
Abstract
Appropriate modulation of imitation according to social context is important for successful social interaction. In the present study we subliminally primed high-functioning adults with ASC and age- and IQ-matched controls with either a pro- or non- social attitude. Following priming, an automatic imitation paradigm was used to acquire an index of imitation. Whereas imitation levels were higher for pro-socially primed relative to non-socially primed control participants, there was no difference between pro- and non- socially primed individuals with ASC. We conclude that high-functioning adults with ASC demonstrate atypical social modulation of imitation. Given the importance of imitation in social interaction we speculate that difficulties with the modulation of imitation may contribute to the social problems characteristic of ASC.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Imitation (also known as ‘mimicry’) is intricately linked with social interaction. Being imitated increases rapport (Chartrand and Bargh 1999), altruistic behavior (van Baaren et al. 2004) and trust (Bailenson and Yee 2005). Furthermore, individuals imitate more when in possession of a positive social attitude (Lakin and Chartrand 2003; Leighton et al. 2010). For example, subliminal pro-social, compared to non-social, priming results in significantly higher levels of imitation (Cook and Bird 2011; Leighton et al. 2010). Thus, imitation is bi-directionally associated with good social interaction and is therefore a key component in building social relationships with others. Crucially, successful social interaction relies on appropriate modulation of the degree of imitation according to the demands of the social situation (Lakin and Chartrand 2003).
Autism Spectrum Conditions (ASC) are characterised by impairments in social interaction, language, and communication (American Psychiatric Association 1994). A number of studies have demonstrated reduced imitation and Mirror Neuron System (MNS) activity in individuals with ASC compared to control participants (Williams et al. 2004). The MNS is a network of brain areas active when an individual both executes and observes an action (Catmur et al. 2008; Iacoboni et al. 1999) and has been argued to comprise the neural mechanism that underpins imitation (Catmur et al. 2007; Heiser et al. 2003; Iacoboni et al. 1999). It has been hypothesised that a ‘broken MNS’ and corresponding imitation impairment is a core feature of ASC (Williams et al. 2001). However, experimental evidence both supports (Avikainen et al. 2003; Dapretto et al. 2006; McIntosh et al. 2006; Oberman et al. 2005; Rogers et al. 2003) and opposes (Bird et al. 2007; Dinstein et al. 2010; Gowen et al. 2008; Hamilton et al. 2007; Leighton et al. 2008; Press et al. 2010; Spengler et al. 2010) the presence of an imitation impairment in ASC. Furthermore, clinical observations of high levels of echolalia (automatic repetition of speech patterns) and echopraxia (automatic imitation of observed actions) in individuals with ASC (Russell 1997; Rutter 1974; Williams et al. 2004) are incompatible with an imitation deficit in ASC, and instead suggest problems with inhibition of imitation.
In response to the inconsistent literature it has been suggested that, rather than an imitation deficit per se, individuals with ASC may have difficulties with appropriately modulating levels of imitation (Hamilton 2008; Kana et al. 2011; Spengler et al. 2010). Although this hypothesis has not previously been tested, it is consistent with studies of individuals with ASC that report hypoactivity in parts of the brain thought to be involved in the modulation of imitation (Castelli et al. 2000; Spengler et al. 2010). Given the importance of appropriate levels of imitation for positive social interaction (Lakin and Chartrand 2003) this hypothesis may go some way towards explaining difficulties with social interaction in individuals with ASC.
The present study used a behavioural measure of imitation, as opposed to a measure of MNS activity, to directly test the hypothesis that the social modulation of imitation is atypical in individuals with ASC. High-functioning adults with ASC and age and IQ-matched controls first completed a previously-validated (Bargh and Chartrand 2000; Cook and Bird 2011; Leighton et al. 2010) technique to unconsciously prime either a pro-social, or non-social attitude. Participants were asked to ‘unscramble’ re-arranged sentences, a proportion of which were related to either pro-social attitudes (“she is my friend”) or non-social attitudes (“he is often alone”). Following this subliminal priming, participants completed an automatic imitation task. We predicted that, as in previous studies (Cook and Bird 2011; Leighton et al. 2010), pro-socially primed control participants would show increased levels of imitation relative to non-socially primed control participants. In line with the impaired modulation of imitation in ASC hypothesis, we predicted no significant difference in levels of imitation for pro-socially and non-socially primed ASC groups.
