coined the term “secondary product revolution,” and it seems evident that the role of primary versus secondary products has varied between domestication processes and regions where domesticates were integrated into human economies (Greenfield 2010; Marciniak 2011). The use of animals for traction in the past can be approached via the analysis of working-related skeletal changes, as well as material culture (Greenfield 2010).
Reindeer have worked together with people for millennia, and the central role of working reindeer in reindeer domestication and early reindeer herding of the Sámi has been hypothesized (e.g., Bately 2007, p. 46; Bjørklund 2013; Ingold 1986). Material culture related to draught use is often ambiguous, because the same harnessing and traction technology may have been used for several species (e.g., reindeer, humans, or dogs). Sled runners dating from the Mesolithic onward have been discovered in northern Fennoscandia (Kuokkanen 2000). Sled runners dating to 1500 BC and resembling those in later Sámi sleds have been found in northwest Russia (Murashkin et al. 2016). Harness parts dating to between ca. 200 BC and AD 160 and resembling those the Nenets use for training draught reindeer have been found on the Yamal Peninsula (Losey et al. 2021). They occur at the same time as the increase in portable imagery depicting reindeer (Nomokonova et al. 2021). In northern Fennoscandia, sleds similar to later Sámi reindeer sleds have been discovered in graves dating from the 10th century onward (Svestad 2018). Undated reindeer harness parts from medieval and early modern Sámi dwelling sites are evidence of the use of draught reindeer (Carpelan 1991, 1993), as are 16th century and later historical sources describing the use of draught and cargo reindeer (e.g., Magnus 1996 [1555], Rheen 1897 [1671]; Schefferus 1979 [1674]; Tornaeus 1900 [1672]).
New methodological possibilities of analyzing the early use of working reindeer have emerged through the analysis of working-related skeletal changes such as pathological lesions and entheseal changes in the reindeer skeleton (Salmi et al. 2020b). These skeletal markers reflect the habitual physical activities and extra loading associated with working the reindeer and form clear patterns of skeletal changes that are different from non-working reindeer (Salmi et al. 2020b). Working reindeer typically have joint diseases such as new bone growth, articular surface erosion and lipping, and joint fusion, particularly in the vertebrae, phalanges, and tarsal bones, that form in response to the extra stress and loading caused by working. Although joint disease also can occur in aging animals in the wild, the combined analysis of frequency, severity, and skeletal distribution of the changes can be indicative of working (Salmi et al. 2020b).
An analysis of these skeletal markers in reindeer remains from three Sámi dwelling sites in northern Finland dating to the 14th to 18th centuries indicated that working animals were already integrated into the economy of the Sámi residing in northern Finland by the 14th century (Salmi et al. 2021). An analysis of paleopathological lesions in reindeer bones in the faunal assemblages from three sites, Juikenttä, Nukkumajoki, and Autiokenttä, revealed new bone growth, articular surface lipping, and bone fusion, particularly in the phalanges, vertebrae, tarsal bones, and humerus (Salmi et al. 2021). Based on location of the lesions in the same skeletal elements as in modern working reindeer, working is the probable cause for the observed pattern (Fig. 3). Moreover, the severity of the phalangeal pathologies in the archaeological assemblages resembled that of modern working reindeer, further supporting the hypothesis that the pathologies were related to working (Salmi et al. 2021). The pattern was especially clear for Juikenttä and Nukkumajoki, with larger faunal assemblages, but pathological specimens were also discovered in Autionkenttä (Salmi et al. 2021).

Relative abundances of skeletal elements of all the skeletal elements with pathological lesions in archaeological assemblages from Juikenttä, Nukkumajoki, and Autiokenttä. Data from Salmi et al. 2021 (illustration by A. Salmi)
Osteometric analysis of the reindeer bones from these three assemblages indicates that both subspecies of reindeer, R. t. tarandus and R. t. fennicus, were present. Although osteometric analysis is not very reliable at identifying individual skeletal elements from reindeer subspecies due to the extensive overlap in size (Puputti and Niskanen 2009), the overall size range of the archaeological specimens compared with modern specimens implied the presence of both subspecies (Salmi et al. 2021). The pathologies were mostly observed in skeletal elements in the middle or upper part of the size range, primarily reflecting the use of large, castrated males for traction (Salmi et al. 2021). The oldest samples with working-related pathologies date to the late 13th and 14th centuries, while most of them date between AD 1400 and 1700 (Salmi et al. 2021).
The analysis of working-related skeletal lesions in the assemblages in association with radiocarbon dates, reindeer bone osteometric measurements, and species diversities of the assemblages showed that wild reindeer hunting and other hunting activities were probably practiced by the Sámi residing in these sites throughout the period of occupation (Salmi et al. 2021). Small-scale reindeer herding, including working reindeer, was integrated into mixed subsistence patterns that also included hunting, fowling, and fishing (Harlin et al. 2019; Salmi et al. 2021). The results confirm the early roots of the forest Sámi subsistence pattern of small-scale reindeer herding combined with hunting, fishing, and gathering (Fjellström 1986; Hansen and Olsen 2014, pp. 192–95; Hultblad 1968; Kortesalmi 2008, p. 26; Paulaharju 1922). The archaeological faunal assemblages analyzed thus far do not go back far enough in time to precisely define the role of draught reindeer in incipient reindeer domestication among the Sámi. However, it is possible that the subsistence adaptation observed at the studied sites from the 14th century onward resembles that of the early reindeer herders, because early reindeer herders employed a similar set of mixed subsistence strategies as the Sámi in the studied sites (e.g., Halinen et al. 2013; Hedman et al. 2015; Vretemark 2019).
Working relationships between people and reindeer are indicative of domestication, but they are also embedded in interpersonal interspecies learning and cooperation processes and have important implications for human–reindeer relationships. In the first half of the 20th century, the Sámi used reindeer to pull and carry loads (Itkonen 1948, pp. 388–412; Näkkäläjärvi and Pennanen 2000). Draught and cargo reindeer were used to transport people and their possessions during seasonal migrations and reindeer-herding tasks (Itkonen 1948, pp. 388–412; Korhonen 2008, p. 58). Today, reindeer herders train reindeer for tourism and competitive racing (Soppela et al. 2020). Working reindeer are usually castrated males, which are selected for training and castrated at the age of three to four years (Itkonen 1948, pp. 419–422). The training, taking place over several seasons, is a two-way interspecies communication and learning process, in which reindeer personality and personhood play crucial parts (Soppela et al. 2020; Vuojala-Magga 2010). Itkonen (1948, pp. 422), an ethnographer, reported that some reindeer were faster to learn than others, and that the personality and teaching style of the trainer affected both how the reindeer learned and how good a transportation reindeer it became. Reindeer are selected for draught training based on their appearance and physical abilities, but above all for their personality (Soppela et al. 2020). The consideration of traditional knowledge of reindeer herders affords new opportunities to reflect on the nature of human–reindeer relationships in past working reindeer training and use. The interspecies learning and communication experiences likely included similar practices, perceptions, and sensory experiences to those of today, although their exact forms have changed over time according to the specifics of the working and training practices and frames of thinking about non-human animals and their personhood.
Reindeer-Herding Landscapes and Architectures
The daily chores of the reindeer herders are always performed in specific landscapes. These landscapes and their architectural and natural features, as well as the tasks performed in them, form the taskscapes and landscapes of reindeer herding. A taskscape consists of interrelated tasks, everyday activities that construct the lived-in worlds of human and non-human actors and their temporalities (Ingold 1993, p. 158, 1997, pp. 29–30; Mazzullo and Ingold 2008). Similarly, domestication has also been described as a process whereby both humans and animals modify their behavior to learn to live together in a shared landscape (Stépanoff 2017). Domestication relationship is played out and constructed using structures and architectures such as tethers and enclosures (Anderson et al. 2017a).
The landscapes of early reindeer herding are indicative of the ways reindeer herding was incorporated into the daily lives of the Sámi. Sámi reindeer-herding practices have varied widely temporally and spatially, but as a coarse generalization, the mobility patterns of the Sámi of the historical period have been divided into forest and mountain Sámi patterns (e.g., Fjellström 1986). The mountain Sámi migrated between different ecological zones seasonally, the main migrations being between the taiga in the winter and mountains in the summer. On the other hand, the forest Sámi mobility pattern consisted of hunting, fishing, and herding activities at different locations in the landscape (e.g., Fjellström 1986, p. 149; Itkonen 1948, p. 93; Rheen 1897 [1671]). Recent GIS analyses focusing on the landscapes and taskscapes of the early reindeer herders indicate that many Sámi communities practiced small-scale reindeer herding in a mixed hunter–herder economy with a nuanced combination of residential and logistical mobilities between ca. AD 600 and AD 1400 (Seitsonen and Viljanmaa 2021). Reindeer feeding and pasture rotation needs also affected people’s lives, as the analysis of the settlement patterns of the early reindeer herders implies that the proximity of good-quality lichen pastures and bogs was already an important factor in the selection of the winter settlement sites between AD 900 and AD 1300 (Halinen et al. 2013; Hedman et al. 2015; Seitsonen and Viljanmaa 2021). From the 15th century onward, major changes in site locations and features indicate a transition to nomadic pastoralism with high residential mobility (Halinen 2016; Mulk 1994; Seitsonen and Viljanmaa 2021). At this time, the settlement pattern changed to temporary settlements in various environments (Bergman et al. 2013; Seitsonen and Viljanmaa 2021).
The mobilities of reindeer and their herders have also been approached recently with the aid of stable isotope analysis. The variations of values of stable isotopes of carbon (δ13C), nitrogen (δ15N), and sulfur (δ34S) imply different patterns of mobilities of people and reindeer (Fjellström et al. 2020; Salmi et al. 2020a). The sites studied thus far vary widely in dating, location, and character, which is why any conclusions about the relationship of mobilities, settlement patterns, and stable isotopes are preliminary at this point. Reindeer bone samples from the 11th to 13th century settlement in Vivallen showed a relatively high variation of stable isotope values, probably reflecting the combination of trade contacts and a mobile way of life (Fjellström et al. 2020). The low variation in the stable isotope values of reindeer bone samples from the settlement of the 17th to 18th century mining community in Silbojokk probably reflect the fact that the mining community’s food was procured from the surrounding area (Fjellström et al. 2020). The high variation in the stable isotope values from reindeer bone samples from the marketplace in Arjeplog is probably related to the role of the marketplace as a place of trade and social exchange (Fjellström et al. 2020). For offering places, the variation in stable isotope values indicates different mobility patterns. Usually, samples from a single offering place are clustered together, indicating that most sites were used mainly by local communities. However, there also were sites with reindeer bone samples with broad isotopic variation: Unna Saiva and Viddjavárri were communal sites likely visited by people from various home regions (Salmi et al. 2020a). Both offering places were large and well known, with rich archaeological finds indicating wide contact networks (Hallström 1932; Manker 1957, pp. 45–52, 167–168; Serning 1956, pp. 87, 119–133). The reindeer offered at Unna Saiva and Viddjavárri were therefore probably domesticated reindeer accompanying people from different areas on their annual migrations between seasonal habitations.
The use of structures and architectures such as fences and enclosures in various reindeer-herding tasks is evident in ethnographic accounts of 19th century and later reindeer herding (e.g., Itkonen 1948; Korhonen 2008). Enclosures were built in connection with habitation sites for milking, supplementary feeding, and keeping reindeer out of the yard (Korhonen 2008, pp. 43–44; Norstedt et al. 2017). Fences were utilized to restrict the movements of reindeer across national and community borders, gather reindeer for counting, marking, and castration, and selecting animals for slaughter in the fall (Korhonen 2008, p. 74; Norstedt et al. 2017) (Fig. 4). However, there is relatively little ancient evidence of such structures in connection with early reindeer herding of the Sámi.

