
Abstract
Understanding population connectivity helps inform resource and conservation managers about appropriate boundaries for management units. However, for many species facing recent increases in fishing or environmental pressure, accurate information on population structure is lacking. Reef Bugs (Thenus australiensis), commonly known as Moreton Bay Bugs, are iconic shovel-nosed lobsters in northern Australia and an increasingly important fisheries resource. Around 80% of landings occur in the Queensland East Coast Otter Trawl Fishery (ECOTF) where the first stock assessment of the species is underway. However, knowledge about population structure to inform stock assessment is lacking. We used genome complexity reduction-based sequencing to characterise high-quality Single Nucleotide Polymorphisms (SNPs) used to evaluate population structure among three management zones of the ECOTF. Post-filtering, a total of 3,031 SNPs were used to infer no genetic differences among locations indicating strong genetic population connectivity. The presence of a single panmictic population was further supported by cluster and kinship analyses. Broad-scale genetic connectivity likely results from pelagic larval dispersal due to limited adult movements. Findings of high gene flow among connected populations indicate a single biological stock of T. australiensis on Australia’s east coast and will inform future fisheries management initiatives.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Identifying appropriate management units for wild animal populations is key to both sustainable resource management and conservation (Palsbøll et al. 2007; Funk et al. 2012). In marine taxa, biological stocks can be defined as networks of breeding groups sufficiently connected and well mixed to form self-replenishing units (Jennings and Kaiser 1998). The stock concept recognises that isolated populations may be more vulnerable to stressors than populations connected by dispersal processes and should be managed accordingly (Begg et al. 1999; Waples et al. 2008). Therefore, efficacy of management measures, e.g. stock assessment or quota allocation, and relevance of monitoring activities are best achieved at the scale of biological stocks (Ovenden et al. 2015). Failure to account for population structure can also lead to detrimental effects on the productivity, biology, and genetic diversity of populations (Altukhov 1981; Ricker 1981; Smith et al. 1991). Identifying population connectivity, i.e. mixing among populations via dispersal processes, is, therefore, necessary to define boundaries of homogeneous stocks and inform effective, evidence-based management.
Marine populations are often considered to be highly connected due to frequently large population sizes, a lack of geographic barriers that filter gene flow, and physical processes that support broad-scale dispersal of eggs, larvae, or spores (Cowen and Sponaugle 2009; Crawford and Oleksiak 2016). This is particularly true of species with long pelagic larval durations (i.e. months or years), including many lobsters, which can facilitate transport of larvae over large distances (Silva et al. 2019). Although some lobsters exhibit extensive adult movements (e.g. in the order of hundreds of kilometres: Bell et al. 1987), others display very limited adult movements (e.g. < 1 km per year: Gardner et al. 2003). Given the bipartite life cycle of lobsters (pre- and post-settlement), populations with limited adult movement rely mostly on passive larval dispersal for dispersal and population mixing. Advection by dominant current regimes is usually the main physical process driving horizontal larval dispersal. However, larval behaviour may alter these effects, by selecting currents that assist transport in favourable directions in response to environmental stimuli (Kingsford et al. 2002; Metaxas and Saunders 2009). Therefore, despite potential sources of variation among species and locations based on pelagic larval duration, current regimes at varying spatial scales, and larval behaviour, population structuring in lobsters is often limited, yet important to investigate to support evidence-based management.