Methods
Participants
19 adults (mean 40.9 years) with ASC and 22 age- and IQ-matched control individuals participated in this experiment (see Table 1 for further details). All participants had normal or corrected-to-normal vision and were screened for exclusion criteria (dyslexia, epilepsy, and any other neurological or psychiatric conditions) prior to taking part. Participants with ASC had a written diagnosis from an independent clinician, which they received no more than 5 years before taking part in this experiment, and all participants (save one for whom data was not available) scored above threshold for Autism Spectrum Disorder on the Autism Diagnostic Observation Schedule (ADOS; Lord et al. 2000). We were unable to distinguish between participants with Asperger Syndrome and Autism, as we did not have information about early development of language in our participants. Participants were randomly assigned to either the Pro- or Non-social Prime Group. ANOVAs demonstrated no main effect of, or interaction between, Prime Group and Diagnostic Group on age or full scale IQ (all ps > 0.1). The two ASC groups did not differ with respect to ADOS score (non-social mean (SEM) = 10.00 (1.00), pro-social mean (SEM) = 9.88 (1.01), t(16) = 0.08, p = 0.93), age (non-social mean (SEM) = 41.30 (3.84), pro-social mean (SEM) = 40.56 (4.50), t(17) = 0.13, p = 0.90), full scale IQ (non-social mean (SEM) = 114.44 (4.99), pro-social mean (SEM) = 111.44 (6.11), t(16) = 0.38, p = 0.70), verbal IQ (non-social mean (SEM) = 116.22 (3.33), pro-social mean (SEM) = 112.00 (5.92), t(16) = 0.62, p = 0.54) or performance IQ (non-social mean (SEM) = 108.89 (6.57), pro-social mean (SEM) = 108.78 (6.48), t(16) = 0.12, p = 0.99). Informed consent was obtained from all participants. The study was approved by the local ethics committee, and performed in accordance with the 1964 Declaration of Helsinki.
Priming Task
Participants were asked to select, in order, 4 words from 5 displayed on a computer screen to make a grammatically correct sentence (see Leighton et al. 2010). 24 out of 36 total trials contained a word semantically related to the target attitude (pro-social or non-social; Table 2). Priming words were generated in a pilot session by an independent group of participants. Although they were not identical they overlapped with those employed by Leighton et al. (2010).
Automatic Imitation Task
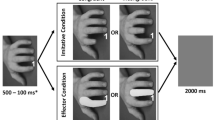
The Automatic Imitation Task was based on that used by Iacoboni et al. (1999) and Brass et al. (2000); for further details see Cook and Bird (2011). Videos (6° visual angle vertically × 9° horizontally, 3,000 ms duration) of a human hand were presented vertically on a computer screen. The participant rested their hand in a horizontal orientation on the computer keyboard, with their index finger holding down the ‘V’ key and their middle finger holding down the ‘B’ key. The participant was required to lift and replace their index or middle finger upon the appearance of a 1 or a 2, respectively. 50% of trials comprised a five-frame video clip of a concurrent lifting action that was either compatible (e.g. the participant was required to make an index finger response and observed an index finger action) or incompatible (e.g. the participant was required to make an index finger response and observed a middle-finger action) with the required movement (Fig. 1). Imitation was calculated as the difference in reaction time (RT) on congruent and incongruent trials. 50% of trials comprised a three-frame ‘baseline’ video clip in which the fingers remained static and either the compatible or incompatible finger acquired a green mask; enabling acquisition of baseline RTs for index and middle finger movements independent of imitation. 120 trials were presented in pseudo-random order. There were no breaks during the task, the duration of which was approximately 15 min.

