
Abstract
Fisheries of the Mekong Basin are crucial to regional food security. They rely on seasonal monsoon rains that inundate rice fields and reconnect water bodies. Species assemblages vary over time and space, but infrastructure developments and climate change are negatively impacting the timing and magnitude of the flood pulse as well as fish migration routes and populations. In lowland rice field landscapes of Cambodia, community fish refuges (CFRs) are used to maintain fish abundance and biodiversity, with varying success. There is little knowledge of the drivers of this variation to guide management interventions. We used beta diversity to explore the effectiveness of 40 CFRs in maintaining species abundance and diversity during the dry seasons from 2012 to 2015. Results show CFR connectivity to the floodplain is important and suggest the type of inlet/outlet influences fish retention. CFRs connected to a large area of rice fields during the wet season had lower losses in species abundance through the dry season. However, large CFRs, or those part of a larger body of water, had higher losses in species abundance. Key design features of CFRs must be coupled with strong management capacity to guide the conservation and fisheries strategy in the Mekong Basin.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
As of 2015, inland fisheries represented 12% of total global capture fisheries production (Funge-Smith, 2018). They represent a substantial and important aquatic food system mostly concentrated in the tropics, which is under continual threat of reduction from power and agriculture needs for water. The capture of wild fish from seasonally flooded rice fields and associated water bodies such as canals, ponds and ditches are collectively termed as “rice field fisheries” (Gregory, 1997), and represent a key food system that crosses the land–water interface. In Cambodia, rice field fisheries contribute substantially to local household food security (Freed et al., 2020). Freshwater fish consumption in Cambodia is among the highest in the world (Hortle et al., 2004; Funge-Smith, 2018) with variation in annual rates by province (from 43.4 to 105.2 kg per capita) depending on proximity to riparian areas and household participation in fisheries (Hortle, 2007).
The majority of Cambodia’s rice fields are rain-fed systems during flood season, and the fish stocks therein are sustained through natural recruitment by freely moving across a variety of aquatic habitats that have remained largely connected, which is a key factor influencing the diversity of fish assemblages and biomass (Welcomme, 1995). However, increasing demand for water resources, coupled with climate-related changes in precipitation and temperature, has diverted and decreased the flow of water (de Wit & Stankiewicz, 2006; Bauer & Scholz, 2015) leading to reduced movement and dispersal of wild aquatic species.
Following recommendations from a study by Gregory (1997), various projects have aimed to enhance the stock and productivity of rice field fisheries by establishing and/or enhancing dry season no-take refuges for brood fish in seasonally inundated rice field landscapes. Specifically, Community Fish Refuges (CFRs) can be constructed reservoirs, a demarcated part of a larger water body, or an entire natural water body that holds water throughout the year (but may also become disconnected from the rest of the floodplain). They are under community ownership and management and are formally recognized and enacted in Cambodian fisheries law and policy. During the dry season, most CFRs become disconnected from the rest of the floodplain and other natural water bodies. Then in the flood season, they are reconnected to these water bodies and to large areas of the seasonally flooded rice fields. The key function of a CFR is to maintain its fish population and diversity through the dry season until the flooding creates seasonal habitat and allows fish to move to areas suitable for feeding and reproduction. During the flood season, fish from CFRs can spawn and feed in the inundated rice fields (Joffre et al., 2012). Fish guilds of the Lower Mekong are largely grouped by migration habits and oxygen requirements: ‘black’ fish are predominantly resident floodplain species that tolerate low-oxygen levels, ‘white’ fish are highly migratory and are sensitive to low oxygen, and ‘grey’ fish are short distance migrators with some tolerance to low oxygen (Welcomme, 2001). Given the varying tolerance of different fish species, the CFR attributes (size, water quality, availability of physical shelter, etc.) and especially its connectivity are likely to affect the species composition of the local fish assemblage and the persistence of certain species over time.
Despite the rich literature on the annual flood cycle and floodplain fisheries in Cambodia (e.g. Junk et al., 1989; Kummu et al., 2008; Kong et al., 2017; Sabo et al., 2017), there has been very limited work on the changes in species abundance and diversity through seasons, or whether CFRs and other perennial aquatic habitats influence the magnitude of seasonal variation in fish assemblages. Yet, it is important to understand these dynamics to inform best management practices and guidance for the CFR programme in Cambodia, and to ensure the diversity and viability of rice field fisheries within broader aquatic food systems development.
Temporal beta diversity is defined as the change in community composition through time and can be measured using data from repeated surveys (Legendre and Gauthier, 2014). Analysis of temporal beta diversity can be used to investigate the effects of environmental change (both natural disturbances and human impacts) on community composition (Legendre 2019). This type of research has been conducted to understand natural patterns of seasonal change in fish and aquatic communities (e.g. Penha et al., 2017; Petsch et al., 2021) and to recommend conservation approaches for fish and aquatic species (e.g. Langer et al., 2016; Dong et al., 2021). Similar research has been conducted for open water fisheries in Cambodia, with important findings on the drivers of seasonal changes in species composition. For example, in a recent study of spatial and temporal variations in fish species assemblages in the Tonle Sap Lake (TSL), Kong et al. (2017) concluded that fish species composition is heterogeneously distributed in space and time, which is likely to be due to spatial variation in environmental conditions and the seasonal migration of particular species reliant on habitat connectivity for their reproduction and survival. Further relevant observations have been made in other floodplain systems, such as the influence of connectivity to perennial large water bodies on local fish diversity (Jiang et al., 2021) and the influence of area and connectivity of local habitats in maintaining fish diversity within floodplain sites (Liu & Wang, 2010).