Reindeer fence, Muonio, Finland (photo by Ilmari Manninen, 1929, Finnish Heritage Agency, Kansatieteen Kuvakokoelma)
It is possible that the ancient Sámi herders gathered reindeer without fences, using people, dogs, and bellwethers, or natural features such as gorges (Korhonen 2008, pp. 72–73). On the other hand, subfossil finds of axe-cut trees that date to ca. AD 600–1200 and may have been parts of fence structures have been discovered in the far north of Finland (Eronen et al. 1994; Zetterberg et al. 1994). While researchers have suggested that these finds were wild reindeer hunting fences (Eronen et al. 1994; Zetterberg et al. 1994), the possibility remains that they were related to the handling of domesticated reindeer. Moreover, dendrochronological evidence indicates that barrier fences constructed of local materials such as whole trees and boulders were used from the mid-18th century onward in forested areas of Sweden (Norstedt et al. 2017). Light and small fence structures made of branches are already described in 18th century historical sources (Leem 1956 [1767]). Fences also were constructed of stone, logs, or poles (Korhonen 2008, pp. 85–96; Norstedt et al. 2017). Based on soil chemical evidence, small herds of reindeer may have been kept in enclosures at or near settlement sites between AD 900 and AD 1300 (Jerand and Linderholm 2019). Similar practices of keeping small herds close to the settlement have been documented in Siberia during the first millennium AD (Anderson et al. 2019). Architectures such as enclosures and fences were meeting points for reindeer and people, points of intimacy and care in the landscape shared by people and domesticated reindeer (Anderson et al. 2017a, 2019; Birke 2017). Although little can be said about the temporalities of these architectures based on current knowledge, there seem to have been various meeting points such as enclosures, passageways, and fences for the Sámi and domesticated reindeer in the landscape since the Late Iron Age (Jerand and Linderholm 2019; Seitsonen and Viljanmaa 2021).
Spiritual Entanglements
In addition to everyday animal management practices, human–reindeer entanglements were played out in the religious ritual of the Sámi. In the Sámi worldview, ritual and subsistence activities were not separate but deeply intertwined spheres of life: everyday practices were imbued with spiritual meaning, and spiritual means were used to secure success in subsistence activities (Äikäs et al. 2009). The Sámi worldview was characterized by the idea of a reciprocal relationship between humans, non-humans, and the supernatural (Helander-Renvall 2010). People communicated with deities and spirits of the land and animals by giving offerings such as animals and animal body parts, metal objects, alcohol, and tobacco at sacred sites (Manker 1957; Mebius 2003, pp. 148–153; Serning 1956; Zachrisson 1984) (Fig. 5).

Sieiddakeädgi offering site in Utsjoki, Finland (photo by A. Salmi)
Reindeer feature prominently in the archaeological assemblages from Sámi offering places. Faunal assemblages analyzed from these sites are usually dominated by reindeer bones, especially antlers, crania, and the uppermost vertebrae (Salmi et al. 2018). The presence of postcranial elements, especially marrow-split long bones, indicates that in addition to antlers and crania, reindeer meat and marrow were sometimes consumed as part of the offering ritual (Salmi et al. 2011). The offered reindeer were usually large individuals, probably males, based on osteometric analyses (Salmi et al. 2015). The predominance of heads of large prime-age male individuals is probably related to the impressive antlers of these individuals, emphasizing the symbolic relationship between reindeer antlers and the regeneration of the reindeer species in the Sámi worldview (Olofsson 2010; Salmi et al. 2015).
While the Samí began to make offerings of wild animals such as brown bears and swans ca. AD 500–700, the earliest reindeer offerings date to the late 12th and early 13th centuries (Salmi et al. 2015, 2018). At that time, reindeer herding was probably practiced by some Sámi communities at offering places in a vast geographical area of northern Fennoscandia (Salmi et al. 2018; see also Bergman et al. 2013; Hedman et al. 2015). Settlement patterns indicate that wild reindeer hunting continued to affect habitation patterns (Halinen et al. 2013; Hedman et al. 2015; Seitsonen and Viljanmaa 2021). The presence of wild mammal and bird bones in faunal assemblages (Hedman et al. 2015) further emphasizes the importance of hunting in this period. On the other hand, settlement patterns also may have been affected by small-scale reindeer herding; the age profiles of reindeer bones from archaeological sites, though not reliable indicators of hunting versus herding, imply that small-scale herding was practiced (Hedman et al. 2015). The role of early reindeer herding at the time that offerings of reindeer began to be made is a matter of debate, although emerging evidence indicates that both wild reindeer hunting and reindeer herding were then ritually important for the Sámi (Heino et al. 2021; Salmi et al. 2020a). Based on ancient mtDNA, the reindeer offered in Finland between AD 1200 and AD 1400 were probably wild (Heino et al. 2021), whereas stable isotope analysis has shown that reindeer killed at offering places in Sweden beginning in 13th century were domesticated with human-influenced changes in their diet (Salmi et al. 2020a).
Between AD 1400 and AD 1650, the number of reindeer at Sámi offering places in Finland, Sweden, and Norway peaked (Salmi et al. 2018). There is evidence of a growing emphasis on reindeer herding at this time. Reindeer herding was the main source of livelihood and the basis of social organization for many Sámi communities, especially those residing in mountainous areas (e.g., Bergman et al. 2013), although hunting, fishing, and gathering remained the main sources of livelihood for many Sámi communities, especially the forest Sámi (Harlin et al. 2019). The ritual importance of domesticated reindeer also increased at this time. Stable isotope analyses indicate that reindeer bones at offering places in Finland and Sweden were from domesticated individuals (Salmi et al. 2020a), and that the relative abundance of domesticated reindeer as offerings grew (Núñez et al. 2020). Based on ancient mtDNA, domesticated reindeer were first part of ritual assemblages at offering places in Finland at this time (Heino et al. 2021) (Fig. 6).

After 1650, the importance of animal offering rituals decreased, although some animal bone finds are dated to after AD 1650 (Salmi et al. 2018). This decrease was probably the result of the intensifying Christianization of the Sámi, especially from the 18th century (Äikäs and Salmi 2015; Kylli 2012). However, offering places still retained their meaning, as is testified by the oral tradition attached to these places and several 19th-to-21st century finds such as bottle glass, coins, and the skulls of freshly killed reindeer (Äikäs and Salmi 2015). Today, many offering places are important to various users such as local communities, neo-pagans, and the tourism industry (Äikäs and Spangen 2016).
The investigation of the entanglements of religious ritual and reindeer herding reflects the multifaceted relationships between humans, reindeer, the land, and the supernatural among the Sámi. For the period between ca. AD 1200 and AD 1400, the different lines of archaeological evidence imply that emerging reindeer herding was to some extent intertwined with changes in ritual practice, i.e., the beginning of reindeer offerings, but that wild reindeer hunting was equally if not more important during this early phase. The reindeer offerings given at this time were a means to ensure success in both hunting and herding activities through ritual activity. The growing economic and societal importance of reindeer herding between AD 1400 and 1650 was clearly reflected in the animal rituals at Sámi offering places. Despite the differences between Sámi communities in terms of the emphasis on reindeer herding, hunting, or fishing, the ritual importance of reindeer grew simultaneously across northern Fennoscandia, which indicates there were shared ways of relating to the landscape and its resources among the Sámi practicing differing modes of subsistence (Salmi et al. 2018). The connection between reindeer herding and offering rituals is less evident in the post-AD 1650 archaeological evidence. It is possible that the modern reindeer bones and other modern finds such as euro coins are related to continuing ideas about the relationship between the land, animals, and the supernatural, but they also may reflect neo-pagan or tourist performances at the sites (Äikäs and Spangen 2016).
Chronology for the Evolution of Reindeer-Herding Practices in Northern Fennoscandia
The Hunter–Herder Phase (AD 700–1400)
Previous research on historical and archaeological sources has established that reindeer herding was probably incorporated into the economy of the Sámi of northern Fennoscandia beginning in the Late Iron Age (ca. AD 600/700) with varying regional developments (e.g., Bergman et al. 2013; Halinen et al. 2013; Hedman et al. 2015; Seitsonen and Viljanmaa 2021). Apart from the tentative mention of decoy or draught reindeer by Ohthere (Bately 2007) and reasoning based on ethnographic analogy (Bjørklund 2013), little is known of the reindeer-herding practices in this early stage. Recent archaeological research on reindeer feeding, draught use, settlement patterns, and the ritual practices of the Sámi has considerably deepened our understanding of the integration of reindeer herding into the Sámi economy and social life between AD 700 and AD 1400.
Evidence of care for the well-being of reindeer includes paleoenvironmental data (Hörnberg et al. 2018) and settlement patterns (Halinen et al. 2013; Hedman et al. 2015), which indicate concern about the availability of lichen pastures. Moreover, stable isotope and feeding behavior indicators show that reindeer herding was complemented by feeding from at least the 13th century (Salmi et al. 2020a). Winter feeding was probably supplemented with dried grasses, sedges, and branches when hard-covered snow prevented reindeer from digging for lichen, and/or feeding connected with taming individuals selected for draught training (Salmi et al. 2020a; Soppela et al. 2020). Evidence of reindeer feeding from the 13th to 14th century has been gathered from offering places in northern Sweden. There is also tentative evidence of fence structures and corralling as part of early reindeer herding (Eronen et al. 1994; Jerand and Linderholm 2019; Zetterberg et al. 1994). Although archaeological evidence does not indicate the extent and reason for the use of fences and enclosures, they were used for multiple purposes such as milking, supplemental feeding, and guiding the movements of wild and domesticated reindeer during hunting and roundups by later reindeer herders (Korhonen 2008; Norstedt et al. 2017). Working-related skeletal markers identified in reindeer bone assemblages from Finland show that reindeer were utilized for traction at from at least the 14th century, and probably earlier (Salmi et al. 2021).
Reindeer herding was practiced alongside hunting before AD 1400: faunal assemblages dated to this period include bones of wild birds, mammals (including reindeer), and fish (Bjørnstad et al. 2012; Harlin et al. 2019; Salmi et al. 2021; Vretemark 2019); dwelling sites were located on grounds suitable for the hunting of wild game (Hedman et al. 2015); and offering rituals featured only wild birds, reindeer, and other mammals (Heino et al. 2021; Salmi et al. 2018). Recent GIS analyses have revealed a complex pattern of mobilities related to hunting and herding, with certain meeting points in the landscape for people and reindeer during their seasonal migrations (Seitsonen and Viljanmaa 2021).
Based on this evidence, I argue that reindeer herding between ca. AD 700 and 1400 was probably small in scale, and the subsistence pattern of the herders is best described as a mixed hunting and herding economy (Fig. 7). The proportion of domesticated individuals out of the total number of reindeer in the faunal assemblage is impossible to evaluate osteologically, but stable isotope analyses indicate only a small number of individuals with human-influenced diets (Núñez et al. 2020; Salmi et al. 2020a), and samples analyzed for ancient mtDNA do not show domesticated haplotypes before AD 1400 (Bjørnstad et al. 2012; Heino et al. 2021). Osteometric analyses cannot measure precise numbers of wild and domesticated reindeer, but they do indicate the presence of both (Salmi et al. 2021). These lines of evidence indicate that domesticated herds were small and that wild reindeer hunting played a major role in subsistence. The evidence for supplemental feeding, the use of reindeer for traction, and possible corralling imply close relationships with domesticated herds or specific individuals. Personal bonds were created in association with daily tasks such as feeding and training, involving care, cooperation, and interspecies communication (Salmi et al. 2021). For example, the close bonds between people and domesticated reindeer were manifested in ritual offerings of foddered individuals (Salmi et al. 2020a).

