
Abstract
Agave is one of the most important genera of plants in Mexico in terms of biodiversity and cultural and economic impact; however, its reproductive traits and breeding systems are poorly known. In addition, clonal propagation of agave cultures and the interruption of sexual reproduction may lead to differences not only in reproductive traits but also in seed set and seed viability across species and between wild plants and Agave landraces. Hence this research evaluated the morphological variation of flowers, fruits, and seeds throughout the infructescence development of pulquero agave landraces (Agave americana, A. mapisaga, and A. salmiana) and their wild relatives (A. salmiana) using statistical analyses. Additionally, seed set and seed viability (tetrazolium test) were estimated for traditional (metepantle) and intensive (monoculture) cultivation systems. The analysis of flowers allowed differentiating between species (A. mapisaga and A. salmiana), while the analysis of fruits separated wild from cultivated plants. Wild agaves attained seed set values close to 0.5, and cultivated agaves, less than 0.2; the lowest seed set values (0.04) were observed under monoculture. Seed viability percentages depended on the taxon analyzed, with no clear differences between wild and cultivated taxa. There were positive correlations between branch length, fruit number, black (potentially viable) seed number, and the percentage of viable seeds. The highest percentage of viable seeds was recorded in the low and middle parts of the infructescence. Our results demonstrate that reproductive traits are suitable for recognizing morphological groups across species and between wild and landrace agaves. They also show that intensive and prolonged clonal propagation is associated with low seed set and seed viability values in Agave landraces.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The genus Agave is endemic to the Americas, with Mexico as its species diversity center. This country is home to 79% of the species, 84% of which are endemic (Eguiarte et al. 2021; Gentry 1982). Agave plants have up to 92 different reported uses (Colunga-GarcíaMarín et al. 2017; Trejo et al. 2022), including the production of fiber and distilled and fermented beverages. One of the most ancient uses of this emblematic plant, which has recently raised interest, is the production of pulque, an alcoholic beverage produced from the fermentation of aguamiel—a sweet sap that flows out from the agave stem when it is scraped with a sharp instrument (Valdivieso-Solis et al. 2021).
Although pulque has been produced since pre-Hispanic times across a large part of the Mexican territory, the central region (Hidalgo, Tlaxcala, Puebla, Mexico City, Edo. Mexico, Querétaro) is recognized as the core of the pulque industry (Gentry 1982). In this region, we can find large agave plants with the following domestication syndrome: gigantism, which in Agave is commonly associated with polyploidy, flexible leaves, sweet aguamiel (9.57–9.8°Brix), reduced apical and lateral spines, and a low saponin content (Álvarez-Ríos et al. 2020; Figueredo-Urbina et al. 2017; Gentry 1982; Mora-López et al. 2011).
Research on the morphological diversity of agaves revealed clear differences between species (Figueredo-Urbina et al. 2017 (A. inaequidens, A. cupreata and A. hookeri); Rivera-Lugo et al. 2018 (Agave angustifolia complex); Trejo et al. 2020 (A. salmiana and A. mapisaga)), between wild and cultivated plants (Colunga-GarcíaMarín et al. 1996 (A. fourcroydes and A. angustifolia); Figueredo-Urbina et al. 2014 (A. inaequidens and A. hookeri), and between management levels (Figueredo-Urbina et al. 2021a (A. salmiana and A. mapisaga). However, the differences between landraces are slight (Álvarez-Ríos et al. 2020 (A. americana, A. salmiana and A. mapisaga); Figueredo-Urbina et al. 2021a; Trejo et al. 2020). Most research has focused on vegetative traits of the stem, and only a few have addressed reproductive traits. In addition, research on the latter usually involved small sample sizes, consisting of 5 to 10 flowers, fruits, or seeds from 1 to 10 plants per population or taxon (Colunga-GarcíaMarín et al. 1996; Rivera-Lugo et al. 2018; Vázquez-Pérez 2015 (Agave karwinskii complex)). This is possibly because reproductive traits are not the target of artificial selection and, therefore, do not provide information that allows recognizing the morphological divergence associated with the domestication process (Colunga-GarcíaMarín et al. 1996; Vargas-Ponce et al. 2007 (Agave angustifolia complex, A. rhodacantha and A. tequilana)). However, Colunga-GarcíaMarín et al. (1996), by including reproductive traits, better described the divergence between wild and cultivated plants and probable ecotypes among wild plants. Rivera-Lugo et al. (2018) recognized morphological divergence between taxonomic species and varieties and cultivated plants within the Agave angustifolia complex based on reproductive traits. The above suggests that reproductive characters have been little explored and that these may display diversity associated with artificial selection.
There are several complexities in measuring reproductive traits in agaves used for pulque production and their wild relatives. In this regard, it should be noted that in some wild and cultivated agave populations, floral scapes are cut and used as building materials or food. To extract aguamiel, the apical meristem of landraces is removed to prevent flowering and promote sugar accumulation in the stem. On the other hand, the flowers of some wild and cultivated plants are used to prepare various regional dishes, although the flowers of cultivated plants are preferred for this purpose because they are softer and less fibrous than those of wild plants (Figueredo-Urbina et al. 2021b; Trejo et al. 2022). A single plant not scraped to obtain aguamiel represents losses of 12 to 47 dollars (when sold whole) or hundreds to thousands of liters of aguamiel, worth 200 to over 500 US dollars. In addition, in recent decades, the exploitation of plants has surpassed their cultivation, resulting in a low availability of adult plants from which to obtain aguamiel (Trejo et al. 2022).
One issue observed in agave crops deals with a decreased seed set, that is, the ability to produce potentially viable seeds (black seeds), estimated as the ratio between the number of black seeds and the total number of seeds (black and white). This decrease, also considered a part of the domestication syndrome (Colunga-GarcíaMarín et al. 1996; Escobar-Guzmán et al. 2008; Romano-Grande 2022), implies that cultivated plants produce fewer black seeds than their wild counterparts, attributed to prolonged and intensive asexual propagation (Escobar-Guzmán et al. 2008; Huerta-Lovera et al. 2018). Cultivation in agave crops is commonly asexual, from basal and rhizomatous clonal stems that retain the characters selected by farmers; when these stems reach an approximate height of 70 cm to 80 cm, they are easily established in new sites (Figueredo-Urbina et al. 2021a; Gentry 1982; Trejo et al. 2020). An extreme case of intensive clonal propagation in monoculture, carried out for hundreds of years, is Agave tequilana Weber, also known as 'blue agave.' This species shows the lowest genetic diversity found in agaves (Gil-Vega et al. 2001; Trejo et al. 2018), in addition to abnormal embryonic development, structural alterations related to low pollen and seed viability, deformed embryos, empty seeds, and low embryo viability (Escobar-Guzmán et al. 2008; Ramírez-Tobías et al. 2016). The above indicates that agave reproduction based solely on cloning and intensive crop systems adversely affects genetic diversity and may increase inbreeding depression (Escobar-Guzmán et al. 2008; Gil-Vega et al. 2001; Ramírez-Tobías et al. 2016; Trejo et al. 2018).
Most Agave species display sexual and asexual reproduction, balancing their diversity and the permanence of populations. In wild agaves, fruit production relative to the number of flowers (fruit set) is approximately 0.45 (Figueredo-Urbina et al. 2017; Gonçalves de Lima 1986; Rocha et al. 2005), and only a small percentage of the resulting seedlings reach the adult stage (Arizaga et al. 2000; Molina-Freaner and Eguiarte 2003; Nobel 1977; Rocha et al. 2005). Arizaga and Ezcurra (2002) reported that only 1.5% of Agave macroacantha plants grown from seeds managed to survive seven years in their natural environment. In wild populations, most plantlets are likely to include different types of vegetative propagules: basal, rhizomatous, axillary, or aerial. However, a few cross-fertilization events may increase genetic diversity by producing genotypes that could offer new and faster adaptation while reducing the accumulation of deleterious genes or increased inbreeding depression that may threaten the survival of individuals or populations (Darwin 1876; Eguiarte et al. 2015). Genetic diversity from sexual reproduction may be a major reservoir of functional diversity allowing species to respond to extraordinary events related to climate change, such as droughts (Murray-Tortarolo 2021). If maintaining high genetic diversity in agave populations becomes a priority, reproduction through seeds must be incorporated into management practices. In this context, finding traits to identify the 'best seeds' is crucial.
Some research reports indicate that agaves used for pulque production are characterized by medium to high genetic diversity levels; however, these investigations have been carried out in crops with low plant production intensity (Álvarez-Ríos et al. 2020; Figueredo-Urbina et al. 2014, 2021a). It has not been explored whether seed set in intensive pulquero agave crops decreases, as reported in henequen (Colunga-GarcíaMarín et al. 1996). The pulquero agave cultivation age, dating back to pre-Hispanic times, may also influence seed set since some current plants may have descended through several generations of asexual propagation. An approach to solve this question would be to quantify the number of potentially fertile (black) seeds and determine their viability in crops of different ages and maintained under different management intensity levels. Seed viability could reflect the selection process and inbreeding levels in crops.Scanty research in this regard noticed lower viability associated with greater cultivation intensity, as in 'blue agave' (Agave tequilana) and A. americana (Escobar-Guzmán et al. 2008).
