
Abstract
The capulín, or black cherry tree (Prunus serotina Ehrh.) is a tree with edible fruits native to North America that has been used not just as a source of food but also as a source of energy (its wood) since prehispanic times. Mexico has three of the five botanical subspecies in the P. serotina botanical complex, which have not yet been characterized for agricultural purposes. It is desirable to determine whether capulín trees comprise a homogeneous group or whether there are separate morpho-types. This study focuses on the morphological variability of seven populations from the western and central regions of Mexico. We used the Hill & Smith Analysis and graphical methods to analyze and interpret data with respect to 22 quantitative morphological characters and 17 qualitative morphological characters, all from different plant organs. Two major geographically and morphologically related groups were clearly identified, and the morphological variability in the study zone was related to taxonomic, genetic and agronomic factors. The results are discussed from a genetic resources perspective. The genetic, cultural, ecological and agronomic implications of these results are also considered, as well as the potential uses for the species. From these results, an agro-morphological characterization can be developed, in order to identify interesting types of capulín for fruit and seed production in agro-industry, or as rootstock for related species and forestry uses. Our results have implications for the conservation and sustainable use of capulín genetic resources in the central and western regions of Mexico.
Similar content being viewed by others

Introduction
The Capulín (Prunus serotina Ehrh.) is a tree with edible fruits species that grows in temperate zones, from southern Canada to southern Bolivia, and in altitudes between 1000 and 3200 meters in Mexico (Niembro 1990). It is a species that has been known and used in Mexico since prehispanic times. The actual importance of this species is a result of its consumption in central Mexico (Muratalla 1984). The fruit of the tree is a type of cherry, often called capulín, capuli or capolin in Mexico (Popenoe and Pachano 1922). Agronomic studies regarding to productivity and phenology of capulín (Avitia and Muratalla 1982; Muratalla 1984; Buendía 1988; Hernández and Muratalla 1990) have been directed by the Muratalla group in Mexico.
In 2003, a study of the agro-morphological diversity of the capulín was performed in Michoacán, Mexico (Agustín et al. 2003). In that study, agronomic variables were evaluated for trees under different conditions of in situ vegetation management: toleratedFootnote 1 trees and wild trees. The tolerated trees could be considered as semi-cultivated plants because these trees were not planted by the farmers, but since they represented a source of fruits and wood, the farmers then decided to conserve them. Casas et al. (2007) have proposed the term “let standing” to denominate such plants. They defined this term as a plant-human interaction which “includes practices directed to maintain within human-made environments useful plants that occurred in those areas before the environments were transformed by humans.” For simplicity, we decided to use the term “tolerated” as a synonym for the term “let standing”.
The zone in which Agustín et al. performed their study contains the Purepecha ethnic group. Several trees grow in their agricultural fields; additionally, wild capulín can be found in the forest adjacent to those fields. Agustín and collaborators conclude that variables such as fruit weight and yield are not static through time, and studies to identify useful descriptors for this plant species are needed.
In the United States, P. serotina has been studied for forestry purposes because of its capacity to regenerate in perturbed environments (Buendía 1988 and Maynard et al. 1991). The existing studies reporting on in vitro regeneration (Maynard et al. 1991 and Espinosa et al. 2006) have focused on genetic transformation and reproductive sterility induction (Xiaomei and Pijut 2008).
One molecular diversity study of samples from the United States, Mexico and Ecuador identified microsatellite markers specific to P. serotina (Downey and Iezzoni 2000); however, the taxonomic identities of the plants used were not taken into account in the study. With respect to the botanical relationships with other Prunus species, Aradhya et al. (2004) performed molecular characterization and found that P. serotina is related to Prunus mahaleb L., an important species in rootstock generation. The use of P. serotina for food stands out among its potential uses, since its fruit and seeds are more nutritious than those of the sweet cherry or sour cherry tree, according to Avendaño (2000). In Mexico, the fruit of capulín is used for making food or liquors, the wood is used in furniture manufacture because of its hardness and durability, and the leaves and flowers have been noted to have therapeutic properties for respiratory diseases (Losoya 1982).
Important studies by Popenoe and Pachano (1922) in South America, McVaugh (1951) in North America, and Rzendowski and Calderón de Rzendowski (2005) in Mexico have described the diversity of capulín, with the intention of assigning botanical identities to each species. P. serotina is composed of five botanical subspecies (McVaugh 1951). In Mexico, three subspecies are present: P. serotina ssp. capuli (Cav.) McVaugh, P. serotina ssp. serotina (Ehrh.) McVaugh and P. serotina ssp. virens (Wooton et Standl.) McVaugh. The botanical systematics of the subspecies take into account primarily the morphological characteristics of the leaves, flowers and fruit. The botanical classification is ambiguous for some subspecies. For example, in the capuli and serotina subspecies, morphological differences with respect to the size and shape of the leaves are minimal. Some other intra-species differences are based on the agricultural handling of the tree. The subspecies capuli is described as cultivated, and the subspecies serotina is considered to be in the process of being domesticated (Avendaño 2000). The fruit size, seed size, length of flower branch and number of flowers on each branch are the main morphological characteristics that are used in determining domestication. Although these systematic elements are used in distinguishing the capulín subspecies, identification of intra-specific variation requires very specific examination.
