
Abstract
Characterizing genetically distinct populations of primates is important for protecting biodiversity and effectively allocating conservation resources. Skywalker gibbons (Hoolock tianxing) were first described in 2017, with the only confirmed population consisting of 150 individuals in Mt. Gaoligong, Yunnan Province, China. Based on river geography, the distribution of the skywalker gibbon has been hypothesized to extend into Myanmar between the N’Mai Kha and Ayeyarwaddy Rivers to the west, and the Salween River (named the Thanlwin River in Myanmar and Nujiang River in China) to the east. We conducted acoustic point-count sampling surveys, collected noninvasive samples for molecular mitochondrial cytochrome b gene identification, and took photographs for morphological identification at six sites in Kachin State and three sites in Shan State to determine the presence of skywalker gibbons in predicted suitable forest areas in Myanmar. We also conducted 50 semistructured interviews with members of communities surrounding gibbon range forests to understand potential threats. In Kachin State, we audio-recorded 23 gibbon groups with group densities ranging between 0.57 and 3.6 group/km2. In Shan State, we audio-recorded 21 gibbon groups with group densities ranging between 0.134 and 1.0 group/km2. Based on genetic data obtained from skin and saliva samples, the gibbons were identified as skywalker gibbons (99.54–100% identity). Although these findings increase the species’ known population size and confirmed distribution, skywalker gibbons in Myanmar are threatened by local habitat loss, degradation, and fragmentation. Most of the skywalker gibbon population in Myanmar exists outside protected areas. Therefore, the IUCN Red List status of the skywalker gibbon should remain as Endangered.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Effective designation and management of conservation areas for wildlife is increasingly important in a world of limited resources and intensifying anthropogenic pressure (Torres-Romero et al., 2023). Basic estimates of animal populations, such as presence/absence and density, are essential to select appropriate conservation areas and inform wildlife management and international conservation policy. The International Union for Conservation of Nature (IUCN) uses estimates of the population size and distribution of a species as essential criteria for defining conservation status (IUCN, 2022). Such estimates often are difficult to obtain, particularly with cryptic or highly morphologically similar species. Animals with similar morphology can represent genetically distinct and divergent lineages, despite their similar appearance. To protect biodiversity, particularly among Hylobatidae, which have extraordinarily high genome plasticity (Carbone et al., 2014), it is important to identify genetically distinct populations and their distributions.
Skywalker hoolock gibbons (Hoolock tianxing) (hereafter referred to as skywalker gibbons) were first described in 2017, with the only confirmed population of 150 individuals occurring in Mt. Gaoligong, Yunnan Province, China (Fan et al., 2017). Based on gibbon ecology and the biogeographic patterns of large rivers acting as dispersal barriers that demarcate speciation boundaries in gibbons (Groves, 1967; Thinh et al., 2010a, b), the distribution of the skywalker gibbon is hypothesized to extend into Myanmar between the N’Mai Kha and Ayeyarwaddy Rivers to the west and the Salween River (named the Thanlwin River in Myanmar and the Nujiang River in China) to the east (Fan et al., 2017) (Fig. 1). A historical gibbon specimen at the Natural History Museum, London, collected in 1913 from the Goteik region of Shan State, Myanmar, which is located between these two riverine boundaries, was morphologically identified as a skywalker gibbon (Fan et al., 2017). However, no live skywalker gibbons have been confirmed as occurring in Myanmar over the past century. According to historical records and a limited population survey conducted in 2010, before the recognition of the skywalker gibbon as a distinct species, eastern hoolock gibbons (Hoolock leuconedys) were described as present in Kachin, Shan, Kayah, and Kayin States in Myanmar, which are now known to be within the potential range of the skywalker gibbon (Geissmann et al., 2013). During the 2010 assessment, the estimated gibbon population numbers were 89,000 in the Northern Kachin Forest complex, 150,000 in Southern Kachin-Northern Sagaing forests, 50,000 in Shan state subtropical forests, and 16,000 in Kayah-Kayin Montane rainforests (Geissmann et al., 2013).

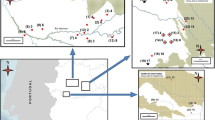
Survey site locations for acoustic gibbon monitoring conducted between December 2021 and March 2023. Site locations were located between the Ayeyarwaddy/N’Mai Kha and Salween (named Thanlwin in Myanmar and Nujiang in China) rivers. Habitat suitability determined according to Ngwe Lwin et al. (2021).
Primate surveys have historically been limited in Myanmar given its complex history of civil ethnic conflict concentrated near protected areas and reserve forests. Many of the historical hoolock gibbon records for Myanmar were collected by the Bombay Natural History Society’s “Mammal Survey of India, Burma and Ceylon” conducted between 1913 and 1915 (Wroughton, 1915), the Vernay-Hopwood Chindwin expedition of 1935 (Carter, 1943), and the Vernay-Cutting expedition conducted between 1938 and 1939 (Anthony, 1941). There is a considerable gap in records between 1940 and the 1980s. The most recent assessments in 2010 were limited in several geographic regions due to security issues. The accuracy of gibbon population estimates generated in 2010 is unknown, with the recognition of the skywalker gibbon as a separate species from the eastern hoolock gibbon further complicating the interpretation of these population estimates.
Our overarching goal is to provide accurate information for the skywalker gibbon species distribution and threats to guide updating of the IUCN Red List status. Our objectives were to: (1) conduct acoustic population surveys of gibbons in potential skywalker gibbon range; (2) conduct morphological and DNA identification of any individuals found; and (3) assess threats to new gibbon populations identified in Myanmar through the use of key informant interviews.
Methods
Evaluation of Predicted Suitable Habitat and Selection of Gibbon Population Survey Sites
To select survey sites between the Ayeyarwaddy and N’Mai Kha Rivers to the west and Salween River to the east, we collated current and historical records of hoolock gibbons from peer reviewed publications (Fan et al., 2011, 2017, Lwin et al., 2021, 2022) and published reports (Geissmann et al., 2013; Htoo & Grindley, 2010). In a previous study, we used 414 independent locations with documented gibbon presence in 1914–2019 to model the predicted range of hoolock species (Lwin et al., 2021). We identified nine forest areas with predicted hoolock species presence that fell within the estimated range of the skywalker gibbon in Myanmar. The largest forest areas within the theoretical skywalker gibbon range were in Kachin, Shan, Kayah, and Kayin States, including Paung Laung Reserve Forest (Fig. 1) (Lwin et al., 2021).
To improve the chance of locating gibbons in selected survey areas, we gathered additional information through interviews with local wildlife conservation organizations (Wapalaw Wildlife Watch and Friends of Wildlife) and members of the Myanmar Timber Enterprise, which conducts elephant logging operations throughout the proposed skywalker gibbon habitat. We telephoned local wildlife researchers, bird guides, and forestry staff to ask them questions about gibbon presence and hunting pressures. We selected sites to survey if they had: 1) a high likelihood of gibbon presence based on occupancy models; 2) a confirmed recent report of a gibbon sighting based on telephone interviews; and 3) were in close proximity to known populations of hoolock gibbon species and/or collection localities for historical museum specimens of skywalker gibbons. We also tried to select locations as far west and south as feasible to determine the western and southern-most borders of the range.