One of the difficulties in studying seed set, seed viability, and reproductive traits in general is seed collection. Few individuals are allowed to flourish in plantations, and field collection is also a challenge. A monocarpic agave plant produces seeds once in its lifetime, which can take 8 to 20 years for pulquero agave. An inflorescence can be more than 10 m in height, making it difficult to collect fruits. In addition, fruits and seeds do not ripen synchronously in a given population but over several weeks. In this scenario, knowing which characters of infructescence branches and fruits correlate with the number of black seeds and their viability along the infructescence would be important in optimizing seed collection for research and propagation purposes.
Aiming to contribute to a deeper understanding of the topics mentioned above, the present study had the following objectives: (i) to analyze reproductive traits (flowers, fruits, and seeds) for recognizing morphological groups in pulquero agave landraces and their wild counterparts; (ii) to compare the seed set between taxa and landraces grown in traditional and intensive production systems; (iii) to analyze the variability of reproductive traits (fruits and seeds), seed set, and seed viability along the infructescence; and (iv) to recognize the reproductive traits associated with black seeds and their viability and assess whether these characteristics change along the infructescence.
Materials and methods
Study site
A large part of the state of Tlaxcala has a temperate subhumid climate with summer rains. Phaozem, Regosol, and Cambisol are the predominant soil types. The plants included in this study were collected at different sampling sites to obtain the largest number of individuals for the study. Two sites corresponded to the urban area of the Tlaxcala capital and Tlaxco municipalities; four sites were located in the Atlangatepec, Chiautempan, Santa Cruz Tlaxcala, and Nanacamilpa municipalities, where land use is dedicated to agriculture, which is mainly mechanized; last, the site located in Carmen Tequexquitla has a temperate semidry climate and is covered by xeric shrubland (INEGI 2009). Nonsignificant differences have been reported for agave plants growing at the collection sites (Trejo et al. 2020) (Fig. 1; Table S1).

The state of Tlaxcala is located in the eastern center of Mexico at coordinates 19° 06′ 18″–19° 43′ 44″ N and 97° 37′ 31″–98° 42′ 30″ W. Municipalities where the sites of agave for pulque production were studied are shown
Plant material
Agave plants used for pulque production and wild plants of the same species, all with a developed floral scape, were collected from 2016 to 2021 and identified with taxonomic keys (García-Mendoza 2011; Gentry 1982). Previous research (Trejo et al. 2020, 2022;Muñóz-Camacho et al. 2023) found that landraces of A. salmiana, A. mapisaga, and A. americana are used for pulque production in Tlaxcala. In this state, the only species for which wild plants have been found is A. salmiana, which is rarely used for pulque production. Although plant owners allowed us to study their plants, not all the variables could be recorded from the same individuals for various reasons, including exploitation of the plant to extract sap, floral scape removal for consumption of the scape and flowers, or because some of the plants studied aborted all fruits. This situation posed methodological challenges that made it necessary to construct different datasets with the data obtained so that each could be analyzed separately to retain important biological information. Table 1 shows the number of plants included in each dataset. Even if the reproductive structures (flowers, fruits, and seeds) used in each analysis could belong to the same plants, the data were treated as separate datasets. The vegetative and reproductive traits obtained were analyzed as described below.
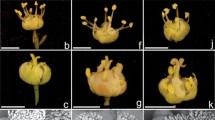
Flower dataset. The importance of flower characters for studying morphological diversity in pulquero agave was explored using 14 plants. Twenty-five flowers were randomly selected along each inflorescence and measured, for a total of 350 flowers; for each flower, 15 morphological characters were recorded (https://doi.org/10.5281/zenodo.8088373). The individuals belonged to five landraces of the species Agave salmiana Otto ex Salm-Dyck subsp. salmiana and Agave mapisaga Trel., var. mapisaga and three wild plants of Agave salmiana and A. salmiana subsp. tehuacanensis. Plants were collected in the seven localities of Tlaxcala mentioned above (Fig. 1, Table 1). For the variables filament length (FLm), anther length (ALm), filament diameter (FDm), and anther diameter (ADm) (Fig. 2, Table 2), the average of six stamens and six pistils of each flower was calculated to obtain a single data point per flower.

Scheme of the measurements of reproductive morphological traits of branches, flowers, fruits, and seeds. Flower: Total flower length (TFL), tepal length (TL), flower tube diameter (FTD), ovarian tube length (OTL), pedicel length (PL), anther diameter (ADm), anther length (ALm), filter diameter (FDm), filament length (FLm), pistil head length (PHL), pistil length (PIL), pistil diameter (PD), distance from the ovary to the pistil (DOP), distance from the ovary to the filament (DOF), oval length (O); Fruits: Capsule length (CL), apicule (A), capsule width (CW), carpel leaf width (CLW), stipe width (SW), stipe length (SL); Seeds: seed length (SEL), seed width (SEW); Branch: Total number of floral buds (TNB), total number of fruits (TNF), branch length (BL)
Fruit dataset 1. The importance of analyzing fruit characters to study morphological diversity in pulquero agave was assessed in 31 plants (Table 1). Nine ripe fruits per plant were randomly selected along the infructescence for a total of 279 fruits. Ten characters were measured for each fruit (Fig. 2, Table 2), with two corresponding to seed traits, namely, seed length and seed width (SEL and SEW, respectively; please refer to Table 2) (https://doi.org/10.5281/zenodo.8088373). These measurements were obtained for seven landraces of two species: Agave americana L. var. marginata, Agave salmiana subsp. salmiana, and a wild plant of Agave salmiana subsp. tehuacanensis. The number of plants in each landrace is shown in Table 1.
To analyze fruit characteristics, we assessed the number of black seeds, reproductive capacity, and seed viability along the infructescence. To assess the correlation between the characteristics of fruits and the presence of black seeds, we used two additional fruit datasets, Fruit dataset 2 and Fruit dataset 3, which recorded the fruit characters capsule length (CL), capsule width (CW), carpel leaf width (CLW), stipe length (SL), stipe width (SW), and number of black seeds (NBS). Differences in the sample size of these datasets allowed us to test whether we could obtain a similar trait pattern along the infructescence with samples of different sizes.
Fruit dataset 2. This set included three plants (1292, 871, 709; Table 1) of the Agave salmiana subsp. salmiana 'Manso' landrace from Nanacamilpa, Tlaxcala, from which three fruits were randomly collected for each infructescence branch, for a total of 237 fruits (https://doi.org/10.5281/zenodo.8088373).
Fruit dataset 3. This included three plants of Agave salmiana subsp. salmiana 'Manso' (720, 729, 731; Table 1). From these plants, we collected all fruits in the infructescence, for a total of 1986 fruits. In addition to the common variables measured in Fruit datasets 1 and 2, for these plants, we measured the characters infructescence height (IH), number of branches (NBR), branch length (BL), number of floral buds (TNB), number of fruits per branch (TNF), number and weight of black seeds (TNBS and BSW, respectively), and viability per branch (VpB; refer to Table 2). White seeds were also counted, although these were small and stuck to each other, so counting them was difficult; therefore, white seeds were counted only from the fruits of plant 720. Data from this database were used to analyze the correlation between reproductive traits and the number and viability of black seeds along the infructescence and the relationship with branch characteristics (Fig. 2, Table 2; https://doi.org/10.5281/zenodo.8088373).
Seed dataset. We assessed the importance of analyzing seed traits for studying morphological diversity in pulquero agave. To this end, we measured 200 seeds randomly collected from each plant. Four traits were measured for each seed: length (SEL2), width (SEW2), weight (SWG), and thickness (ST, see Tables 1 and 2) (https://doi.org/10.5281/zenodo.8088373). The plants analyzed belonged to three landraces of Agave salmiana subsp. salmiana: 'Ayoteco' (one plant), 'Manso' (two plants), and 'Prieto' (one plant), in addition to Agave salmiana Amarillo (one plant) and A. salmiana subsp. tehuacanensis De Cerro (one plant) (Table 1).
Importance of reproductive characters (flowers, fruits, and seeds) in defining morphological groups
The correlations between flower, fruit, and seed traits were explored through Spearman's correlation analyses. Subsequently, a principal component analysis (PCA) was performed for the three databases to reduce the dimensions. Data were centered to perform the analysis. To simplify the expression of the most important variables for each component and facilitate interpretation, we performed varimax rotation after the PCA with the psych package in R (Revelle 2021). After PCA, hierarchical clustering on principal components (HCPC) was performed (Husson et al. 2015) using Euclidean distances and Ward's criterion (Ward 1963). HCPC combines PCA and cluster analysis; HCPC clusters are produced from the principal components determined by PCA. We used the elbow and average silhouette width methods to select the number of clusters. The resulting clusters grouped the flowers, fruits, and seeds of the plants analyzed and allowed visualization of potential groups. The PCA and HCPC were performed with the packages factoextra (Kassambara and Mundt 2020), FactoMineR (Husson et al. 2015), and Factoshiny in R v. 4.2.1 (R Core Team 2022).