Moreton Bay Bugs (Thenus spp.) are Scyllarid lobsters distributed around the northern coastline of Australia. Like other Scyllaridae, Moreton Bay Bugs have evolved a dorso-ventrally flattened morphology allowing exploitation of open sedimental habitats where they find refuge by burying in soft sediments during the day before emerging to forage nocturnally. Around 80% of landings in Australia occur in the East Coast Otter Trawl Fishery (ECOTF) on the east coast of Queensland. Reef Bugs (Thenus australiensis Burton & Davie, 2007) comprise ~ 93% of Moreton Bay Bug landings in the ECOTF (McMillan et al. 2023), with the remainder comprising the congeneric Mud Bug (T. parindicus Burton & Davie, 2007). Within the ECOTF, ~ 80% of landings occur from two main grounds off Townsville in the Central management zone and off Gladstone–Bundaberg in the Southern Inshore zone (Fig. 1). Remaining landings occur from Far North Queensland to the Sunshine Coast in the south, with total landings amounting to ~ 400 to 700 tonnes annually. Originally viewed as a bycatch species in the ECOTF, T. australiensis has gained considerably in market value in recent years leading to increased targeted fishing (McMillan et al. 2023). Stock depletion of the co-located Saucer Scallop (Ylistrum balloti Bernardi, 1861) in the Southern Inshore zone has also led to large-scale effort shifts toward targeting of T. australiensis using similar fishing gear as previously employed in the scallop fishery. These combined effects have led to increased management focus on T. australiensis and a need for the first stock assessment of the species to understand the status of the resource, but information on the spatial scale of appropriate management units has been lacking.

Average annual harvest of T. australiensis in the Queensland East Coast Otter Trawl Fishery, Australia, from 1988 to 2021. The Townsville and Gladstone–Bundaberg regions each account for ~ 40% of total landings. Landings are filtered by the top 95% of reporting grids and top 99% of fishers to be representative of the fishery. Landings in Princess Charlotte Bay (PCB) fall outside the top 95% of grids but are included to demonstrate the relatively low abundance of T. australiensis in Far North Queensland
Like many lobsters, T. australiensis displays limited adult movements (Jones 1988; McMillan et al. 2023) and likely relies on the extended duration of its pelagic larval phase (up to 45 days: Mikami and Greenwood 1997) as the primary mode of dispersal and population mixing. Two spawning peaks occur over Austral summer, but low-level spawning continues year-round (Jones 1988). Larval transport is influenced by a complex regime of currents including the poleward flowing East Australia Current and counterflows caused by southeast trade winds as well as numerous eddies caused by the abundant reefs and islands of the Great Barrier Reef system. The mediating effects of local forcing caused by wind and tides acting on prevailing currents as well as seasonal variations in current velocity and direction add further complexity to flows in the Great Barrier Reef lagoon (Benthuysen et al. 2022). The extent and spatio-temporal complexity of these flows, as well as year-round spawning and the relatively long pelagic larval duration of T. australiensis likely support extensive larval dispersal and population mixing. This study investigates the genetic population structure in T. australiensis to support stock assessment and future management by determining the appropriate spatial scale for management.
Modern genetic tools provide the capacity to examine population structure and identify appropriate boundaries of management units. Recent advances in genome-wide sequencing allow high-resolution comparison of Single Nucleotide Polymorphisms (SNPs) at thousands of loci, with the increased power often permitting detection of fine-scale structuring among populations (Georges et al. 2018; Silva et al. 2019; Woodings et al. 2018). Here we use a genome complexity reduction-based sequencing approach to investigate population structure of T. australiensis throughout their main distribution within the ECOTF area to inform appropriate spatial scales for stock assessment and future management.
Materials and methods
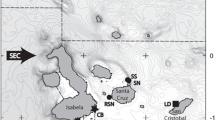
Samples were collected from individual T. australiensis caught using conventional otter trawl gear by commercial vessels operating in the ECOTF from March to August 2022. Tissue samples were taken from pleopods (abdominal appendages used for swimming and brooding) of 70 individuals at four locations. Sampling locations included Bundaberg (BBG; n = 30) in the Southern Inshore management zone, Townsville (TSV; n = 30) in the Central zone, and Mission Beach (MB; n = 6) and Princess Charlotte Bay (PCB; n = 4) in the Northern zone (Fig. 2). Smaller sample sizes from the Northern zone reflect the relatively low abundance of T. australiensis in this zone compared to the main grounds located off Townsville and Bundaberg (Fig. 1).