Dating of archaeological evidence and the chronology of the development of reindeer herding among the Sámi of northern Fennoscandia (illustration: by A. Salmi)
The underlying reasons behind the beginnings of reindeer herding among the Sámi are not well understood, but it seems that its gradual adoption was connected to several societal, economic, and environmental changes that took place in northern Fennoscandia in the Late Iron Age. Trade contacts with European markets intensified in the ninth century, creating a heightened need for trade products such as furs, hides, and fish (Kuusela et al. 2020). Climatic fluctuations like the Late Antique Little Ice Age (ca. AD 536–660) and the subsequent climatic warming during the Medieval Climate Anomaly (ca. AD 950–1250) may have instigated changes in resource use patterns (Seitsonen and Viljanmaa 2021). Such changes in economy and resource use may have instigated the intensification of resource management that gradually developed toward domestication. Hansen and Olsen (2014, pp. 203–214) have suggested that the management of wild reindeer hunting infrastructure, and especially hunting fence systems, may have led to the emergence of social organization with leadership and the concept of ownership of resources including animals. It is also possible that the use of reindeer for traction was an important driver of reindeer domestication among the Sámi, and that transportation reindeer were important for maintaining social and trade contacts over vast areas. Sámi settlement patterns show a shift from locations connected by water to locations farther away between ca. AD 700 and AD 800 (Hedman 2003; Seitsonen and Viljanmaa 2021), and transportation reindeer would have been important for the mobility of land-connected societies (Halinen 2022).
The Transition to Mobile Pastoralism (AD 1400–1600 Onward)
The 15th century seems to have been a turning point in the development of reindeer herding in northern Fennoscandia as mobile pastoralism emerged, although with considerable regional variations (Fig. 7). Historical sources increasingly shed light on reindeer-herding practices, as historical sources from the 16th and 17th centuries describe a mobile pastoralist reindeer-herding society in which reindeer milking, draught reindeer, and pasture rotation were in use (Leem 1956 [1767]; Magnus 1996 [1555]; Rheen 1897 [1671]). Nevertheless, archaeological research has revealed additional details about herding practices. The stable isotope data on reindeer feeding patterns show that some reindeer received supplemental fodder and others did not (Salmi et al. 2020a). However, the variation in dietary stable isotope decreased between AD 1400 and AD 1600, which perhaps indicates more uniform feeding patterns (Núñez et al. 2020). Skeletal indicators show that working reindeer also were utilized at this time (Salmi et al. 2021). The religious role of the domesticated reindeer also grew, as the first individuals carrying haplotypes common in modern domesticated reindeer appear in the faunal assemblages from offering places between AD 1400 and AD 1600. Archaeological evidence of reindeer-herding practices thus clearly shows that although there was a transition to mobile reindeer pastoralism, many cultural practices adopted earlier continued, possibly with adaptations to changing economic and social conditions.
The combined effect of colonialism, economic factors, and taxation may have been an important factor in the transition to mobile pastoralism. Economic needs may have led to such an increase in wild reindeer hunting that the populations crashed, creating a need for new ways of resource management in the form of domestication (e.g., Odner 2001; Vorren 1974). Genetic bottlenecks in reindeer populations dated to the 11th–14th centuries indicate overhunting, which may have triggered the need to intensify resource management by managing domesticated herds (Bergstøl 2020; Hansen and Olsen 2014: 204; Røed et al. 2018). A related line of reasoning stresses that the transition to larger-scale reindeer herding in the 15th to 17th centuries created a surplus for the demands of trade networks and taxation by the states that increased their political and economic grip on the north at the same time (Hedman 2003, pp. 223–230; Lundmark 1982; Wallerström 2000). Furthermore, the cooling of the climate during the Little Ice Age (ca. AD 1300–1850) and the Black Death in Scandinavia in the mid-14th century may have caused societal and economic crises, instigating changes in the subsistence strategies of the Sámi (e.g., Carpelan 2003; Seitsonen 2020).
These environmental, societal, and economic changes may have created a need to intensify the production of goods such as reindeer meat and hides for trade, taxation, and livelihood. Herd structure and the emphasis of herding strategy are difficult to evaluate based on current archaeological knowledge, especially because the mixed hunting and herding livelihood complicates the interpretation of faunal assemblages with regard to herding strategies. However, there is tentative evidence, such as a slightly smaller proportion of working reindeer (Hull and Salmi n.d.) and a slightly larger proportion of younger individuals slaughtered (Harlin et al. 2019), indicating that the importance of meat production may have risen slightly even within communities practicing a mixed livelihood of hunting and herding. However, the use of reindeer for transportation was still immensely important, as draught and cargo reindeer enabled large-scale seasonal migrations and long-distance trade journeys (Salmi and Heino 2019). However, it is clear that the adoption of mobile pastoralism happened at different times among different Sámi groups, and some did not adopt it at all, retaining a hunter–herder livelihood until the 20th century (Carpelan 2003; Hansen and Olsen 2014: 35; Harlin et al. 2019; Itkonen 1948). Moreover, the transition to large-scale mobile reindeer pastoralism occurred not only among the Sámi but also among other Eurasian peoples such as the Nenets, Chukchi, Koryak, Enets, and northern Khanty (Krupnik 1993; Stépanoff 2017). In the 19th century, herd sizes had hugely increased across the Eurasian Arctic, leading to a highly specialized subsistence strategy described as the “the reindeer revolution” (Krupnik 1993; Stépanoff 2017). At this point, the main activity of most households was reindeer herding with large herds, which provided herders with most of their food and other resources (Stépanoff 2017).
Pathways to Reindeer Domestication
The chronology of reindeer domestication in Fennoscandia has puzzled scientists, so the wider context of the process and its connections with other animal domestication processes merit scientific scrutiny. The interdependencies of reindeer-herding traditions in Fennoscandia and Siberia have long been debated (e.g., Storli 1996). Whereas evolutionary explanations of the origins of reindeer domestication stress that reindeer herding developed separately in Fennoscandia and Siberia, diffusionist explanations maintain that reindeer herding developed first in Siberia and spread from there (e.g., Gordon 1990; Kortesalmi 2008, pp. 20–21; Mirov 1945; Storli 1996). Although the debate remains open, recent archaeological evidence has shed some new light on the possible similarities, dissimilarities, and connections between the development of reindeer herding in these areas.
There likely are several origins of reindeer herding in Siberia. Early 20th century literature cites Chinese historical sources and rock art depicting possibly harnessed and ridden reindeer as evidence of reindeer domestication centers in the Sayan Mountains or the Lake Baikal area, possibly around the turn of the first millennium AD, and with influences from the pastoral peoples of the steppes farther south (e.g., Laufer 1917; Mirov 1945). Recently, researchers have also argued for an early onset of reindeer herding in the Yamal Peninsula (Anderson et al. 2019; Losey et al. 2021). Multiple sources of evidence such as geochemical analyses, objects interpreted as transportation reindeer headgear, and portable reindeer imagery indicate that domesticated reindeer were held and trained there from the early centuries AD (Anderson et al. 2019; Losey et al. 2021; Nomokonova et al. 2021). The reindeer economy seems to have included small-scale reindeer herding as part of a mixed economy of hunting, fishing, fowling, and gathering berries (Anderson et al. 2019). Reindeer herding therefore seems to have begun in Siberia earlier than in Fennoscandia, which may in turn indicate that the Sámi got the idea and expertise of reindeer herding from Siberian reindeer-herding communities.
Genetic lineages common in today’s domestic herds in Fennoscandia occurred in Siberia as early as 3900 BC, and researchers have argued that these lineages spread to Fennoscandia from Siberia (Røed et al. 2018, 2020). However, when the results of analyses of nuclear and mitochondrial DNA are compared, the situation is more complicated. The Fennoscandian domestic reindeer nuclear DNA lineages are clustered with native Norwegian mountain reindeer lineage, while the mitochondrial DNA lineages seem non-native to Fennoscandia (Røed et al. 2008; Weldenegodguad et al. 2020). This pattern may be a result of controlled breeding, in which wild mountain reindeer males were deliberately or incidentally allowed to breed with domestic reindeer females (Heino and Pelletier 2022). A similar practice has been observed for the Skolt Sámi (Helle 1982) and the Evenki (Anderson et al. 2017b). Furthermore, there remains a possibility that domestic reindeer belonging to different lineages than those common today were extant before the spread of haplotypes II and Ib, the spread of which is probably related to the emergence of mobile pastoralism with large reindeer herds (Heino and Pelletier 2022).
The exact relationship between the Sámi and various Siberian reindeer-herding traditions therefore remains unsolved, but it is clear there were cultural connections between the human groups populating these areas. For example, the material culture and biological evidence from the Early Metal Age (ca. 1500–1100 BC) burial site of Oleneostrovskiy in the Kola Peninsula indicates contacts to both the west and the east (Murashkin et al. 2016). Furthermore, material culture and especially metal objects analyzed from Sámi habitation and offering sites dating to ca. AD 1000–1300 indicate a vast contact network to both the west and the east (e.g., Halinen 2016; Serning 1956; Zachrisson 1984). It therefore seems plausible that there were connections between the reindeer-herding traditions of the Sámi and Siberian reindeer-herding peoples, possibly entailing the transfer of technological expertise and domestic reindeer stock. This also seems reasonable given the similarity of the early reindeer-herding adaptations in Siberia and Fennoscandia, both probably being a mix of small-scale herding inclusive of working reindeer use, hunting, and gathering.
The models of transitions in reindeer domestication in Fennoscandia resonate with general models of animal domestication, especially with the prey and directed pathways to domestication (Zeder 2012, 2015). The prey pathway to domestication entails a process in which humans deliberately aim to increase the supply of a game species (Zeder 2012, 2015). The need to increase the efficiency of resource management has often resulted from overhunting (Larson and Fuller 2014). The theories of the origin of reindeer domestication that stress economic factors leading to possible overhunting, resource management intensification, and related societal changes are concurrent with the prey pathway to domestication. The fitness of the prey pathway model to reindeer domestication is also supported by the fact that wild reindeer was an important game species for the Sámi before the onset of reindeer herding (e.g., Hansen and Olsen 2014, pp. 80–81; Kuusela et al. 2020). Moreover, the religious role of wild reindeer offerings before the intensification of reindeer herding testifies to the importance of wild reindeer to the Sámi economy and worldview (Heino et al. 2021). The dwelling patterns of the Sámi at the time of reindeer domestication may have been adapted to wild reindeer hunting, as well as reindeer herding (e.g., Halinen et al. 2013; Hedman et al. 2015).
The prey pathway model of domestication emphasizes the importance of reindeer meat and hides, as opposed to traction, as a driving force behind the adoption of reindeer herding. On the other hand, if the hypothesis of the importance of draught reindeer in early reindeer domestication (Bjørklund 2013; Ingold 1986) holds true, elements related to another pathway to domestication, the directed pathway (Zeder 2012, 2015), would be relevant for reindeer domestication. The directed pathway begins with a deliberate attempt to domesticate a species to fulfill specific needs, for example, traction (Zeder 2012, 2015). The directed pathway requires prior familiarity with other domesticated animal economies (Zeder 2012). The directed pathway theory is supported by the earlier adoption of reindeer herding and draught reindeer use in Siberia (Anderson et al. 2019; Losey et al. 2021; Nomokonova et al. 2021), which would then have acted as a potential model for the Sámi to adopt reindeer herding. The possible Siberian genetic origins of Fennoscandian domesticated reindeer (Røed et al. 2018) also supports the directed pathway model of reindeer herding among the Sámi. The osteological record at our disposal does not go far enough in time to allow us to conclude whether draught reindeer were indeed an important factor in the early reindeer domestication of the Sámi (Bjørklund 2013; Ingold 1986), but the evidence of the use of working reindeer in small-scale reindeer herding in Finland in the 14th century may point in that direction, because it is possible that the early reindeer herders employed a similar set of mixed subsistence strategies as the Sámi in northeastern Fennoscandia did from the 14th century onward (Salmi et al. 2021).