To reduce dimensionality and analyze the possibility of classifying flowers, fruits, and seeds according to taxon and crop system, we conducted a linear discriminant analysis (LDA) with the MASS package (Venables and Ripley 2002). We defined a training set with the data and then fitted a linear model. As the dependent variable, we used the ‘source’ of plants (landrace or wild) for flowers and species for fruits and seeds. These categories were suggested by the previous analysis. We examined the variation explained by each linear discriminant function and analyzed the coefficient obtained for different traits.
After identifying the variables determining the linear discriminant (LD) function in each analysis, these variables were compared between the proposed groups. We performed Kruskal‒Wallis tests to compare the traits identified as important in the LDA: three flower traits (flower tube diameter [FTD], total flower length [TFL], and ALm; please refer to Table 2) and two fruit traits (NBS and total number of seeds [TNS]). Differences between groups were explored with pairwise Wilcoxon rank sum tests. To compare the four traits measured for seeds between the groups tested in the LDA, we used a one-way ANOVA followed by a post hoc Tukey’s test when significant differences were observed.
The potential effect of crop management (intensive or traditional) on the expression of seed traits was assessed with Welch’s t tests for each trait in the 'Manso' landrace. Prior to the tests, the data were assessed for normality and homoscedasticity. Variables were square-root transformed as needed to achieve normality; if the normality assumption was not met, we used a nonparametric test.
Seed set
Seed set was calculated individually for the 35 plants from Fruit datasets 1 to 3. We calculated the mean and 95% confidence intervals (CIs) of seed set by species, landrace, and crop system (Fruit datasets 1–2). Significant differences between crop systems (intensive or traditional) within the salmiana subspecies (Fruit dataset 1) were assessed with a Mann‒Whitney U test.
Seed set was also estimated at the within-individual level along the infructescence (Fruit datasets 2–3); to this end, each infructescence was divided into five levels (a–e) in the acropetal direction. A Kruskal–Wallis test followed by post hoc Dunn–Bonferroni correction was performed to explore potential differences in seed set along the infructescence.
Seed viability
To analyze the viability per plant (VpP), 600 seeds were randomly collected from each plant in the seed dataset. In addition, VpB was determined for seeds produced by the three plants of A. salmiana subsp. salmiana 'Manso' in Fruit dataset 3 (plants 720, 729, and 731). For these plants, we tested an average of 19 black seeds randomly selected (minimum 5, maximum 20) from each branch.
Viability was determined using the tetrazolium test (ISTA 2010). Seeds were first disinfected; afterward, they were immersed in a commercial liquid detergent solution for 20 min, rinsed with distilled water, immersed in a 70% ethanol solution for one minute, immersed in a 3% sodium hypochlorite solution for 10 min, and finally rinsed with distilled water (Flores-García et al. 2008; Santacruz-Ruvalcava et al. 1999). Seeds were conditioned by placing them in glass jars and leaving them to imbibe in distilled water for 24 h. An incision parallel to the hypocotyl axis was made on each seed with a scalpel to facilitate the entry of the tetrazolium solution. Afterward, seeds were placed on filter paper in Petri dishes, and 15 mL of a 1% 2,3,5-triphenyl tetrazolium solution, pH 7, was added (Van-Waes and Debergh 1986). Seeds were incubated at 25 °C for 24 h in darkness. Embryo viability was determined based on the embryo staining pattern. Viable embryos were mostly (2/3) or entirely stained; otherwise, they were deemed nonviable. Viability was expressed as a percentage.
We evaluated the following. (a) The potential relationship between seed viability and agave landrace. To this end, a Xi2 test of independence was carried out. Pearson's residuals were calculated to test for significant differences between observed and expected values. (b) The relationships between SEL2, SEW2, ST, and SWG and VpP were tested with Pearson's correlation. The percentage of viable seeds per branch was compared between the three plants of Fruit dataset 3 (720, 729, and 731) using a Kruskal‒Wallis test followed by a Wilcoxon Rank Sum test.
Correlations between characters of branches, seeds, and fruits and differences along the infructescence
To identify predictors of TNBS and VpP within the infructescence, from the data recorded for the three plants of Fruit dataset 3, Spearman's correlations were run for BL, TNF, TNBS, and BSW with TNB and VpB.
We also examined whether BL, TNF, TNBS, TNB, and VpB changed along the inflorescence. To this end, branches were classified according to their position on the inflorescence, assigning them to one of five categories in the acropetal direction (A–E). For each trait, we performed Kruskal‒Wallis and pairwise Wilcoxon rank sum tests to investigate potential differences between categories along the infructescence.
Fruit characters as predictors of the presence of black seeds and differences along the infructescence
With Fruit datasets 1 to 3, which shared six variables (CL, CW, CLW, SL, SW, and NBS), we explored which fruit traits were correlated with NBS using Spearman's correlations between each variable and NBS. We also analyzed how the values of these characters changed along the infructescence (Fruit datasets 2–3). Fruit dataset 3 contains a census of fruits for the three plants, while Fruit dataset 2 contains data from a random sample of nine fruits per branch. The different datasets allowed us to test which fruit variables could be used to predict the presence of black seeds and whether these correlations were maintained in samples of different sizes. To test for changes along the infructescence, branches were classified in the same way described above, and the six variables were compared between levels with a Kruskal‒Wallis test and a Wilcoxon test, as needed. All statistical analyses were carried out in R v. 4.2.1 (R Core Team 2022).
Results
Importance of reproductive characters (flowers, fruits, and seeds) in defining morphological groups
The significant correlations between the variables of Flower dataset, Fruit dataset 1, and Seed dataset support the feasibility of performing PCA (Figure S1).
Flower dataset. In the case of floral traits, the first three PCA components explain approximately 65% of the total variation observed. The first component (RC1) accounts for 31% and is defined mainly by tepal length (TL), total flower length (TFL), and FLm (please refer to Table 2). The second component (RC2) explains 18% of the variability between groups and is defined mainly by the distance from the ovary to the pistil (DOP) and ovarian tube length (OTL). RC3 accounts for 15% of the variability and is defined by DAm and pedicel length (PL) (Table 3).
Figure 3a depicts the PCA built from the flower characters, showing two main clouds: one represents Agave salmiana and the other, Agave mapisaga, along with most individuals of the A. salmiana subsp. salmiana 'Ayoteco' landrace. The Agave salmiana cloud shows a continuous distribution with two ends. Plants 1191 and 1182, identified as wild A. salmiana, are located in the lower left quadrant, and plants 1176 and 1530 of the subspecies salmiana 'Manso' are positioned in the upper right quadrant. Another cloud groups the flowers of plants 1187 and 1188 of 'Ayoteco' and plant 1565 of A. mapisaga var. mapisaga, separated to the right of the main cloud (Fig. 3a); these three plants are cultivated. An HCPC was performed with the first three principal components (Fig. 3b). Three clusters are apparent: from left to right, Cluster A includes flowers of plants identified as wild and cultivars of the two subspecies of Agave salmiana; in this cluster, flowers have high values of PL, TFL, FTD, and TL. Cluster B mainly groups flowers of the 'Manso' salmiana subspecies; the flowers in this cluster have high values of OTL, DOP, and ovary length (O) and low values of pistil head length (PHL) and ADm. Cluster C contains flowers of plants 1187, 1188, and 1565 (A. mapisaga and 'Ayoteco'); the flowers in this cluster have high values of ALm, TL, and FTD, and low values of PL and DOP (Fig. 3b).

Flower dataset. a Principal component analysis (PCA) built from flower traits. The larger cloud includes mainly Agave salmiana plants. Flowers mainly of wild plants of the two A. salmiana subspecies are positioned in the bottom left. Flowers mainly of cultivated agaves (landraces) are positioned in the top right. The cloud to the right includes flowers of Agave. mapisaga var. mapisaga + 'Ayoteco'. b Dendrogram generated by hierarchical clustering on principal components (HCPC) from three principal components (PCs) based on floral traits. The dendrogram produced three clusters. A and B contain most A. salmiana flowers. C includes flowers of A. mapisaga var. mapisaga + 'Ayoteco'. Tags correspond to the ID assigned to each flower for analysis. Colors do not correspond between panels
LDA was carried out with the data from the flower dataset. The categories tested with this analysis were landraces of A. salmiana (plants 680, 1008, 1010, 1176, 1180, 1186, 1192, 1530, and 1963), wild A. salmiana (plants 1191 and 1182), and A. mapisaga + 'Ayoteco' (plants 1187, 1188, and 1565) (Figure S2a). The analysis revealed that the first two discriminant functions explained 97% of the variation (LD1: 75.4%; LD2: 24.5%). The first linear discriminant function was strongly related to positive values of ALm and FTD, while LD2 was positively related to ALm and negatively to TFL. From the previous analyses and the Kruskal‒Wallis (K-W) tests, the characteristics of flowers of the groups tested in the discriminant analysis can be summarized as follows. The flowers of wild A. salmiana plants (1191 and 1182) are shorter and thinner than those of the rest; the flowers of cultivated A. salmiana plants are long and thin; and those of 'Ayoteco' and A. mapisaga var. mapisaga (1187. 1188, and 1565) are the longest and widest (p < 0.05).