Sampling sites for T. australiensis collected from commercial trawl vessels operating in Queensland’s East Coast Otter Trawl Fishery. Sampling locations include Princess Charlotte Bay (PCB), Mission Beach (MB), Townsville (TSV) and Bundaberg (BBG). TMZs = Trawl Management Zones
Tissue samples were preserved in 95% ethanol solution and stored at – 18 °C. Small amounts of biological tissue (< 15 mg) were transferred to a well plate and sent to Diversity Arrays Technology for DNA extraction and sequencing (DArT Pty Ltd, Canberra, Australia). Samples were genotyped to acquire SNPs using DArTseqTM technologies, a complexity-reduced sequencing method as described by Grewe et al. (2015). DNA samples were processed in digestion/ligation reactions, ligating two adaptors corresponding to the combination of PstI and SphI restriction enzyme overhangs. The PstI-compatible adapter includes the barcode. The PCR primers were designed to add the required sequences for enabling sequencing in a single-read Illumina flowcell, and an Illumina Hiseq2500 (Illumina, San Diego) was used to sequence on average 3.5 million reads per sample. The generated sequences were processed using proprietary DArT analytical pipelines to assess read quality and undertake read assembly and SNP calling (Kilian et al. 2012; Georges et al. 2018). The marker extraction pipeline consisted of de novo variant-calling using DArTsoft v.14, a software package developed by DArT P/L. The pipeline first clustered DArT-tags with three or fewer base pair differences using DArT P/L’s C++ algorithm and parsed into separate SNP loci.
After the DArTseqTM quality control stage of filtering, SNP data and metadata were transformed into a genlight object for further filtering in R Studio (R version 4.2.1, R Core Team 2022) using the package ‘dartR’ (Gruber et al. 2018). SNPs were tested for repeatability (≥ 99%), loci call rate (≥ 70%), and individual call rate (≥ 85%), resulting in the removal of two individuals. Filtering for individual heterozygosity resulted in removal of two further individuals (see Table S1 for specific filtering procedures).
Population structure was visualised through principal component analysis (PCA) and Discriminant Analysis of Principal Components (DAPC) to plot genetic similarity between individuals and locations using ‘dartR’ and ‘adegenet’ (Jombart 2008; Gruber et al. 2018). Statistical methods to test for genetic differentiation included genetic distance and isolation by distance tests, pairwise fixation indices (FST), observed (HO) and expected (HE) heterozygosity values and inbreeding coefficients (FIS). Using the R package ‘SNPRelate’ (Zheng et al. 2012), kinship coefficients were calculated based on estimates of maximum likelihood that genes shared by pairs of individuals are identical by descent. Coefficients of relatedness (r) ranging from 0 to 1 (where 0.5 = full siblings or parent-offspring, 0.25 = half-siblings, 0.125 = first cousins etc.) were then produced by doubling kinship coefficients (Wright 2022) and a heatmap was produced to visualise relatedness of individuals. Using the ‘LEA’ R package (Frichot and François 2015), cross-entropy Delta-K values were plotted to assess the optimal number of ancestral populations. Admixture plots were also created using the snmf() function in ‘LEA’ to visualise individual admixture coefficients from a genotypic matrix. The sparse nonnegative matrix factorisation (sNMF) provides least-squares estimates of ancestry proportions assuming K ancestral populations (Frichot and François 2015). This strategy estimated an entropy criterion that provides an optimal fit of the statistical model to the data for factors ranging from K = 1 to K = 5 for 10 repetitions.
Results
A total of 18,655 loci were characterised as part of the DarTseq™ approach. During the filtering process a total of 15,624 loci were excluded due to non-conforming minor allele frequencies, call rate and repeatability, leading to a final dataset of 3,031 loci for population analysis. Of the 70 individuals sequenced, a total of 59 individuals were retained from four locations (Bundaberg n = 27, Townsville n = 23, Mission Beach n = 5, and Princess Charlotte Bay n = 4) after quality control and filtering.
A lack of pairwise genetic differentiation was observed between locations (mean FST = − 0.005), with no significant p-values observed (p > 0.05). Negative FST values were found between all paired locations except Townsville–Bundaberg (FST = 0.0008), possibly reflecting greater within-group variation than inter-group variation for the small sample sizes from Princess Charlotte Bay and Mission Beach (Table 1). No significant deviations in heterozygosity were found in any location (HE = 0.1545–0.0.1818; HO = 0.1599–0.1634, Table S2 Supplementary Material).