The capulín is a fruit-bearing tree distributed widely throughout the central and western regions of Mexico (central-western region). It is necessary to create a program to exploit this tree, starting from an accurate characterization of its different potential uses. Basically, it is necessary to determine whether the capulín trees in central Mexico comprise a homogeneous group, or whether there are geographically separated morpho-types. The present study is a description of the morphological variability of P. serotina Ehrh. in the central-western region of Mexico. It serves to delineate approaches for the management of the genetic resources of the species and to enable an agromorphological characterization for the identification of promising varieties of the capulín for fruit production (fresh fruit, rootstocks) and other uses.
Materials and methods
Plant material collection
Plant specimens from seven tree populations of P. serotina Ehrh. from four states of Mexico, Michoacán, Querétaro, México State and Tlaxcala (Table 1), were collected for the herbarium. The number of sampled trees was directly proportional to the size of the tree population. The dispersion and reproduction characteristics of the capulín populations, as well as their agricultural handling, suggest a count of 20 individuals per 10,000 m2 in the central and western regions of Mexico. For the wild populations, 25% of the trees in a 10,000 m2 area were sampled (Mich1 = 6, Mich2 = 5, Mich3 = 4, Qro = 6, Tlax1 = 5 and Tlax2 = 5 individuals). Of the samples collected from the capulín collection of Chapingo Autonomus University, ten individuals, with desirable agronomic characteristics, such as fruit size (diameter ≥2 cm.) and color, a sweet flavor, and a productivity percentage greater than 50% (flowers/fruits), were recognized. These features were taken into account during the analysis.
Methods
The likely distribution of the species in Mexico was estimated with the use of information from 295 P. serotina subspecies accessions from the herbarium databases and botanical reports analyzed in the Geographical Information System (GIS) FloraMap ver. 1.02 (Arnold 2000). The western and central regions of Mexico were selected as the study regions, based on the GIS results. The determination of the collection sites was aided by botanical reports and communication with Rzendowski and Calderón de Rzendowski (2005). For the climate data and formulas, the Köppen Climate Classification, as modified by Garcia, was used (García 1988).
The descriptor list was developed from the works of McVaugh (1951), Avendaño (2000), Rzendowski and Calderón de Rzendowski (2005) and the Protocol of Distinctness, Uniformity, and Stability Test of Prunus avium L. Sweet Cherry (CPVO 2006). The complete list of capulín descriptors contains 54 general characters (morphological, cytogenetic and agricultural management-related). However, for the present study, only 39 were used. These descriptors are the morphological characters that can be measured in the field and easily evaluated with software analysis. These 39 morphological characters include 22 quantitative variables and 17 qualitative variables (Table 2). These characters were used to evaluate 41 herbarium samples of the capulín from seven western and the central Mexico populations. A descriptive analysis and multivariate methods were used in the data analyses. The flower filaments were pressed, dissected, photographed and measured with the aid of UTHSCSA ImageTool 3.0® Software (UTHSCSA 1998).
For each tree, three herbarium samples were taken from three different zones in the middle part of the treetop. Each sample included a reproductive branch with a minimum of three floral branches. The branches without fruit were collected during a later visit. Around 200 fruits were collected and photographed. Such images were analyzed for color determination and as a source of morphological data (diameter, roundness, size) through image analysis. Trunk color was determined at the collection site with the use of the Munsell Plant Tissue Color Chart (Munsell Color Company 1997), at a point one meter from the ground. Finally, 41 herbarium specimens (with three replicates) were collected, and their descriptor values and passport data were obtained using the mean values of three repetitions for each one. Each collected herbarium sample was labeled, pressed and mounted as a herbarium plant specimen to verify its taxonomic identity.
The Hill and Smith analysis was used for the statistical analysis (Hill and Smith 1976). With this method, simultaneous analysis of the quantitative and qualitative multi-state data was possible. The statistical language R ver. 2.5.1 (R Development Core Team 2007) for Mac OS 10.4.9 and the library ADE4 version 1.4.3 for R (Thioulouse et al. 1996) were used to obtain dual diagrams (Dray and Dufour 2007). The 3D scatterplot was visualized with Normal Contour Ellipsoids in the JMP software (SAS Institute 2007). To determine the variables involved in differentiating the populations, an ANOVA, the Wilcoxon-Kruskal–Wallis test and the HSD Tukey test were used.
Results
Potential distribution and sampling results
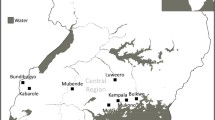
The map generated by FloraMap (Fig. 1) shows that P. serotina is distributed in four main regions in Mexico: northern, western, central and south-eastern, with the central and western regions having the greater incidence of the species. In addition, the three subspecies present in Mexico share ecological niches in the western and central regions, which includes the states of Michoacán, Mexico State, Tlaxcala and Querétaro.