The archaeological evidence at hand therefore shows that the domestication of reindeer among the Sámi probably followed a path that was a combination of elements from the prey and directed pathways. It is clear that wild reindeer hunting predates reindeer herding, and it is likely that intensification of resource management played a role in the transition to reindeer herding. On the other hand, the Sámi may have had prior knowledge of reindeer herding acquired through contacts with reindeer-herding Siberian peoples, and this knowledge may have prompted the adoption of reindeer herding, possibly the introduction of genetic lineages common in today’s herds, as well as the use of draught reindeer.
Negotiating the Domestication Relationship
As reindeer were domesticated, they acquired new roles in human economies and societies. A common thread in the animal domestication literature posits that animal domestication presents a profound change in one way or another in the relationship between humans and animals (e.g., Ingold 2000; Knight 2012; Oma 2010). The change may entail domesticated animals becoming private property controlled by humans (e.g., Ingold 2000). Although the relationships with both wild and domesticated animals can be close and intimate, closeness and intimacy may take different forms when animals are domesticated (e.g., Knight 2012; Oma 2010). In addition to changes, the continuations in the relationships between people and animals through the transition to domestication merit attention.
One of the potential continuous processes relevant to reindeer domestication is the management of game animal stock and its possible transformation into a domestication relationship. As outlined earlier, reindeer hunting had been incorporated into the prehistoric economies of people in northern Fennoscandia since the area was populated after the last Ice Age. The use of various hunting strategies and structures such as pitfall systems, fences, and decoy animals (e.g., Halinen 2005, pp. 82–94; Helskog 2011b, 2012; Hennius 2020; Jordhøy 2008) shows a detailed and intimate knowledge of reindeer ecology and behavior and the management of their movements and actions using this knowledge. Researchers have even argued that the management of the fence structures has been one of the driving forces behind the societal changes leading to reindeer herding (Hansen and Olsen 2014, pp. 203–214). Such practices can be seen as a continuum from wild animal management and hunting to the corralling, fencing, and guiding of the movements of domesticated reindeer in a reindeer-herding economy. Another example of a continuous practice is the use of transportation technology. Sleds and sled parts have been discovered in northern Fennoscandia dating from the Mesolithic onward, and the sleds may have been pulled by dogs, people, or reindeer (e.g., Kuokkanen 2000). Although an ambiguous source of information for the study of reindeer domestication, it shows a continuum in the use of transportation technology that may have been and probably was used with several species. The continuation between hunting and herding is also emphasized by the fact that wild reindeer hunting was long incorporated into the subsistence economy of the Sámi groups also practicing reindeer herding (e.g., Salmi et al. 2021). Wild reindeer also retained their religious importance throughout the various transitions in reindeer herding (Salmi et al. 2018).
It is not possible to pinpoint a definite point when hunting changed into herding, because the transition was a long process in which cultural practices and technologies were adapted to new forms of human–reindeer interaction. However, we can try to outline some of the defining characteristics of the relationship between domesticated reindeer and herders. An outline of these characteristics is also potentially helpful for understanding how human–animal relationships may have evolved in the domestication processes of other animal species. First, care and trust building have been crucially important in the relationships between reindeer and herders. An archaeologically documented example of care for the well-being of the reindeer is supplemental feeding. In today’s reindeer herding, and probably also in the past, feeding has enabled herders to monitor and affect the well-being of the reindeer, build trust between people and reindeer, and help in taming and training draught reindeer (Soppela et al. 2020; Salmi et al. 2022). Reindeer also learn to anticipate feeding and other forms of care from the herders (Salmi et al. 2022; Turunen and Vuojala-Magga 2014).
Moreover, the relationships between herders and reindeer are and probably always have been constructed though interspecies practice and learning. For example, the training of working reindeer is based on the mutual learning and trust that come with regular contact and attuning to each other’s feelings and reactions. Thus, reindeer personhood plays an important role in this mutual learning and cooperation (Soppela et al. 2020; Vuojala-Magga 2010). The pastoral Sámi understood animals as persons, capable of communication, emotions, intentional action, and meaningful relationships with people, although animal personhood was considered different from human personhood, arising in certain situations and variable between individuals (Helander-Renvall 2010). Domesticated reindeer were creatures with personhood and agency, played out in various ways in reindeer herders’ practical tasks, social interactions, and religious rituals. In the Sámi worldview, it was understood that reindeer shared a common environment, as well as moments of communication and reciprocity with humans (Helander-Renvall 2010). The Sámi, the reindeer, and other living creatures therefore constituted a multispecies society in which humans and non-humans both took part in social interactions and engagement (Boyd 2017; Kirksey and Helmreich 2010; Pilaar Birch 2018; Salmi and Heino 2019).
In a broader sense, the relationship between reindeer and herders also was constructed through the herding tasks performed in the landscape with the animals. The landscape-level characteristic of animal domestication has been emphasized by Stépanoff (2017), who argues that reindeer domestication is a process in which both humans and animals modify their behavior to learn to live together in a shared landscape. It is emphasized that this learning and adaptation affects all parties: reindeer have motivation and agency to enter the human association; humans balance the needs of the reindeer with their own needs; and the landscape undergoes changes associated with the new forms of human–reindeer interaction (Beach and Stammler 2006; Istomin and Dwyer 2010; Stépanoff 2017). The landscapes of reindeer herding consisted of the daily tasks and mobilities that were designed to meet the various needs of reindeer and herders. They also were shaped by spiritual concerns for the well-being of humans and reindeer that were negotiated with the supernatural powers at various sacred sites in the landscape. Moreover, reindeer can be seen as essential creatures in the construction of the herders’ lived-in worlds. As the world constantly comes into being and is transformed when different beings act in it in relation to other beings (Ingold 2011, pp. 67–75), and humans and animals become who they are in relation to each other (Haraway 2008, pp. 15–27), reindeer herding among the Sámi was and remains a constant process of inhabiting the world with the reindeer. Domesticated reindeer were active participants in the constant creation of the herders’ lived-in worlds. This co-creation occurred in shared spaces and meeting places, and it constituted the daily tasks of reindeer herding.
Some of the features of reindeer domestication in northern Fennoscandia may be unique to the reindeer species, regional environmental conditions, and Sámi culture. For example, the interplay of climate and reindeer migratory behavior contributed to the development of Sámi mobile pastoralism based on following the seasonal movements of reindeer herds between habitats. However, some of the insights learned from reindeer domestication in northern Fennoscandia can inform us about the more general characteristics of animal domestication potentially present in the domestication processes of other species. First, care for the well-being of animals is an important characteristic of the domestication relationship and animal management and is probably present more widely in domestication processes. Second, interpersonal relationships and cooperation with some or all domesticated animals are likely to form in the daily practices shared by people and animals. These aspects of animal domestication are manifested not only at an interpersonal level but more broadly in the landscape, society, and religion. Animal domestication can therefore be seen as a social relationship characterized by communication, cooperation, and care, maintained and constructed through interspecies encounters in everyday activities and spiritual entanglements. This relationship is constantly renegotiated in everyday interaction with the animals and is therefore ever-changing and never complete. In these daily encounters, the domestication relationship and the shared world of humans and animals are built. Their archaeological research therefore holds the key to understanding the transformations of the lives of humans and animals participating in the domestication process. The various interactions visible in the archaeological record have the potential to enhance our understanding of the social components of animal domestication, adding nuances to the chronologies and narratives of animal domestication.
Conclusion
The domestication of the reindeer and the development of various reindeer-herding traditions in northern Eurasia have been the focus of archaeological, historical, and ethnographic scrutiny for more than 100 years. In this review, I have concentrated on these processes among the Indigenous Sámi of northern Fennoscandia, especially referencing new archaeological results that have become available in recent years through methodological advances and theoretical insights. As my focus has been on the development of various reindeer-herding practices and their social and religious dimensions, I have especially discussed the archaeological evidence of reindeer feeding, draught reindeer use, reindeer-herding landscapes and architectures, and the role of reindeer in religious ritual. These insights were incorporated into the general chronology of reindeer domestication and the understanding of the adoption of reindeer herding among the Sámi and in a wider northern Eurasian context. They also were utilized to explore the implications of reindeer domestication on human–reindeer sociality and to discuss how animal domestication studies can benefit from understanding how the domestication relationship is constructed in daily encounters and interactions with the animals.
Several important issues remain for future studies of reindeer domestication. First, the recent development in archaeological methodologies and combination of different methodologies, including analyses of stable isotopes, ancient DNA, physical activity markers, and landscape archaeology have opened new possibilities to trace human–reindeer interactions in the domestication process. In particular, osteological methods such as paleopathology, geometric morphometrics, and the study of bone cross-sectional properties and muscle attachment site morphology offer promising avenues for elucidating various physical activity patterns such as draught and cargo use, corralling, and feeding behavior relevant for animal domestication studies. Some of these methods have yet to be applied to archaeological data, and none has been applied in a large-scale study. Method development targeting a more nuanced understanding of physical activity patterns based on more comprehensive data sets with animals with various activity patterns also will be needed. In the future, these methodologies should be combined with other analytical possibilities with the potential to reveal past human–animal interactions, such as various soil analyses. In addition, future research should address the problems and potential in combining various data sets for animal domestication studies. For example, landscape archaeological analyses, faunal analyses, and the analysis of ancient DNA operate at very different levels and timescales. While the analysis of ancient DNA reflects population-level changes over a long period, the study of phenotypically plastic morphological changes through geometric morphometrics and physical activity markers can more efficiently target incipient domestication in a more restricted timescale. Moreover, approaches such as landscape archaeology and the archaeology of religion can reveal culturally significant aspects of the human–domesticate relationship. Current reindeer domestication studies often approach the problem using a single perspective or methodology. Comprehensive multi-proxy analyses will therefore be important for the future study of both reindeer domestication and animal domestication in general.
The second important topic to address is regional variations in the development of reindeer herding. The archaeological and historical evidence shows there were regional variations in reindeer-herding traditions among the Sámi. In the 17th century and later literary sources, the different subsistence patterns of the forest and mountain Sámi are mentioned. Earlier variation in reindeer-herding traditions is more difficult to pinpoint due to the patchiness of the archaeological data. Some of the transitions such as the changing mobility patterns related to the development of a hunting–herding adaptation and mobile pastoralism and the shifts in the foci of religious ritual seem to have happened simultaneously in relatively large areas in northern Finland, Sweden, and Norway (Salmi et al. 2018; Seitsonen and Viljanmaa 2021). Reindeer feeding seems to have been a regionally and temporally variable practice (Salmi et al. 2020a). Because many of the other reindeer-herding practices such as the draught use and corralling of reindeer are presently only documented from one or a handful of sites (Jerand and Linderholm 2019; Salmi et al. 2021), the representativeness of the archaeological data allows no specific conclusions about how widespread they were. As the current data sets are patchy in their regional and temporal representativeness, it will be important to build larger and more representative data sets in the future to elucidate the local histories and variations in the adoption and development of reindeer herding among the Sámi. Such studies also will need to consider the integration of reindeer herding into other forms of land use such as hunting, fishing, cultivation, and gathering. Future research will need to address the interdependencies, similarities, and differences in the development of reindeer herding in Fennoscandia and Siberia, with more comprehensive and representative data sets from both areas.