Our main objective was to assess changes in community composition during the dry season because the faunal communities that remain at the end of the dry season and are re-assembled in the wet season are the foundation of the fisheries diversity. We tested the following hypotheses related to CFR performance in maintaining fish populations: (1) Perennial connection of CFRs (e.g. those prone to flooding and/or in larger water bodies) and the length of time since CFR establishment will positively influence CFR beta diversity. (2) Beta diversity will be negatively correlated with the distance to the Tonle Sap Lake, the largest perennial water body and reservoir of most fish species present in the ecosystem. (3) A CFR’s area and connectedness to larger water bodies influence its performance in maintaining biodiversity and fish abundance through the dry season.
Methods
Study area background
The Tonle Sap and its floodplain in the Mekong Basin in Cambodia represent the 5th most productive inland fishery in the world with an annual production of ~ 4–500,000 metric tonnes almost entirely from natural recruitment of wild fish (Funge-Smith, 2018). This productivity is driven by seasonal rainfall fluctuations and resulting nutrient dynamics and primary productivity of the floodplain, known as the “flood pulse” (Junk et al., 1989). Previous research using catch monitoring data from the large river channels and the TSL has highlighted the importance of the flood pulse in fisheries productivity (Sabo et al., 2017) and the overall decline in the medium to large-sized fish species in the catch over 15 years (Ngor et al., 2018a).
Approximately, 30% of the total land area of Cambodia is subjected to seasonal flooding, fed directly by rainfall or by the associated flooding of lakes and rivers, lasting from a few weeks to 6 months. Much of these seasonally flooded areas are used for rice cultivation and rice field fisheries (Chheng et al., 2016). These rice field fisheries are open to all fishers during flood season, as long as non-commercial gear is used. Aquatic life in the flooded rice cultivation areas is a common pool resource, regardless of land ownership. Small fish (bream, cyprinids, gourami) caught by artisanal fishers throughout the year are a major source of food security for households in riparian and floodplain areas (Shams, 2007; Garaway et al., 2013; Tweddle et al., 2015), and larger predatory fish (e.g. snakehead and catfish) are both consumed and sold for income (Hortle et al., 2008; Freed et al., 2020).
The flood pulse of the TSL has been affected by development of land and waters in the upstream Mekong and its catchment basin (De Xun Chua et al., 2022). As a result, the productivity of Mekong floodplains may decline, calling for management innovations to mitigate the loss of floodplain ecosystem services, including fisheries (Kummu & Sarkkula, 2008; Dugan et al., 2010) and regional species genetic pools (Arrington & Winemiller, 2003; Food and Agriculture Organization of the United Nations, 2019).
Study sites and CFR characterization
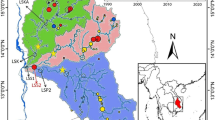
This study focuses on 40 CFRs surrounding the TSL that were selected in 2012 to receive interventions including biophysical modifications and strengthening of the management committee operations (Fig. 1). The CFRs are located in the provinces of Pursat, Battambang, Siem Reap and Kampong Thom.

Map of 40 community fish refuges (red dots) around the Tonle Sap Lake in Cambodia
The Cambodian Fisheries Administration (FiA) divides water bodies that can house CFRs into two main categories: (i) a community pond surrounded by rice fields, and (ii) a conservation area within a large water body that is surrounded by rice fields. However, for the purpose of better management and interventions, the CFRs were further classified into four categories (Brooks et al., 2015): (1) irrigation reservoir, (2) community pond within agricultural land not prone to flooding, (3) community pond within agricultural land prone to flooding and (4) a demarcated area within a larger water body.
Well-functioning CFRs are managed by local communities to enhance fish productivity and biodiversity by ensuring aquatic habitat remains throughout the dry season and protecting a community of fishes from fishing mortality year-round (Fig. 2). Communities undertake various habitat improvement interventions with the support of WorldFish and other NGO partners. This study reports findings based on the first phase of the Rice Field Fisheries Enhancement Project that supported the 40 CFRs, funded by USAID from 2012 to 2016.

Reproduced with permission from Joffre et al. (2012)
A community fish refuge in different seasons. ©WorldFish.
Water availability and flooding vary seasonally at the sites. The dry season occurs from January to May, and floods occur from July to November (Freed et al., 2020). The average water depth in the study site rice fields was 0.1 cm and 18 cm during dry and flood seasons, respectively.
Data collected during the site selection process and used in analyses for this study included water body area (included the total water body for CFRs that were a demarcated area within a larger water body), distance from the CFR to the nearest village, and the rice field area connected to the CFR in the wet season (the area was determined during the delineation of the rice field ecosystem fishing ground area that was described in Freed et al., 2020).
Monitoring protocol
Fish biodiversity and environmental variables were sampled at 3-month intervals at the 40 CFR sites from November 2012 to November 2015. The sampling methods are described in detail in Fiorella et al. (2019). Standardised sampling was conducted using gill nets four times per year (13 sampling occasions in total). Depending on the size and depth of the CFR, five to eight gill nets were used during a sampling occasion. The sample consisted of a maximum of 8 gill nets, 2.5 m in height, composed of 1.5 or 3 m long panels of mesh sizes of 1.5, 2, 4, 6, 8 and 10 cm. The number and length of gill nets were adjusted to seasonal variations in CFR water depth and bottom structure, to minimise excessive sampling in small water bodies. Each gill net was set perpendicular to the shoreline, with a minimum of 20 m between gill nets. The submerged net area was calculated per set using the water depth at each end of the net, and the length of the net. Gill nets were set between 16:00 and 17:00 h and remained in the water for 12–14 h. Captured fish were grouped by species and the number of individuals, the minimum and maximum fork length, and the minimum and maximum weight, and total weight per species were recorded.