a The five-frame action video clip. Frame one was displayed for a variable interval (range: 800–2,400 ms). Frames two and three were displayed for 34 ms each and frame four for 500 ms. These display durations ensured the appearance of a short video clip. The fifth frame (a black screen) remained on screen until the duration of the trial had reached 3,000 ms and the participant had returned both fingers to the letters V and B on the keyboard. b The three frames of a ‘baseline’ trial. Frame one was displayed for a variable interval. Frame two was displayed for 568 ms and the final frame was displayed until the duration of the trial had reached 3,000 ms and the participant had returned both fingers to the letters V and B
Before data acquisition participants received standardized instructions, completed 5 practice trials of the Priming Task, and made 5 correct consecutive responses [p (chance) < 0.05] on the Automatic Imitation Task. Finally participants completed a debriefing questionnaire (Leighton et al. 2010).
Results
Priming Task
Errors were infrequent on the Priming Task (mean error rate was l0%). A 2 × 2 ANOVA on Priming Task error data showed no main effect of Diagnostic Group or Prime Group and no interaction (all F(1,37) < 1, all p > 0.40; see Table 2 for mean and SEM values).
Automatic Imitation Task
Automatic Imitation Task error-trials and reaction times (RTs) < 150 ms and > 2,000 ms were removed from the analysis. Paired-samples t tests demonstrated that all four groups exhibited significant imitation, i.e. RTs on Compatible trials were significantly faster than on Incompatible trials (all ps < 0.01, see Table 2). A 2 × 2 ANOVA on the magnitude of Automatic Imitation shown on this task (RT on Incompatible Trials minus RT on Compatible Trials, see Fig. 2) showed an interaction between Diagnostic- (ASC, Control) and Prime Group (Non-social, Pro-social), F(1,37) = 2.92, p = 0.048 (1-tailed), η 2 p = 0.07). Simple effects analyses demonstrated that the effect of social priming was clearly shown in the Control Group: participants who were unconsciously primed with a pro-social attitude imitated more [mean (SEM) = 82.51 ms (22.20)] than those primed with a non-social attitude (35.42 ms (10.18); F(1,37) = 5.87, p = 0.02 η 2 p = 0.14). However, the ASC Group showed no such social modulation of their automatic imitative behaviour: the degree of imitation shown by the pro-socially primed participants with ASC [34.83 ms (8.76)] was not different from that shown by the non-socially primed participants with ASC (36.61 ms (8.01); F(1,37) = 0.007, p = 0.93, η 2 p = 0.00). In addition, simple effects analysis showed that the degree of imitation shown by the Control Pro-Social Group was significantly greater than that shown by the ASC Pro-social Group [F(1,37) = 5.42, p = 0.03, η 2 p = 0.13]. In contrast, the Control and ASC Non-social Groups did not differ [F(1,37) = 0.004, p = 0.95, η 2 p = 0.00].

Pro-socially primed participants in the Control Group imitated more than non-socially primed participants. Participants with ASC showed no such social modulation of imitation: the degree of imitation shown by the ASC Pro-social Group did not differ from that shown by the ASC Non-social Group. Furthermore the Control Pro-Social Group showed significantly greater imitation than the ASC Pro-social Group. In contrast, the Control and ASC Non-social Groups did not differ. *Indicates p < 0.05
Previous research has shown that the magnitude of imitation is modulated by mean RT (Press et al. 2005). A 2 × 2 ANOVA on baseline trial RT (mean RT across all baseline trials—i.e. trials in which the finger remained static and acquired a green mask) showed no significant effects of Prime Group, Diagnostic Group nor interaction (all ps > 0.1, see Table 2). A 2 × 2 ANOVA on incongruent (e.g. participant lifted index finder and middle finger acquired green mask) minus congruent (e.g. participant lifted index finder and index finger acquired green mask) baseline trial RTs showed no significant effects of Prime Group, Diagnostic Group nor interaction (all ps > 0.1). Therefore, we are confident that the interaction between Prime Group and Diagnostic Group on the magnitude of imitation is not a product of mean RT differences.