Fruit dataset 1. In the case of fruit traits, the PCA showed that the first three components explained 73% of the total variation (Table 4). The first component accounted for 38% and was defined mainly by CW and SEL. The second component explained 21% of the variability between the groups and was defined by the total number of seeds (TNS) and number of white seeds (NWS); the third component explained 13% of the variation and was defined by SL. Only the fruits produced by\plant 2020 of A. salmiana subsp. tehuacanensis from De Cerro tended to differ from those of the rest (Figure S2b).
An HCPC was performed with the first two PCA (Fig. 4a). Clusters A and C contained the fruits of A. salmiana subsp. salmiana and A. americana. The analysis grouped the fruits of A. salmiana subsp. tehuacanensis in a third cluster (B). The linear discriminant analysis of fruit characters, through which we sought to separate the taxa (A. salmiana subsp. salmiana, A. salmiana subsp. tehuacanensis and A. americana), shows that the first discriminant function accounts for 92% of the variation (Fig. 4b). This first LD factor is related to TNS and NBS. The K-W test shows that the fruits of A. salmiana subsp. tehuacanensis produce significantly more seeds (Xi2 = 21.55, p < 0.05) and black seeds (Xi2 = 20,796, p < 0.05) than the fruits of the A. salmiana subsp. salmiana and A. americana plants studied. In addition, the fruits of A. salmiana subsp. tehuacanensis have wider carpel leaves than those of A. salmiana subsp. salmiana and A. americana. The seeds of A. salmiana subsp. tehuacanensis are longer than those of the other two species (p < 0.05).

Fruit dataset 1. a Dendrogram derived from the hierarchical clustering on principal components (HCPC) showing three clusters. Clusters A and B contain most of the fruits of Agave salmiana subsp. salmiana and A. americana. Cluster C contains fruits of A. salmiana subsp. tehuacanensis. b Plot showing the first two linear discriminant functions of the LDA, which account for 92% of the variation. Tags correspond to the IDs assigned to fruits for analysis. Colors do not correspond between panels
Seed dataset. The PCA performed with seed traits showed that the first three components accounted for 91.1% of the variation (Table 5, Figure S2c). The results reveal two major axes of seed morphological variation: PC1 explains 45.3% of the variation and is best defined by SEL2, whereas PCA2 explains 33.8% of the variation and is mainly defined by ST. The HCPC carried out from the PCA is shown in Fig. 5a. Cluster A mainly includes seeds of the 'Prieto' and 'Ayoteco' landraces; this cluster is characterized by high ST values and low SEL2 and ST values. Cluster B mainly includes seeds of the cultivars 'Manso' and 'Amarillo', characterized by high SEL2 and SEW2 values and low ST and SWG values. Finally, Cluster C mainly includes seeds of wild A. salmiana subsp. tehuacanensis and is characterized by high values of all the variables. The linear discriminant analysis for seed traits of wild and landrace plants revealed that the two first linear discriminant functions accounted for 97.4% of the variation (LD1: 74.8%; LD2: 22.6%). The first linear discriminant function was strongly related to positive SWG values, while LD2 was strongly but negatively related to SEL2 (Fig. 5b). Therefore, we found that seeds of wild plants were clearly separated from seeds of landraces along the LD2 axis (i.e., greater length values).

Seed dataset. a Dendrogram plotted after the hierarchical clustering on principal components (HCPC). Cluster A mainly includes seeds of the landraces 'Prieto' and 'Ayoteco'; Cluster B mainly includes seeds of the landraces 'Manso' and 'Amarillo'. All these are landraces of the subspecies salmiana; Cluster C mainly includes seeds from wild Agave. salmiana and the subspecies tehuacanensis. b Plot showing the first two linear discriminant functions of the LDA, which account for 97.4% of the variation. Colors mark the sources of seeds (wild or landrace plant). Tags correspond to the IDs assigned to flowers for analysis. Colors do not correspond between panels
One-way ANOVAs revealed that the seeds of landraces have higher ST (F = 67.54, p < 0.001) but lower SEL2 and SEW2 values than the seeds of wild plants (F = 96.524, p < 0.001; F = 9.62, p < 0.001, respectively). Regarding the influence of management on the expression of seed traits, the t tests applied to the 'Manso' landrace revealed that seeds obtained from plants grown under intensive management were smaller (t = 5.34, p < 0.0001), thinner (t = -3.63, p < 0.001), and lighter (t = -3.19, p < 0.01) than those from plants grown under traditional management.
Seed set
The ornamental plant Agave americana var. marginata yielded a seed set of 0.15 ± 0.08 [95% CI 0.9–0.21] (Table S2), while Agave salmiana subsp. salmiana (which integrates all landraces) showed an average of 0.09 ± 0.09 [95% CI 0.03–0.27]. The seed set obtained for plants in Fruit datasets 2 and 3 was similar to that for plants in Fruit dataset 1 (Table S3). The wild plant of Agave salmiana subsp. tehuacanensis De Cerro had the highest value (seed set = 0.45 ± 0.21 [95% CI 0.27–0.61]) (Table S2).
The seed set of plants differs significantly according to the culture system (U = 24.50, p = 0.005) (Table S2). The seed set under the traditional culture system, metepantle (\(\overline{X}\) = 0.10 ± 0.07 [95% CI 0.07–0.13]), is higher than the value estimated for plants grown under an intensive cultivation system (\(\overline{X}\) = 0.04 ± 0.01 [95% CI 0.03–0.05]). The same pattern is observed when analyzing 'Manso' plants in Fruit datasets 2 and 3 (K-W = 49.34, p < 0.05).
The distribution of seed set along the infructescence was analyzed for all 'Manso' plants in Fruit dataset 2. A decreasing trend in the acropetal direction is observed; i.e., basal branches tend to have a high seed set (\(\overline{X}\) = 0.08 ± 0.07 [95% CI 0.07–0.10]), while more distal branches show a lower seed set (\(\overline{X}\) = 0.05 ± 0.03 [CI [0.04–0.06], (K-W = 11.01, p < 0.001). Fruit dataset 3, for which we recorded the total seed count of a single plant, shows a similar trend, with a higher seed set on the first basal branches (\(\overline{X}\) = 0.05 ± 0.03 [95% CI 0.04–0.05]) and a significant difference between the highest branches (K-W = 42.32, p < 0.001).
Seed viability
In the Seed dataset, the viability per plant (VpP) was 67.6% for A. salmiana subsp. salmiana and 94.5% for A. salmiana subsp. tehuacanensis. Plants of A. salmiana subsp. salmiana under traditional management (metepantle) showed high viability percentages: 'Prieto' (88%) and 'Ayoteco' (79.3%). 333 For the 'Manso' landrace, very similar values were obtained for plants grown under metepantle and intensive management: 57.8% and 57.7%, respectively. Wild A. salmiana Amarillo showed the lowest VpP, at 55.8%, while wild A. salmiana tehuacanensis De Cerro showed 94.5%—the highest of all taxa (Table S4). The difference in VpP depends on each taxon analyzed (X2 = 443.35, df = 5, p < 0.001).
Seed nonviability values depended on the taxon (X2 = 356.26, df = 15, p < 0.001). The local ‘Manso’ landrace showed the highest proportion of nonviable seeds in both management systems. Most corresponded to ‘empty’ seeds, representing more than 45%; this contrasts with the other landraces, for which the percentage of ‘empty’ seeds barely exceeded 15%. The A. salmiana tehuacanensis De Cerro wild plant showed the lowest percentage of nonviable seeds (5.5%) (Table S4).
The correlation analysis between seed and plant viability revealed that only seed weight was correlated with the percentage of viable seeds (r = 0.909, p < 0.01; Table 6).
Viability per branch (VpB) was analyzed for the three plants in Fruit dataset 3 ('Manso'), which revealed an average of 450 black seeds per plant. VpB was below 50% in all plants (plant 720, 42.3%; plant 729, 40.76%; plant 731, 30.18%), lower than the values mentioned above. No significant differences in VpB among plants were found (H = 0.219, p = 0.896). In the three plants studied, approximately 60% of nonviable seeds were empty, 26% had endosperm but no embryo, 10% had an embryo but no endosperm, and 4% had an embryo and endosperm. We observed significant differences in seed viability along the infructescence; these are described in the following section.