The third potential avenue for future research is the examination of animal domestication through the concept of everyday social interaction. The examination of human–reindeer interactions and sociality in the context of reindeer-herding tasks has shown that interspecies communication, cooperation, and care were important components of reindeer herding. The social bonds between people and reindeer were maintained and constructed through various interspecies and interpersonal encounters in everyday and ritual activities performed in the landscape. By emphasizing these aspects of the domestication relationship, I have argued that reindeer domestication was and remains an ongoing process of everyday interaction between people and animals through which social bonds are constructed and constantly renegotiated. Although Zeder (2015) has suggested that the social components of the human–domesticate relationships are characteristics of animal management practices rather than domestication itself, I argue that these aspects cannot be separated from the physiological, genetic, and ecological components of animal domestication but should be examined as an integral part of the way in which the animals gradually entered into human association and remained there. When animals entered the domestication relationship, their roles as participants of the multispecies society changed, as well as how they participated in the construction of the lived-in worlds of the humans domesticating them. The meeting points, spaces, and everyday cultural practices through which these lived-in worlds were constructed can be analyzed through the archaeological record. Some potentially promising avenues for future research, currently poorly covered in the archaeological literature on reindeer domestication, would be subtle landscape modifications and manipulations to attract reindeer and cater to their needs, such as the controlled burning of vegetation to create feeding opportunities or the use of smoke to repel insects.
Fourth, an important aspect to consider in future reindeer domestication studies is the involvement of local communities and the integration of reindeer herders’ traditional knowledge about the research process. Such approaches have proven to offer extremely valuable insights into domestication histories among other reindeer herders (e.g., Anderson et al. 2019; Losey et al. 2021), but they are still less widespread than they could be in the Sámi archaeology practiced in Finland, Sweden, and Norway (Harlin 2019; Spangen et al. 2020). The traditional knowledge of today’s reindeer herders cannot be projected on the past as such because cultural practices are ever-changing (e.g., Politis 2014; Stump 2013), although elements of traditional knowledge can persist with a high degree of coherence over a long period (Smith 2012). The traditional knowledge of reindeer herders can therefore inform us about how human–reindeer relationships were built and maintained in herding practices. As shown by some of the examples discussed in this paper, especially regarding feeding and draught reindeer use, the integration of reindeer herders’ traditional knowledge has the potential to add important nuances to the understanding of reindeer domestication in Fennoscandia. Although the specific rationales and techniques of reindeer supplemental feeding or draught reindeer training may have changed, the elements of care, interspecies communication, and trust building were probably part of past reindeer herding in a similar way to today.
The understanding of elements of change and continuity in reindeer-herding practices is especially pertinent today, as the social, political, climatic, and economic environment of reindeer herding is changing quickly. Climate change and competing forms of land use have necessitated a growing dependence on supplemental feeding, instigating changes in the economic and social conditions of reindeer herding (Heikkinen et al. 2012; Horstkotte et al. 2020). Herd structure has undergone marked changes with the emphasis of reindeer herding changing from a multipurpose herd to one intended primarily for meat production (Holand 2007). There are therefore economic and environmental pressures to increase herd sizes and increase supplemental feeding, but herders do not necessarily see these as viable options for their long-term livelihood (Heikkinen et al. 2012; Horstkotte et al. 2020). However, traditions once close to disappearance, such as draught reindeer training, have been revived in connection with the tourism boom that has created a need for reindeer safaris (Soppela et al. 2020). In the future, the combination of meat trade and processing, crafts, and tourism is expected to support the economic and cultural sustainability of reindeer-herding communities (Rantamäki-Lahtinen 2008). An understanding of changes in these traditions provided by archaeological research is important for today’s reindeer herders for situating the past, present, and future of their livelihood in the context of the long-term history of reindeer herding.
References Cited
Amundsen, H., and Os, K. (2020). Funnel-shaped reindeer trapping systems in Hedmark - Saami or Norse? In Spangen, M., Salmi, A., Äikäs, T., and Fjellström, M. (eds.), Currents of Saami Pasts: Recent Advances in Saami Archaeology, Monographs of the Archaeological Society of Finland 9, Archaeological Society of Finland, Helsinki, pp. 16–33.
Anderson, D. G., Harrault, L., Milek, K. B., Forbes, B. C., Kuoppamaa, M., and Plekhanov, A. V. (2019). Animal domestication in the high Arctic: Hunting and holding reindeer on the I͡Amal peninsula, northwest Siberia. Journal of Anthropological Archaeology 55: 101079.
Anderson, D. G., Kvie, K. S., Davydov, V. N., and Røed, K. H. (2017b). Maintaining genetic integrity of coexisting wild and domestic populations: Genetic differentiation between wild and domestic Rangifer with long traditions of intentional interbreeding. Ecology and Evolution 7: 6790–6802.
Anderson, D., Loovers, J. P. L., Schroer, S. A., and Wishart, R. P. (2017a). Architectures of domestication: On emplacing human-animal relations in the North. Journal of the Royal Anthropological Institute 23: 398–418.
Arbuckle, B. S., and Makarewicz, C. A. (2009). The early management of cattle (Bos taurus) in Neolithic central Anatolia. Antiquity 83: 669–686.
Äikäs, T., Puputti, A., Núñez, M, Aspi, J., and Okkonen, J. (2009). Sacred and profane livelihood: Animal bones from sieidi sites in northern Finland. Norwegian Archaeological Review 42: 109–22.
Äikäs, T., and Salmi, A. (2015). North/south encounters at Sámi sacred sites in northern Finland. Historical Archaeology 49: 90–109.
Äikäs, T., and Spangen, M. (2016). New users and changing traditions—(Re)defining Sami offering sites. European Journal of Archaeology 19: 95–121.
Aronsson, K. (1991). Forest Reindeer Herding AD 1–1800: An Archaeological and Palaeoecological Study in Northern Sweden, Archaeology and Environment 10, University of Umeå, Umeå.
Baglo, C. (2019). The disappearance of the Sea Sámi as a cultural display category. Nordic Museology 27(3): 25–44.
Banfield, A. W. F. (1961). A Revision of the Reindeer and Caribou, Genus Rangifer, National Museum of Canada, Ottawa.
Barboza, P. S., and Parker, K. L. (2006). Body protein stores and isotopic indicators of N balance in female reindeer (Rangifer tarandus) during winter. Physiological and Biochemical Zoology: Ecological and Evolutionary Approaches 79: 628–644
Bately, J. (2007). Text and translation. In Bately, J., and Englert, A. (eds.), Ohthere’s Voyages: A Late 9th Century Account of Voyages Along the Coasts of Norway and Denmark and Its Cultural Context, The Viking Ship Museum, Roskilde, pp. 40–58.
Beach, H., and Stammler, F. (2006). Human-animal relations in pastoralism. Nomadic Peoples 10(2): 6–30.
Beck, A., and Mayr, C. (2012). Nitrogen and carbon isotope variability in the green-algal lichen Xanthoria parietina and their implications on mycobiont–photobiont interactions. Ecology and Evolution 2: 3132–3144.
Bergman, I., Zackrisson, O., and Liedgren, L. (2013). From hunting to herding: Land use, ecosystem processes, and social transformation among Sami AD 800–1500. Arctic Anthropology 50(2): 25–39.
Bergstøl, J. (2020). Hunting native reindeer, while herding imported ones? Some thoughts on the development of Saami pastoralism. In Spangen, M., Salmi, A., Äikäs, T., and Fjellström, M. (eds.), Currents of Saami Pasts: Recent Advances in Saami Archaeology, Monographs of the Archaeological Society of Finland 9, Archaeological Society of Finland, Helsinki, pp. 34–45.
Bezard, P., Brilland, S., and Kumpula, J. (2015). Composition of late summer diet by semi-domesticated reindeer (Rangifer tarandus tarandus) in different grazing areas in northernmost Finland. Rangifer 35: 39–52.
Birke, L. (2017). Meeting points: Choreographies of horses and humans. Trace 3: 54–70.
Bjørklund, I. (2013). Domestication, reindeer husbandry and the development of Sámi pastoralism. Acta Borealia 30: 174–189.
Bjørnstad, G., Flagstad, Ø., Hufthammer, A., and Røed K. H. (2012). Ancient DNA reveals a major genetic change during the transition from hunting economy to reindeer husbandry in northern Scandinavia. Journal of Archaeological Science 39: 102–108.
Bocquet-Appel, J. (2002). Paleoanthropological traces of a Neolithic demographic transition. Current Anthropology 43: 637–650.
Bogaard, A., Allaby, R., Arbuckle, B. S., Bendrey, R., Crowley, S., Cucchi, T., et al. (2021). Reconsidering domestication from a process archaeology perspective. World Archaeology 53: 56–77.
Bökönyi, S. (1989). Definitions of animal domestication. In Clutton-Brock, J. (ed.), The Walking Larder: Patterns of Domestication, Pastoralism, and Predation, Unwin Hyman, London, pp. 22–27.
Boyd, B. (2017). Archaeology and human–animal relations: Thinking through anthropocentrism. Annual Review of Anthropology 46: 299–316.
Brännström, M. (2020). The enhanced role of archaeological and historical research in court proceedings about Saami land rights. In Spangen, M., Salmi, A., Äikäs, T., and Fjellström, M. (eds.), Currents of Saami Pasts: Recent Advances in Saami Archaeology, Monographs of the Archaeological Society of Finland 9, Archaeological Society of Finland, Helsinki, pp. 177–188.
Britton, K. H. (2009). Multi-Isotope Analysis and the Reconstruction of Prey Species Palaeomigration and Palaeoecology, Doctoral thesis, Department of Archaeology, Durham University, Durham.
Carpelan, C. (1991). Peuranpyytäjien talvikylä Inarissa. Raito 2: 20–28.
Carpelan, C. (1993). Juikenttä – Näkökulma saamelaiseen yhteiskuntaan, Erikoispainossarja No. 287, Oulun yliopiston Historian laitos, Oulu.
Carpelan, C. (2003). Inarilaisten arkeologiset vaiheet. In Lehtola, V.-P. (ed.), Inari – Aanaar, Inarin historia jääkaudesta nykypäivään, Inarin kunta, Oulu, pp. 28–95.
Cauvin, J. (2000). The Birth of the Gods and the Origins of Agriculture, translated by T. Watkins, Cambridge University Press, Cambridge.
Clark, N. (2007). Animal interface: The generosity of domestication. In Cassidy, R., and Mullin, M. (eds.), Where the Wild Things Are Now? Domestication Reconsidered, Berg, Oxford, pp. 49–70.
Clutton-Brock, J. (1992). A Natural History of Domesticated Mammals, Cambridge University Press, Cambridge.
Eronen, M., Zetterberg, P., and Okkonen, J. (1994). Kirveen ja vaakanävertäjän (Tomicus minor, Hart.) jälkiä Lapin subfossiilisissa männyissä. Terra 106: 238–248.
Evans, R. D. (2007). Soil nitrogen isotope composition. In Michener, R. H., and Lajtha, K. (eds.), Stable Isotopes in Ecology and Environmental Science, Malden, Blackwell, pp. 83–98.
Fjellström, M., Eriksson, G., Angerbjörn, A., and Lidén, K. (2020). Approaching historic reindeer herding in northern Sweden by stable isotope analysis. Journal of Nordic Archaeological Science 19: 63–75.
Fjellström, P. (1986). Samernas samhälle i tradition och nutid, Norstedt, Stockholm.
Germonpré, M., Sablin, M. V., Stevens, R. E., Hedges, R. E. M., Hofreiter, M., Stiller, M., and Després, V. R. (2009). Fossil dogs and wolves from Palaeolithic sites in Belgium, the Ukraine and Russia: Osteometry, ancient DNA and stable isotopes. Journal of Archaeological Science 36: 473–490.
Gillis, R., Bréhard, S., Bălăşescu, A., Ughetto-Monfrin, J., Popovici, D., Vigne, J., and Balasse, M. (2013). Sophisticated cattle dairy husbandry at Borduşani-Popină (Romania, fifth millennium BC): The evidence from complementary analysis of mortality profiles and stable isotopes. World Archaeology 45: 447–472.