Debriefing Questionnaire
Examination of the debriefing questionnaire data indicated that no participant correctly guessed the purpose of either the Priming or Automatic Imitation Task. Furthermore, no participant correctly identified a link between the tasks or a theme among the words presented in the Priming Task. Therefore, we can conclude that no participant was aware of the type of priming they had received or that the purpose of the study was to examine imitation and its relationship with social attitudes.
Discussion
In agreement with previous studies (Cook and Bird 2011; Leighton et al. 2010) we found that control participants primed with words promoting pro-social attitudes (e.g. friend, crowded, team, talkative) showed significantly higher levels of imitation than control participants primed with words promoting non-social attitudes (e.g. himself, solo, one, private). There was no significant difference between imitation levels shown by individuals with ASC primed with pro-social words compared with those primed with non-social words. These results comprise the first experimental evidence of atypical social modulation of imitation in individuals with ASC. The ability to appropriately modulate levels of imitation to suit the social situation is important in social interactions (Lakin and Chartrand 2003) hence we speculate that difficulties with the modulation of imitation may contribute to the social problems characteristic of ASC. In addition, this finding suggests that future studies of imitation and MNS function in ASC should consider the extent to which the task includes cues that may act as unconscious social primes. Efforts should be made to either eliminate these cues (and therefore investigate un-modulated MNS function), or include social cues as a factor in the experimental design.
Although no previous studies have directly tested the hypothesis that social modulation of imitation is atypical in ASC, Oberman et al. (2008) investigated MNS activity whilst participants observed hand actions conducted by a familiar (self or parent) and unfamiliar (stranger) other. Oberman et al. (2008) measured mu wave suppression as an indirect measure of MNS activity. They demonstrated that children with ASC showed significantly less mu wave suppression compared to typically-developing children whilst observing actions performed by the unfamiliar actor. However, when observing actions performed by the familiar actor there was no difference between the groups. If familiarity is considered a social prime these results may be interpreted as evidence of social modulation of the MNS in ASC and therefore incompatible with the results of the present experiment. However, different stimuli were used in familiar and unfamiliar conditions in the study by Oberman and colleagues. Therefore, rather than being a product of social modulation, the results may be a consequence of stimulus-specific characteristics. For example, observation of familiar and unfamiliar actors may prompt differing levels of attention or motivation. In the present study identical automatic imitation paradigms were employed for both pro-social and non-social groups and therefore there were no differences in stimulus characteristics that might constitute different bottom-up signals for attentional engagement or motivation.
Although there is no bottom-up role for attention in our results, it is possible that attention may play a ‘top-down’ role in our observed effect: pro-social priming may increase imitation by enhancing attention to biological stimuli. However, as we have previously argued (Cook and Bird 2011), there is little evidence to support this hypothesis in the context of this paradigm. For both the pro-social and non-social groups, the imitation paradigm required attention to the same part of the screen as the movement stimuli (i.e. the cue to move was presented equidistant from the index and middle fingers of the video hand); any trials in which participants did not attend were detected by checking for incorrect responses, and for abnormally long or short reaction times, and these trials were excluded from the analysis. Furthermore, using this same paradigm we have previously reported evidence (Cook and Bird 2011) that the distribution of reaction times does not fit with a model of social priming affecting attentional process (where pro-social and non-social groups should differ even for the fastest responses). Rather, the distribution of reaction times fits a model of social priming affecting an inhibitory process (where pro-social and non-social groups need not necessarily differ for the fastest responses but should differ for the slowest responses). Therefore, it is likely that, rather than attentional mechanisms, social priming affects the inhibition of imitative responses; that is the pro-social group, compared to the non-social group are less likely to inhibit automatic imitative responses.