We ultimately conducted acoustic population monitoring in nine forest blocks between the Ayeyarwaddy/N’Mai Kai and Salween rivers in Kachin and Shan States (Table I). During the study period, sites in Kayah and Kayin States were inaccessible due to conflict (Fig. 1).
-
1.
Jid Lwai (26°11’ N, 98°19’), Myitkyina District, Kachin State, in a forest block adjacent to Imawbum National Park. The forest consists of broadleaved evergreen forest surrounded by shifting cultivation, predominantly cardamon.
-
2.
Lang Yang and 3. Kyauk Phyu Taung (25°58’ N, 98°19’ and 25°57’ N, 98°17’), Hsaw Law Township, Myitkyina District, Kachin State. The forest consists of mountainous broadleaved evergreen forest south of Imawbum National Park.
-
3.
Tamu Kone (25°41’ N, 98°11’), Chibwe Township, Myitkyina District, Kachin State. The forest consists of broadleaved evergreen forest with shifting cultivation for corn and cardamon plantations.
-
4.
Chaung Maw (25°39’ N, 98°02’), Wine Maw Township, Myitkyina District, Kachin State. The forest is located east of Sha Ngaw River, a tributary of the N’Mai Kha River, and is classified as remnant broadleaved evergreen forest surrounded by shifting cultivation.
-
5.
Sei Law (25°27’ N, 97°45’), Wine Maw Township, Myitkyina District, Kachin State. The forest is located in community managed forest near the N’Mai Kha River, and is a small mixed evergreen forest block on a north-south mountain range surrounded by a plantation area.
-
6.
Paung Laung Reserve Forest (19°45’–20°28’ N, 96°22’–96°49’), Pinlaung Township, Taunggyi District, Southern Shan State. The forest is classified as mixed deciduous in lower elevations and broadleaved evergreen forest in higher elevations (Kitahara et al., 2019; Mon et al., 2009).
-
7.
Sat Taw Reserve Forest (20°36’–20°43’ N, 97°45’–97°52’), Mongnei Township, Langkho District, Southern Shan State. The forest is classified as mixed deciduous forest, with some conifers present at higher elevations, with shifting cultivation.
-
8.
Mae Nei Taung Mountain Range (20°38’–20°48’ N, 97° 18’–97°23’), Hopong Township, Taunggyi District, Southern Shan State. The forest is classified as vacant land, with the remaining forested areas managed by local communities using traditional practices. The area is hilly and classified as evergreen forest, with mixed coniferous forests, hill savannas and Rhododendron forests found at higher elevations along the mountain range. Crops are grown in the mountain area, predominantly Assyrian plum (Cordia myxa), tea (Camellia sinensis), opium poppy (Papaver somniferum), and coffee (Coffea arabica).
Acoustic Population Survey Methods
We surveyed three forest blocks in Shan State between December 2021 and March 2023 and six forest blocks in Kachin State between March and May 2022. We used acoustic point-count sampling methods (i.e., triangulation), using established methods (Brockelman & Srikosamatara, 1993; Buckley et al., 2006; Cheyne et al., 2008; Brockelman et al., 2020; Hankinson et al., 2021), to investigate the presence of gibbons and to estimate gibbon density within each area (Cheyne et al., 2008). We established three listening posts in a triangular array, 300–600 m apart, within each site within forest blocks. We established sites 1–2 km apart to form an array. To avoid sampling bias due to elevation, we set up listening posts on hills/ridges on both sides of a mountain/valley. We avoided listening posts at the edge of forests or large rivers, where less gibbons are expected to be present. Gibbons require continuous canopy so degraded areas, rivers, and areas with highly fragmented canopy are not ideal habitats for gibbons and will yield an inaccurate estimate of the population (Cheyne et al., 2016; Hamard et al., 2010). We visited each array of three listening posts on 4–5 consecutive days from 06h00 to 11h00 am and recorded all song types at 3-min intervals. Because of local regulations and safety concerns, field teams could not enter forest areas long before or after dark. For each song heard, we recorded bearing, distance, song type (e.g., solo, duet, great calls (female vocalization)), and start and end time.
Other Field Sampling Methods
Following morning acoustic monitoring periods, we attempted to visually locate individual gibbons to take photographs of them and collect noninvasive biospecimens. We collected chewed plant samples (n = 11) to recover saliva from 11 individual gibbons observed feeding in Paung Laung using methods described previously (Smiley Evans et al., 2016). Specifically, we collected samples of dropped plants or fruits when a gibbon moved to a new foraging tree, cut out the portion of the discarded plant or fruit material that was most masticated using disposable sterile scalpel blades, and placed plant material in DNA/RNA Shield (Zymogen, Inc). We also opportunistically obtained a dry skin sample from a hunter in Lon Than village in Tamu Kone area. We preserved the dry specimen in a plastic bag at ambient temperature. We also obtained a single piece of skin from a museum specimen from the Natural History Museum of the United Kingdom (NHMUK 1933.7.29.15) collected in 1913 from Gokteik, northern Shan State (22°21’N, 96°55’E) that was provisionally classified as a skywalker gibbon based on morphology (Fan et al., 2017).
Photographic Morphologic Assessment
We examined photographs from individual gibbons from Shan State. We examined photographs visually for discrete external characteristics that distinguish between skywalker gibbons and other hoolock species based on descriptions from Mt. Gaoligong, China (Fan et al., 2017). We examined the following characteristics that are unique to H. tianxing and distinguish the species from eastern hoolock gibbons (Hoolock leuconedys): 1) the eyebrow streaks are thinner and separated by a large gap; 2) the beard is completely black or brown instead of white; 3) white hair is absent in the suborbital area; 4) the genital tuft is black, brown, or dark gray instead of whitish; and 5) adult females have incomplete white face rings, with only sparse white hairs present on the lateral orbital and suborbital regions (Fan et al., 2017).
Molecular Analyses
We used saliva, skin, and museum samples to molecularly verify species identity. We extracted total genomic DNA from saliva and the skin sample using Qiagen DNeasy kits (Qiagen, Inc.). From the saliva we amplified a 284-bp fragment of the mitochondrial cytochrome b (cytb) gene using primers 5’-ATGATACGAAAAACCATCGTTG-3’ and 5’-GCGAATAATTCAACCATAGTTC-3’. We performed amplification for 45 cycles, each with a denaturation step at 94 °C for 30 s, annealing at 52 °C for 30 s, and extension at 72 °C for 30 s, followed by a final extension step at 72 °C for 5 min. From the skin sample, we amplified the complete cytb gene by using previously described protocols (Thinh et al., 2010a, b). We checked PCR amplifications on 1.5% agarose gels. Subsequently, we cleaned PCR products with SpinPrep PCR Clean-up kits (Milipore) and sent them for Sanger-sequencing at Eurofins Genomics using the corresponding amplification primers. We checked sequence electropherograms with 4Peaks 1.8 and aligned sequences in SeaView 5.0.4 (Gouy et al., 2010). We also extracted the complete cytb sequence from an unpublished (C. Roos) complete mitochondrial genome of an adult male gibbon specimen in the NHMUK. We deposited this and newly generated sequences in GenBank, which are available under accession numbers OR159231-OR159233. We performed species identification using the online Basic Local Alignment Search Tool (BLAST) of the National Center for Biotechnology Information (NCBI) using standard settings.