Gordon, B. C. (1990). World Rangifer communal hunting. In Davis, L. B., and Reeves, B. O. K. (eds.), Hunters of the Recent Past, Unwin Hyman, London, pp. 277–303.
Greenfield, H. J. (2010). The secondary products revolution: The past, the present and the future. World Archaeology 42: 29–54.
Gustine, D. D., Barboza, P. S., Lawler, J. P., Adams, L. G., Parker, K., Arthur, S. M., and Shults, B. S. (2012). Diversity of nitrogen isotopes and protein status in caribou: Implications for monitoring northern ungulates. Journal of Mammalogy 93: 778–790.
Halinen, P. (2005). Prehistoric Hunters of Northern-most Lapland: Settlement Patterns and Subsistence Strategies, Iskos 14, Suomen Muinaismuistoyhdistys, Helsinki.
Halinen, P. (2016). The inland Sámi societies of northern Fennoscandia during the Late Iron Age and early medieval period: An archaeological approach. In Uino, P., and Nordqvist, K. (eds.), New Sites, New Methods: Proceedings of the Finnish-Russian Archaeological Symposium, Helsinki, 19–21 November 2014, Finnish Antiquarian Society, Helsinki, pp. 160–174.
Halinen, P. (2022). Viewpoints on the disintegration of the Late Iron Age and early medieval Sámi societies and the emergence of reindeer herding in northern Fennoscandia. In Halinen, P., Heyd, V., and Mannermaa, K. (eds.), Odes to Mika: Festschrift of Professor Mika Lavento on the Occasion of his 60th Birthday, Archaeological Society of Finland, Helsinki, pp. 147–155.
Halinen, P., Hedman, S., and Olsen, B. (2013). Hunters in transition: Sámi hearth row sites, reindeer economies and the organization of domestic space, 800–1300 AD. In Anderson, D. G., Wishart, R. P., and Vaté, V. (eds.), About the Hearth: Perspectives on the Home, Hearth and Household in the Circumpolar North, Berghahn Books, New York, pp. 152–182.
Hallström, G. (1932). Lappska offerplatser. In Edén, N. (ed.), Arkeologiska studier tillägnade H.K.H. Kronprins Gustaf Adolf, Norstedt, Stockholm pp. 111–131.
Hambleton, E., and Rowley-Conwy, P. (1997). The medieval reindeer economy at Gæccevaj’njar’ga 244 B in the Varanger Fjord, north Norway. Norwegian Archaeological Review 30: 55–70.
Hansen, L. I., and Olsen, B. (2014). Hunters in Transition: An Outline of Early Sami History, Brill, Leiden.
Haraway, D. (2008). When Species Meet, University of Minnesota Press, Minneapolis.
Harbers, H., Zanolli, C., Cazenave, M., Theil, J., Ortiz, K., Blanc, B., et al. (2020). Investigating the impact of captivity and domestication on limb bone cortical morphology: An experimental approach using a wild boar model. Scientific Reports 10: 19070.
Harlin, E. (2019). Sámi archaeology and the fear of political involvement: Finnish archaeologists’ perspectives on ethnicity and the repatriation of Sámi cultural heritage. Archaeologies 15: 254–284.
Harlin, E., Mannermaa, K., and Ukkonen, P. (2019). Animal bones from medieval and early modern Saami settlements in Finnish Lapland. In Mannermaa, K., Manninen, M. A., Pesonen, P., and Seppänen, L. (eds.), Helsinki Harvest: Proceedings of the 11th Nordic Conference on the Application of Scientific Methods in Archaeology, Monographs of the Archaeological Society of Finland 7, Archaeological Society of Finland, Helsinki, pp. 149–177.
Hedman, S. (2003). Boplatser och offerplatser: Ekonomisk strategi och boplatsmönster bland skogssamer 700–1600 AD, Umeå University, Umeå.
Hedman, S., Olsen, B., and Vretemark, M. (2015). Hunters, herders and hearths: Interpreting new results from hearth row sites in Pasvik, arctic Norway. Rangifer 35: 1–24.
Heikkinen, H. I., Kasanen, M., and Lépy, E. (2012). Resilience, vulnerability and adaptation in reindeer herding communities in the Finnish-Swedish border area. Nordia Geographical Publications 41(5): 107–121.
Heino, M., and Pelletier, M. (2022). Impact of human selection on the genetics and morphology of the reindeer. In Salmi, A. (ed.), Domestication in Action: Human-Reindeer Interaction in Past and Present Reindeer Herding, Palgrave Macmillan, Cham, pp. 35–61.
Heino, M. T., Salmi, A., Äikäs, T., Mannermaa, K., Kirkinen, T., Sablin, M., et al. (2021). Replacement of wild reindeer genetic lineages by domestic ones at Sámi offering sites in Finland from 1400–1600 AD onwards. Journal of Archaeological Science Reports 35: 102691.
Helander-Renvall, E. (2010). Animism, personhood and the nature of reality: Sami perspectives. Polar Record 46(1): 44–56.
Helle, T. (1982). Peuran ja poron jäljillä, Kirjayhtymä, Helsinki.
Helle, T. P., and Jaakkola, L. M. (2008). Transitions in herd management of semi-domesticated reindeer in northern Finland. Annales Zoologici Fennici 48: 81–101.
Helskog, K. (2011a). Humans and reindeer. Quaternary International 238: 1–3.
Helskog, K. (2011b). Reindeer corrals 4700–4200 BC: Myth or reality? Quaternary International 238: 25–34.
Helskog, K. (2012). Ancient depictions of reindeer enclosures and their environment. Fennoscandia Archaeologica XXIX: 29–54.
Hennius, A. (2020). Towards a refined chronology of prehistoric pitfall hunting in Sweden. European Journal of Archaeology 23: 530–546.
Hodder, I. (2011). The role of religion in the Neolithic of the Middle East and Anatolia with particular reference to Çatalhöyük. Paléorient 37(1): 111–122.
Holand, Ø. (2007). Flokkstruktur og slaktestrategi i reindrifta – Et historisk perspektiv. Rangifer Report 12: 21–33.
Horstkotte, T., Lépy, É., and Risvoll, C. (eds.) (2020). Supplementary Feeding in Reindeer Husbandry – Results from a Workshop with Reindeer Herders and Researchers from Norway, Sweden and Finland, Umeå University, Umeå.
Horwitz, L. K., Tchernov, E., Ducos, P., Becker, C., von den Driesch, A., Martin, L., and Garrard, A. (1999). Animal domestication in the southern Levant. Paléorient 25(2): 63–80.
Hörnberg, G., Josefsson, T., DeLuca, T. H., Higuera, P. E., Liedgren, L., Östlund, L., and Bergman, I. (2018). Anthropogenic use of fire led to degraded scots pine-lichen forest in northern Sweden. Anthropocene 24: 14–29.
Hull, E. H., and Salmi, A. (n.d). Activity reconstruction of Rangifer tarandus hooves in Fennoscandian archaeology: Methodological considerations and application to archaeological material from two Sámi habitation sites, Manuscript on file, Research Unit of History, Culture and Communication, University of Oulu, Oulu.
Hultblad, F. (1968). Övergång från nomadism till agrar bosättning i Jokkmokks socken, Almqvist and Wiksell, Stockholm.
Indrelid, S., and Hufthammer, A. K. (2011). Medieval mass trapping of reindeer at the Hardangervidda mountain plateau, south Norway. Quaternary International 238(1–2): 44–54.
Ingold, T. (1986). Reindeer economies and the origins of pastoralism. Anthropology Today 2(4): 5–10.
Ingold, T. (1993). The temporality of the landscape: Conceptions of time and ancient society. World Archaeology 25: 152–174.
Ingold, T. (1997). The picture is not a terrain: Maps, paintings and the dwelt-in world. Archaeological Dialogues 4(1): 29–31.
Ingold, T. (2000). Perception of the Environment: Essays on Livelihood, Dwelling and Skill, Routledge, London.
Ingold, T. (2011). Being Alive: Essays on Movement, Knowledge and Description, Routledge, London.
Istomin, K. V., and Dwyer, M. J. (2010). Dynamic mutual adaptation: Human-animal interaction in reindeer herding pastoralism. Human Ecology 38: 613–623.
Itkonen, T. I. (1948). Suomen lappalaiset vuoteen 1945, Söderström, Porvoo.
Jerand, P., and Linderholm, J. (2019). Possible traces of reindeer corralling at a hearth-row site from 1000–1300 CE in northern Norway. In Halinen, P., and Olsen, B. (eds.), In Search of Hearths: A Book in Memory of Sven-Donald Hedman, Iskos 22, Finnish Antiquarian Society, Helsinki, pp. 31–39.
Jordhøy, P. (2008). Ancient wild reindeer pitfall trapping systems as indicators for former migration patterns and habitat use in the Dovre region, southern Norway. Rangifer 28: 79 –87.
Kirksey, S. E., and Helmreich, S. (2010). The emergence of multispecies ethnography. Cultural Anthropology 25: 545–576.
Knight, J. (2012). The anonymity of the hunt: A critique of hunting as sharing. Current Anthropology 53: 334–355.
Kohler, T. A., Smith, M. E., Bogaard, A., Feinman, G. M., Peterson, C. E., Betzenhauser, A., et al. (2017). Greater post-Neolithic wealth disparities in Eurasia than in North America and Mesoamerica. Nature 551: 619–622.
Korhonen, T. (2008). Poroerotus: Historia, toiminta ja tekniset ratkaisut, Suomalaisen Kirjallisuuden Seura, Helsinki.
Korpijaakko, K. (1989). Saamelaisten oikeudellinen asema Ruotsi-Suomessa: Oikeushistoriallinen tutkimus Länsi-Pohjan Lapin maankäyttöoloista ja –oikeuksista ennen 1700-luvun puoltaväliä, Lakimiesliiton kustannus, Helsinki.
Kortesalmi, J. J. (2008). Poronhoidon synty ja kehitys Suomessa, Suomalaisen Kirjallisuuden Seura, Helsinki.
Krupnik, I. (1993). Arctic Adaptations: Native Whalers and Reindeer Herders of Northern Eurasia, University Press of New England for Dartmouth College, Hanover.
Kuokkanen, T. (2000). Stone Age sledges of central-grooved type: Finnish reconstructions. Fennoscandia Archaeologica XVI(I): 37–56.
Kuusela, J., Nurmi, R., and Hakamäki, V. (2016). Co-existence and colonisation: Re-assessing the settlement history of the pre-Christian Bothnian Bay coast. Norwegian Archaeological Review 49: 177–203.
Kuusela, J., Salmi, A., and Äikäs, T. (2020). Hunters, fishers, traders – An archaeological and zooarchaeological perspective on the development of the Late Iron Age and medieval northern Fennoscandian trade network. Estonian Journal of Archaeology 24: 141–160.
Kylli, R. (2012). Saamelaisten kaksi kääntymystä: Uskonnon muuttuminen Utsjoen ja Enontekiön lapinmailla 1602–1905, Suomalaisen Kirjallisuuden Seura, Helsinki.
Larson, G., and Fuller, D. Q. (2014). The evolution of animal domestication. Annual Review of Ecology, Evolution, and Systematics 66: 115–36.
Laufer, B. (1917). The Reindeer and Its Domestication, Corinthian Press, Lancaster.
Leem, K. (1956 [1767]). Beskrivelse over Finnmarkens Lapper, Halvorsen & Borsum, Oslo.
von Linné, C. (1969 [1732]). Carl von Linnés lappländska resa, Natur och Kultur, Stockholm.