Prunus serotina Ehrh. probability incidences map in Mexico. Zones in dark have a high statistical probability of exhibiting the presence of the species according to climatic and geographic conditions. Map made with 295 accessions analyzed in FloraMap ver. 1.02
The capulín in the west and center of Mexico grow in areas near the Quercus and Pinus forests with 400–900 mm of rainfall per year. The species prefers acidic volcanic soils. The estimated distribution corresponds to the ecological niches described by Venero (1966) and Argueta et al. (1994). McVaugh (1951) recognized the central and western regions as the zones where the three subspecies of P. serotina (capuli, serotina and virens) converge; in those areas, adult capulín trees are commonly found in areas without vegetation, probably propagated by means of birds. The presence of adult capulín trees in glades agrees with the reports of Venero (1966) and Avendaño (2000), in which the capulín is considered a pioneer species (Buendía 1988).
Hill & Smith analysis results
The capulín’s morphological variation, described by the Hill & Smith analysis, clearly demonstrates the relationships among the populations studied, as well as the variability present within the study area. The first two principal components (Factors 1 and 2), with the greatest diversity, explained approximately 18% of the total variance in the data set. The first three principal components (Factors 1, 2 and 3) explained 24.5% of the total variance. These results in the multivariate analysis reveal great diversity among the Capulín accessions used in this study.
Two geographical groups of capulín were distinguished (Fig. 2a): the central group, including four populations from three states in the center of Mexico (Tlaxcala, Querétaro and Mexico State), and the western group, including three populations of Michoacán State. This division is a result of geographical factors; morphological differences between the capulín populations from the center and west of Mexico were found, including differences in characteristics such as productivity percentage (POP), internode length in young stems (ILS), thickness of young stems (TYS), length/thickness quotient length of second basal leaf of flowering branch (LT2), petiole length of second basal leaf of flowering branch (PL2), stamen length (STL), fruit diameter (FRD) and seed diameter (SED). These differences could be a result of the different domestication processes having occurred in the areas, as shown by the differentiation between the cultivated (domesticated) and tolerated trees (Fig. 2b). In the 3D projection (Fig. 2d), three sub-groups were evidenced in the group from central Mexico, with the sub-group Qro being an intermediate between the Mex and Tlax sub-groups. The Mex and Tlax groups shared morphologically related individuals, mainly according to their fruit and seed size and flower branch length. Most of the individuals with anthropocentrically interesting traits were included in a single morphological group (Fig. 2c). Relationships among the individuals from populations Mich1 and Mich2 were evident, but not for the individuals of the Mich3 population.

Visualization of the individuals with respect to three principal components that explain the greater diversity from seven capulín populations’ data from Western and Central regions of Mexico. a and b images show two ecoregions; the groups were formed by States (provinces) of origin in Mexico. c This projection shows three groups, separating a group with desirable fruits characteristics (size, color, flavor, and seed size). d This projection explains population relationships, showing morphological relationships among Querétaro, Mexico and Tlaxcala populations
We determined correlations among the quantitative variables, such as that between the fruit mooring (PFM) and fruit number per flower branch (FIB) (correlation coefficient: 0.850), the thickness of young stems (TYS) and internode length (ILS) (correlation coefficient: 0.522), and the fruit diameter (FRD) and fruit pedicel length (FPL) (correlation coefficient: 0.736). These results highlight the importance of vegetative growth to the final yield of fruits, which is common in other edible fruit species. The main results of the analysis were the relationships among the qualitative and quantitative variables (Table 3) from the use of the Hill & Smith correlation coefficient (ρHS). All of the values of ρHS are positive, and all of the variables have the same direction. For example, the color of the young leaves (YLC) and the tooth number for second leaf border (TQM) both have taxonomic interest, and accordingly McVaugh (1951) used them for his classification and subspecies division of P. serontina. The implications of the YLC-TQM relationship and others are addressed in the Discussion section.
Finally, 21 descriptors, such as the determinant for distinguishing capulín populations (Table 4), were recognized based on significant differences for an ANOVA test or Wilcoxon-Kruskal–Wallis test and their importance to botanical distinctness and agronomic characterization. Of these 21 descriptors, 16 were quantitative variables and 5 were qualitative. However, the list of 39 morphological descriptors can be used for morphological analysis.
Discussion
The present study provides a description of the morphological variability of the capulín in the central and western regions of Mexico and an analysis of the factors affecting their structure in this region. The discussion focuses on the management, use and breeding of the capulín in Mexico.
Distribution of the capulín in the central and western regions of Mexico
We observed that the capulín grows in the ecological niches described by Venero (1966) and Avendaño (2000), that is to say, in a temperate climate, with annual precipitation between 500 and 1000 mm, with deep soils and in the margins of coniferous forests. In addition, adult trees are commonly found in vegetation clearings, as the capulín is sensitive to shading by taller and more vigorous species (Niembro 1990).