Why might the effect of social priming on the inhibition of imitative responses differ between control participants and those with ASC? A recent set of studies suggests that the control of imitation relies on social cognitive processes for distinguishing one’s own actions from the actions of another individual (Brass et al. 2005, 2003; Spengler et al. 2010). These social cognitive processes and imitation-inhibition both elicit activity in medial prefrontal cortex (MPFC) and temporo-parietal junction (TPJ): key nodes in the social brain network (Brothers 1990; Frith 2007; Frith and Frith 2010). Spengler et al. (2010) recently showed that, in individuals with ASC, low levels of MPFC and TPJ activity during a mentalising task were associated with poor imitation-inhibition. Although the neural correlates of pro-social priming have not been elucidated and it is not clear that imitation-enhancement and imitation-inhibition depend on overlapping brain areas, the work of Spengler and Brass suggests a testable hypothesis for future investigation: compared to control participants, individuals with ASC have a reduced social brain response to pro-social primes and hence exhibit atypical modulation of imitation. This hypothesis bears similarities with the ‘social relevance hypothesis’ proposed by Oberman et al. (2008) which suggests that, compared to typically-developing individuals, those with ASC require stimuli with greater social relevance in order to elicit comparable levels of MNS activity.
It is also possible that atypical modulation of imitation following pro-social priming is an instance of a more general failure of top-down modulation in individuals with ASC (Frith 2003). Studies of functional connectivity using magnetic resonance imaging report both greater and lesser connectivity between frontal and posterior areas in individuals with ASC compared to control participants. Bird et al. (2006) reported a reduced top-down influence of attention on face processing. Similarly, Kana et al. (2009) demonstrated underconnectivity between frontal and posterior regions during a mentalising task. More recently, greater task-independent connectivity between prefrontal cortex (PFC) and MNS regions has been reported in individuals with ASC compared to controls (Shih et al. 2010). Accordingly, atypical functional connectivity between brain areas that underpin the modulation of imitation (e.g. PFC) and those that underpin imitation itself (e.g. MNS) may be responsible for the impaired social modulation of imitation evidenced in the present study.
Conclusion
The present study found that control participants primed with words promoting pro-social attitudes showed significantly higher levels of imitation than control participants primed with words promoting non-social attitudes: this difference between pro- and non-social groups was absent for individuals with ASC. These results comprise the first demonstration of atypical social modulation of imitation in individuals with ASC. The ability to appropriately modulate levels of imitation to suit the social situation is important in social interactions hence this finding may help to explain some of the social problems characteristic of ASC.
References
American Psychiatric Association. (1994). Diagnostic and statistical manual of mental disorders. Washington, DC: American Psychiatric Association.
Avikainen, S., Wohlschläger, A., Liuhanen, S., Hänninen, R., & Hari, R. (2003). Impaired mirror-image imitation in Asperger and high-functioning autistic subjects. Current Biology, 13(4), 339–341.
Bailenson, J. N., & Yee, N. (2005). Digital chameleons: Automatic assimilation of nonverbal gestures in immersive virtual environments. Psychological Science, 16(10), 814–819.
Bargh, J. A., & Chartrand, T. L. (2000). The mind in the middle: A practical guide to priming and automaticity research. In H. T. Reis & C. M. Judd (Eds.), Handbook of research methods in social and personality psychology (pp. 253–285). New York: Cambridge University Press.
Bird, G., Catmur, C., Silani, G., Frith, C., & Frith, U. (2006). Attention does not modulate neural responses to social stimuli in autism spectrum disorders. NeuroImage, 31(4), 1614–1624.
Bird, G., Leighton, J., Press, C., & Heyes, C. (2007). Intact automatic imitation of human and robot actions in autism spectrum disorders. Proceedings of Biological Sciences/The Royal Society, 274(1628), 3027–3031.
Brass, M., Bekkering, H., Wohlschläger, A., & Prinz, W. (2000). Compatibility between observed and executed finger movements: comparing symbolic, spatial, and imitative cues. Brain and Cognition, 44(2), 124–143.
Brass, M., Derrfuss, J., Matthes-von Cramon, G., & von Cramon, D. Y. (2003). Imitative response tendencies in patients with frontal brain lesions. Neuropsychology, 17(2), 265–271.
Brass, M., Derrfuss, J., & von Cramon, D. Y. (2005). The inhibition of imitative and overlearned responses: a functional double dissociation. Neuropsychologia, 43(1), 89–98.