Liu, X., Koba, K., Koyama, L. A., Hobbie, S. E., Weiss, M. S., Inagaki, Y., et al. (2018). Nitrate is an important nitrogen source for Arctic tundra plants. Proceedings of the National Academy of Sciences 115: 3398–3403.
Losey, R., Nomokonova. T., Arzyutov, D. V., Gusev, A. V., Plekhanov, A. V., Fedorova, N. V., and Anderson, D. G. (2021). Domestication as enskilment: Harnessing reindeer in Arctic Siberia. Journal of Archaeological Method and Theory 28: 197–231.
Lundmark, L. (1982). Uppbörd, utarmning, utveckling: Det samiska samhällets övergång till rennomadism i Lule lappmark, Arkiv för Studier i Arbetarrörelsens Historia, Lund.
Magnus, O. (1996 [1555]). A Description of the Northern Peoples, Ashgate Publishing, Farnham.
Manker, E. (1957). Lapparnas heliga ställen: Kultplatser och offerkult i belysning av nordiska museets och landsantikvariernas fältundersökningar, Acta Lapponica XIII, Gebers, Uppsala.
Marciniak, A. (2011). The secondary products revolution: Empirical evidence and its current zooarchaeological critique. Journal of World Prehistory 24: 117–130.
Mazzullo, N., and Ingold, T. (2008). Being along: Place, time and movement among Sámi people. In Bærenholdt, J., and Granås, B. (eds.), Mobility and Place: Enacting Northern European Peripheres, Ashgate, Burlington, pp. 27–38.
Mebius, H. (2003). Bissie. Studier i samisk religionshistoria, Jengel, Östersund.
Michelsen, A., Quarmby, C., Sleep, D., and Jonasson, S. (1998). Vascular plant 15N natural abundance in heath and forest tundra ecosystems is closely correlated with presence and type of mycorrhizal fungi in roots. Oecologia 115: 406–418.
Miller, A. V., and Makarewicz, C. (eds.) (2019). Isotopic Investigations of Pastoralism in Prehistory, Routledge, New York.
Mirov, N. T. (1945). Notes on the domestication of reindeer. American Anthropologist 47: 393–408.
Mulk, I. (1994). Sirkas – ett samiskt fångstsamhälle i förändring Kr. f. – 1600 e.Kr., Studia Archaeologica Universitatis Umensis 6, Umeå: Umeå universitet.
Murashkin, A. I., Kolpakov, E. M., Shumkin, V. Y., Khartanovich, V. I., and Moiseyev, V. G. (2016). Kola Oleneostrovskiy grave field: A unique burial site in the European Arctic. In Uino, P., and Nordqvist, K. (eds.), New Sites, New Methods: Proceedings of the Finnish-Russian Archaeological Symposium, Helsinki, 19–21 November 2014, Iskos 21, Finnish Antiquarian Society, Helsinki, pp. 186–199.
Näkkäläjärvi, K., and Pennanen, J. (2000). Poronhoito perustuu pohjoisen luonnon vuotuiskiertoon. In Pennanen, J., and Näkkäläjärvi, K. (eds.), Siiddastallan – Siidoista kyliin: Luontosidonnainen saamelaiskulttuuri ja sen muuttuminen, Pohjoinen, Oulu, pp. 76–79.
Nieminen, M., and Helle, T. (1980). Variations in body measurements of wild and semi-domestic reindeer (Rangifer tarandus) in Fennoscandia. Annales Zoologici Fennici 17: 275–283.
Nieminen, M., and Heiskari, U. (1989). Diets of freely grazing and captive reindeer during summer and winter. Rangifer 9: 17–34.
Niinimäki, S., Härkönen, L., Puolakka, H., van den Berg, M., and Salmi, A. (2021). Cross-sectional properties of reindeer long bones and metapodials allow identification of activity patterns. Archaeological and Anthropological Sciences 13: 146.
Niinimäki, S., and Salmi, A. (2016). Entheseal changes in free-ranging versus zoo reindeer – Observing activity status of reindeer. International Journal of Osteoarchaeology 26: 314–323.
Nomokonova, T., Losey, R. J., Fedorova, N. V., Gusev, A. V., and Arzyutov, D. V. (2021). Reindeer imagery in the making at Ust’-Polui in Arctic Siberia. Cambridge Archaeological Journal 31: 161–181.
Norstedt, G., Rautio, A., and Östlund, L. (2017). Fencing the forest: Early use of barrier fences in Sami reindeer husbandry. Rangifer 37: 69–92.
Núñez, M., Äikäs, T., Aspi, J., Eriksson, G., Heino, M., Oinonen, M., Okkonen, J., and Salmi, A. (2020). Animal remains from Saami offering places – Glimpses of human-animal relations from Finnish Lapland AD 1000–1900. In Spangen, M., Salmi, A., Äikäs, T., and Fjellström, M. (eds), Currents of Saami Pasts: Recent Advances in Saami Archaeology, Monographs of the Archaeological Society of Finland 9, Archaeological Society of Finland, Helsinki, pp. 61–78.
O'Connor, T. P. (1997). Working at relationships: Another look at animal domestication. Antiquity 71: 149–156.
Odner, K. (2001). Trade, tribute and household responses: The archaeological excavation at Geahèevájnjárga 244B in Varanger fjord, northern Norway. Acta Borealia 18: 25–50.
Olofsson, C. (2010). Making new antlers: Depositions of animal skulls and antlers as a message of regeneration in South Sámi grave contexts. Norwegian Archaeological Review 43: 97–114.
Olsen, B. (2019). Brodkorbneset and Steintjørna: Two hearth-row sites in Pasvik, Arctic Norway. In Halinen, P., and Olsen, B. (eds.), In Search of Hearths: A Book in Memory of Sven-Donald Hedman, Iskos 22, Finnish Antiquarian Society, Helsinki, pp. 9–30.
Oma, K. A. (2010). Between trust and domination: Social contracts between humans and animals. World Archaeology 42: 175–187.
Parker, K. L., Barboza, P. S., and Stephenson, T. R. (2005). Protein conservation in female caribou (Rangifer tarandus): Effects of decreasing diet quality during winter. Journal of Mammalogy 86: 610–622
Paulaharju, S. (1922). Lapin muisteluksia, Kustannusosakeyhtiö Kirja, Helsinki.
Pelletier, M., Kotiaho, A., Niinimäki, S., and Salmi, A. (2020). Identifying early stages of reindeer domestication in the archaeological record: A 3D morphological investigation on forelimb bones of modern populations from Fennoscandia. Archaeological and Anthropological Sciences 12: 169.
Pelletier, M., Kotiaho, A., Niinimäki, S., and Salmi, A. (2021a). Impact of selection and domestication on hindlimb bones of modern reindeer populations: Archaeological implications for early reindeer management by Sámi in Fennoscandia. Historical Biology 34: 802–820.
Pelletier, M., Niinimäki, S., and Salmi, A. (2021b). Influence of captivity and selection on limb long bone cross-sectional morphology of reindeer. Journal of Morphology 282: 1533–1556.
Pickard, C., Schoop, U. D., Bartosiewicz, L., Gillis, R., and Sayle, K. L. (2017). Animal keeping in Chalcolithic north-central Anatolia: What can stable isotope analysis add? Archaeological and Anthropological Sciences 9: 1349–1362.
Pilaar Birch, S. E. (ed.) (2018). Multispecies Archaeology, Routledge, London.
Politis, G. G. (2014). Ethnoarchaeology. In Smith, C. (ed.), Encyclopedia of Global Archaeology,Springer, New York, https://doi.org/10.1007/978-1-4419-0465-2_284.
Puputti, A., and Niskanen, M. (2009). Identification of semi-domesticated reindeer (Rangifer tarandus tarandus, Linnaeus 1758) and wild forest reindeer (R.t. fennicus, Lönnberg 1909) from postcranial skeletal measurements. Mammalian Biology 74: 49–58.
Rankama, T., and Ukkonen, P. (2001). On the early history of the wild reindeer (Rangifer tarandus L.) in Finland. Boreas 30: 131–147.
Rantamäki-Lahtinen, L. (ed.) (2008). Porotalouden taloudelliset menestystekijät, Maa- ja Elintarviketalouden Tutkimuskeskus, Helsinki.
Rheen, S. (1897 [1671]). En kortt Relation om Lapparnes Lefwarne och Sedher: Wijd-Skiepellsser; sampt I manga Stycken Grofwe wildfarellsser, Harald Wretman, Uppsala.
Roslund, Y. (1989). Den arkeologiska undersökningen. Silvret från Nasafjäll. In Awebro, K., Björkenstam, N., Norrman, J., Petersson, S., Sten, S., and Wallquist, E. (eds.), Silvret från Nasafjäll: arkeologi vid Silbojokk, Stockholm, Riksantikvarieämbetet, pp. 71–132.
Røed, K. H., Flagstad, Ø., Nieminen, M., Holand, Ø., Dwyer, M. J., Røv, N., and Vilà, C. (2008). Genetic analyses reveal independent domestication origins of Eurasian reindeer. Proceedings of the Royal Society B: Biological Sciences 275: 1849–1855.
Røed, K. H., Bjørklund, I., and Olsen, B. J. (2018). From wild to domestic reindeer – Genetic evidence of a non-native origin of reindeer pastoralism in northern Fennoscandia. Journal of Archaeological Science Reports 19: 279–286.
Røed, K. H., Bjørnstad, G., Flagstad, Ø., Haanes, H., Hufthammer, A. K, Jordhøy, P., and Rosvold, J. (2014). Ancient DNA reveals prehistoric habitat fragmentation and recent domestic introgression into native wild reindeer. Conservation Genetics 15: 1137–1149.
Røed, K. H., Kvie, K. S., Losey, R. J., Kosintsev, P. A., Hufthammer, A. K., Dwyer, M. J., et al. (2020). Temporal and structural genetic variation in reindeer (Rangifer tarandus) associated with the pastoral transition in northwestern Siberia. Ecology and Evolution 10: 9060–9072.
Salmi, A., Äikäs, T., Fjellström, M., and Spangen, M. (2015) Animal offerings at Sámi offering site Unna Saiva—Changing religious practices and human-animal relationships. Journal of Anthropological Archaeology 40: 10–22.
Salmi, A., Äikäs, T., and Lipkin, S. (2011). Animating rituals at Sámi sacred sites in northern Finland. Journal of Social Archaeology 11: 212–35.
Salmi, A., Äikäs, T., Spangen, M., Fjellström, M., and Mulk, I. (2018). Tradition and transformation in Sámi animal-offering practices. Antiquity 92: 472–489.
Salmi, A., van den Berg, M., Niinimäki, S., and Pelletier, M. (2021). Earliest archaeological evidence for domesticated reindeer economy among the Sámi of northeastern Fennoscandia AD 1300 onwards. Journal of Anthropological Archaeology 62: 101303.
Salmi, A., Fjellström, M., Äikäs, T., Spangen, M., Núñez, M., and Lidén, K. (2020a). Zooarchaeological and stable isotope evidence of Sámi reindeer offerings. Journal of Archaeological Science Reports 29: 102129.
Salmi, A., Fjellström, M., Niinimäki, S., Soppela, P., Kynkäänniemi, S., and Wallén, H. (2022). Human-reindeer relationships and reindeer feeding: Perspectives from archaeology and reindeer herders’ knowledge. In Salmi, A. (ed.), Domestication in Action: Human-Reindeer Interaction in Past and Present Reindeer Herding, Palgrave Macmillan, Cham, pp. 123–149.
Salmi, A., and Heino, M. T. (2019). Tangled worlds: The Swedish, the Sámi, and the reindeer. International Journal of Historical Archaeology 23: 260–282.