Correlation between the characters of branches, seeds, and fruits and differences along the infructescence
Branch length showed a significant and positive correlation with the number of fruits per branch (TNF), number of black seeds (TNBS), number of floral buds (TNB), and VpB. 349 In addition, TNF showed a significant and positive correlation with TNBS (p < 0.05; Table S5).
When exploring how these characters change along the infructescence, we found significant differences in branch length (BL) between the different levels of the infructescence (Xi2 = 58.851, p < 0.001); the lower and middle branches were the longest (levels A–C; Fig. 6).

Fruit dataset. a From left to right, plant 731 (11.35 m), plant 720 (11.24 m), and plant 729 (11.06 m). b Length of branches along the inflorescence. Letters represent homogeneous groups (post hoc test, p < 0.001)
The highest number of fruits was found in infructescence levels A to D (Xi2 = 28.353, p < 0.01). TNBS showed no significant differences in the first four levels (Xi2 = 20.324, p < 0.1) and was higher in these than in the top level. VpB and TNB were higher for levels A, B, and C than for the upper infructescence levels (D and E). Seeds from fruits at level E had significantly lower VpB values than those from the bottom three levels, with no significant differences between them (p < 0.05; Fig. 7). The anatomical characteristics of plants in Fruit dataset 3 are summarized in Table S6.

Number of fruits per branch, number of black seeds per branch, percent viability per branch, and number of shoots per branch along the inflorescence for the three plants in Fruit dataset 3. Letters represent homogeneous groups (post hoc test, p < 0.05)
Fruit characters as predictors of the presence of black seeds and differences along the infructescence
We explored how the six common variables in Fruit datasets 1 to 3 (CL, CW, CLW, SL, SW, and NBS) correlated with each other and how they changed along the infructescence (Fruit datasets 2–3). NBS was correlated (p < 0.05) mainly with CW and CLW. These results were obtained from data for plants in Fruit datasets 1 to 3 (Fig. 8, Tables S7-9) and 'Manso' plants in Fruit dataset 1 (Table S10) and by analyzing the three plants in Fruit dataset 3 separately (data not shown).

Correlations between the number of black seeds and fruit traits in the plants of Fruit dataset 1. The circle size and color intensity reflect the correlation strength. Blue means a positive correlation; red, a negative correlation. Capsule length (CL), capsule width (CW), carpel leaf width (CLW), stipe length (SL), stipe width (SW), number of black seeds (NBS)
When we explored the differences in fruit variables and the number of seeds along the infructescence in Fruit datasets 2 and 3, the variables CW, CLW, and NBS showed no significant differences along the infructescence (K-W test, p < 0.05); the pattern was similar for CL, SL, SW, and TNS in the two fruit datasets (Fig. 9, Figure S3).

Boxplots representing capsule width (CW), capsule leaf width (CLW), and number of black seeds (NBS) values for the five levels along the infructescence. Red dots represent mean values. Letters represent homogeneous groups, p < 0.05. Images a, c, and e correspond to Fruit dataset 2; images b, d, and f, to Fruit dataset 3
Discussion
Importance of reproductive characters (flowers, fruits, and seeds) in defining morphological groups
Flower dataset. The flower characters with the highest loadings for the first and second principal components in the PCA of flowers were related to flower size. A differentiation trend was observed from the small flowers of wild plants to the large flowers of landraces. However, differences occurred mainly between species. There may be an allometric relationship between flower size and rosette size, suggesting that artificial selection has been a driver of the morphological diversity of flowers. Unfortunately, the relationship between flower size and stem size in agaves has not been sufficiently analyzed to infer any allometric relationship between them. This topic deserves to be investigated in future research.
In some previous research analyzing vegetative and reproductive characters, floral traits were not the most relevant, although some appeared to be significant for the first to the third principal component or discriminant function. ALm and TL appear to be significant in all works, including the present study (Colunga-GarcíaMarín et al. 1996; Figueredo-Urbina et al. 2017; Rivera-Lugo et al. 2018; Vázquez-Pérez 2015).
The inclusion of floral characters in the morphological diversity analyses of agaves has allowed differentiation between species, botanical landraces, and ecotypes (Colunga-GarcíaMarín et al. 1996; Figueredo-Urbina et al. 2017; Rivera-Lugo et al. 2018; Vázquez-Pérez 2015). However, as observed in this work, the analysis of just flowers only allowed differentiating between species; the exception was the salmiana subspecies 'Ayoteco' landrace, which was grouped with A. mapisaga. This was also observed in the analysis of vegetative characters and chloroplast and nucleus intergenic sequences in relation to pulque production in Tlaxcala (Trejo et al. 2020). Based on the above, A. salmiana and A. mapisaga are recognized as different species in terms of morphological and genetic features, and the hypothesis that 'Ayoteco' is a hybrid between the two species is supported.
Fruit dataset 1. The analysis of fruit and seed characters revealed differences mainly between wild and cultivated plants. The wild subspecies tehuacanensis has wider capsules with greater TNS and NBS values. These characters have been relevant for identifying differences between species and landraces (Colunga-GarcíaMarín et al. 1996; Figueredo-Urbina et al. 2017; Rivera-Lugo et al. 2018). The fruits of wild plants are the widest, being related to the greatest number of black seeds. In turn, NBS is positively correlated with larger seeds (length and weight). However, since we analyzed only a single wild plant and one individual of A. americana, the above finding should be interpreted with caution; this exercise should be replicated including different species and taxonomic levels.
Seed dataset. When comparing seeds produced by 'Manso' plants under metepantle and monoculture, we observed that agave plants grown under intensive cultivation (monoculture) produced shorter, narrower, lighter, and thinner seeds (fewer black seeds) than plants grown under metepantle. It has been reported that seed weight and seed thickness are related to a greater amount of endosperm and a larger embryo, which may translate into greater seed viability and seedling survival and establishment (Garrido et al. 2005; Ortiz-Hernández et al. 2018; Wolf et al. 1986).
Other research has also stressed the significance of seed length, thickness, and weight in recognizing groups across taxa (Colunga-GarcíaMarín et al. 1996; Sánchez-Salas et al. 2017; Vázquez-Díaz et al. 2011). In cacti, Guillén et al. (2013) observed that selection has not translated into larger seeds. It is considered that larger seeds may have greater energy reserves, leading to higher germination rates, seedling vigor, and successful establishment (Guillén et al. 2011 2013; Loza-Cornejo and Terrazas 2011). The advantages of larger seeds are discussed below.
Seed Set
There are few reported data on the seed set of agaves. The seed set ranged between 0.1 and 0.6, with an average of 0.35. The lowest seed set values were observed in species grown under high cropping intensity, such as A. tequilana and A. americana (0.1). In agaves (Table S11), reported data show seed set values from 0.1 to 0.4 (\(\overline{X}\) = 0.2) for landraces and from 0.3 to 0.6 (\(\overline{X}\) = 0.4) for wild plants. This work recorded the same trend of higher seed set values in wild than in cultivated plants. In addition, plants grown under low crop intensity (metepantle) showed higher seed set values than plants grown in monoculture.
Except for the Agave salmiana subsp. tehuacanensis wild plant analyzed, the salmiana landraces measured had seed set values below the average reported above; these values may also be related to the subspecies and are not contrary to the trends described above. The lowest values correspond to plants grown under high cropping intensity (PULMEX, Nanacamilpa) and propagated from a small number of plants introduced from the state of Hidalgo (northwest of Tlaxcala, 50 km from Nanacamilpa) some 100 years ago (pers. comm. Senobio Becerra 2019). The above may affect the lowest seed set values (0.04) reported to date. Seed set values per plant were similar when measuring nine fruits per plant (Fruit dataset 1), nine fruits per branch (Fruit dataset 2), and all fruits from the plant (Fruit dataset 3; Table S3).
Seed set was estimated from a small number of fruits per plant (Table S11). The greatest sampling effort was previously reported by Huerta et al. (2018) when counting black and white seeds in 11% of fruits per umbel. Our work is the first in which all the black seeds in all fruits of three Agave plants and all the white seeds of a single individual have been counted. For future research, we recommend recording a larger number of measurements to estimate the adequate number of fruits that yield a close approximation of the seed set without having to count all seeds of all fruits, i.e., with a lower sampling effort.
There are several hypotheses on seed production in agaves. As most Agave species are monocarpic, a high reproductive cost is suggested with no guarantee that fruits are formed (Nobel 1977) since most seeds are either consumed (García-Meneses 2004; Nobel 1977) or do not develop into adult individuals (Jordan and Nobel 1979). However, the establishment of some recombinant individuals increases population genetic diversity and replaces the parent plant (Eguiarte et al. 2013).