Data analysis
We used the temporal beta diversity index (TBI; Legendre, 2019) to understand the changes in community composition between early and late dry season. Specifically, for each CFR, we measured the dissimilarity in community composition (described by a matrix of species by sites) between February and May in the same year. We employed one matrix to represent the three occasions in February and a separate matrix for the occasions in May. In other words, each combination of refuge and year was considered as a distinct site in the analysis.
We assembled the community composition matrices through a measure of catch per unit effort (CPUE). We summed the number of individuals captured within a sampling occasion for each refuge and then standardised the values to account for the number of gill nets used in the sample. Because there were large differences across the abundances of different species, CPUE data were log + 1 transformed.
We used the percentage difference coefficient (also known as Bray–Curtis dissimilarity) to calculate the temporal beta diversity index. We used 9,999 permutations to test the significance of the temporal beta diversity indices in each site. Specifically, we examined if any site-year combination showed significant changes in composition during the dry season.
Mean differences in beta diversity
The temporal beta diversity index proposed by Legendre (2019) can be decomposed into gains and losses of abundances-per-species or gains and losses of species (when using occurrence data). ‘Species loss’ refers to a species being absent from a repeat survey, not that it is locally extinct. In the same way, species gain is where it was once absent but then appears in a subsequent sampling occasion. We used a Bayesian regression to examine the extent to which differences in community composition are driven predominantly by gains or losses. We were also interested in testing whether gains and losses differ across refuge categories. To that end, we used a multivariate model in which the response variables were the percentage difference due to gains and due to losses (Ieno et al., 2007). In addition, to control for unmeasured differences across CFRs and years, we calculated a group-level intercept for each of these levels. Using a multivariate model, as opposed to two separate models, allow us to acknowledge that gains across refuges and years (as well as their variability) is correlated with losses. Furthermore, a multivariate model provides the residual correlation between response variables which assess whether there is an unmodelled dependency between the response variables (Zuur et al. 2009). The formula notation for this regression is
Percentage differences were modelled using a zero inflated beta distribution. The model was fitted using four Markov chains with 5,000 iterations each; half of those as a warm-up for the Hamiltonian Monte Carlo method (Hoerl and Snee, 2015). We used weakly informative priors for all parameters. We performed a Moran’s test (Moran, 1950) using 999 permutations to test for residual spatial autocorrelation and found that residuals were spatially independent (Moran’s I = − 0.004; P = 0.298).
Covariates of beta diversity
We investigated how the sampling occasion and refuge covariates were associated with the differences in community composition as measured by both the gain and loss components of temporal beta diversity. We performed a PCA to identify any collinearity among parameters and detect the most relevant parameters in describing the data. After discarding collinear variables and selecting only those that represented variation in the data set, we selected refuge-level covariates of TBI. These covariates were as follows: (1) the area of the whole water body, (2) the proportion of area covered by aquatic plants in February, (3) the CFR depth in May, (4) the depth difference between May and February, (5) the distance of the CFR to the nearest village, (6) the rice field area connected in the wet season and (7) the type of inlets/outlets that connect the CFR to the rice field and the area of the refuge’s water body. The reason for selecting these covariates was to account for: the potential of fish in the CFRs to evade predation and/or sampling (covariates 1–3), the severity of drought and/or water use affecting the fish habitat (covariate 4), the remoteness and level of difficulty to monitor the CFR for poachers (covariate 5), and the ease and/or likelihood of fish species returning to the CFR at the start of the dry season (covariates 6 and 7). Inlet/outlet types were earthen channels or ditches, or culverts made from a pipe or reinforced concrete or other material (and some CFRs connect to rice fields without the need for artificially constructed inlet/outlets). We did not include the CFR category as a covariate for two main reasons. First, in this model we were mainly interested in the site characteristics that may drive differences in beta diversity. Second, these site characteristics and the CFR categories are not independent from each other, and including both would invalidate the assumptions of our modelling framework.
Distances and area variables were log-transformed, and numerical variables were scaled to have a mean of zero and unit standard deviation. We modelled response variables (i.e. gains and losses in a multivariate model), hierarchy structure (random effects for refuge and year), priors and model chains in the same way as in the model in which we compare mean differences in beta diversity.
We calculated the temporal beta diversity index and the species’ paired t tests using the implementations in the R package ‘adespatial’ 0.3-8. Bayesian models were implemented using ‘brms’ 2.14.4 (Bürkner, 2017). All analyses were performed in R version 4.0.2 (R Core Team 2020). Code is available at https://github.com/WorldFishCenter/cambodian-fish-refuges.
Differences across species types
We also looked at the role that different species may play in the change in temporal beta diversity. Specifically, we computed paired t tests of differences between the abundance observed at the beginning and end of the dry season (February and May) for each species. We used the same community composition matrices as for the computation of the temporal beta diversity index. That is, keeping each refuge–year combination as an independent site in the analysis. We used 9,999 permutations to test statistical significance and used the Holm method to correct for multiple testing. We then computed t tests for the total abundance of each group of species (black, grey, white and other species) at the beginning and end of the dry season (February and May). To do this, we aggregated the abundance of all species within each group at the beginning and end of the dry season and then conducted a t test to check for significant difference in the total abundance of each group.