Threat Assessment Surveys
We conducted key informant interviews in each site to assess overall anthropogenic activities present. Because there are no protected areas in Shan State and wildlife threat assessments have not previously been performed in this region, we conducted these key informant interviews to determine whether similar threats may impact gibbons in this region compared with others in Myanmar. We conducted interviews in 12 villages in Shan State between December 2021 and July 2022 to gather local ecological knowledge about gibbons and determine potential anthropogenic threats impacting any existing populations. We asked respondents about their religious affiliation, because it potentially had bearing on the level of hunting threat posed. We randomly selected villages based on proximity to two forest areas (Paung Laung and Mae Nei Taung Forest Reserves) where skywalker gibbon populations were confirmed to occur based on the combined results of our acoustic population surveys and DNA analyses. Villages ranged from 1 to 4 km from the edge of the forest. Within each village, we prioritized interviews with hunters or former hunters to obtain the most accurate information on gibbon threats. We invited individuals to be interviewed if they were aged 18 years or older and if they were likely to be engaged in activities posing threats to gibbons (e.g., hunting, logging, or forest disturbance). Given the sensitive nature of hunting, we identified potential respondents within a household by asking less direct questions, such as “who spends the most time in the forest?” and “who has the most contact with animals?” We conducted only one interview per household to ensure independence of responses and limit biases. All respondents gave informed oral consent to partake in the survey after being informed of the purpose and description of the study. Participation was voluntary and respondents were able to terminate their participation in the study at any time.
We created a standard Burmese-language questionnaire for all interviews, consisting of four sections comprised of 39 individual questions in total (Supplementary Material). Section one gathered basic demographic respondent information; section two focused on occupation and potential hunting activities in the forest; section three gathered information on gibbon knowledge, including respondents’ awareness and personal encounters with gibbons and potential threats to gibbon populations; and section four asked questions on forest cover changes and the potential reasons for such changes. We used Burmese research assistants with extensive experience working in hunting and agriculturally based communities to conduct questionnaires. We selected research assistants based on their ability to empathize with the human desire for hunting to increase relatability with respondents.
Statistical Methods
To estimate gibbon group density, we used the Acoustic Spatial Capture-Recapture (ASCR) R package developed for the IUCN SSC Primate Specialist Group Section on Small Apes (http://gibbons.asia/tag/ascr) (Stevenson, 2014). This package fits spatially explicit capture-recapture (SECR) models to acoustic data and has been used as an efficient density estimation method in previous gibbon studies (Kidney et al., 2016). SECR models require the use of a mask—a fine grid of latitude and longitude coordinates around each trap (array of listening posts)—to provide a set of plausible locations for each detected call. We set the maximum distance between each mask point and the closest trap to 2,000 m. We set the distance between adjacent mask points to 200 m. We determined the most appropriate detection function based on the Akaike Information Criterion (AIC). We ultimately used the half-normal detection function to model density: \(g\left(d;g0,\sigma \right)=g0{\text{exp}}\left(\frac{-{d}^{2}}{2 {\sigma }^{2}}\right)\), where d is the density of groups that call on a single day. Further division by the probability that a group calls on a given day (estimated from external data) yields an estimate of gibbon group density (in groups per hectare). g0 indicates the intercept for either the halfnormal or hazard-rate detection function, and σ is the scale parameter for either the halfnormal or hazard-rate detection function. \(g0\) was fixed as 1 for the single occasion surveys and \(d\) at 0 m (Kidney et al., 2016). We conducted analyses separately for each survey site (four sites in Paung Laung and one site in each of the eight remaining locations).
To calculate suitable skywalker gibbon habitat existing within and outside protected areas, we based overall habitat suitability estimates on earlier hoolock gibbon occupancy models that included skywalker gibbon data (misclassified as eastern hoolock gibbons, because the species had not yet been identified in Myanmar) (Lwin et al. 2021) overlapped with the new skywalker gibbon range proposed through this study according to the IUCN Red List Assessment Guidelines (https://www.iucnredlist.org/assessment/process). We calculated km2 of suitable habitat both within and outside of protected areas by overlaying shape files from Lwin et al. (2021) with Protected Area System boundaries provided by the Myanmar Forest Department.
To better understand dynamics contributing to threats in Skywalker gibbon forest sites, we assessed whether respondent sociodemographic characteristics, hunting activities, gibbon knowledge, and forest cover change varied between Paung Laung and Mae Nei Taung Forest sites. Interviews collected data on respondents’ age, gender, ethnicity, religion, occupation, education level, and frequency of visiting the local forest, all of which have been shown to influence knowledge and awareness of gibbons and other biodiversity within indigenous communities in China (Turvey et al., 2017; Zhang et al., 2020), and data on local interactions with gibbons and associated environmental change (perceptions of local gibbon population decline, reported hunting of gibbons, taboos against hunting gibbons, perceptions of local forest loss). To investigate variation in these responses between survey sites, we created generalized linear models (GLMs) with Gaussian or binomial error structures in R Version 4.2.2 and RStudio. However, most of the variables showed little variation across the overall respondent sample, showed collinearity with survey site (e.g., the two sites contained different ethnic and religious groups, meaning that the relative contribution of these factors to variation in interview responses could not be investigated independently of survey site), or had no responses from one of the two survey sites (e.g., no respondents reported gibbon hunting from Mae Nei Taung). As such, differences between the two survey sites were only investigated statistically for respondent age, whether respondents visited the forest at least once per week, reported gibbon declines, taboos against hunting gibbons, and perceptions of local forest loss. Given the relatively small respondent sample size (n = 24 in Paung Laung and n = 26 in Mae Nei Taung), we investigated relationships between each response variable and survey site using a series of separate models containing a single response variable and survey site as a single predictor variable to maximize statistical power within each model.
Ethical Note
All study procedures involving gibbons were non-invasive in nature. All gibbon survey procedures were approved by the Myanmar Forest Department and the Shan State Forest Department. All human surveys were conducted anonymously and no personal identifying information was collected. All human survey procedures were approved by the Institutional Review Board of the University of California, Davis (IRB #1863876-1) and the Myanmar Forest Department. The authors declare that they have no conflict of interest.
Data Availability
The datasets gathered and analyzed during the current study are available from the corresponding author on reasonable request.
Results
Population Surveys
In Kachin State, we recorded 23 gibbon groups at six survey sites: 12 groups at Kyauk Phyu, three groups each at Jid Lwai, Lang Yang and Tamu Kone, and one group each at Chaung Maw and Sei Law. Group densities estimated using the ASCR method varied between 0.57 and 3.6 group/km2 (Table I). In Shan State, we recorded 21 gibbon groups at two survey sites: 19 groups in Paung Laung, and two groups in Mae Nei Taung. We did not locate any gibbons in Sat Taw Reserve Forest. Group densities estimated using the ASCR method varied between 0.134 and 0.85 group/km2 (Table I; Fig. 2).

Proposed new skywalker gibbon (Hoolock tianxing) distribution area in Myanmar. Locations of survey sites in Kachin State (A) and Shan State (B), showing presence or absence of gibbons as assessed between December 2021 and March 2023. Size of circles in insets A and B represent gibbon densities.