The capulín acts as a pioneering species for perturbed environments, which gives it an additional potential use. The capulins in central Mexico were close to human settlements, since their fruits and wood are of anthropocentric interest. In the western region, human beings historically did not handle the capulín trees very much, although that appears to be changing. It is possible to find capulins in vegetation clearings, gorges, and agricultural fields and on roads. Thus, given the species diversification concepts put forth by Vavilov (1927) and Harlan (1992), the studied regions have the environmental requirements and cultural features to support the diversification of the species.
Avendaño (2000) suggests that capulín diversity is obtained through the continuous processes of diversification and domestication. In a context of improvement and plant breeding in situ, as well as conservation, the diversity of the species is desirable because (1) diversity allows the application of artificial selection to one or several traits, (2) diversity helps prevent inbreeding, overall, if the species tends to it in commercial production, and (3) diversity supplies material for future breeding programs or in germplasm regeneration programs, e.g. after a catastrophic event.
The diversity reported in this study, along with our knowledge of biological processes such as domestication and other forms of capulín diversification, as well as the traditional knowledge of the species in some regions of Mexico, suggests that the capulín could be considered as a potential commercial crop, supported by a wide genetic base, which could be useful in modern breeding programs and farmer-participatory plant breeding models.
Morphological variability of Capulín in the central and western region of Mexico
Taxonomic reports identify the Mich1 and Mich2 populations as the serotina subspecies, whereas the Mich3 population is reported as the virens subspecies (Rzendowski and Calderón de Rzendowski 2005). However, the collation of the morphological information with the McVaugh classification (1951) shows that the correspondence is not exact. Calderón de Rzendowski and Rzendowski (2001) have reported the possibility of intermediate individuals between the subspecies (intra-specific hybrids). In our analysis, individuals from the Michoacán populations present these characteristics.
Cordeiro et al. (2008) observed a similar morphological pattern in European sweet and sour cherry collections, in which some autochthonous varieties of cherry in Portugal have the morphological characteristics of hybrids between the sweet cherry (Prunus avium L. and diploid, AA, 2n = 2x = 16) and the sour cherry (P. cerasus L. and allotetraploid, AAFF, 2n = 4x = 32). This result makes sense in relation to our observations. Three polyploidy levels for P. serotina have been reported (4x, 5x, and 6x) by Dickson et al. (1992) and diploid individuals reported by Forbes (1990). Taking into account that the three species are from the same botanic genus, that the phenomenon of allopoliploidy has been reported for Capulín (Pairon and Jacquemart 2005), and that P. serotina is a botanic complex distributed over a wide area, we likely observed intra-specific hybrids.
The phenomenon of hybridization has been considered an important component in the speciation of wild and domesticated plants (Arnold 2004). Recalling Arnold (2004), we are considering natural hybridization to be a phenomenon that implies the occurrence of individuals (hybrids) resulting from a mating between individuals originating in two different populations. Such hybrids are distinguishable from their progenitors with respect to some heritable characters, such as morphological, biochemical, and genetic traits. However, hybrid individuals may be misidentified when the parental groups are morphologically similar to the hybrids and if the distribution of the hybrids overlaps with that of the parental groups (Mahelka et al. 2007).
Hybridization and allopoliploidy may have played important roles in the speciation and colonization of capulín in its native range of distribution and in the environments in which it is considered an invasive species (central Europe). The model of homoploid hybrid speciation can be used in order to understand the morphological intermediacy between groups of related plants (Gross and Rieseberg 2005). However, molecular analyses are necessary in order to verify this application.
We must also consider the influence of human beings on the evolution of capulín. In addition to the ecological speciation and natural selection of capulín, what role has artificial selection played? Is the “hybrid trait” implied in the domestication process of capulín? Is the “hybrid trait” a target of selection? In order to answer these questions, molecular analysis, and cytogenetic and ethnobotanic techniques implicating the other subspecies of P. serotina are desirable. Hybrids are important in plant breeding programs because they are used in the search for new varieties, in understanding the genetic relationships of some traits, and, in the case of Prunus species, in agriculture, as rootstocks for fruit tree varieties.
Environmental conditions also influence some characteristics of trees, for example, the action of humidity at the collection sites. The Mich 2 population is located on the western side of the “Cerro Burro” Mountain (Donkey Hill) in Pátzcuaro, with an agricultural zone bordering the coniferous forest (Quercus and Pinus). The Mich 3 population is located in the municipality of Erongarícuaro, in an agricultural zone with stony ground that does not retain humidity efficiently. In addition, the winds circulating around the Pátzcuaro Lake are not favorable. These environmental differences were translated into variable values for the productivity percentage, number of flowers per branch, and length and diameter of the seed, as well as characteristics inherent to the thickness of the fruit’s skin such as POP, number of flowers per branch (NFB), seed length (SEL), SED, thickness of the fruit skin (TFS) and susceptibility to fruit cracking (SFC).