Results
Across the 40 CFR sites over three years sampling, 272,032 individuals were sampled belonging to 161 species in 82 genera and 35 families. The total number of species captured by site varied from the low of 33 (Boeng Tramses, Cat. 4) to the high of 85 (Tumnub Kandole, Cat. 1 and Boeng Rolum, Cat. 4), while the total number of individuals by site across all sampling ranged from 2,379 (Boeng Kampeng, Cat. 4) to 12,509 (Damnak Kranh, Cat. 1). In dry season sampling (February and May) across all CFRs and years, six species accounted for over 50% of individuals sampled: Parambassis siamensis (Fowler 1937; glass fish), Esomus metallicus (Ahl 1923; striped flying barb), Cyclocheilichthys lagleri (Sontirat, 1989; Lagler carp), Puntius cf. masyai (barb species), Puntius brevis (Bleeker, 1849; swamp barb) and Parambassis apogonoides (Bleeker 1851; Iridescent glassy perchlet).
The community composition of CFRs between February and May was quite different, and this difference was, generally, due to losses in per-species abundance during this period. The mean total dissimilarity (measured using the percentage difference) ranged between 50% [28%, 72%] (mean and 95% credible intervals) for CFRs in ponds within agricultural land not prone to flooding (Cat. 2) and 57% [34%, 77%] for CFRs in reservoirs for irrigation (Cat. 1) (Fig. 3, top panel). When partitioning these dissimilarities into those due to losses and gains in species abundances, we found that dissimilarity losses account for a larger amount than gains across all CFR categories (probability of direction, p.d., 0.95; Fig. 3, middle and bottom panels). When looking at the random effects, we also found that there is no more substantial difference between the variability of beta diversity index across CFRs than across years (Supplementary Fig. 2). In addition, none of the site-year combinations changed in exceptional ways when compared to all others (all P values of the temporal beta diversity index corrected for multiple testing were larger than 0.05). In other words, the length of time since initial CFR interventions (since 2012) did not seem to affect the pattern of community composition change.

Percentage total dissimilarity and dissimilarity due to gains and losses according to CFR type (category)
The model that explored covariates of beta diversity had a R2 of 0.36 [0.21, 0.49] for the dissimilarity in per-species abundance due to losses and 0.2 [0.09, 0.33] for the dissimilarity due to gains. From the variables we analysed, we found that three of them were likely associated with either of the response variables: the channel type (a categorical variable), the area of the water body (positively correlated with percent dissimilarity due to species abundance losses) and the area of the rice field connected during the wet season (negatively correlated with percent dissimilarity due to species abundance losses). CFRs connected to the adjacent rice fields through culverts were less likely to experience species abundance losses during the dry season particularly when compared to CFRs without channels or those with earth channels (p.d. 0.96 and 0.72; Fig. 4A). Similarly, CFRs with culverts experienced larger species abundance gains when compared to CFRs without channels (p.d. 0.999).

Percentage dissimilarities: A due to species abundance losses (blue) and gains (green) in CFRs with different types of inlet/outlet connections to adjacent rice fields, B due to species abundance losses in correlation with (log) area of the CFR water body, and C due to species abundance losses in correlation with area of rice field connected during the wet season
We found that the (log) area of the CFR water body had a positive association with the dissimilarities due to species abundance losses (p.d. 0.99; Fig. 4B). In contrast, the rice field area connected to the refuge during the wet season had a negative association with losses (p.d. 0.99; Fig. 4C). In addition, we also found some evidence for negative association between the refuge’s area at the end of the dry season and the loss of species abundance as well as a negative association between the refuge’s distance to the nearest village and the species abundance gains. However, the effect sizes of these associations were considerably smaller (Supplementary Fig. 2).
We found that out of the 149 species sampled during the dry season, seven species showed significant (log) differences in their abundances (namely Puntius cf. masyai, Pristolepis fasciata, Trichogaster microlepis, Notopterus notopterus, Hampala macrolepidota, Labiobarbus siamensis and Parambassis apogonoides (Table 1). Another 24 species showed important differences but were not considered significant at the 0.05 level after correcting for multiple testing (Table 1). The average difference in log species abundances between the end and the beginning of the dry season was − 0.63, − 0.74 and 0.03 for black, grey and white species, respectively.
Discussion
The range of species richness (33–85 species) and number of individuals (2,379–12,509) found across the 40 CFRs sites illustrates that these systems support and harbour the natural species diversity available throughout the TSL region (with estimates of between 149 and 296 fish species having been recorded in the Tonle Sap Lake Campbell et al., 2006; Allen et al., 2012; Baran et al., 2013). The differences seen in community composition of CFRs between February and May were more due to fish being lost from CFRs than fish being gained. These differences were generally due to species abundance losses during the dry season period rather than a reduction in species richness (‘whole’ populations disappearing from samples). The six species that accounted for over 50% of individuals sampled are all grey species, broadly categorised as short distance migrators with some tolerance to low oxygen (Welcomme et al., 2001). This result is at least partly due to the sampling method, which favours capture of non-predatory species and small-sized individuals. On average, black and grey fish species were more likely to experience losses in abundances than white fish species. Five of seven species that showed different abundances during the dry season were grey and were likely to have been trapped in the CFR, unable to migrate back to larger water bodies.