The PCA plot indicated no population structuring or differentiation between locations in the dataset, with all individuals centred around a single cluster in which ellipses overlap (Fig. 3). The DAPC plot confirmed a lack of population structuring with overlapping ellipses for all four groups (Fig. S1 Supplementary material). Coefficients of relatedness (r) ranged from 0 to 0.062 with 28 pairs of third cousins identified (r = 0.03125–0.0624) spread throughout all sampled regions (Fig. 4). Delta-K cross-entropy analysis indicated no clear contrast between values of K = 1 to K = 5, supporting a lack of population structure and a single likely ancestral population (Fig. S2 Supplementary material). LEA plots of genetic admixture for cluster ranges from K = 2 to K = 4 also indicated mixing throughout all populations with no population structuring (Fig. S3 Supplementary material).

Principal Component Analysis plot of genetic similarity among individual T. australiensis from the four sampled regions. Ellipses represent variance around group centroids

Heatmap matrix of pairwise coefficients of relatedness (r) with individuals grouped by sampling locations (‘Populations’). For full sibling or parent–offspring pairs: r = 0.5, half-siblings: r = 0.25, first cousins: r = 0.125 and so on. Third cousins (r = 0.03125) were the closest relationship identified in this study. Where r = 0, pairs are unrelated. PCB = Princess Charlotte Bay. Sampling ID of individuals are given on the x and y axes
Discussion
This study provides the first findings on population connectivity in T. australiensis throughout the ECOTF on Australia’s east coast, revealing high gene flow within the population and a single well-mixed stock. The four locations assessed cover the main fishing grounds for T. australiensis, particularly the important fishing grounds off Bundaberg and Townsville that yield ~ 80% of Reef Bug landings. The strong population clustering and small genetic differences observed between populations support strong genetic connectivity throughout the study area. The occurrence of related individuals in populations with wide geographic separation, e.g. third cousins between Bundaberg and Princess Charlotte Bay further support broad-scale genetic population connectivity. Mark-recapture studies of adult T. australiensis have found short-range adult movements insufficient to support the high degree of genetic mixing observed here among widely dispersed populations (Jones 1988; McMillan et al. 2023). The observed genetic population connectivity, therefore, likely results from larval dispersal processes.
Similar well-mixed genetic connectivity at broad spatial scales has been observed in many lobster species. Like T. australiensis, the congeneric shovel-nosed lobster Thenus unimaculatus (Burton & Davie, 2007) displayed a lack of population structuring over a likewise large geographic area spanning the east and west coasts of India (Jeena et al. 2016). Other lobsters have also demonstrated comparable instances of high gene flow over large distances, including the Ornate Spiny Lobster Panulirus ornatus (Fabricius, 1798) over a similar geographic area as examined here (Queensland’s east coast to the Torres Strait: Farhadi et al. 2022), as well as the Spiny Lobster Panulirus argus (Latreille, 1804) in the Gulf of Mexico and the Caribbean (Silberman et al. 1994), and the European Lobster Homarus gammarus (Linnaeus, 1758) across large areas of the Northeast Atlantic (Ellis et al. 2017; Jenkins et al. 2019). These species all share long pelagic larval durations (PLDs) that likely facilitate high gene flow over such large distances.
The PLD of T. australiensis extends up to 45 days, transitioning through four phyllosomal instars during which the transparent larvae grow from ~ 4 mm to ~ 20 mm total length before developing into a post-larval nisto instar prior to settlement on the seafloor (Barnett et al. 1984; Mikami and Greenwood 1997). All larval stages involve passive dispersal, i.e. lacking pleopods to assist motility and dependent on environmental transport by ocean currents (Barnett et al. 1984). Despite this lack of motility, T. australiensis appears to utilise current regimes at favourable depth strata to maintain position in preferred coastal habitats, avoiding advection to oceanic areas where survival to recruitment is unlikely (Booth et al. 2005).