The group density estimates for non-protected forest areas where gibbons were present (Jid Lwai, Lang Yang, Kyuak Phyu, Tamu Kone, Chaung Maw, Sei Law, Paung Laung, Mae Nei Taung) were not substantially different from the group density estimates for two of the more recently designated protected areas in Myanmar containing eastern hoolock gibbons, the Indawgyi Biosphere Reserve (0.71 group/km2, 95% confidence interval [CI] 0.4–1.27) and Mahamyaing Wildlife Sanctuary (0.97 group/km2, 95% CI 0.71–1.32) (Lwin et al., 2022, Tun et al., 2023). The main forest area in Shan State that we surveyed, Paung Laung Forest Reserve, was similar in size (1,337 km2) to Indawgyi Biosphere Reserve (1,700 km2) and Mahamyaing Wildlife Sanctuary (1,180 km2) but has a much lower level of legal conservation protection.
Morphological Descriptions
We evaluated 60 photographs of five individual gibbons (15 photographs of two adult females and one adult male from Paung Laung Reserve Forest, and 45 photographs of two adult males in Mae Nei Taung). All five individuals were very similar and could be clearly identified as skywalker gibbons based on distinct visual morphological traits. In the adult males, the eyebrows are well separated and white-colored hair is scant or absent around the eyes, chin, and genital tuft, making these areas appear brown or black (close to the color of the rest of the body). These features distinguish them from eastern hoolocks, which have many white hairs around their eyes and a white chin and genital tuft. In the adult females, there is less white fur around the eyes and the white fur does not form the conspicuous white face ring (connecting the chin and eyebrows) that is seen in the eastern hoolock gibbon (Fig. 3).

Morphological comparison of hoolock gibbon species and subspecies. Photographs of Hoolock hoolock, Hoolock hoolock mishmiensis, Hoolock leuconedys, and Hoolock tianxing (China) sourced and adapted from Fig. 2 in “Description of a new species of Hoolock gibbon (Primates: Hylobatidae) based on integrative taxonomy” (Fan et al., 2017). H. tianxing (Myanmar) photographs by Nature Conservation Society Myanmar and additional H. tianxing (China) photographs by P-F Fan. Top row: Male gibbons. Male H. tianxing have less convex tops of head, straighter white brows, and less conspicuous white beards compared with H. hoolock, H. h. mishmiensis, and H. leuconedys. Bottom row: Female gibbons. Female H. tianxing have fainter white face rings compared with H. hoolock, H. h. mishmiensis, and H. leuconedys.
Molecular Identification
We obtained a partial (217 bp) cytb sequence from one chewed plant sample collected in Paung Laung and the complete (1140 bp) cytb sequence from the skin sample from Tamu Kone. We also obtained the complete cytb sequence from the mitochondrial genome of the skin museum specimen from Gokteik. Ten chewed plant specimens collected did not contain sufficient DNA to detect gibbon genetic material. All three sequences obtained were identical in the 217 bp partial cytb sequence, whereas the two full-length cytb sequences differed in three positions (0.26%). Using BLAST search, we identified all three sequences unambiguously as skywalker gibbons. For the partial cytb sequence from Paung Laung we revealed 99.54–100% (0–1 differences) identity with skywalker gibbon, 97.70–98.62% (3–5 differences) identity with eastern hoolock gibbon and 97.24–97.70% (5–6 differences) identity with western hoolock gibbon (Hoolock hoolock) (100% query coverage). For the complete cytb sequence from Tamu Kone, we obtained 99.74–99.91% (1–3 differences) identity with skywalker gibbon, 97.70–98.42% (11–18 differences) identity with eastern hoolock gibbon and 96.84–97.19% (32–36 differences) identity with western hoolock gibbon (100% query coverage). For the complete cytb sequence of the museum specimen from Gokteik, BLAST results revealed 99.65–99.82% identity (2–4 differences) with skywalker gibbon, 98.33–98.95% (12–19 differences) identity with eastern hoolock gibbon, and 96.93–97.28% (31–35 differences) identity with western hoolock gibbon (100% query coverage).
Threat Assessments
We interviewed 50 people in Paung Laung (five villages, 24 respondents; mostly Karen ethnicity) and Mae Nei Taung (seven villages, 26 respondents; mostly Pa’O ethnicity). People in Paung Laung were equally divided between Buddhist and Christian religious affiliation and people in Mae Nei Taung were all Buddhist. Respondents varied in age from 19 to 85 years, 96% (n = 48/50) were male, and 94% (n = 47/50) practiced farming as their primary occupation. Respondents had considerable experience with local forests; 88% (n = 44/50) reported that they had visited the forest at least once per month, and 44% (n = 22/50) visited at least once per week for farming, collection of fuelwood and nontimber forest products, timber extraction, and hunting (for subsistence and sale). Hunting was reported by 82% (n = 41/50) of respondents. There was no statistically significant difference between the two forest areas in respondent age (GLM with Gaussian error structure: estimate = −0.56, standard error [SE] = 4.16, t = −0.13, p = 0.894) or whether respondents visited the forest at least once per week (GLM with binomial error structure: estimate = 0.09, SE = 0.60, z = 0.15, p = 0.884). These were the only two sociodemographic indices that showed substantial variation across the respondent sample, indicating that responses can be compared directly between the two areas without needing to control for additional potential confounding effects of respondent characteristics.
Respondents had a high level of awareness and experience of skywalker gibbons. In total, 82% (n = 41/50) could identify a skywalker gibbon from a photograph, and 98% (n = 49/50) could identify it from a playback of its call. Overall, only one respondent could not recognize the species at all. In total, 82% (n = 41/50) had seen and/or heard gibbons in the forest close to their village. Only 34% (n = 17/50) of respondents reported the date when they had last seen a gibbon in the local forest with dates ranging from within the past year to 30 years earlier.
Most respondents (70%; n = 35/50) felt that gibbons had declined, and there was no statistically significant difference in numbers of respondents reporting gibbon declines between the two forest areas (GLM with binomial error structure: estimate = −0.69, SE = 0.63, z = −1.10, p = 0.270). When asked about local hunting of gibbons, 46% (n = 11/24) reported that people hunted gibbons around Paung Laung; two respondents specifically stated that juvenile gibbons were hunted. In contrast, no respondents around Mae Nei Taung reported local hunting of gibbons. Awareness of local hunting in Paung Laung was reported by both Buddhists (n = 7/12) and Christians (n = 4/12), and from all five villages surveyed around this forest area. Considerably more respondents reported the existence of taboos against hunting gibbons around Mae Nei Taung (n = 17/26) compared with Paung Laung (n = 9/24); however this difference was not statistically significant (GLM with binomial error structure: estimate = −1.15, SE = 0.59, z = −1.95, p = 0.052).
Of the 34 respondents who described their perceptions for causes of gibbon decline, 85% (n = 29/34) of respondents considered that forest cover had decreased locally, with local forest loss reported to have occurred because of expansion of agriculture (including shifting cultivation) and agroforestry, logging, forest burning, and fuelwood collection. More respondents considered that forest cover had decreased in Paung Laung (n = 23/24) compared with Mae Nei Taung (n = 19/26); however, this difference was not statistically significant (GLM with binomial error structure: estimate = 2.14, SE = 1.11, z = 1.92, p = 0.055). Only two respondents thought that gibbon declines could be attributed to hunting.