The seed size, from the Tlaxcala populations, suggests that capulíns have been treated and selected by humans on the basis of their seed characteristics at that site, as Avendaño (2000) indicates. In Michoacán, the capulín is used as an edible-fruit species. The Mex population presents morphological variation with an agronomic interest. The Qro individuals are projected to approximate the Mex individuals. Thus, the individuals of Querétaro and Mexico State have a morphological relationship (Fig. 2d). During the collecting trip in Pinal de Amoles, Querétaro, the inhabitants suggested that, 30 years ago, agricultural workers in Puebla State carried capulín seeds with interesting characteristics (flavor, color and fruit size) to Quéretaro. This information does not agree with the findings of Rzendowski and Calderón de Rzendowski (2005), who identified the capulín located on this site as members of the serotina subspecies.
The morphological variability of capulín in central Mexico likely results from human selection directed to traits with anthropocentric interest, such as flavor and size of the fruit, as well as the size of the seed. Thus, we can conclude that P. serotina is undergoing domestication, as Avendaño (2000) suggests. This interpretation also agrees with that of Cordeiro et al. (2008). They found that more intra-specific morphological variability is found in the sweet cherry (P. avium L.) than in the sour cherry (P. cerasus L.); they state that this pattern is the product of an intensive domestication process, based on the fruit traits of sweet cherry.
According to McVaugh (1951), samples collected in Querétaro (Qro) and Tlaxcala (Tlax) cannot be considered as a serotina subspecies, because those populations have been selected (Qro) and treated (Tlax), indicating that their evolution does not correspond to that of organisms undergoing a natural process.
We consider as treated plants, those selected plants that are targets of strategies directed to promote their growth and production. These strategies include the elimination of weeds or pruning, for example. Casas and collaborators (2007) provide two terms to refer to this type of in situ management of plants under domestication: “Encourage growing” relates to increasing the density of the population (mostly in annual crops), and “Protection,” referring to the deliberate actions favoring the competitiveness of the species under domestication. We consider it convenient to unify these terms, because in a perennial species such as Capulín, before propagation, the people, in the communities that we visited, after selecting the plants, protected them, i.e., they eliminated weeds and branches with symptoms of some diseases or pest infestation. This treatment occurs mostly in the wild or in areas near to agricultural fields with other crops, such as maize, bean or squash.
When the people identified trees showing favorable responses (a greater expression of productivity and/or health), they decided to propagate those plants, by means of seeds or grafting. Commonly, the people decide to move the selected plants and their progenies to an orchard or, occasionally, to their backyards. The management of these plants them changed to an ex situ management, in which the expression of characteristics such as fruit yield became the focus of manipulation; we can say that these plants are cultivated plants, because they have been deliberated produced in order to provide satisfactory products for human consumption.
In the case of the plants from Querétaro, the reason for transferring seeds from a place with selected plants (Puebla) to a place without the species is the desire to get fruits. The implicit reason is propagation of the species, but manipulation for yield is not the main focus yet. Therefore, we can consider that the process of domestication for plants from Querétaro is different from that for populations from Michoacán and Puebla, places in which we could see wild, tolerated, treated and cultivated plants. The plants from Querétaro are evolving from a deliberate transfer, a colonization induced by humans, which involves the introduction of a narrow genetic base. However, these plants are no longer being propagated or manipulated by humans. Additionally, evidence of natural dispersion was seen in Querétaro; it could be a result of the consumption of the fruits by people and birds. Thus, this population can reveal the ecological dynamic of a selected-induced population undergoing colonization under natural selection and the implications for taxonomy.
In contrast, Avendaño (2000) suggests that those populations correspond to P. serotina ssp. capuli, are undergoing domestication. This suggestion is valid if we consider management as the main criterion for differentiation of the subspecies of P. serotina, as McVaugh suggested. However, it is ambiguous in some situations. Therefore, we propose some morphological criteria to differentiate among P. serotina subspecies; however, the utilization of additional techniques (e.g. molecular markers in nuclear and organelles and determining of genome sizes) is desirable.
The relationships among the variables
Characteristics such as the thickness of young stems (TYS), internode length (ILS), fruit diameter (FRD), and fruit pedicel length (FPL) mark potential handling strategies, as pruning is of interest for commercial producers. These features indicate to the breeder or agronomist the type of irrigation, tolerance to wind, rain, hail, and birds, some soil types, pruning intensity and support of the fruit. With markers of this type, ways can be devised to exploit the physiology of the species through pruning or some other agronomic techniques of crop physiology manipulation. Additionally, with such markers, it is possible to assess photosynthesis capability and the ability to be used as a rootstock and determine some of the industrial forestry characteristics, such as wood hardness, wood flexibility, wood porosity, volumetric retractability, compression resistance, traction resistance, and shear stress.
With respect to the quantitative correlations, the variable stamen length (STL) is important because it yields high correlation values for the next variables: the productivity percentage (POP-STL = 0.539), thickness of the fruit pedicel (TFP-STL = 0.534), number of flowers per branch (NFB-STL = 0.505) and seed diameter (SED-STL = 0.534). Descriptors such as the STL, TYS and ILS can be taken as morphological traits for elite individual selection, allowing inferences about the characteristics of yield components and phenotype.