The occurrence of related individuals between Princess Charlotte Bay (PCB) and other sampled locations is intriguing because PCB lies north of the bifurcation of the west flowing North Vanuatu Jet where it approaches the Great Barrier Reef and forms the equatorward flowing Gulf of Papua Current and the poleward flowing East Australia Current between ~ 14.4–14.7° S (Ridgway et al. 2018). However, larval dispersal between PCB and the rest of the population may be facilitated intermittently by the mediating effects of local forcing (e.g. tide and wind) on these currents and seasonal variability in the velocity and direction of flows over the inner shelf within the Great Barrier Reef lagoon (Benthuysen et al. 2022). Year-round spawning in combination with the relatively long PLD of T. australiensis are, therefore, likely to be important in facilitating broad-scale larval dispersal and genetic population connectivity throughout the study area.
Other Scyllaridae have comparable PLDs to T. australiensis, though in some species PLD extends to several months (Booth et al. 2005). Despite this, the ~ 45-day dispersal of T. australiensis larvae appears sufficient to enable thorough mixing throughout their range on Australia’s east coast due to the extent and complexity of current regimes in the study area. The Spanner Crab Ranina ranina (Linnaeus, 1758) overlaps with the southern distribution of T. australiensis in Queensland and displays a similar PLD lasting 36–62 days (Minagawa 1990). During this short period, larvae spawned primarily in central Queensland supply the entire distribution of the species with recruits, dispersing as far as northern New South Wales (Schilling et al. 2022). Physical forcing of current regimes on Queensland’s east coast, therefore, appears sufficient to drive larval transport over large areas during PLDs similar to that of T. australiensis.
Conclusion
A lack of genetic population structuring was observed in T. australiensis on the Australian east coast, supporting assessment and monitoring as a homogeneous management unit throughout the ECOTF. High gene flow and population connectivity among locations over a broad scale likely resulted from pelagic larval dispersal due to the limited movements of adult T. australiensis. Better understanding of the underlying physical processes driving the large-scale dispersal and thorough mixing of T. australiensis among regions may be achieved by particle dispersal modelling. Our findings of a single well-mixed biological stock of T. australiensis on Queensland’s east coast will inform the first stock assessment for this species as well as future management measures.
Data availability
The datasets generated during and/or analysed during the current study are available from the corresponding author on reasonable request.
References
Altukhov, Y. P., 1981. The stock concept from the viewpoint of population genetics. Canadian Journal of Fisheries and Aquatic Sciences 38(12): 1523–1538.
Barnett, B., R. Hartwick & N. Milward, 1984. Phyllosoma and nisto stage of the Morton Bay Bug, Thenus orientalis (Lund)(Crustacea: Decapoda: Scyllaridae), from shelf waters of the Great Barrier Reef. Marine and Freshwater Research 35(2): 143–152.
Begg, G. A., K. D. Friedland & J. B. Pearce, 1999. Stock identification and its role in stock assessment and fisheries management: an overview. Fisheries Research 43(1–3): 1–8.
Bell, R. S., P. W. Channells, J. W. MacFarlane, R. Moore & B. F. Phillips, 1987. Movements and breeding of the ornate rock lobster, Panulirus ornatus, in Torres Strait and on the north-east coast of Queensland. Australian Journal of Marine and Freshwster Research 38(2): 197–210.
Benthuysen, J. A., M. J. Emslie, L. M. Currey-Randall, A. J. Cheal & M. R. Heupel, 2022. Oceanographic influences on reef fish assemblages along the Great Barrier Reef. Progress in Oceanography 208: 102901.
Booth, J. D., W. R. Webber, H. Sekiguchi & E. Coutures, 2005. Diverse larval recruitment strategies within the Scyllaridae. New Zealand Journal of Marine and Freshwater Research 39(3): 581–592.
Cowen, R. K. & S. Sponaugle, 2009. Larval dispersal and marine population connectivity. Annual Review of Marine Science 1: 443–466.
Crawford, D. L. & M. F. Oleksiak, 2016. Ecological population genomics in the marine environment. Briefings in Functional Genomics 15(5): 342–351.
Ellis, C. D., D. J. Hodgson, C. L. Daniels, M. Collins & A. G. Griffiths, 2017. Population genetic structure in European lobsters: implications for connectivity, diversity and hatchery stocking. Marine Ecology Progress Series 563: 123–137.