Brothers, L. (1990). The social brain: A project fr integrating primate behaviour and neurophysiology in a new domain. Concepts in Neuroscience, 1, 27–51.
Castelli, F., Happé, F., Frith, U., & Frith, C. (2000). Movement and mind: a functional imaging study of perception and interpretation of complex intentional movement patterns. NeuroImage, 12(3), 314–325.
Catmur, C., Gillmeister, H., Bird, G., Liepelt, R., Brass, M., & Heyes, C. (2008). Through the looking glass: counter-mirror activation following incompatible sensorimotor learning. The European Journal of Neuroscience, 28(6), 1208–1215.
Catmur, C., Walsh, V., & Heyes, C. (2007). Sensorimotor learning configures the human mirror system. Current Biology, 17(17), 1527–1531.
Chartrand, T. L., & Bargh, J. A. (1999). The chameleon effect: the perception-behavior link and social interaction. Journal of Personality and Social Psychology, 76(6), 893–910.
Cook, J., & Bird, G. (2011). Social attitudes differentially modulate imitation in adolescents and adults. Experimental Brain Research: Special Issue on Joint Action, 211(3–4), 601–612.
Dapretto, M., Davies, M. S., Pfeifer, J. H., Scott, A. A., Sigman, M., Bookheimer, S. Y., et al. (2006). Understanding emotions in others: mirror neuron dysfunction in children with autism spectrum disorders. Nature Neuroscience, 9(1), 28–30.
Dinstein, I., Thomas, C., Humphreys, K., Minshew, N., Behrmann, M., & Heeger, D. J. (2010). Normal movement selectivity in autism. Neuron, 66(3), 461–469.
Frith, C. D. (2003). What do imaging studies tell us about the neural basis of autism? Novartis Foundational Symposium, 251, 149–166.
Frith, C. D. (2007). The social brain? Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences, 362(1480), 671–678.
Frith, U., & Frith, C. (2010). The social brain: allowing humans to boldly go where no other species has been. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences, 365(1537), 165–176.
Gowen, E., Stanley, J., & Miall, R. C. (2008). Movement interference in autism-spectrum disorder. Neuropsychologia, 46(4), 1060–1068.
Hamilton, A. F. D. C. (2008). Emulation and mimicry for social interaction: a theoretical approach to imitation in autism. Quarterly Journal of Experimental Psychology, 61(1), 101–115.
Hamilton, A. F. D. C., Brindley, R. M., & Frith, U. (2007). Imitation and action understanding in autistic spectrum disorders: how valid is the hypothesis of a deficit in the mirror neuron system? Neuropsychologia, 45(8), 1859–1868.
Heiser, M., Iacoboni, M., Maeda, F., Marcus, J., & Mazziotta, J. C. (2003). The essential role of Broca’s area in imitation. The European Journal of Neuroscience, 17(5), 1123–1128.
Iacoboni, M., Woods, R. P., Brass, M., Bekkering, H., Mazziotta, J. C., & Rizzolatti, G. (1999). Cortical mechanisms of human imitation. Science, 286(5449), 2526–2528.
Kana, R. K., Keller, T. A., Cherkassky, V. L., Minshew, N. J., & Just, M. A. (2009). Atypical frontal-posterior synchronization of theory of mind regions in autism during mental state attribution. Social Neuroscience, 4(2), 135–152.
Kana, R. K., Wadsworth, H. M., & Travers, B. G. (2011). A systems level analysis of the mirror neuron hypothesis and imitation impairments in autism spectrum disorders. Neuroscience and Biobehavioral Reviews, 35(3), 894–902.
Lakin, J. L., & Chartrand, T. L. (2003). Using nonconscious behavioral mimicry to create affiliation and rapport. Psychological Science, 14(4), 334–339.
Leighton, J., Bird, G., Charman, T., & Heyes, C. (2008). Weak imitative performance is not due to a functional ‘mirroring’ deficit in adults with autism spectrum disorders. Neuropsychologia, 46(4), 1041–1049.