The low seed set of agaves has been associated with limited resource allocation to seeds (Garrido et al. 2005; Sutherland 1987), low production of poor-quality pollen (Estrella 2005; Sutherland 1987), a lack of pollinators (Howell and Roth 1981; Molina-Freaner and Eguiarte 2003), the position of flowers (Garrido et al. 2005; Marquis 1988; Sutherland 1987), an advantage to male fitness (Sutherland 1987), and selective abortion (Sutherland 1987; Wiens 1984). In addition, the lower seed set of cultivated plants has been related to high cropping intensity, which may lead to inbreeding depression (Ramírez-Tobías et al. 2016). In monoculture, we observed plants with deformed flowers and malformed branches, which aborted all fruits (Fig. 6a). Future research will measure genetic diversity and inbreeding in these plants.
Seed viability
Seed viability is associated with the taxonomic identity of individuals. The subspecies tehuacanensis (wild plants) showed higher viability values than the subspecies salmiana (landraces). Within salmiana, differences were observed between landraces under traditional management and less intensive use ('Prieto' and 'Ayoteco', respectively), which showed the highest viability (VpP and VpB, see Table 2). In the case of 'Manso', the most used landrace in Tlaxcala and central Mexico (Reyes-Agüero et al. 2019; Trejo et al. 2022),past intensive use led to the landrace showing the lowest viability and highest nonviability regardless of the current crop modality. However, the low viability of wild A. salmiana Amarillo suggests that this is a wild trait, although it would not explain the high viability recorded in 'Prieto' and 'Ayoteco'.
Some research explored the viability of agave seeds using tetrazolium salts, showing a pattern in which higher cropping intensity is associated with lower viability. Agave tequilana var. Azul showed 19% viability and a nonviability of 81%; 28% of the seeds did not contain an embryo (Ramírez-Tobías et al. 2016). In addition, it has been reported that pollen grains of 'blue agave' develop abnormal pollen tubes, which may lead to weak embryos with high short-term mortality and subsequent abortion of the seed (Escobar-Guzmán et al. 2008).
In the case of A. mapisaga, a species used for pulque production in central Mexico, one study reported 68% viability, 31% nonviability, and 1% of seeds with no embryo (Ramírez-Tobías et al. 2016). This species shows lower production intensity and use than A. salmiana subsp. salmiana 'Manso'. For 'Manso' grown in Nanacamilpa, Tlaxcala, plants grown under metepantle showed 68.8% viability (13.6% seeds with no embryo), whereas plants under monoculture had 44.8% viability (11.6% seeds with no embryo); in the latter system, 26% of seeds were empty (Romano-Grande 2022). Prolonged and intensive clonal propagation may increase inbreeding depression, which reduces the genetic diversity of populations and influences seed viability. This effect may be greater in crops formed from a handful of plants without introducing plants from other sources.
In the present study, seed viability was positively correlated with seed weight, as observed in other agaves and perennial plants (Garrido et al. 2005; Ortíz-Hernández et al. 2018; Vogler and Bahnisch 2006; Wolf et al. 1986). Seed weight is related to the amount of available reserves and other functional characteristics that positively affect germination and seedling establishment. At the intraspecific level, a higher seed weight has been associated with higher survival percentages after pathogen attack; additionally, larger seeds germinate faster than smaller seeds (Fricke et al. 2019; Larios et al. 2014, Larios and Venable 2018; Litchi et al. 2017; Sánchez-Salas et al. 2017; Wang and Ives 2017), and seedlings from larger seeds are more likely to be successfully established (Garrido et al. 2005; Ortiz-Hernández et al. 2018; Wolf et al. 1986). Therefore, greater viability of large seeds could translate into greater fitness.
Correlations between the characters of branches, seeds, and fruits and differences along the infructescence
As some taxonomists have indicated (García-Mendoza 2011; Gentry 1982), Agave salmiana has a pyramidal inflorescence—clearly exemplified in the present work—characterized by longer basal branches that become gradually shorter toward the tip of the floral peduncle. The largest number of black seeds and highest seed viability are found in the lower three levels of the infructescence.
Variations in seed size within the same individual or the same infructescence have been reported previously. Different explanations have been proposed to explain this phenomenon: an architectural effect, resource availability, and variation in pollen quantity and quality (Hove et al. 2016; Medrano et al. 2000; Pelabon et al. 2021).
According to the architecture-effect hypothesis, basal flowers usually ripen, while distal flowers and fruits rarely do. Natural selection may lead to increased preanthesis inversion in ovaries of basal flowers and less allocation to distal flowers, with a positional decline in ovary size regardless of the fruit-ripening status in the plant or inflorescence (Larios and Venable 2018; Medrano et al. 2000). If this were the case in Agave, we could expect this same pattern in all individuals, regardless of landrace, resource availability, pollinators, and environmental conditions, with no interannual differences.
A second hypothesis concerns competition for spatially and temporarily limited resources. This competition would give rise to very strong selection pressure that may influence all components of plant fitness, growth, survival, and fertility (Larios et al. 2014). The seeds that ripen first, produced on the most basal branches of agaves, would be the first to receive the available resources; these resources would decrease over time, producing the observed pattern. The process from the emergence of the flower peduncle to the formation of fruits can take up to a year; the formation of flowers can begin in April, with fruits fully formed by November (García-Mendoza 2011). Considering a long period, the first fruits to ripen will likely develop.
The third hypothesis concerns the variation in pollen quality or its availability and reception along the inflorescence. Agave salmiana has been reported as a 'generalist' species since its pollinators include insects, birds, and bats (Estrella 2005; Gómez-Aíza and Zuria 2010). By having a greater number of flowers, the lower branches offer more resources (which are attractive) to pollinators. In our study, the first branches were twisted, possibly because the plants studied came from several generations of clones with no sexual reproduction; nonetheless, the largest number of flowers were produced on the lower branches (Fig. 6a). At the peak of the flowering season, when most wild or cultivated plants have flowers, a large volume of pollen is available; this will decline as the growth season progresses, along with the number of pollinators.
Attempts have been made to identify the most likely of the three hypotheses, and the hypothesis concerning resource availability is favored (Hove et al. 2016). These investigations have not yet been carried out in Agave; however, under the environmental conditions in which the genus thrives, this variability in fruit and seed characteristics would safeguard plants against environmental fluctuations. An 'excess' of flowers would act as a reservoir in the face of herbivory and environmental unpredictability. The possibility of intra-individual selection with selective abortion may function as a strategy to increase species fitness (Hove et al. 2016; Medrano et al. 2000; Pelabon et al. 2021). Our results create the possibility of investigating this phenomenon in the genus Agave.
Fruit characters as predictors of the presence of black seeds and differences along the infructescence
In the present work, the subspecies of Agave salmiana show a clear relationship between capsule width and the number of black seeds. Capsule length was irrelevant since long, very thin fruits with a greater number of nonviable seeds were observed. This may change according to the species studied. Colunga-GarcíaMarín et al. (1996) reported correlations of fruit length and width with the number of black seeds. This suggests that larger fruits produce more black seeds. In species of the angustifolia complex, a trend is observed only between fruit length and the number of black seeds (Rivera-Lugo et al. 2018). In wild and cultivated populations of Agave karwinskii, there is no trend relating fruit size to the number of black seeds (Vázquez-Pérez 2015). The cultivated species Agave hookeri has larger fruits and seeds than its supposed wild relative A. inaequidens as a likely outcome of artificial selection (gigantism), and the cultivated species produces fewer black seeds (Figueredo-Urbina et al. 2017).
To harvest the largest number of highly viable seeds, the following is suggested. The first three (basal) infructescence levels have the largest number of fruits, the widest fruits have the largest number of seeds, and the heaviest (“chubbiest”) seeds are the most viable; in addition, according to Ortiz-Hernández et al. (2018), the brightest seeds are the most viable ones.
As for the minimum sampling effort, although a census of flowers, fruits, and seeds provides valuable information with reliable results on the trends of variables measured between and within individual plants, this implies a large investment of resources (human, temporal, and monetary). Our results indicate that sampling three fruits per branch reflects the general trend; even sampling a total of nine fruits may provide information. In this case, however, the information regarding the variability within the infructescence would be missed, which is significant according to the findings of the present study. If only minimal sampling is feasible, we recommend recording identity defined according to the position of flowers, fruits, or seeds along the infructescence.
Conclusions
The analysis of reproductive traits allows differentiating between species, wild plants, and landraces in agaves from which sap is extracted for pulque production. However, a greater understanding of the differentiation between taxa will be achieved if vegetative and reproductive traits are analyzed together. Investigating potential allometric relationships between traits may reflect the effect of artificial selection on reproductive characters. The present work supports the hypothesis that asexual, intensive, and prolonged propagation leads to lower seed set and seed viability in agaves. This work reports for the first time the count of all seeds produced by a single agave plant, which may differ from the number of seeds estimated by sampling only a few fruits. Finally, we found that the most viable seeds were the heaviest seeds produced by the widest fruits in the three basal levels of the infructescence.