The performance of the model that explored covariates of beta diversity was remarkable given it did not consider the potentially large effect of species competitive and predatory interactions. However, the data did not support the first hypothesis. Neither CFR connection to perennial water bodies nor the length of time since initial CFR interventions significantly influenced CFR beta diversity. Beta diversity differences between CFRs were similar to the differences across time (years) (Supplementary Fig. 2). Three CFR attributes showed association with the gain and loss of species abundance: the channel type, the area of the water body and the area of the rice field connected during the wet season. While we provide some potential explanations for these associations, it is not yet clear whether these attributes are related to a more direct cause of gain and loss of species abundance, such as management measures including fishing prohibition during the dry season. This would be supported by existing evidence of the effect of management measures, especially community fundraising, on CFR biomass reported by Fiorella et al. (2019). Good management was also associated with improved water quality, and in turn increased species richness (Fiorella et al., 2019). In addition, anthropogenic changes in agroecosystems can have unexpected effects and decouple the correlation between beta diversity and environmental conditions (Dala-Corte et al., 2019). For example, water infrastructure developments can have substantial effects on seasonal flow patterns and drive towards assemblages dominated by generalist species (Ngor et al., 2018b).
Since beta diversity differences between CFRs appear to be substantial, we explore some potential explanations for this spatial variability. Fish behaviour may also drive the spatial variability of temporal beta diversity. There is a great body of literature evidencing the ability of fishes, from salmon (Clemens et al., 1939) to stingrays (Tilley et al., 2013), to navigate and return to a particular location. A recent study used otolith chemistry to show catfish migrated up to 1,400 km into the Mekong and to return to the same estuary (Vu et al., 2022), and work in Australia with Lates calcarifer and Neoarius leptaspis demonstrated “homing back to previously occupied dry season refugia during the wet-to-dry transition, even though other potential refugia were available in closer proximity to wet season activity areas” (Crook et al., 2020). The authors’ suggestion that it is this homing behaviour that might regulate temporal variation in fish community composition and abundance, supports Kong et al. (2017) and our own findings of spatial variation driving heterogeneity more than changes over time. Finally, the large differences of temporal beta diversity between CFRs would justify the “all-inclusive” conservation approach of CFRs throughout the landscape, rather than a “hotspot” approach with fewer conservation sites, as recommended by Langer et al. (2016) for ecosystems with unique localized species assemblages.
The hypothesis that distance to the Tonle Sap floodplain drives temporal beta diversity was not confirmed. However, distance alone is a relatively poor descriptor and other distance measures may respond better than linear distance (Landeiro et al. 2011). Half of the 40 CFRs lie inside the east and west bordering roads. The average distance of CFRs from the Tonle Sap Lake (mean ± SD) is 38.05 ± 15.6 km. The closest CFR lies 9 km away, while the furthest is located 72 km away. However, some CFRs may be close to the lake but higher up and out of the floodplain, i.e. less connected than expected by direct distance, whereas some may be far from the lake but close to other streams or large water bodies, thereby being more connected than expected by distance alone. Aside from distance from TSL, CFRs connected with a main river might have more grey and white fish that depend on oxygenated waters and short migration routes, but those far from rivers might have less.
In support of our third hypothesis, CFRs that were well connected with a large area of rice fields during the wet season had lower losses in species abundance through the dry season. This role of connectivity in species assemblages is supported from findings in wetland floodplains in Brazil (Penha et al., 2017). However, those CFRs that were large themselves, or were part of a larger body of water had higher losses in species abundance, suggesting that area alone is not positively associated with safeguarding abundant fish populations. This effect might be indicative of fishing where the water body is not protected as a CFR and may also be due to effects of multiple water uses in large water bodies, such as for rice irrigation, domestic water and livestock. Also, fish can disperse in the larger water bodies, while the smaller CFRs that lose both depth and area during the dry season will concentrate fish at higher densities, which may mask actual losses. There was some evidence that greater water depth in CFRs at the end of the dry season led to lower losses in fish abundance, which seems logical as this implies more available habitat, higher oxygenation and the presence of more ecological niches.
As might be expected, community fish refuges with culverts connecting them to neighbouring rice fields had higher species abundance gains than those without channels (Fig. 4A). The presence of culverts also had a positive effect in retaining fish through the dry season compared to CFRs without channels, or with only earthen channels. Field observations indicate that culverts were installed more often in CFRs that were poorly connected to the floodplain. In this sense, the type of inlet/outlet intervention correlates with the level of CFR connectivity. It is likely that the CFRs without channels or with earth channels are well enough connected to other water bodies that outmigration can still occur during the dry season, while CFRs with culverts are in areas with sparse water during the dry season and so would be more likely to retain or even attract fish searching for dry season habitat. In this case, CFRs earthen channels or no channels could gain more fish at the start of the dry season relative to the CFRs with culverts, and so then also lose more during the dry season (Fig. 4A). However, at the species level, the effects of various attributes of culvert and fishways such as substrate type (soft/hard, organic/artificial) roughness and length have been shown to be highly variable (Thorncraft et al., 2006; Goodrich et al., 2018). Explaining the mechanisms behind these field observations and species level effects were beyond the scope of this study, but could be further investigated through differences between flood and dry season data. Alternatively, culverts may again be merely an indicator of better management practices, where well-managed CFRs are more likely to have culverts, and hence show less fish losses for a variety of management-related reasons. This may also account for fish gains decreasing with increasing distance between the CFR and village, because the opposite interaction was not seen with distance for fish losses.