Leighton, J., Bird, G., Orsini, C., & Heyes, C. (2010). Social attitudes modulate automatic imitation. Journal of Experimental Social Psychology, 46(6), 905–910.
Lord, C., Risi, S., Lambrecht, L., Cook, E. H., Leventhal, B. L., DiLavore, P. C., et al. (2000). The autism diagnostic observation schedule-generic: A standard measure of social and communication deficits associated with the spectrum of autism. Journal of Autism and Developmental Disorders, 30(3), 205–223.
McIntosh, D. N., Reichmann-Decker, A., Winkielman, P., & Wilbarger, J. L. (2006). When the social mirror breaks: deficits in automatic, but not voluntary, mimicry of emotional facial expressions in autism. Developmental Science, 9(3), 295–302.
Oberman, L. M., Hubbard, E. M., McCleery, J. P., Altschuler, E. L., Ramachandran, V. S., & Pineda, J. A. (2005). EEG evidence for mirror neuron dysfunction in autism spectrum disorders. Brain Research. Cognitive Brain Research, 24(2), 190–198.
Oberman, L. M., Ramachandran, V. S., & Pineda, J. A. (2008). Modulation of mu suppression in children with autism spectrum disorders in response to familiar or unfamiliar stimuli: the mirror neuron hypothesis. Neuropsychologia, 46(5), 1558–1565.
Press, C., Bird, G., Flach, R., & Heyes, C. (2005). Robotic movement elicits automatic imitation. Cognitive Brain Research, 25(3), 632–640.
Press, C., Richardson, D., & Bird, G. (2010). Intact imitation of emotional facial actions in autism spectrum conditions. Neuropsychologia, 48(11), 3291–3297.
Rogers, S. J., Hepburn, S. L., Stackhouse, T., & Wehner, E. (2003). Imitation performance in toddlers with autism and those with other developmental disorders. Journal of Child Psychology and Psychiatry and Allied Disciplines, 44(5), 763–781.
Russell, J. (1997). Autism as an executive disorder. New York: Oxford University Press.
Rutter, M. (1974). The development of infantile autism. Psychological Medicine, 4, 147–163.
Shih, P., Shen, M., Ottl, B., Keehn, B., Gaffrey, M. S., & Müller, R. (2010). Atypical network connectivity for imitation in autism spectrum disorder. Neuropsychologia, 48(10), 2931–2939.
Spengler, S., Bird, G., & Brass, M. (2010a). Hyperimitation of actions is related to reduced understanding of others’ minds in autism spectrum conditions. Biological Psychiatry, 68(12), 1148–1155.
Spengler, S., von Cramon, D. Y., & Brass, M. (2010b). Resisting motor mimicry: control of imitation involves processes central to social cognition in patients with frontal and temporo-parietal lesions. Social Neuroscience, 5(4), 401–416.
van Baaren, R. B., Holland, R. W., Kawakami, K., & van Knippenberg, A. (2004). Mimicry and prosocial behavior. Psychological Science, 15(1), 71–74.
Williams, J. H. G., Whiten, A., & Singh, T. (2004). A systematic review of action imitation in autistic spectrum disorder. Journal of Autism and Developmental Disorders, 34(3), 285–299.
Williams, J. H., Whiten, A., Suddendorf, T., & Perrett, D. I. (2001). Imitation, mirror neurons and autism. Neuroscience and Biobehavioral Reviews, 25(4), 287–295.
Acknowledgments
Jennifer L. Cook was supported by a Wellcome Trust Ph.D. studentship.
Open Access
This article is distributed under the terms of the Creative Commons Attribution Noncommercial License which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This is an open access article distributed under the terms of the Creative Commons Attribution Noncommercial License (https://creativecommons.org/licenses/by-nc/2.0), which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
About this article
Cite this article
Cook, J.L., Bird, G. Atypical Social Modulation of Imitation in Autism Spectrum Conditions. J Autism Dev Disord 42, 1045–1051 (2012). https://doi.org/10.1007/s10803-011-1341-7
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10803-011-1341-7