Data availability
The data generated and analyzed in the present study are available at https://doi.org/10.5281/zenodo.8088373
References
Álvarez-Ríos G, Pacheco-Torres F, Figueredo-Urbina CJ, Casas A (2020) Management, morphological and genetic diversity of domesticated agaves in Michoacán, México. J Ethnobiol Ethnomed 16:3. https://doi.org/10.1186/s13002-020-0353-9
Arizaga S, Ezcurra E (2002) Propagation mechanisms in Agave macroacantha (Agavaceae), a tropical arid-land succulent rosette. Am J Bot 87:632–641. https://doi.org/10.3732/ajb.89.4.632
Arizaga S, Ezcurra E, Peters E, Ramírez de Arellano F, Vega E (2000) Pollination ecology of Agave macroacantha (Agavaceae) in a Mexican tropical desert. I. Floral biology and pollination mechanisms. Am J Bot 87:1004–1010. https://doi.org/10.2307/2657000
Colunga-GarcíaMarín P, Estrada-Loera E, May-Pat F (1996) Patters of morphological variation, diversity, and domestication of wild and cultivated populations of Agave in Yucatán, México. Am J Bot 87:1069–1082. https://doi.org/10.2307/2445997
Colunga-GarcíaMarín, P, Torres-García I, Casas A, Figueredo-Urbina CJ, Rangel-Landa S, Delgado-Lemus A et al (2017) Los agaves y las prácticas mesoamericanas de aprovechamiento, manejo y domesticación. In: Casas A, Torres-Guevara J, Parra F (eds) Domesticación en el Continente Americano. CDMX, UNAM/UNALM, pp 274–309.
Darwin RC (1876) The effects of cross and self-fertilization in the vegetable kingdom. John Murray, Londres
Eguiarte LE, Aguirre-Planter E, Aguirre X, Colín R, González A, Rocha M, Scheinvar E, Trejo L, Souza V (2013) From isozymes to genomics: population genetics and conservation of Agave in México. Bot Rev 79:483–506. https://doi.org/10.1007/s12229-013-9123-x
Eguiarte LE, Núñez-Farfán J, Souza V, Domínguez C, Cordero C (2015) La reproducción en las angiospermas: Perspectivas ecológicas y evolutivas en el bicentenario de Darwin. In: Morrone JJ, Magaña P (eds) Evolución biológica: una visión actualizada desde la revista Ciencias. UNAM, Facultad de Ciencias, México, pp 135–165.
Eguiarte LE, Jiménez-Barrón OA, Aguirre-Planter E, Scheinvar E, Gámez N, Gasca-Pineda J, Castellanos-Morales G, Moreno-Letelier A, Souza V (2021) Evolutionary ecology of Agave: distribution patters, phylogeny, and coevolution (an homage to Howard S. Gentry). Am J Bot 108:1–20. https://doi.org/10.1002/ajb2.1609
Escobar-Guzmán R, Zamudio-Hernández F, Gil-Vega K, Simpson J (2008) Seed production and gametophyte formation in Agave tequilana and Agave americana. Botany 86:1343–1353. https://doi.org/10.1139/B08-099
Estrella JP (2005) Biología de la polinización de Agave salmiana Otto & Salm—Dick en el valle de Tehuacán-Cuicatlán, México. BSc Dissertation. Universidad Nacional Autónoma de México
Figueredo CJ, Casas A, Colunga-GarcíaMarín P, Nassar JM, González-Rodríguez A (2014) Morphological variation, management and domestication of ‘maguey alto’ (Agave inaequidens) and ‘maguey manso’ (A. hookeri) in Michoacán, México. J Ethnobiol Ethnomed 10:66
Figueredo-Urbina CJ, Casas A, Torres-García I (2017) Morphological and genetic divergence between Agave inaequidens, A. cupreata and domesticated A. hookeri. Analysis of their evolutionary relationships. PLoS ONE 8:1–24. https://doi.org/10.1371/journal.pone.0187260
Figueredo-Urbina CJ, Álvarez-Ríos G, García-Montes MA, Octavio-Aguilar P (2021a) Morphological and genetic diversity of traditional of agave in Hidalgo State, Mexico. PLoS ONE 16:124. https://doi.org/10.1371/journal.pone.0254376
Figueredo-Urbina CJ, Álvarez-Ríos GD, Zárraga-Cortés L (2021b) Edible flowers commercialized in local markets of Pachuca de Soto, Hidalgo, Mexico. Botan Sci 1:120–138. https://doi.org/10.17129/botsci.2831
Flores-García A, Álvarez-Moctezuma JG, Rodríguez de la O JL, Corona-Ambris A (2008) Germinación in vitro de semillas de Nolina parviflora (H.B.K.) Hemsl. Foresta Veracruzana 10:27–33
Fricke EC, Tewksbury JJ, Rogers HS (2019) Linking intra-specific trait variation and plant function: seed size mediates performance tradeoffs within species. Oikos 128:1716–1725. https://doi.org/10.1111/oik.06494
García-Mendoza AJ (2011) Agavaceae. Flora del Valle de Tehuacán-Cuitlatán. Fascículo 88. Instituto de Biología, UNAM, México
García-Meneses PM (2004) Reprodución y germinación en Agave cupreata Trel. & Berger (Agavacea) en la localidad de Ayauco, Guerrero. Bsc Dissertation, UNAM, México
Garrido JL, Rey PJ, Herrera CM (2005) Fuentes de variación en el tamaño de la semilla de la herbácea perenne Helleborus foetidus L. (Ranunculaceae). Anal Jard Bot Mad 62:115–125. https://doi.org/10.3989/ajbm.2005.v62.i2.10
Gentry HS (1982) Agaves of continental North America. The University of Arizona Press, Tucson
Gil-Vega K, González-Chavira MM, Martínez de la Vega O, Simpson J, Vandemark G (2001) Analysis of genetic in Agave tequilana var. Azul using RAPD markers. Euphytica 119:335–341. https://doi.org/10.1023/1017553107303
Gómez-Aíza L, Zuria I (2010) Aves visitantes a las flores del maguey (Agave salmiana) en una zona urbana del centro de México. Ornitol Neotrop 21:17–30
Gonçalves O (1986) El maguey y el pulque en los códices mexicanos. Fondo de Cultura Económica, México City
Guillén S, Terrazas T, De la Barrera E, Casas A (2011) Germination differentiation patters of wild and domesticated columnar cacti in a gradient of artificial selection intensity. Genet Resour Crop Evol 58:409–423. https://doi.org/10.1007/s10722-010-9586-0
Guillen S, Casas A, Terrazas T, Vega E, Martínez-Palacios A (2013) Differential survival and growth of wild and cultivated seedlings of columnar cacti: consequences of domestication. Am J Bot 100:2364–2379. https://doi.org/10.3732/ajb.1300241
Howell DJ, Roth BS (1981) Sexual reproduction in agaves: the benefits od bats: the cost of semelparous advertising. Ecology 62:1–7
Huerta-Lovera M, Peña-Valdivia CB, García-Esteva A, Kohashi-Shibata J, Campos-García H, Aguirre-Rivera JR (2018) Maguey (Agave salmiana) infructescence morphology and its relationship to yield components. Genet Resour Crop Evol 65:1649–1661. https://doi.org/10.1007/s10722-018-0641-6
Husson F, Josse J, Le S, Mazet J (2015) FactoMineR: multivariate exploratory data analysis and data mining. R package version 1.31.4. 24:1–18
Hove AA, Mazer SJ, Ivey CT (2016) Seed set variation in wild Clarkia populations: teasing apart the effects of seasonal resource depletion, pollen quality, and pollen quantity. Ecol Evol 6:6524–6536. https://doi.org/10.1002/ece3.2372
Instituto Nacional de Estadística, Geográfica e Informática (2009) Proanuario de información municipal de los Estados Unidos Mexicanos, Nanacamilpa de Mariano Arista. INEGI. México
International Seed Testing Association (ISTA) (2010) International Rules for Seed Testing. Bassersdorf, CH Switzerland.
Jordan PW, Nobel PS (1979) Infrequent establishment of seedlings of Agave deserti (Agavaceae) in the northwestern Sonora desert. Am J Bot 66:1079–1084
Kassambara A, Mundt F (2020) factoextra: Extract and Visualize the Results of Multivariate Data Analyses. Available from: https://cran.r-project.org/package=factoextra.