Limitations and future research
Our results suggest that small CFRs of at least 2 m depth that are well connected to surrounding rice fields, but are close to villages, are optimal for maintaining biodiversity. Local management practices are likely to be significant, and highly contextualised, and another study using the same dataset failed to draw significant conclusions based on biophysical modifications and management interventions in CFRs (Fiorella et al., 2019). In Hilsa fisheries in Bangladesh, management approaches including community fish guards were considered instrumental in the recovery of the fishery (Khan et al., 2020; Rahman et al., 2020). In further CFR research, a first step would be to incorporate community interviews into sampling and ask if fishing occurs in CFRs and surrounding areas during the dry season as part of sampling. This would help to answer if CFR species gains and losses are determined by the species present (and community trophic dynamics), or by (illegal) fishing events in CFR in the dry season, and also if these species gains and losses correspond to catch composition from nearby rice field habitats.
The effects of the local practice of stocking CFRs with predators (black fish) on community abundance and diversity (specifically white and grey fish) throughout the dry season are unknown. Additional sampling methods will be required to more reliably target black fish species and determine whether these high value species are being retained—and finding ample food during the dry season—and what effects this has on overall biodiversity in the CFR. This would also allow more study across the three fish guilds and an analysis on the influence of low-oxygen tolerance characteristics on response to seasonality alteration.
Data availability
The dataset analysed during the current study is available in the Zenodo Repository, https://zenodo.org/record/7041592.
References
Allen, D. J., K. G. Smith & W. R. T. Darwall (Compilers), 2012. The Status and Distribution of Freshwater Biodiversity in Indo-Burma. IUCN, Cambridge. ISBN 9782831714240.
Arrington, D. A. & K. O. Winemiller, 2003. Diel changeover in sandbank fish assemblages in a Neotropical floodplain river. Journal of Fish Biology. https://doi.org/10.1046/j.1095-8649.2003.00167.x.
Baran, E., N. So, P. Degen, X. Y. Chen & P. Starr, 2013. Updated information on fish and fisheries in the Mekong Basin. Catch Culture 19: 24–25.
Bauer, S. & I. Scholz, 2015. Adaptation to Climate Change in Southern Africa: New Boundaries for Development. Routledge [available on internet at: https://play.google.com/store/books/details?id=ESSoBgAAQBAJ].
Brooks, A., M. Kim, C. Sieu, V. Sean & V. Try, 2015. A Characterization of Community Fish Refuge Typologies in Rice Field Fisheries Ecosystems. WorldFish [available on internet at: http://aquaticcommons.org/18464/1/2015-37.pdf].
Bürkner, P.-C., 2017. BRMS: an R package for Bayesian multilevel models using Stan. Journal of Statistical Software 80: 1–28 [available on internet at: https://www.jstatsoft.org/v080/i01].
Campbell, I. C., C. Poole, W. Giesen & J. Valbo-Jorgensen, 2006. Species diversity and ecology of Tonle Sap Great Lake, Cambodia. Aquatic Sciences 68: 355–373.
Chheng, P., S. Un, J. Tress, V. Simpson & C. Sieu, 2016. Fish productivity by aquatic habitat and estimated fish production in Cambodia [available on internet at: https://www.worldfishcenter.org/resource_centre/WF-4078.pdf].
Clemens, W. A., R. E. Foerster & A. L. Pritchard, 1939. The Migration of Pacific Salmon in British Columbia Waters. The Migration and Conservation of Salmon. American Association for the Advancement of Science, Lancaster: 51–59.
Crook, D. A., D. J. Buckle, J. R. Morrongiello, Q. A. Allsop, W. Baldwin, T. M. Saunders & M. M. Douglas, 2020. Tracking the resource pulse: movement responses of fish to dynamic floodplain habitat in a tropical river. The Journal of Animal Ecology 89: 795–807. https://doi.org/10.1111/1365-2656.13146.
Dala-Corte, R. B., L. F. Sgarbi, F. G. Becker & A. S. Melo, 2019. Beta diversity of stream fish communities along anthropogenic environmental gradients at multiple spatial scales. Environmental Monitoring and Assessment 191: 288. https://doi.org/10.1007/s10661-019-7448-6.
de Wit, M. & J. Stankiewicz, 2006. Changes in surface water supply across Africa with predicted climate change. Science 311: 1917–1921. https://doi.org/10.1126/science.1119929.
De Xun Chua, S., X. X. Lu, C. Oeurng & T. Sok, 2022. Drastic decline of flood pulse in the Cambodian floodplains (Mekong River and Tonle Sap system). Hydrology and Earth System Sciences. hess.copernicus.org [available on internet at: https://hess.copernicus.org/articles/26/609/2022/].
Dong, R., Y. Wang, C. Lu, G. Lei & L. Wen, 2021. The seasonality of macroinvertebrate β diversity along the gradient of hydrological connectivity in a dynamic river–floodplain system. Ecological Indicators 121: 107112.
Dugan, P. J., C. Barlow, A. A. Agostinho, E. Baran, G. F. Cada, D. Chen, I. G. Cowx, J. W. Ferguson, T. Jutagate, M. Mallen-Cooper, G. Marmulla, J. Nestler, M. Petrere, R. L. Welcomme & K. O. Winemiller, 2010. Fish migration, dams, and loss of ecosystem services in the Mekong Basin. Ambio 39: 344–348. https://doi.org/10.1007/s13280-010-0036-1.
Fiorella, K. J., E. R. Bageant, M. Kim, V. Sean, V. Try, H. J. MacDonell, E. Baran, Y. Kura, A. C. Brooks, C. B. Barrett & S. H. Thilsted, 2019. Analyzing drivers of fish biomass and biodiversity within community fish refuges in Cambodia. Ecology and Society [available on internet at: https://www.jstor.org/stable/26796983].
Food and Agriculture Organization of the United Nations, 2019. The State of the World’s Biodiversity for Food and Agriculture: FAO Commission on Genetic Resources for Food and Agriculture Assessments. FAO [available on internet at: https://play.google.com/store/books/details?id=NamMDwAAQBAJ].