Discussion
We report confirmation of skywalker gibbon presence in Myanmar, extending the species’ current range beyond China, and further supporting the hypothesis that the species range is demarcated by the N’Mai Kha and Ayeyarwaddy Rivers to the west and Salween River (Thanlwin in Myanmar and Nujiang in China) to the east. Our findings confirm that skywalker gibbons occur as far south as Shan State. Surveys in Kayin and Kayah States were not possible at the time of this study (due to security issues); however, it is likely that gibbon populations previously identified as eastern hoolock gibbons in Kayin and Kayah States are in fact skywalker gibbons. Based on these findings, we conclude that most of the world’s skywalker gibbon populations occur in Myanmar.
We confirmed skywalker gibbon presence noninvasively without access to voucher specimens or captive individuals, using chewed, naturally discarded plants. Recent gibbon voucher specimens are not preserved anywhere in Myanmar and historical specimens in international museums come with a degree of uncertainty regarding specific collection sites. Similarly, gibbons coming from zoological institutions within Myanmar may have inaccurate reported locations of origin. Fecal samples have long been the most commonly targeted noninvasive sample type for the identification of gibbon DNA (Oka & Takenaka, 2001; Bradley et al., 2001). However, feces often are a challenging specimen type to collect, given the frequency gibbons defecate in the presence of humans or other predatory animals. Identifying the origin of fecal samples in densely forested locations also can be challenging. Scat detection dogs have recently improved the likelihood of detection of gibbon fecal samples (Orkin et al., 2016); however, this resource was not available in Myanmar. We therefore used chewed, naturally discarded plants for DNA identification (Smiley Evans et al., 2016). We attempted to collect fecal samples during our surveys, but we did not find any. We did recover gibbon DNA from one out of 11 chewed plant specimens. This was a lower rate of DNA detection from chewed plants compared with similar studies performed using these methods with habituated mountain gorillas (Gorilla beringei beringei) and golden monkeys (Cercopithecus mitis kandti) (Smiley Evans et al., 2016). Our lower rate of detection was likely due to the greater distance at which wild gibbons had to be observed compared to habituated individuals from other species. Prioritizing collection of longer chewed plant specimens is easier the closer they can be observed (Smiley Evans et al., 2016). While we had a low rate of DNA detection, given the abundance of chewed plant samples available, we found this to be a viable method—showing that saliva recovered from this specimen type can be used for DNA identification of wild, nonhabituated gibbons.
We used both molecular identification and knowledge of gibbon ecology to determine the new proposed species range. We selected site locations for gibbon surveys located between the major north-south rivers of the Ayerwaddy/N’Mai Hka and Salween rivers and did not conduct gibbon surveys outside of this river-bound landscape. We made this decision based on other gibbon speciation records that show large rivers represent barriers for gibbon dispersal (Groves, 1967; Thinh et al., 2010a, b). Gibbons do not swim and are largely restricted to the forest canopy making river valleys generally unfavorable for gibbons (Groves, 1967; Thinh et al., 2010a, b). Other gibbon species in Myanmar have species distributions bounded by rivers: the Chindwin River is the boundary between western hoolock and eastern hoolock gibbons (Groves, 1967), and the Lohit River may act as the boundary between western hoolock and Mishmi Hills hoolock gibbon (Hoolock hoolock mishmiensis) (Choudhury, 2013). Based on this information, we did not conduct surveys west of the Ayerwaddy or east of the Thanlwin; however, for further confirmation of the species boundary, this survey effort could be attempted in future.
We found skywalker gibbons in a wide range of habitat types, extending from high elevational seasonal montane forests in the Mt. Gaoligong region of China (Fei et al., 2019) to lower elevation mixed deciduous forests of Shan State, Myanmar, including disturbed forest areas, demonstrating the adaptability of the species. We document an elevational range of 600–2000 meters above mean sea level (MAMSL) on the Myanmar side of the border. There are lower elevation forests in Myanmar within the hypothesized skywalker gibbon range, such as the Bago Yoma. None of these forests, however, were surveyed, because they had no recent gibbon sightings, a criterion for our selection of acoustic population monitoring locations. Skywalker gibbons have been documented between 1596–2685 MAMSL in China (Fan et al., 2011). This wide elevational range is similar to other hoolock gibbons: the western hoolock gibbon has been documented between 15 MAMSL (Ahsan, 2000) and 2300 MAMSL (Geissmann et al., 2013). The highest recorded elevation of a gibbon species is recorded in China, where Nomascus concolor has been documented above 2600 MAMSL (Fan & Jiang, 2010; Haimoff et al., 1987).
As further efforts are put toward revising the protected area system in Myanmar, we recommend that areas that support confirmed skywalker gibbon populations be included under protection to curb extremely high hunting and land conversion pressures from surrounding communities. We propose better protection for skywalker gibbons in the southernmost, lower elevation region of the newly defined skywalker gibbon range (Fig. 3). For example, in Shan State, where there are no protected areas, we identified gibbon groups in forested areas that are heavily degraded by logging and other anthropogenic activities. This finding indicates the extreme conservation urgency for this region, as skywalker gibbons remain but in low-quality habitats with high hunting pressures. Much less focus has been placed on protecting remaining forests in this region compared with more northern regions of Myanmar, where large areas of undisturbed forest landscape still exist. Restoration of habitats is a critical component of a multidimensional approach to primate conservation (Chapman et al., 2020) and identifying skywalker gibbons within degraded forests in Myanmar is an opportunity to help support conservation and reforestation in some of these areas. We therefore recommend that a portion of the southernmost habitat for skywalker gibbons also be prioritized for inclusion in the Myanmar protected area system (Fig. 4). Protected areas are positively associated with safeguarding primate species diversity globally (Torres-Romero et al., 2023) and should be incorporated into skywalker gibbon conservation action planning in Myanmar.

Proposed new protected area(s) to conserve the southern population of skywalker gibbons in Myanmar.
Community-based conservation efforts are also urgently needed and might be enacted on a much shorter time frame than that required to establish and manage new government organized protected areas. Based on discussions with communities who live in the survey areas in Kachin State, two communities (Jid Lwai, Site 1; and Tamu Kone, Site 4) stated their willingness to establish community protected areas to protect gibbons and to prevent land grabs for mining. In addition, one community (Sei Law, Site 6) wanted to start a gibbon conservation awareness program for their community to prevent hunting of gibbons. Shan State communities were enthusiastic for the conservation of gibbons and communities preferred to encourage indigenous and community conserved areas rather than the designation of a government operated protected area given current concerns around indigenous people’s rights in Shan State. Forested habitat within indigenous peoples’ lands (Garnett et al., 2018) is positively associated with safeguarding primate species diversity globally (Torres-Romero et al., 2023) and has had proven success in conserving primates in other parts of Myanmar (Thompson et al., 2023). Community-managed forests are one option to integrate forest management into national poverty reduction programs in rural areas. These forests have lower and less variable annual deforestation rates than protected ones (Porter-Bolland et al., 2012). Increasingly, a collective effort of stakeholders, including government-led protected areas as well as communities and indigenous peoples’ groups, is being recognized as the only effective way to protect and save our closest living relatives (Estrada et al., 2017; Fan, 2017; Pan et al., 2016; Li et al., 2018).