The Hill & Smith correlation coefficient (ρHS), allows for the introduction of relationships between the quantitative and qualitative variables. For example, McVaugh uses the YLC and TQM at the same time in his taxonomic classification to distinguish subspecies. Our analysis demonstrates that both variables give the same information; however, these are different types of variables (YLC = qualitative, TQM = quantitative). Relationships of this type make it possible for the researcher to choose convenient variables because they avoid the duplication of information. Relationships like the one between flowering season and fruit diameter (FWS – FRD) show that, though they give similar information, it is important to take into account traditional knowledge. The settlers of the zones adjacent to the capulín populations indicated the variable FWS, whereas the authors determined FRD. Situations of this type justify the use of multivariate methods with variables of different natures and origins. These methods permit us to maximize the usefulness of the information, linking the scientific and the traditional knowledge. This model can be applied to other underutilized species for which traditional knowledge is available.
The variables showing important differences among the states, localities and populations are the petiole length of the second basal leaf of the flowering branch (PL2) and the number of teeth in the margin of the second basal leaf of the flowering branch (TQM). It is these two variables that McVaugh (1951) used to describe the subspecies. The stamen length, flower diameter, number of flowers per branch and thickness of fruit petiole, seed length and equatorial diameter of seed (STL, FLD, NFB and TFP, SEL, SED, respectively) variables are good descriptors of the differences between the groups. The first four variables are always used in taxonomic classifications. Variables such as SEL and SED are important for germplasm characterization because seed variables are usually stable in time and space (González-Andrés and Ortiz 1995).
The list of descriptors for cultivated species includes quantitative and qualitative characteristics. Those of Cordeiro and his collaborators (2008), who have used both types of characteristics in their analyses, are important for an understanding of the role of native and autochthonous germplasm in intra-specific variability. Methods that permit us to analyze both types of variables at the same time, and the relationships among the variables, are important in providing adequate scientific support for the results, easy interpretations of them, and maximization of the information from plant material with anthropocentric relationships.
Implications for the genetic resource management of the capulín
The capulín is a fruit-bearing tree widely distributed throughout the central-western region of Mexico. From our results, two ecoregions are differentiated: the populations from Tlaxcala, Querétaro and México State comprise the central region, where the capulín is used to produce fresh fruits and seeds for consumption, and the western region (Michoacán), where the species provides seeds for consumption, fresh fruit and agro-ecological services.
We consider agro-ecological service to be the direct or indirect benefits resulting from incorporating a native species under domestication into the agroecosystem. In the case of the capulín, maize producers from the area of Pátzcuaro use it as a windbreak, but they have also selected trees fructifying in August (out of season) for their windbreak walls. The consequence of this deliberate action is that the birds prefer to consume capulín fruits rather than the maize grain (a base for human food), because the fruits are bigger, sweeter and more easily available than the maize grains. The implication of this use of the capulín is that the losses in maize yield are less than would occur when the capulín is not present in the vicinity of the maize fields; therefore, the capulín acts as a distracter for potential pests. Thus, the phenotypes of capulín exhibiting delayed fructifying are valuable, and their management and conservation is part of the traditional knowledge of the community today.
The capulín seeds are considered to be orthodox with latency (Muratalla 1984; Camacho 1980). Their loss of viability has not been evaluated. The clonal propagation of capulín is easy through grafting onto another capulín tree. In situ conservation of capulín in Mexico is necessary. Our work contributes to the descriptions of individuals having special morphological characteristics for agronomic purposes and to the methods for analyzing these characteristics. In situ conservation is urgently needed and recommended for the Michoacán populations. The wild variation is valuable, but it is threatened by the increasing introduction of germplasm from the central region, because such fruit characteristics as fruit size, flavor and color are different in the two populations. Moreover, agricultural land division is severely disturbing the environment of the species being developed.
The genetic relationships among the subspecies of capulín have not yet been clarified. Forbes (1990) suggests the possibility of weak gametophitic self-incompatibility. Dickson et al. (1992) report polyploidy at different levels for capulín. The morphological, physiological and biochemical variations of some subspecies seem to be related to polyploidy. The allopolyploidization process could explain the presence of individuals with characteristics of different subspecies. Additionally, epigenetic phenomena such as mutations are recurrent in the allopolyploidization and could be considered part of the evolutionary process in capulín. Pairon and Jacquemart (2005) reports values of observed heterozygocity (Ho) of 0.987 and an average number of alleles per locus (A) of 10.33 for P. serotina (probably ssp. serotina). These suggest that the capulín is a highly heterozygotic species with allopolyploid origins, explaining its tetraploidy.
The populations from central Mexico exhibit important morphological similarities, possibly as a consequence of human selection. The evolution of populations from the western Mexico region has been less linked to anthropocentric interest related to fruit characteristics, in comparison to populations from central Mexico; instead western Mexico populations appear to be undergoing an incipient domestication process.
The 39 morphological descriptors used could be useful in the agronomic description of the capulín. Descriptors such as the stamen length (STL), thickness of young stems (TYS) and internode length (ILS) have been linked with fruit quality variables. These descriptors can be used as indirect morphological traits for the selection of individuals for breeding programs.