Freed, S., Y. Kura, V. Sean, S. Mith, P. Cohen, M. Kim, S. Thay & S. Chhy, 2020. Rice field fisheries: wild aquatic species diversity, food provision services and contribution to inland fisheries. Fisheries Research 229: 105615 [available on internet at: https://www.sciencedirect.com/science/article/pii/S0165783620301326].
Funge-Smith, S., 2018. Review of the State of the World Fishery Resources: Inland Fisheries. FAO [available on internet at: http://www.fao.org/3/CA0388EN/ca0388en.pdf].
Garaway, C. J., C. Photitay, K. Roger, L. Khamsivilay & M. Halwart, 2013. Biodiversity and nutrition in rice-based ecosystems; the case of Lao PDR. Human Ecology. https://doi.org/10.1007/s10745-013-9602-z.
Goodrich, H. R., J. R. Watson, R. L. Cramp, M. A. Gordos & C. E. Franklin, 2018. Making culverts great again. Efficacy of a common culvert remediation strategy across sympatric fish species. Ecological Engineering 116: 143–153 [available on internet at: https://www.sciencedirect.com/science/article/pii/S0925857418300831].
Gregory, R., 1997. Ricefield Fisheries Handbook, Cambodia-IRRI-Australia Project. IRRI, Cambodia [available on internet at: https://agris.fao.org/agris-search/search.do?recordID=XF2015042593].
Hoerl, R. W. & R. D. Snee, 2015. Statistical thinking: Improving business performance. Wiley. ISBN 978-1-19-20272-1.
Hortle, K. G., 2007. Consumption and the yield of fish and other aquatic animals from the Lower Mekong Basin. MRC Technical Paper 16: 1–88.
Hortle, K. G., S. Lieng & J. Valbo-Jorgensen, 2004. An Introduction to Cambodia’s Inland Fisheries. Mekong River Commission [available on internet at: http://www.mrcmekong.org/assets/Publications/report-management-develop/Mek-Dev-No4-Mekong-Fisheries-Cambodia-Eng.pdf].
Hortle, K. G., R. Troeung & S. Lieng, 2008. Yield and Value of the Wild Fishery of Rice Fields in Battambang Province, Near the Tonle Sap Lake, Cambodia. Mekong River Commission.
Ieno, E. N., A. F. Zuur, R. Bastida, J. P. Martin, M. Trassens & G. M. Smith, 2007. Multivariate analyses of South-American zoobenthic species – spoilt for choice. In Analysing Ecological Data. Springer, New York: 503-513.
Jiang, X., P. Zheng, L. Cao & B. Pan, 2021. Effects of long-term floodplain disconnection on multiple facets of lake fish biodiversity: decline of alpha diversity leads to a regional differentiation through time. The Science of the Total Environment 763: 144177.
Joffre, O., M. Kosal, Y. Kura, P. Sereywath & N. Thuok, 2012. Community Fish Refuges in Cambodia: Lessons Learned. WorldFish [available on internet at: http://pubs.iclarm.net/resource_centre/WF_3147.pdf].
Junk, W., P. B. Bayley & R. E. Sparks, 1989. The flood pulse concept in river–floodplain systems. In International Large River Symposium, 1989.
Khan, M. A., M. A. Wahab, A. B. M. M. Haque, M. Nahiduzzaman & M. J. Phillips, 2020. Value chain impact of the increased hilsa shad (Tenualosa ilisha) harvest in Bangladesh. International Food and Agribusiness Management Review 23: 355–368. https://doi.org/10.22434/IFAMR2019.0201.
Kong, H., M. Chevalier, P. Laffaille & S. Lek, 2017. Spatio-temporal variation of fish taxonomic composition in a South-East Asian flood–pulse system. PLoS ONE 12: e0174582. https://doi.org/10.1371/journal.pone.0174582.
Kummu, M. & J. Sarkkula, 2008. Impact of the Mekong River flow alteration on the Tonle Sap flood pulse. Ambio 37: 185–192. https://doi.org/10.1579/0044-7447(2008)37[185:iotmrf]2.0.co;2.
Landeiro, V. L., W. E. Magnusson, A. S. Melo, H. M. V. Espírito-Santo & L. M. Bini, 2011. Spatial eigenfunction analyses in stream networks: do watercourse and overland distances produce different results? Freshwater Biology 56: 1184–1192. https://doi.org/10.1111/j.1365-2427.2010.02563.x.
Langer, T. A., B. A. Murry, K. L. Pangle & D. G. Uzarski, 2016. Species turnover drives β-diversity patterns across multiple spatial and temporal scales in Great Lake Coastal Wetland Communities. Hydrobiologia 777: 55–66.
Legendre, P., 2019. A temporal beta-diversity index to identify sites that have changed in exceptional ways in space-time surveys. Ecology and Evolution 9: 3500–3514. https://doi.org/10.1002/ece3.4984.
Legendre, P. & O. Gauthier, 2014. Statistical methods for temporal and space–time analysis of community composition data. Proceedings of the Royal Society B: Biological Sciences 281: 20132728. https://doi.org/10.1098/rspb.2013.2728.
Liu, X. & H. Wang, 2010. Estimation of minimum area requirement of river-connected lakes for fish diversity conservation in the Yangtze River floodplain. Diversity and Distributions 16: 932–940.
Moran, P. A., 1950. A test for the serial independence of residuals. Biometrika 37(1/2): 178–181.