We also identified variation in local gibbon hunting, taboos, and forest loss across different social-ecological landscapes in Myanmar that highlight important local-scale differences in attitudes and interactions with gibbons and their habitat, indicating the need for region-specific community outreach and forest protection as part of future conservation efforts. These findings suggest that the skywalker gibbon population at Paung Laung may be more threatened by both hunting and habitat loss in comparison to other areas, and further research of this nature should be conducted as a matter of urgency to assess the status and viability of this population. More widely, our interview findings also demonstrate the high potential for using an interview survey approach to collect indigenous knowledge about the status of local gibbon populations more widely across Myanmar in the future as part of the conservation toolkit for strengthening the evidence-base for this poorly known species. These methods have been successful in measuring local ecological knowledge to detect cryptic threatened primates, such as the Hainan gibbon (Turvey et al, 2017), and could be expanded in Myanmar.
Skywalker gibbons are currently considered by the IUCN in its Red List assessment as Endangered A4cd (IUCN, 2022) because of the suspected population decline caused by significant and accelerating rates of deforestation and overexploitation for both food and medicinal purposes (Fan et al., 2020). Having confirmed the existence of skywalker gibbon populations in Myanmar, we propose the expansion of the species’ range to the Ayerwaddy River as the western demarcation and the Salween River as the eastern demarcation of the range (Fig. 4). Based on this updated distribution range, there is currently an estimated 7,248 km2 of suitable habitat in China and 31,703 km2 of suitable habitat in Myanmar (38,951 km2 in total) for skywalker gibbons. Of the suitable skywalker gibbon habitat within the Myanmar and China Protected Area Systems (areas with official conservation protection status), we determined that there are 1,012 km2 in China and 1,368 km2 in Myanmar. Overall, a very small fraction of the suitable habitat for skywalker gibbons in Myanmar exists within the protected area system (~4%).
While skywalker gibbons have now been confirmed to occur in Myanmar, thus increasing the species’ global population size and confirmed distribution, threats to the survival of the species across the region are extensive, including local habitat loss, degradation and fragmentation, and hunting (both for meat and for live-trade) in both China and Myanmar. Nearly half of the remnant population in China lives in landscapes used for agroforestry (Fei et al., 2017), and based on our data, greater than 95% of the population in Myanmar exists outside protected areas. A suspected population decline caused by significant rates of deforestation in Myanmar (estimated at 0.94% annually between 2002 and 2014, totaling >2 million ha forest loss) is projected to continue and potentially accelerate (Bhagwat et al., 2017). A population loss equaling or exceeding 50% can thus be anticipated over three generations (between 2000 and 2045). Therefore, the conservation status of the skywalker gibbon should remain as Endangered A4cd.
References
Ahsan MF (2000) Socio-ecology of the hoolock gibbon (Hylobates hoolock) in two forests of Bangladesh. In: Chicago Zoological Society (Ed.), The apes: challenges for the 21st century (pp. 286–299). Chicago Zoological Society.
Anthony, H. E. (1941). Mammals collected by the Vernay-Cutting Burma expedition. Field Museum of Natural History, Zoological Series, 27, 37–123.
Bhagwat, T., Hess, A., Horning, N., Khaing, T., Thein, Z. M., Aung, K. M., Aung, K. H., Phyo, P., Tun, Y. L., Oo, A. H., Neil, A., Thu, W. M., Songer, M., Connette, K. L., Bernd, A., Huang, Q., Connette, G., & Leimgruber, P. (2017). Losing a jewel – Rapid declines in Myanmar’s intact forests from 2002–2014. Plos One, 12(5), e0176364. https://doi.org/10.1371/journal.pone.0176364
Bradley, B. J., Chambers, K. E., & Vigilant, L. (2001). Accurate DNA-based sex identification of apes using non-invasive samples. Conservation Genetics, 2, 179–181.
Brockelman, W. Y., & Srikosamatara, S. (1993). Estimating density of gibbon groups by use of the loud songs. American Journal of Primatology, 29(2), 93–108. https://doi.org/10.1002/ajp.1350290203
Brockelman, W. Y., Tun, A. Y., Pan, S., Naing, H., & Htun, S. (2020). Comparison of point transect distance and traditional acoustic point-count sampling of hoolock gibbons in Htamanthi Wildlife Sanctuary, Myanmar. American Journal of Primatology, 82(12), e23198. https://doi.org/10.1002/ajp.23198
Buckley, C., Nekaris, K. A., & Husson, S. J. (2006). Survey of Hylobates agilis albibarbis in a logged peat-swamp forest: Sabangau catchment, Central Kalimantan. Primates, 47(4), 327–335. https://doi.org/10.1007/s10329-006-0195-7
Carbone, L., Alan Harris, R., Gnerre, S., et al. (2014). Gibbon genome and the fast karyotype evolution of small apes. Nature, 513, 195–201. https://doi.org/10.1038/nature13679
Carter, D. T. (1943). The mammals of the Vernay-Hopwood Chindwin expedition, Northern Burma. Bulletin of the American Museum of Natural History, 82(4), 95–114.
Chapman, C., Bicca-Marques, J. C., Dunham, A., Fan, P., Fashing, P. J., Gogarten, J., Guo, S., Huffman, M., Kalbitzer, U., Li, B., Ma, C., Batsuda, I., Omeja, P., Sarkar, D., Sengupta, R., Serio-Silva, J. C., Tsuji, Y., & Stenseth, N. (2020). Primates can be a rallying symbol to promote tropical forest restoration. Folia Primatologica, 91(6), 669–687.
Cheyne, S. M., Thompson, C. J., Phillips, A. C., Hill, R. M., & Limin, S. H. (2008). Density and population estimate of gibbons (Hylobates albibarbis) in the Sabangau catchment, Central Kalimantan, Indonesia. Primates, 49(1), 50–56. https://doi.org/10.1007/s10329-007-0063-0
Cheyne, S. M., Gilhooly, L., Hamard, M. C., Hoing, A., Houlihan, P. R., KursaniLoken, B., Philips, A., Rayadin, Y., Capila, B. R., Rowland, D., Sastramidjaja, W. J., Spehar, S., Thompson, C., & Zrust, M. (2016). Population mapping of gibbons in Kalimantan, Indonesia: correlates of gibbon density and vegetation across the species’ range. Endangered Species Research, 30, 133–143.
Choudhury, A. (2013). Description of a new subspecies of Hoolock gibbon Hoolock hoolock from north east India. Newsletter & Journal of the Rhino Foundation for Nature India, 9, 49–59.