The morphologic variability of the capulín in the central-western region of Mexico described in this study suggests a wide genetic base, useful for improving this species by genetic breeding. Breeding could be directed to the obtaining of selections with better fruit or seed quality, rootstocks for other related species and forestry uses.
Ex situ conservation for the capulín is possible because of the characteristics of its seeds, as well as its simple propagation through grafting. Some communities in Michoacán and Puebla have adopted the practice of grafting as a method for propagating plants with big, sweet fruits, but it has not been generalized to the establishment of orchards. Methods such as cryoconservation and the development of synthetic seeds and cell suspensions must be evaluated. In situ conservation is urgent for the western populations, due to the changes in the environment of the species during its development and the introduction of germplasm from the central region of Mexico. These changes imply the loss and displacement of wild germplasm and tolerated individuals. For both types of conservation, our results can be used to help in the selection of individuals and sites.
Notes
Plants or trees that have existed prior to when the farmer began to work the land. Trees in agricultural fields or parcels, where the farmer has permitted their development for any reason. Now, the farmer uses these trees, but he does not perform agricultural maintenance (e.g. pruning or fertilization) on these trees (Casas et al. 1997).
References
Agustín JA et al (2003) Caracterización morfológica y selección de fenotipos criollos de capulín (Prunus serotina var. capuli (Cav.) McVaugh) en la región templada de Michoacán, In: Avances del Programa de Investigación en Fruticultura (Report). Chapingo, México, p 1
Aradhya MK et al (2004) Molecular characterization of variability and relationships among seven cultivated and selected wild species of Prunus L. using amplified fragment length polymorphism. Sci Hort 103:131–144
Argueta V, Cano AL, Rodarte ME (1994) Capulín (Prunus serotina Ehrh.). In: Argueta V, Cano AL, Rodarte ME (eds) Atlas de la flora medicinal de México. Instituto Nacional Indigenista, México, pp 319–320
Arnold RH (2000) FLORAMAP—predicting distributions. Divers Distrib 6:271–272
Arnold ML (2004) Natural hybridization and the evolution of domesticated, pest and disease organisms. Mol Ecol 13:997–1007
Avendaño GA (2000) Manejo y aprovechamiento de Prunus serotina subsp. capuli Ehrh. (Rosaceae) en comunidades campesinas de Tlaxcala y el Estado de México. MSc. Thesis, Instituto de Recursos Naturales, Colegio de Postgraduados. Mexico
Avitia GE, Muratalla LA (1982) Estudio de germinación de semillas de capulín. Avances en la enseñanza y la investigación. Colegio de Posgraduados, México, pp 256–267
Buendía S (1988) Cultivo y fisiología de tres selecciones de capulín en el suroeste del Estado de México. BSc. thesis, Departamento de Fitotecnia. Universidad Autónoma Chapingo, Mexico
Calderón de Rzendowski G and Rzendowski J (2001) Flora fanerogamica del valle de Mexico, edition. CONABIO. Patzcuaro. Michoacán, Mexico
Camacho MF (1980) Eliminación de la dormancia de semillas de capulín Prunus serotina ssp. capuli (Cav.) mediante el remojo y secado. Cienc Forestal 15:63–74
Casas A et al (1997) Ethnobotany and domestication in Xoconochtli, Stenocereus stellatus (Cactaceae), in the Tehuacán Valley and La Mixteca Baja. México Econ Bot 51:279–292
Casas A et al (2007) In situ management and domestication of plants in Mesoamérica. Ann Bot 100:1101–1115
Community Plant Variety Office (CPVO) (2006) Protocol for distinctness, uniformity and stability test of Prunus avium L. sweet cherry. CPVO Europe, Belgium
Cordeiro RL, Morales MR, Bartolo JA, Ortiz JM (2008) Morphological characterization of sweet and sour cherry cultivars in a germplasm bank at Portugal. Genet Resour Crop Evol 55:593–601
Dickson EE, Arumuganathan K, Kresovich S, Doyle JJ (1992) Nuclear DNA content variation within the Rosaceae. Am J Bot 79:1081–1086
Downey SL, Iezzoni FA (2000) Polymorphic DNA Markers in Black Cherry (Prunus serotina) are identified using sequences from sweet cherry, peach, and sour cherry. J Am Soc Hortic Sci 125:76–80
Dray S, Dufour AB (2007) The ade4 package: implementing the duality diagram for ecologists. J Stat Softw 22:1–20
Espinosa AC, Pijut PM, Michler CH (2006) Adventitious shoot regeneration and rooting of Prunus serotina in vitro cultures. Hort Sci 41:193–201
Forbes D (1990) Self incompatibility and cross-incompatibility in black cherry (Prunus serotina). Dissertation, University of Florida
García E (1988) Modificaciones al sistema de clasificación climática de Köppen. Ed. Universidad Nacional Autónoma de México. México