Larios E, Venable DL (2018) Selection for seed size: the unexpected effects of water availability and density. Funct Ecol 32:2216–2224. https://doi.org/10.1111/1365-2435.13138
Larios E, Búrquez A, Becerr JX, Venable DL (2014) Natural selection on seed size through the life cycle of a desert annual plant. Ecology 95:3213–3220. https://doi.org/10.1890/13-1965.1
Lichti NI, Steele MA, Swihart RK (2017) Seed fate and decision-making processes in scatter-hoarding rodents. Biol Rev 92:474–504. https://doi.org/10.1111/brv.12240
Loza-Cornejo S, Terrazas T (2011) Morfo-anatomía de plántulas en especies de Pachycereeae: ¿Hasta cuándo son plántulas? Bol Soc Bot 88:1–13
Marquis RJ (1988) Intra-crown variation in leaf herbivory and seed production in Striped Maple, Acer pensylvanicum L. (Aceraceae). Oecología 77:51–55
Medrano M, Guitián P, Guitián J (2000) Patterns of fruit and seed set within inflorescences of Pancratium maritimum (Amaryllidaceae): nonuniform pollination, resource limitation, or architectural effects? Am J Bot 87:493–501. https://doi.org/10.2307/2656592
Molina-Freaner F, Eguiarte LE (2003) The pollination biology of two paniculate agaves (Agavaceae) from northwestern Mexico: contrasting roles of bats as pollinators. Am J Bot 90:1016–1024. https://doi.org/10.3732/ajb.90.7.1016
Mora-López JA, Reyes-Agüero JA, Flores-Flores JL, Peña-Valdivia CB, Aguirre-Rivera JR (2011) Variación morfológica y humanización de la Sección Salmianae del género Agave. Agrociencia 45:465–477
Murray-Tortarolo GN (2021) Seven decades of climate change across Mexico. Atmósfera 34:217–226. https://doi.org/10.20937/atm.52803
Muñoz-Camacho LL, Bello-Cervantes E, Romano-Grande E, Trejo L (2023). Diversity and traditional knowledge of pulque agave landraces in the community of San Predro Tlalcuapan, Tlaxcala, Mexico. Rev Mex Biodiversity 94:e944979. https://doi.org/10.22201/ib.20078706e.2023.94.4979
Nobel PS (1977) Water relations of flowering of Agave deserti. Bot Gaz 138:1–6
Ortiz-Hernández YD, Gutiérrez-Hernández GF, Corzo-Ríos LJ, García-Ramírez E, Martínez-Tomás SH (2018) Varietal and germinative characterization of Agave potatorum (Asparagaceae) seeds with different origins. Botan Sci 96:628–639. https://doi.org/10.17129/botsci.1914
Pélabon C, De Giorgi F, Opedal ØH et al (2021) Is there more to within-plant variation in seed size than developmental noise? Evol Biol 48:366–377. https://doi.org/10.1007/s11692-021-09544-y
R Core Team (2022) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Revelle W (2021) psych: procedures for psychological, psychometric, and personality research. https://CRAN.R-project.org/package=psych
Ramírez-Tobías HM, Niño-Vázquez R, Aguirre-Rivera JR, Flores J, De-Nova-Vázquez JA, Jarquin-Gálvez R (2016) Seed viability and effect of temperature on germination of Agave angustifolia subsp. tequilana and A. mapisaga; two useful Agave species. Genet Resour Crop Evol 63:881–888. https://doi.org/10.1007/s10722-015-0291-x
Reyes-Agüero JA, Peña-Valdivia CB, Aguirre-Rivera JR, Mora-López JA (2019) Infraespecific variation of Agave mapisaga Trel. and A. salmiana Otto ex Salm-Dyck. (Asparagaceae) related to ancestral usages at the hñähñu region in central Mexico. Agrociencia 53:563–579
Rivera-Lugo M, García-Mendoza A, Simpson J, Solano E, Gil-Vega K (2018) Taxonomic implications of the morphological and genetic variation of cultivated and domesticated populations of the Agave angustifolia complex (Agavoideae, Asparagaceae) in Oaxaca, Mexico. Plant Syst Evol 304:969–979. https://doi.org/10.1007/s00606-018-1525-0
Rocha M, Valera A, Eguiarte LE (2005) Reproductive ecology of five sympatric Agave Littaea (Agavaceae) species in central Mexico. Am J Bot 92:1330–1341. https://doi.org/10.3732/ajb.92.8.1330
Romano-Grande E (2022) Impulso a la conservación y producción de agaves utilizados para la producción de agaves utilizados para la producción de pulque: Diversidad morfológica, germinación y sobrevivencia de Agave salmiana subsp. salmiana Manso. MSc. Dissertation, AUTx, Mexico. https://doi.org/10.4067/S0717-66432017000200251
Sánchez-Salas J, Fores J, Jurado E, Sáenz-Mata J, Orozco-Figueroa P, Muro-Pérez G (2017) Hidrocoria en semillas de Agave victoriae-reginae T. Moore, especie en peligro de extinción: Morfología y anatomía con facilitadores de la hidro-dispersión y germinación. Gayana Bot 74:251–261
Santacruz-Ruvalcaba F, Gutiérrez PH, Rodríguez-Garay B (1999) Efficient in vitro propagation of Agave parrasana Berger. Plant Cell 56:163–167. https://doi.org/10.1023/A:1006232911778
Sutherland S (1987) Why hermaphroditic plants produce many more flowers than fruits: experimental tests with Agave mckelveyana. Evolution 41:750–759
Trejo L, Limones V, Peña G, Scheinvar E, Vargas-Ponce O, Zizumbo-Villarreal D, Colunga-GarcíaMarín C (2018) Genetic variation and relationships among agaves to the production of Tequila and Mezcal in Jalisco. Ind Crops Prod 125:140–149. https://doi.org/10.1016/j.indcrop.2018.08.072
Trejo L, Reyes M, Cortés-Toto D, Romano-Grande E, Muñoz-Camacho LL (2020) Morphological diversity and genetic relationships in pulque production agaves in Tlaxcala, Mexico, by means of unsupervised learning and gene sequencing analysis. Front Plant Sci 11:524812. https://doi.org/10.3389/fpls.2020.524812
Trejo L, Luz-Velázquez ML, Vallejo M, Montoya A (2022) Differentiating knowledge of Agave landraces, uses and management in Nanacamilpa, Tlaxcala. J Ethnobiol 42:3150. https://doi.org/10.2993/0278-0771-42.1.31
Valdivieso-Solís DG, Vargas-Escamilla CA, Mondragón-Contreras N, Galván-Valle GA, Gilés-Gómez M, Bolivar F, Escalante A (2021) Sustainable production of Pulque and Maguey in Mexico: current situation and perspectives. Front Sustain F Syst 5:678168. https://doi.org/10.3389/fsufs.2021.678168
Van-Waes JM, Debergh PC (1986) Adaptation of the tetrazolium method for testing, the seed viability, and scanning electron mRCIoscopy study of some Western European orchids. Physiol Plant 66:435–442. https://doi.org/10.1111/j.1399-3054.1986.tb05947.x
Vargas-Ponce O, Zizumbo-Villarreal D, Martínez-Castillo J, Coello-Coello J, Colunga-GarcíaMarín P (2007) In situ diversity and maintenance of Traditional Agave Landraces used in spirits production in West-Central Mexico. Econ Bot 61:362–375
Vázquez-Díaz E, García-Nava JR, Peña-Valdivia CB, Ramírez-Tobías HM, Morales-Ramos V (2011) Tamaño de la Semilla, emergencia y Desarrollo de la plántula de maguey (Agave salmiana Otto ex Salm-Dyck). Rev Fitotec Mex 34:167–173
Vázquez-Pérez N (2015) Variación morfológica y genética de Agave karwinskii (Agavaceae) en los estados de Oaxaca y Puebla. MSc. Dissertation. UNAM, México
Venables, B, Ripley B (2002) Modern applied statistics with S. https://doi.org/10.1007/b97626
Vogler WD, Bahnisch LM (2006) Effect of growing site, moisture stress and seed size on viability and dormancy of Sporobolus pyramidalis (giant rats tail grass) seed. Aust J Exp Agr 46:1473–1479
Wang B, Ives AR (2017) Tree-to-tree variation in seed size and its consequences for seed dispersal versus predation by rodents. Oecologia 183:751–762. https://doi.org/10.1007/s00442-016-3793-0
Ward JH (1963) Hierarchical grouping to optimize an objective function. J Amer Stat Assoc 58:236–244. https://doi.org/10.1080/01621459.1963.10500845
Wolf LL, Hainsworth FR, Mercier T, Benjamin R (1986) Seed size variation and pollination uncertainty in Ipomopsis aggregate (Polemoniaceae). J Ecol 74:361–371. https://doi.org/10.2307/2266093
Funding
The authors state that they received neither funding nor any other financial support during the preparation of this manuscript.
Author information
Authors and Affiliations
Contributions
LT Study design, data collection, manuscript drafting. DS Design of statistical analysis and manuscript drafting. ERG Data collection and statistical analysis. BSC Data collection. DEDN Statistical analysis. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
No conflicts of financial or any other type of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Trejo, L., Soriano, D., Romano-Grande, E. et al. Diversity of reproductive characters, seed set, and viability of Agave seeds used for pulque production and their wild relatives in Tlaxcala, Mexico. Genet Resour Crop Evol 71, 2877–2903 (2024). https://doi.org/10.1007/s10722-023-01803-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10722-023-01803-5