Estrada, A., Garber, P. A., Rylands, A. B., Roos, C., Fernandez-Duque, E., Di Fiore, A., Nekaris, K. A., Nijman, V., Heymann, E. W., Lambert, J. E., Rovero, F., Barelli, C., Setchell, J. M., Gillespie, T. R., Mittermeier, R. A., Arregoitia, L. V., de Guinea, M., Gouveia, S., Dobrovolski, R., … Li, B. (2017). Impending extinction crisis of the world’s primates: Why primates matter. Science Advances, 3(1), e1600946. https://doi.org/10.1126/sciadv.1600946
Fan, P. F. (2017). The past, present, and future of gibbons in China. Biological Conservation, 210, 29–39. https://doi.org/10.1016/j.biocon.2016.02.024
Fan, P., & Jiang, X. (2010). Altitudinal ranging of black-crested gibbons at Mt. Wuliang, Yunnan: Effects of food distribution, temperature and human disturbance. Folia Primatologica, 81(1), 1–9. https://doi.org/10.1159/000279465
Fan, P.-F., Wen, X., Sheng, H., Huai-Sen, A., Tian-Can, W., & Ru-Tao, L. (2011). Distribution and conservation status of the vulnerable eastern hoolock gibbon Hoolock leuconedys in China. Oryx, 45(1), 129–134. https://doi.org/10.1017/S003060531000089X
Fan, P. F., He, K., Chen, X., Ortiz, A., Zhang, B., Zhao, C., Li, Y.-Q., Zhang, H.-B., Kimock, C., Wang, W.-Z., Groves, C., Turvey, S. T., Roos, C., Helgen, K. M., & Jiang, X.-L. (2017). Description of a new species of Hoolock gibbon (Primates: Hylobatidae) based on integrative taxonomy. American Journal of Primatology, 79(5), e22631. https://doi.org/10.1002/ajp.22631
Fan, P. F., Turvey, S. T., & Bryant, J. V. (2020). Hoolock tianxing (amended version of 2019 assessment). The IUCN Red List of Threatened Species, 2020, e.T118355648A166597159. https://doi.org/10.2305/IUCN.UK.2020-1.RLTS.T118355648A166597159.en. Accessed 21 June 2023.
Fei, H. L., Zhang, D., Yuan, S. D., Zhang, L., & Fan, P. F. (2017). Antipredation sleeping behavior of Skywalker hoolock gibbons (Hoolock tianxing) in Mt. Gaoligong, Yunnan, China. International Journal of Primatology, 38, 629–641. https://doi.org/10.1007/s10764-017-9970-0
Fei, H., Thompson, C. & Fan, P. (2019). Effects of cold weather on the sleeping behavior of Skywalker hoolock gibbons (Hoolock tianxing) in seasonal montane forest. American Journal of Primatology, 81(9), e23049. https://doi.org/10.1002/ajp.23049
Garnett, S. T., Burgess, N. D., Fa, J. E., et al. (2018). A spatial overview of the global importance of Indigenous lands for conservation. Nature Sustainability, 1, 369–374. https://doi.org/10.1038/s41893-018-0100-6
Geissmann, T., Grindley, M. E., Lwin, N., Aung, S. S., Aung, T. N., Htoo, S. B., & Momberg, F. (2013). The conservation status of hoolock gibbons in Myanmar. Gibbon Conservation Alliance.
Gouy, M., Guindon, S., & Gascuel, O. (2010). SeaView version 4: a multiplatform graphical user interface for sequence alignment and phylogenetic tree building. Molecular Biology and Evolution, 27(2), 221–224. https://doi.org/10.1093/molbev/msp259
Groves, C. P. (1967). Geographic variation in the hoolock or white-browed gibbon (Hylobates hoolock Harlan 1834). Folia Primatologica, 7, 276–283.
Haimoff, E. H., Yang, X. J., He, S. E., & Chen, N. (1987). Preliminary observations of wild black-crested gibbons (Hylobates concolor concolor) in Yunnan Province. People’s Republic of China. Primates, 28(3), 319–344. https://doi.org/10.1007/BF02381015
Hamard, M., Cheyne, S. M., & Nijman, V. (2010). Vegetation correlates of gibbon density in the peat-swamp forest of the Sabangau catchment, central Kalimantan, Indonesia. American Journal of Primatology, 71, 1–10.
Hankinson, E., Hill, R. A., Marsh, C. D., Nowak, M. G., Abdullah, A., Pasaribu, N., Supriadi Nijman, V., Cheyne, S. M., & Korstjens, A. H. (2021). Influences of forest structure on the density and habitat preference of two sympatric gibbons (Symphalangus syndactylus and Hylobates lar). International Journal of Primatology, 42(2), 237–261. https://doi.org/10.1007/s10764-021-00199-2
Htoo, S. B., & Grindley, M. (2010). Hoolock Gibbon and Biodiversity Survey on Khe Shor Ter Mountain, Nattaung Range, Luthaw Township, Mudraw District, Karen State, Myanmar. Myanmar Primate Conservation Program Report No. 11. Karen Environmental and Social Action Net (KESAN), and People Resource and Conservation Foundation (PRCF). http://www.gibbonconservation.org/05_projects/2010karen_3final_eng.pdf
IUCN Standards and Petitions Committee. (2022). Guidelines for using the IUCN red list categories and criteria. Version 15.1. Prepared by the Standards and Petitions Committee. Available at: https://www.iucnredlist.org/documents/RedListGuidelines.pdf
Jones-Todd, C. M. (2019). Acoustic spatial capture-recapture (ascr). Available at: https://cmjt.shinyapps.io/ascr_shiny/.
Kidney, D., Rawson, B. M., Borchers, D. L., Stevenson, B. C., Marques, T. A., & Thomas, L. (2016). An efficient acoustic density estimation method with human detectors applied to gibbons in Cambodia. PLoS One, 11(5), e0155066. https://doi.org/10.1371/journal.pone.0155066
Kitahara, F., Sato, T., Win, B. N., Takao, G., Win, T. T., & Maung, K. W. (2019). Examination of bamboo measurement method for biomass estimation in the Paung Laung reserve forest, Myanmar. AGRIVITA, Journal of Agricultural Science, 41(1), 32–39. https://doi.org/10.17503/agrivita.v40i3.2009
Li, B., Li, M., Li, J., Fan, P., Ni, Q., Lu, J., Zhou, X., Long, Y., Jiang, Z., Zhang, P., Huang, Z., Huang, C., Jiang, X., Pan, R., Gouveia, S., Dobrovolski, R., Greuter, C., Oxnard, C., Groves, C., …, Garber, A. (2018). The primate extinction crisis in China: Immediate challenges and a way forward. Biodiversity Conservation, 27, 3301–3327. https://doi.org/10.1007/s10531-018-1614-y
Lwin, K. S. S., Matssudaira, K., & Oo, K. S. (2019). Species identification of hoolock gibbons at zoological gardens in Myanmar. Direct GenBank Submission https://www.ncbi.nlm.nih.gov/nuccore/1913240952
Lwin, N., Sukumal, N., & Savini, T. (2021). Modelling the conservation status of the threatened hoolock gibbon (genus Hoolock) over its range. Global Ecology and Conservation, 29, e01726. https://doi.org/10.1016/j.gecco.2021.e01726
Lwin, N., Ngoprasert, D., Sukumal, N., Browne, S., & Savini, T. (2022). Status and distribution of hoolock gibbon in the newly established Indawgyi Biosphere Reserve: implication for protected area management. Global Ecology and Conservation, 38, e02209. https://doi.org/10.1016/j.gecco.2022.e02209
Mon, M. S., Kajisa, T., Mizoue, N., & Yoshida, S. (2009). Factors affecting deforestation in Paunglaung watershed, Myanmar using remote sensing and GIS. Journal of Forest Planning, 14(1), 7–16. https://doi.org/10.20659/jfp.14.1_7
Oka, T., & Takenaka, O. (2001). Wild gibbons’ parentage tested by non-invasive DNA sampling and PCR-amplified polymorphic microsatellites. Primates, 42, 67–73. https://doi.org/10.1007/BF02640690