Farhadi, A., F. Pichlmueller, B. Yellapu, S. Lavery & A. Jeffs, 2022. Genome-wide SNPs reveal fine-scale genetic structure in ornate spiny lobster Panulirus ornatus throughout Indo-West Pacific Ocean. ICES Journal of Marine Science 79(6): 1931–1941.
Frichot, E. & O. François, 2015. LEA: an R package for landscape and ecological association studies. Methods in Ecology and Evolution 6(8): 925–929.
Funk, W. C., J. K. McKay, P. A. Hohenlohe & F. W. Allendorf, 2012. Harnessing genomics for delineating conservation units. Trends in Ecology & Evolution 27(9): 489–496.
Gardner, C., S. D. Frusher, M. Haddon & C. Buxton, 2003. Movements of the southern rock lobster Jasus edwardsii in Tasmania. Australian Bulletin of Marine Science 73: 653–671.
Georges, A., B. Gruber, G. B. Pauly, D. White, M. Adams, M. J. Young, A. Kilian, X. Zhang, H. B. Shaffer & P. J. Unmack, 2018. Genomewide SNP markers breathe new life into phylogeography and species delimitation for the problematic short-necked turtles (Chelidae: Emydura) of eastern Australia. Molecular Ecology 27(24): 5195–5213.
Gruber, B., P. J. Unmack, O. F. Berry & A. Georges, 2018. Dartr: An r package to facilitate analysis of SNP data generated from reduced representation genome sequencing. Molecular Ecology Resources 18(3): 691–699.
Jeena, N., A. Gopalakrishnan, J. K. Kizhakudan, E. Radhakrishnan, R. Kumar & P. Asokan, 2016. Population genetic structure of the shovel-nosed lobster Thenus unimaculatus (Decapoda, Scyllaridae) in Indian waters based on RAPD and mitochondrial gene sequences. Hydrobiologia 766: 225–236.
Jenkins, T. L., C. D. Ellis, A. Triantafyllidis & J. R. Stevens, 2019. Single nucleotide polymorphisms reveal a genetic cline across the north-east Atlantic and enable powerful population assignment in the European lobster. Evolutionary Applications 12(10): 1881–1899.
Jennings, S. & M. J. Kaiser, 1998. The effects of fishing on marine ecosystems, Advances in Marine Biology, Vol. 34. Elsevier, Amsterdam: 201–352.
Jones, C. M., 1988. The biology and behaviour of bay lobsters, Thenus spp.(Decapoda: Scyllaridae), in northern Queensland, Australia. PhD thesis, University of Queensland, Brisbane, Australia.
Kilian, A., P. Wenzl, E. Huttner, J. Carling, L. Xia, H. Blois, V. Caig, K. Heller-Uszynska, D. Jaccoud & C. Hopper, 2012. Diversity arrays technology: a generic genome profiling technology on open platforms. Data production and analysis in population genomics: Methods and protocols. Methods in Molecular Biology 2012(888): 67–89.
Kingsford, M. J., J. M. Leis, A. Shanks, K. C. Lindeman, S. G. Morgan & J. Pineda, 2002. Sensory environments, larval abilities and local self-recruitment. Bulletin of Marine Science 70(1): 309–340.
McMillan, M., S. Leahy, J. Daniell, N. Louw, E. Roberts, M. Wickens, K. Hillcoat & M. F. O’Neill, 2023. Determining the spatial distribution and abundance indices for Moreton Bay Bugs, Thenus parindicus and Thenus australiensis in Queensland to improve stock assessment and management. Fisheries Research and Development Corporation Report 2020–020. In Press.
Metaxas, A. & M. Saunders, 2009. Quantifying the “bio-” components in biophysical models of larval transport in marine benthic invertebrates: advances and pitfalls. The Biological Bulletin 216(3): 257–272.
Mikami, S. & J. G. Greenwood, 1997. Complete development and comparative morphology of larval Thenus orientalis and Thenus sp.(Decapoda: Scyllaridae) reared in the laboratory. Journal of Crustacean Biology 17(2): 289–308.