González-Andrés F, Ortiz JM (1995) Seed morphology of Cytisophyllum, Cytisus, Chamaecytisus and Genista (Fabaceae: Genisteae) species for characterization. Seed Sci Technol 23:289–300
Gross BL, Rieseberg LH (2005) The ecological genetics of homoploid hybrid speciation. J Hered 96:241–252
Harlan JR (1992) Crops and man. American Society of Agronomy, Madison
Hernández JL, Muratalla LA (1990) Inducción floral del capulín (Prunus serotina ssp. capuli) mediante Paclobutrazol (PP333). In: Actas de Horticultura. I Congreso Ibérico de Ciencias Hortícolas vol. IV. Lisboa, Portugal, p 189
Hill MO, Smith AJ (1976) Principal component analysis of taxonomic data with multi-state discrete characters. Taxon 25:249–255
Losoya X (1982) Capulín. In: Losoya X (ed) Flora Medicinal de México serie Plantas Indígenas. IMSS, México, pp 80–93
Mahelka V, Fehrer J, Krahulec F, Jarolimova V (2007) Recent natural hybridization between two allopolyploid wheatgrasses (Elytrigia, Poaceae): ecological and evolutionary implications. Ann Bot London 100:249–260
Maynard CA, Kavanagh K, Fuerukranz H, Drew AP (1991) Black cherry (Prunus serotina Ehrh.). In: Bajaj YPS (ed) Biotechnology in agricultural forestry, vol 16. Springer, Berlin, pp 3–22
McVaugh R (1951) A revision of the North American black cherries (Prunus serotina Ehrh., and relatives). Brittonia 7:279–315
Munsell Color Company (1977) Munsell plant tissue color chart. In: Munsell Color Company. Modules 17 y 320
Muratalla LA (1984) Factores limitantes en la producción de capulín (Prunus serotina ssp. capuli), Technical Bulletin. Colegio de Postgraduados, pp 18–27
Niembro RA (1990) Árboles y arbustos útiles de México. In: Noriega-Limusa (ed) Ciudad de México, México
Pairon MC, Jacquemart AL (2005) Disomic segregation of microsatellites in the tetraploid Prunus serotina Ehrh. (Rosaceae). J Am Soc Hortic Sci 130:729–734
Popenoe W, Pachano A (1922) The Capulin cherry. J Hered 13:51–62
R Development Core Team (2007) A language and environment for statistical computing. Development Core Team R, Vienna, Austria
Rzendowski J, Calderón de Rzendowski G (2005) Prunus serotina Ehrh. In: Rzendowski J, Calderón de Rzendowski G (eds) Flora del Bajío y regiones adyacentes. Instituto de Ecología A. C. Centro Regional del Bajío, Pátzcuaro
SAS Institute Inc (2007) JMP, Version 7.0, Cary, NC. United States. (Software)
Thioulouse J, Chessel D, Dolédec S, Olivier JM (1996) ADE-4: a multivariate analysis and graphical display software. Stat Comput 7:75–83
UTHSCSA Image Tool (1998) IT Version 3.0. Department of Dental Diagnostic Science, University of Texas Health Science Center, San Antonio, p 57
Vavilov NI (1927) Geographical regularities in the distribution of the genes of cultivated plants (Russ., Engl. summary). Bjull Appl Bot Gen i Sel 17:411–428
Venero A (1966) El capulin, su comportamiento en la provincia de Buenos Aires (Capulin. Its behavior in the province of Buenos Aires). Rev Fac Agron 42:143–160
Xiaomei L, Pijut PM (2008) Plant regeneration from in vitro leaves of mature black cherry (Prunus serotina). Plant Cell Tissue Organ Cult 94:113–123
Acknowledgments
The present paper was developed thanks to the facilities given by the Departamento de Fitotecnia in the Universidad Autónoma Chapingo, Mexico.
We are grateful to Dr. Emmanuel Pérez Calix, researcher in the Instituto de Ecología, A.C. – Bajío, for his recommendations to find wild populations of capulín. Also to Dr. Jerzy Rzendowski, curator of the herbarium of the Instituto de Ecología, A.C. – Bajío, for his comments and recommendations in the use of descriptors for the species.
Special thanks to Palma Lower, writing specialist at UC Davis, for her valuable comments and corrections during the improvement of this paper.
Jonathan Fresnedo Ramírez (first author) is supported by a CONACYT-UCMEXUS (Mexican Council of Science and Technology and University of California Institute for Mexico and the United States) doctoral fellowship at University of California, Davis.
Open Access
This article is distributed under the terms of the Creative Commons Attribution Noncommercial License which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This is an open access article distributed under the terms of the Creative Commons Attribution Noncommercial License (https://creativecommons.org/licenses/by-nc/2.0), which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
About this article
Cite this article
Fresnedo-Ramírez, J., Segura, S. & Muratalla-Lúa, A. Morphovariability of capulín (Prunus serotina Ehrh.) in the central-western region of Mexico from a plant genetic resources perspective. Genet Resour Crop Evol 58, 481–495 (2011). https://doi.org/10.1007/s10722-010-9592-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10722-010-9592-2