Orkin, J. D., Yang, Y., Yang, C., Yu, D. W., & Jiang, X. (2016). Cost-effective scat-detection dogs: unleashing a powerful new tool for international mammalian conservation biology. Scientific Reports, 6, 34758. https://doi.org/10.1038/srep34758
Pan, R., Oxnard, C., Gruerter, C. C., Li, B., Zi, X., He, G., Guo, S., & Garber, P. A. (2016). A new conservation strategy for China- a model starting with primates. American Journal of Primatology, 78(11), 1137–1148. https://doi.org/10.1002/ajp.22577
Porter-Bolland, L., Ellis, E., Guariguata, M. R., Ruiz-Mallen, I., Negrete-Yankelevich, S., & Reyes-Garcia, V. (2012). Community managed forests and forest protected areas: an assessment of their conservation effectiveness across the tropics. Forest Ecology and Management, 268, 6–17. https://doi.org/10.1016/j/foreco.2011.05.034
Smiley Evans, T., Gilardi, K., Barry, P., Ssebide, B., Kinani, J. F., Nizeyimana, F., Noheri, J. B., Byarugaba, D., Mudkikiwa, A., Cranfield, M., Mazet, J., & Johnson, C. (2016). Detection of viruses using discarded plants from wild mountain gorillas and golden monkeys. American Journal of Primatology, 78(11), 1222–1234. https://doi.org/10.1002/ajp.22576
Stevenson, B. (2014). ascr: An R package for acoustic spatial capture-recapture using AD Model Builder. Available at: http://hdl.handle.net/2292/40239
Thinh, V. N., Mootnick, A. R., Geissmann, T., Li, M., Ziegler, T., Agil, M., Moisson, P., Nadler, T., Walter, L., & Roos, C. (2010a). Mitochondrial evidence for multiple radiations in the evolutionary history of small apes. BMC Evolutionary Biology, 10, 74. https://doi.org/10.1186/1471-2148-10-74
Thinh, V. N., Rawson, B., Hallam, C., Kenyon, M., Nadler, T., Walter, L., & Roos, C. (2010b). Phyologeny and distribution of crested gibbons (genus Nomascus) based on mitochondrial cytochrome b gene sequence data. American Journal of Primatology, 72, 1047–1054. https://doi.org/10.1002/ajp.20861
Thompson, C., Lwin, N., Aung, P. P., Aung, H. T., Htike, T., San, A. M., Thant, N. M., Roos, C., Fan, P. F., Rompay, K., Grindley, M., Tin, P. P., Wai, N. N., Lwin, H., Gilardi, K., Momberg, F., Cheyne, S., & Smiley Evans, T. (2023). The status of primates and primatology in Myanmar. Global Ecology and Conservation, 47, e02662. https://doi.org/10.1016/j.gecco.2023.e02662
Torres-Romero, E. J., Nijman, V., Fernandez, D., & Eppley, T. M. (2023). Human-modified landscapes driving the global primate extinction crisis. Global Change Biology, 29, 5775–5787. https://doi.org/10.1111/gcb.16902
Tun, W. K. K. N., Sukumal, N., Ngoprasert, D., Shwe, N. M., & Savini, T. (2023). Gibbon population status and long-term viability: Implication for a newly established protected area management. Global Ecology and Conservation, e02534. https://doi.org/10.1016/j.gecco.2023.e02534
Turvey, S. T., Bryant, J. V., Duncan, C., Wong, M. H. G., Guan, Z., Fei, H., Ma, C., Hong, X., Nash, H. C., Chan, B. P. L., Xu, Y., & Fan, P. (2017). How many remnant gibbon populations are left on Hainan? Testing the use of local ecological knowledge to detect cryptic threatened primates. American Journal of Primatology, 79(2), 1–13.
Wroughton, R. C. (1915). Bombay natural history society’s mammal survey of India, Burma, and Ceylon. The Journal of the Bombay Natural History Society, 23(4), 460–480.
Zhang, L., Guan, Z., Fei, H., Yan, L., Turvey, S. T., & Fan, P. (2020). Influence of traditional ecological knowledge on conservation of the skywalker hoolock gibbon (Hoolock tianxing) outside nature reserves. Biological Conservation, 241, 108267.
Acknowledgments
This research was funded by the Arcus Foundation (G-PGM-1912-3087), the IUCN Section on Small Apes, and the International Centre for Integrated Mountain Development (ICIMOD). TSE was supported by the Fogarty International Center of the National Institutes of Health (Grant no. K01TW010279) and the National Science Foundation (Grant no. 2109860). The authors thank the Forest Department of the Ministry of Natural Resources and Environmental Conservation (MONREC) for their support of gibbon conservation in Myanmar, the Shan State Government and Forest Department for facilitating village interviews and supporting this research, the Myanmar Timber Enterprise (MTE), Wapalaw Wildlife Watch and Friends of Wildlife for providing information on gibbon presence, the Livestock Breeding and Veterinary Department for contributing to molecular species identification as continued technical support from previously implemented NIH funded project with UC Davis (K01TW010279), and JoAnn Yee and the California National Primate Research Center Primate Assay Laboratory Core for guidance on primate sampling. The authors thank the editor and two anonymous reviewers for their constructive feedback on the manuscript.
Author information
Authors and Affiliations
Contributions
PPA: conceptualization, data curation, funding acquisition, formal analysis, writing – original draft; NL: conceptualization, data curation, formal analysis, visualization, writing – original draft; THA: investigation; TSMH: investigation, writing – original draft; CT: conceptualization, data curation, formal analysis, funding acquisition, writing – original draft; CR: formal analysis, writing-original draft; LZL: investigation; WNO: investigation; ZS: investigation; STT: methodology, formal analysis, writing – review & editing; SMZ: investigation; WZ: investigation; MTM: investigation; YTW: resources, supervision; ZMO: methodology, supervision; KAVR: methodology, writing – review & editing; KVG: funding acquisition, project administration, writing – review & editing; ATB: investigation; FM: conceptualization, methodology, supervision; PFF: methodology, supervision, writing – original draft; SMC: conceptualization, data curation, formal analysis, funding acquisition, methodology, supervision, writing-original draft; TSE: conceptualization, formal analysis, funding acquisition, methodology, project administration, supervision, writing-original draft
Corresponding author
Ethics declarations
Inclusion and Diversity Statement
Twelve authors (including joint first, second, and third authors) are contributors from the location where the research was conducted, and participated in study conception, study design, data collection, analysis, and interpretation of the findings.
Additional information
Handling Editor: Joanna (Jo) M. Setchell
Pyae Phyo Aung and Ngwe Lwin joint first authors.
Susan M. Cheyne and Tierra Smiley Evans joint senior authors.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Aung, P.P., Lwin, N., Aung, T.H. et al. Confirmation of Skywalker Hoolock Gibbon (Hoolock tianxing) in Myanmar Extends Known Geographic Range of an Endangered Primate. Int J Primatol (2024). https://doi.org/10.1007/s10764-024-00418-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10764-024-00418-6