Minagawa, M., 1990. Complete larval development of the red frog crab Ranina ranina (Crustacea, Decapoda, Raninidae) reared in the laboratory. Nippon Suisan Gakkaishi 56(4): 577–589.
Ovenden, J. R., O. Berry, D. J. Welch, R. C. Buckworth & C. M. Dichmont, 2015. Ocean’s eleven: a critical evaluation of the role of population, evolutionary and molecular genetics in the management of wild fisheries. Fish and Fisheries 16(1): 125–159.
Palsbøll, P. J., M. Berube & F. W. Allendorf, 2007. Identification of management units using population genetic data. Trends in Ecology & Evolution 22(1): 11–16.
R Core Team, 2021. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. URL https://www.R-project.org/.
Ricker, W., 1981. Changes in the average size and average age of Pacific salmon. Canadian Journal of Fisheries and Aquatic Sciences 38(12): 1636–1656.
Ridgway, K. R., J. A. Benthuysen & C. Steinberg, 2018. Closing the gap between the Coral Sea and the equator: direct observations of the north Australian western boundary currents. Journal of Geophysical Research: Oceans 123(12): 9212–9231.
Schilling, H. T., D. E. Hewitt, N. Malan, M. D. Taylor & D. D. Johnson, 2022. Cross-jurisdictional larval supply essential for eastern Australian spanner crabs (Ranina ranina). Marine and Freshwater Research 73(11): 1352–1367.
Silberman, J., S. Sarver & P. Walsh, 1994. Mitochondrial DNA variation and population structure in the spiny lobster Panulirus argus. Marine Biology 120: 601–608.
Silva, C. N., C. Villacorta-Rath, L. N. Woodings, N. P. Murphy, B. S. Green, K. Hartmann, C. Gardner, J. J. Bell & J. M. Strugnell, 2019. Advancing our understanding of the connectivity, evolution and management of marine lobsters through genetics. Reviews in Fish Biology and Fisheries 29: 669–687.
Smith, P., R. Francis & M. McVeagh, 1991. Loss of genetic diversity due to fishing pressure. Fisheries Research 10(3–4): 309–316.
Waples, R. S., A. E. Punt & J. M. Cope, 2008. Integrating genetic data into management of marine resources: how can we do it better? Fish and Fisheries 9(4): 423–449.
Woodings, L. N., N. P. Murphy, S. R. Doyle, N. E. Hall, A. J. Robinson, G. W. Liggins, B. S. Green, I. R. Cooke, J. J. Bell & J. M. Strugnell, 2018. Outlier SNPs detect weak regional structure against a background of genetic homogeneity in the Eastern Rock Lobster, Sagmariasus verreauxi. Marine Biology 165: 1–17.
Wright, S., 1922. Coefficients of inbreeding and relationship. The American Naturalist 56(645): 330–338.
Zheng, X., D. Levine, J. Shen, S. M. Gogarten, C. Laurie & B. S. Weir, 2012. A high-performance computing toolset for relatedness and principal component analysis of SNP data. Bioinformatics 28(24): 3326–3328.
Acknowledgements
This work was funded by the Fisheries Research and Development Corporation (FRDC) as part of project 2020-020 ‘Determining the spatial distribution and abundance indices for Moreton Bay Bugs, Thenus parindicus and Thenus australiensis in Queensland to improve stock assessment and management’. The authors thank commercial fishers Neil Mogensen and Andrew Redfearn for their assistance in sample collection.
Funding
Open Access funding enabled and organized by CAUL and its Member Institutions.
Author information
Authors and Affiliations
Contributions
MNM was responsible for conceptualisation, administration, methodology, sample collection and processing, investigation, and writing. NB was involved in sample processing, methodology, formal analysis, and writing. SMW contributed to conceptualisation, methodology, review, and editing. BJH was involved in supervision, editing and review.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Additional information
Handling Editor: Diego Fontaneto
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
McMillan, M.N., Brunjes, N., Williams, S.M. et al. Broad-scale genetic population connectivity in the Moreton Bay Bug (Thenus australiensis) on Australia’s east coast. Hydrobiologia 851, 2347–2355 (2024). https://doi.org/10.1007/s10750-023-05460-8
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-023-05460-8