Ngor, P. B., P. Legendre, T. Oberdorff & S. Lek, 2018a. Flow alterations by dams shaped fish assemblage dynamics in the complex Mekong-3S River system. Ecological Indicators 88: 103–114 [available on internet at: https://www.sciencedirect.com/science/article/pii/S1470160X18300232].
Ngor, P. B., K. S. McCann, G. Grenouillet, N. So, B. C. McMeans, E. Fraser & S. Lek, 2018b. Evidence of indiscriminate fishing effects in one of the world’s largest inland fisheries. Scientific Reports 8: 8947. https://doi.org/10.1038/s41598-018-27340-1.
Penha, J., V. L. Landeiro, J. C. G. Ortega & L. Mateus, 2017. Interchange between flooding and drying, and spatial connectivity control the fish metacommunity structure in lakes of the Pantanal wetland. Hydrobiologia 797: 115–126.
Petsch, D. K., K. Cottenie, A. A. Padial, J. D. Dias, C. C. Bonecker, S. M. Thomaz & A. S. Melo, 2021. Floods homogenize aquatic communities across time but not across space in a Neotropical floodplain. Aquatic Sciences. https://doi.org/10.1007/s00027-020-00774-4.
Rahman, M. J., M. A. Wahab, M. Nahiduzzaman, A. B. M. Haque & P. Cohen, 2020. Hilsa fishery management in Bangladesh. IOP Conference Series: Earth and Environmental Science 414: 012018. https://doi.org/10.1088/1755-1315/414/1/012018/meta.
Sabo, J. L., A. Ruhi, G. W. Holtgrieve, V. Elliott, M. E. Arias, P. B. Ngor, T. A. Räsänen & S. Nam, 2017. Designing river flows to improve food security futures in the Lower Mekong Basin. Science. https://doi.org/10.1126/science.aao1053.
Shams, N., 2007. Contribution of rice field ecosytems to food security strategies in northwest Cambodia. Journal of Sustainable Agriculture 29: 109–133. https://doi.org/10.1300/J064v29n04_09.
Thorncraft, G., L. Baumgartner & T. Marsden, 2006. Fish passage and fishways in the Mekong Basin: getting past the barriers. In Proceedings of the 7th Technical Symposium Mekong Fisheries, 15–17 November 2005. MRC Conference Series. archive.iwlearn.net: 237–251 [available on internet at: http://archive.iwlearn.net/mrcmekong.org/download/free_download/proceedings_7TechSym/paper19-Fish-passage-n-fishways.pdf].
Tilley, A., J. López-Angarita & J. R. Turner, 2013. Effects of scale and habitat distribution on the movement of the southern stingray Dasyatis americana on a Caribbean atoll. Marine Ecology Progress Series 482: 169–179 [available on internet at: https://www.int-res.com/abstracts/meps/v482/p169-179/].
Tweddle, D., I. G. Cowx, R. A. Peel & O. L. F. Weyl, 2015. Challenges in fisheries management in the Zambezi, one of the great rivers of Africa. Fisheries Management and Ecology 22: 99–111. https://doi.org/10.1111/fme.12107.
Vu, A. V., L. J. Baumgartner, K. E. Limburg, G. S. Doran, M. Mallen-Cooper, B. M. Gillanders, J. D. Thiem, J. A. Howitt, C. M. Kewish, J. Reinhardt & I. G. Cowx, 2022. Life history strategies of Mekong pangasiid catfishes revealed by otolith microchemistry. Fisheries Research 249: 106239 [available on internet at: https://www.sciencedirect.com/science/article/pii/S0165783622000169].
Welcomme, R. L., 1995. Relationships between fisheries and the integrity of river systems. Regulated Rivers: Research and Management. https://doi.org/10.1002/rrr.3450110110.
Welcomme, R. L., 2001. Inland Fisheries: Ecology and Management. Wiley, Oxford [available on internet at: https://play.google.com/store/books/details?id=Y3TxzAEACAAJ].
Zuur, A. F., E. N. Ieno, J. N. Walker, A. A. Saveliev & G. M. Smith, 2009. In Gail, M., K. Krickeberg, J. M. Samet, A. Tsiatis & W. Wong (eds), Mixed Effects Models and Extensions in Ecology with R. Springer. https://doi.org/10.1007/978-0-387-87458-6.
Funding
This publication is made possible with kind support from GIZ and the Sustainable Aquaculture and Community Fish Refuge Project (SAFR) in Cambodia. This work was undertaken as part of the CGIAR Research Programme on Fish Agri-Food Systems (FISH) and the One CGIAR Initiatives “Securing the Food Systems of Asian Mega-Deltas for Climate and Livelihood Resilience” (AMD) and “Aquatic Foods” supported by contributors to the CGIAR Trust Fund. This work was funded by the United States Agency for International Development (USAID) Grant Number AID-442-IO-12-00001 and Feed the Future through USAID under the terms of Contract No. AID-442-IO-16-00002. The donor had no involvement in study design, execution, or report preparation. The opinions expressed herein are those of the authors and do not necessarily reflect the views of the U.S. Agency for International Development.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
All authors certify that they have no affiliations with or involvement in any organization or entity with any financial interest or non-financial interest in the subject matter or materials discussed in this manuscript.
Additional information
Handling editor: Luis Mauricio Bini
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Tilley, A., Freed, S., Cagua, E.F. et al. Retention of fish biodiversity in a mixed-use agroecosystem in Cambodia. Hydrobiologia 851, 1475–1488 (2024). https://doi.org/10.1007/s10750-023-05400-6
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-023-05400-6