Salmi, A., and Niinimäki, S. (2021). Reindeer physical activity patterns and reconstruction of feeding behaviour: Implications for reindeer domestication and human-reindeer interaction. In Salmi, A., and Niinimäki, S. (eds.), Archaeologies of Animal Movement: Animals on the Move, Springer, New York, pp. 45–57.
Salmi, A., Niinimäki, S., and Pudas, T. (2020b). Identification of working reindeer using palaeopathology and entheseal changes. International Journal of Paleopathology 30: 57–67.
Sarkki, S., Komu, T., Heikkinen, H. I., Garcia, N. A., Lépy, É., and Herva, V. (2018). Applying a synthetic approach to the resilience of Finnish reindeer herding as a changing livelihood. Ecology and Society 21(4): 14.
Schefferus, J. (1979 [1674]). Lapponia, Arvi A. Karisto Osakeyhtiö, Hämeenlinna.
Seitsonen, O. (2020). Ruttoa ylätunturissa? Saamelaisten poronhoitokohteiden ajoittuminen Enontekiön Yliperällä. Suomen Keskiajan Arkeologinen Seura 2/2020: 2–20.
Seitsonen, O, and Viljanmaa, S. (2021). Transnational landscapes of Sámi reindeer: Domestication and herding in northernmost Europe 700–1800 AD. Journal of Field Archaeology 46: 172–191.
Serning, I. (1956). Lapska offerplatsfynd från järnålder och medeltid i de svenska lappmarkena, Almqvist and Wiksell, Uppsala.
Sherratt, A. G. (1981). Plough and pastoralism: Aspects of the secondary products revolution. In Hodder, I., Isaac, G., and Hammond, N. (eds.), Pattern of the Past, Cambridge University Press, Cambridge, pp. 261–306.
Skrzypek, G., Wojtuń, B., Richter, D., Jakubas, D., Wojczulanis-Jakubas, K., and Samecka-Cymerman, A. (2015). Diversification of nitrogen sources in various tundra vegetation types in the high Arctic. PLoS One 10: e0136536.
Slunga, N. (1993). Kirkko ja koulu – Papisto ja kirkollinen elämä. In Hederyd, O., and Alamäki, Y. (eds.), Tornionlaakson historia II. 1600-luvulta vuoteen 1809, Tornionlaakson Kuntien Historiakirjatoimikunta, s.l., pp. 280–313.
Smith, L. T. (2012). Decolonizing Methodologies: Research and Indigenous Peoples, 2nd ed., Zed Books, London.
Soppela, P, Kynkäänniemi, S., and Wallen, H. (2020). Poron käyttö ajokkaana – Perinne jatkuu matkailussa. Poromies 4/2020: 44–47.
Soppela, P., Saarela, S., Heiskari, U., and Nieminen, M. (2008). The effects of wintertime undernutrition on plasma leptin and insulin levels in an arctic ruminant, the reindeer. Comparative Biochemistry and Physiology - B Biochemistry and Molecular Biology 149: 613–621.
Spangen, M., Salmi, A., Äikäs, T., and Fjellström, M. (2020). Introduction: Currents of Saami pasts. In Spangen, M., Salmi, A., Äikäs, T., and Fjellström, M. (eds.), Currents of Saami pasts: Recent Advances in Saami Archaeology, Monographs of the Archaeological Society of Finland 9, Archaeological Society of Finland, Helsinki, pp. 2–15.
Sten, S. (1989). Husdjurshållning, jakt och fiske i Silbojokk – En osteologisk analys av djurbenen. In Awebro, K., Björkenstam, N., Norrman, J., Peterson, S., Roslund, Y., Sten, S., and Wallquist, E. (eds.), Silvret från Nasafjäll: Arkeologi vid Silbojokk, Riksantikvarieämbetet, Stockholm, pp. 167–178.
Stépanoff, C. (2017). The rise of reindeer pastoralism in northern Eurasia: Human and animal motivations entangled. Journal of the Royal Anthropological Institute 23: 376–397.
Storli, I. (1996). On the historiography of Sami reindeer pastoralism. Acta Borealia 13: 81–115.
Stump, D. (2013). On applied archaeology, Indigenous knowledge, and the usable past. Current Anthropology 54: 268–298.
Svestad, A. (2018). Entering other realms: Sámi burials in natural rock cavities and caves in northern Fenno-Scandinavia between 900 BC and AD 1700. In Bergsvik, K.A., and Dowd, M. (eds.), Caves and Ritual in Medieval Europe, AD 500–1500, Oxbow Books, Oxford, pp. 13–31.
Takken Beijersbergen, L. M., Fernandes, R., Mørkvede, P. T., and Hufthammer, A. K. (2021). Temporal and spatial variability of bone collagen stable carbon and nitrogen isotopic ratios of Norwegian reindeer. Journal of Archaeological Science: Reports 37: 102890.
Tegengren, H. (1952). En utdöd Lappkultur i Kemi Lappmark: Studier i Nordfinlands kolonisationshistoria, Åbo Akademi, Åbo.
Tornaeus, J. (1900 [1672]). Berättelse om Lapmarckerna och Deras Tillstånd, K. B. Wiklund (ed.), Harald Wretmans tryckeri, Uppsala.
Turunen, M, and Vuojala-Magga, T. (2014). Past and present winter feeding of reindeer in Finland: Herders’ adaptive learning of feeding practices. Arctic 67: 173–188.
Vigne, J. (2011). The origins of animal domestication and husbandry: A major change in the history of humanity and the biosphere. Comptes Rendus Biologies 334: 171–181.
Virrankoski, P. (1973). Pohjois-Pohjanmaan ja Lapin historia III. Pohjois-Pohjanmaa ja Lappi 1600-luvulla, Pohjois-Pohjanmaan ja Lapin Maakuntaliiton Yhteinen Historiatoimikunta, Oulu.
Vorren, Ø. (1974). Gál'gojáv're-området - Et demonstrasjonsfelt for samisk kulturutvikling. Ottar 81: 5–11.
Vretemark, M. (2019). The faunal remains from two hearth row sites in Pasvik, arctic Norway. In Halinen, P., and Olsen, B. (eds.), In Search of Hearths: A Book in Memory of Sven-Donald Hedman, Iskos 22, Finnish Antiquarian Society, Helsinki, pp. 67–74.
Vuojala-Magga, T. (2010). Knowing, training, learning: The importance of reindeer character and temperament for individuals and communities of humans and animals. In Stammler, F., and Takakura, H. (eds.), Good to Eat, Good to Live with: Nomads and Animals in Northern Eurasia and Africa, Center for Northeast Asian Studies, Tohoku University, Sendai, pp. 43–61.
Wallerström, T. (1983). Kulturkontakter i Norrbottens kustland under medeltiden. Norrbotten 82–83: 16–55.
Wallerström, T. (2000). The Saami between east and west in the Middle Ages: An archaeological contribution to the history of reindeer breeding. Acta Borealia 17: 3–39.
Weinstock, J. (1997). The relationship between body size and environment: The case of Late Pleistocene reindeer (Rangifer tarandus). Archaeofauna 6: 123–135.
Weladji, R. B., and Holand, Ø. (2006). Influences of largescale climatic variability on reindeer population dynamics: Implications for reindeer husbandry in Norway. Climate Research 32: 119–127.
Weldenegodguad, M., Pokharel, K., Ming, Y., Honkatukia, M., Peippo, J., Reilas, T., Røed K. H., and Kantanen, J. (2020). Genome sequence and comparative analysis of reindeer (Rangifer tarandus) in northern Eurasia. Scientific Reports 10: 8980.
Wilkins, A. S., Wrangham, R. W., and Fitch, W. T. (2014). The "domestication syndrome" in mammals: A unified explanation based on neural crest cell behavior and genetics. Genetics 197: 795–808.
Ylimaunu, T. (2007). Aittakylästä kaupungiksi – Arkeologinen tutkimus Tornion kaupungistumisesta 18. vuosisadan loppuun mennessä, Pohjois-Suomen Historiallinen Yhdistys, Rovaniemi.
Zachrisson, I. (1984). De samiska metalldepåerna år 1000–1350 i ljuset av fyndet från Mörtträsket, Lappland, Umeå Universitet, Umeå.
Zachrisson, I. (1997). Möten i gränsland: Samer och germaner i Mellanskandinavien, Statens Historiska Museum, Stockholm.
Zeder, M. A. (2012). The domestication of animals. Journal of Anthropological Research 68: 161–190.
Zeder, M. A. (2015). Core questions in domestication research. Proceedings of the National Academy of Sciences 112: 3191–3198.
Zetterberg, P., Eronen, M., and Briffa, K. R. (1994). Evidence on climatic variability and prehistoric human activities between 165 BC and AD 1400 derived from subfossil Scots Pines (Pinus sylvestris L.) found in a Lake in Utsjoki, northernmost Finland. Bulletin of the Geological Society of Finland 66(2): 107–124.
Zohary, D., Tchernov, E., and Kolska Horwitz, L. (1998). The role of unconscious selection in the domestication of sheep and goats. Journal of Zoology 245: 129–135.
Bibliography of Recent Literature
Andersen, O. (2014). Stone walls as a means of understanding the different types of reindeer herding: A study from the Lule Sámi area on the Norwegian side of the border. Arctic Anthropology 51(2): 23–34.
Äikäs, T. (2015). From Boulders to Fells. Sacred Places in the Sámi Ritual Landscape, Monographs of the Archaeological Society of Finland 5, Archaeological Society of Finland, Helsinki.
Hull, E. H., Niinimäki, S., and Salmi, A. (2020). Differences in entheseal changes in the phalanges between ecotypes of Fennoscandian reindeer. International Journal of Osteoarchaeology 30: 666–687.
Losey, R. J. (2021). Domestication is not an ancient moment of selection for prosociality: Insights from dogs and modern humans. Journal of Social Archaeology https://doi.org/https://doi.org/10.1177/14696053211055475
Salmi, A., and Niinimäki, S. (2016). Entheseal changes and pathological lesions in draught reindeer skeletons – Four case studies from present-day Siberia. International Journal of Paleopathology 14: 91–99.
Seitsonen, O., and Égüez, N. (2022). Here be reindeer: Geoarchaeological approaches to the transspecies lifeworlds of the Sámi reindeer herder camps on the tundra. In Halinen, P. (ed.), Materiality and Objects: Multi-disciplinary Approaches to Archaeological Material and Contexts, Iskos 24, Finnish Antiquarian Society, Helsinki, pp. 124–145.
Sommerseth, I. (2011). Archaeology and the debate on the transition from reindeer hunting to pastoralism. Rangifer 31: 111–127.
Sommerseth, I. (2015). New traces of wild reindeer hunting in the alpine areas in northern Norway. In Indrelid, S., Hjelle, K. L., and Stene, K. (eds.), Exploitation of Outfield Resources – Joint Research at the University Museums of Norway, Universitetsmuseet i Bergen Skrifter No. 32, University Museum of Bergen, Bergen, pp. 19–29.
van den Berg, M., Wallén, H., and Salmi, A. (in press). The identification of castrated reindeer (Rangifer tarandus) bones and the significance of castration to trace human-animal relationships in the north. Archaeological and Anthropological Sciences.
Acknowledgments
This project received funding from the European Research Council (ERC) under the European Union’s Horizon 2020 research and innovation program (grant agreement No. 756431) and the Academy of Finland (project numbers 275635 and 308322). I wish to thank all the researchers working in the Domestication in Action project whose work has contributed to the results presented and cited in this article.
Funding
Open Access funding provided by University of Oulu including Oulu University Hospital.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Salmi, AK. The Archaeology of Reindeer Domestication and Herding Practices in Northern Fennoscandia. J Archaeol Res 31, 617–660 (2023). https://doi.org/10.1007/s10814-022-09182-8
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10814-022-09182-8